Embed Size (px)
Citation preview

This article was downloaded by: [Baylor University Libraries]On: 25 June 2014, At: 02:04Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Toxicology and Environmental Health, PartA: Current IssuesPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/uteh20
Pre- and Postshift Levels of Inflammatory Biomarkersand Dna Damage in Non-Bitumen-Exposed ConstructionWorkers—Subpopulation of the German Human BitumenStudyBenjamin Kendzia a , Beate Pesch a , Boleslaw Marczynski a , Anne Lotz a , Peter Welge a ,Hans-Peter Rihs a , Thomas Brüning a & Monika Raulf-Heimsoth aa Institute for Prevention and Occupational Medicine of the German Social AccidentInsurance–Institute of the Ruhr-Universität Bochum (IPA) , Bochum , GermanyPublished online: 11 Jun 2012.
To cite this article: Benjamin Kendzia , Beate Pesch , Boleslaw Marczynski , Anne Lotz , Peter Welge , Hans-Peter Rihs ,Thomas Brüning & Monika Raulf-Heimsoth (2012) Pre- and Postshift Levels of Inflammatory Biomarkers and Dna Damage inNon-Bitumen-Exposed Construction Workers—Subpopulation of the German Human Bitumen Study, Journal of Toxicology andEnvironmental Health, Part A: Current Issues, 75:8-10, 533-543, DOI: 10.1080/15287394.2012.675304
To link to this article: http://dx.doi.org/10.1080/15287394.2012.675304
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Toxicology and Environmental Health, Part A, 75:533–543, 2012Copyright © Taylor & Francis Group, LLCISSN: 1528-7394 print / 1087-2620 onlineDOI: 10.1080/15287394.2012.675304
PRE- AND POSTSHIFT LEVELS OF INFLAMMATORY BIOMARKERSAND DNA DAMAGE IN NON-BITUMEN-EXPOSED CONSTRUCTIONWORKERS—SUBPOPULATION OF THE GERMAN HUMAN BITUMEN STUDY
Benjamin Kendzia, Beate Pesch, Boleslaw Marczynski, Anne Lotz, Peter Welge,Hans-Peter Rihs, Thomas Brüning, Monika Raulf-Heimsoth
Institute for Prevention and Occupational Medicine of the German Social AccidentInsurance–Institute of the Ruhr-Universität Bochum (IPA), Bochum, Germany
Circadian variations in immune defense and tissue repair may interfere with shift effects ofoccupational exposure when investigating biomarkers in cross-shift studies. This investigationcompared biomarkers of inflammation and DNA damage in 59 nonsmoking and 59 smokingmale construction workers pre- (6–10 a.m.) versus postshift (4–7 p.m.). Cellular composi-tions were analyzed in blood, induced sputum (IS), and nasal lavage fluid (NALF) and solubleinflammatory biomarkers were analyzed in IS and NALF. DNA damage was measured as8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodGuo) adducts and DNA strand breaks (alkalineComet assay) in white blood cells (WBC). Apoptosis was quantified as percent apoptotic cellsby annexin V and 7-amino-actinomycin staining in blood lymphocytes using flow cytome-try. In nonsmokers higher preshift than postshift levels of interleukin-8 (IL-8) in IS and moreDNA strand breaks were detected. However, more DNA adducts were found postshift. Amongsmokers, the cellular composition of IS and NALF differed between pre- and postshift samples,in particular more neutrophils pre- than postshift. In contrast, more cells in early apoptosiswere observed post shift in both smokers and nonsmokers. These results indicate a potentialinfluence of circadian rhythms on several biomarkers used in epidemiological studies. Datasuggest interference with shift-work effects of occupational exposure in cross-shift studies andalso the need to consider smoking as a modifying variable.
Inflammatory biomarkers, especiallycytokines and DNA damage, have beenincreasingly employed in research on theadverse health effects due to occupationalexposures. Cross-shift studies have beendesigned to assess the influence of occupationalexposure during a working shift by comparing
This study is part of the Human Bitumen Study, which was initiated by the German Committee for Hazardous Substances(AGS) and the German Bitumen Forum and financially supported by German Social Accident Insurance (DGUV), Eurobitume,Deutscher Asphaltverband e.V., Concawe, Zentralverband des Deutschen Dachdeckerhandwerks, Industrieverband Bitumen-, Dach-und Dichtungsbahnen e.V., Berufsgenossenschaft Rohstoffe und Chemische Industrie (BG RCI) and Aksys GmbH, BeratungsstelleGussasphaltanwendungen (bga), BG BAU, Arbeitsgemeinschaft der Bitumenindustrie (Arbit). We thank all workers who participated inthe Human Bitumen Study. We gratefully acknowledge the support of the field team, especially Anne Flagge, Anja Molkenthin, BiancaWachter, Dieter Höber, Gerd Zoubek, and in parts of the study Klaus Schott (†) and Hans-Jürgen Schicker and the laboratory staff GerdaBorowitzki, Susanne Freundt, Heike Stubel, Maria Düser Sabine Bernard, Evelyn Heinze, Sigrid Reckinger-Pospiech, and Helga Hut fortheir skilful technical assistance.
Address correspondence to Benjamin Kendzia, Institute for Prevention and Occupational Medicine of the German Social AccidentInsurance–Institute of the Ruhr-Universität Bochum (IPA), Bürkle-de-la-Camp-Platz 1, 44789 Bochum, Germany. E-mail: [email protected]
biomarkers pre- and postshift. However,circadian oscillations of major biological pro-cesses may interfere with a work-shift effect, asmay other confounding exposures like smokingand nutrition. Circadian timing is an importantfactor in the temporal organization of DNArepair and immune defense to synchronize
533
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014

534 B. KENDZIA ET AL.
biological processes with periods of differentactivity (Born et al. 1997; Chen and McKnight2007).
The extent of this potential interferencemay differ between blood and peripheraltissues. Inflammatory biomarkers were investi-gated in blood (Ellingsen et al. 2010), but lessis known regarding these biomarkers in periph-eral tissues and their diurnal rhythm. Inducedsputum (IS) and nasal lavage fluid (NALF) con-tain cells and soluble components of the lowerand upper airways and their analysis offers theopportunity to study inflammation and othereffects in peripheral tissues.
Peripheral immune cells display a diur-nal expression of clock genes, another set ofmajor timekeepers in the temporal coordina-tion of biological processes (Kusanagi et al.2004; 2008). There is growing evidence thatthe circadian clock is influenced by cytokinesserving as pacemakers in neuroimmune sig-nalling (Coogan and Wyse 2008). Cytokinesalso play a physiological role in sleep reg-ulation (Kapsimalis et al. 2005; Opp 2005).Whether sleep strengthens immune defenses(Bryant et al. 2004) and serves as a recoveryperiod for enhanced regeneration of periph-eral tissues depends on the activity patterns ofmammals. Circulating hematopoietic stem cellswere found to fluctuate in antiphase with theexpression of chemokines in the bone marrow(Mendez-Ferrer et al. 2008). Monocyte-derivedtype-1 mediated immune defense was reportedto peak at night (Lange et al. 2006).
White blood cells (WBCs) serve as biolog-ical material for assessing genotoxic effects inhumans (Collins et al. 2008). There is growingevidence for diurnal oscillations of blood com-ponents, as reported for erythropoietin levelsas stimulators of erythropoiesis (Gunga et al.2007), and for the release of hematopoieticstem cells (HSC) from bone marrow. However,less is known about circadian variation in DNAdamage and repair. Oxidative DNA damageincludes strand breaks, apurinic/apyrimidinicsites, and oxidized bases. Within the lat-ter group, interest has focused on 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodGuo),a major DNA product with mutagenic potential
(Kasai 1997). DNA strand breaks are one ofthe most common DNA lesions (Lindahl 1993),and they may occur directly via reactive oxygenspecies (ROS) or indirectly as an intermedi-ate during base (mostly 8-oxoguanine) excisionrepair (Caldecott 2003).
The aim of this study was to determinecross-shift differences in immunological andgenotoxic biomarkers in 59 nonsmoking and59 smoking male construction workers at twotime periods. These workers served as a ref-erence group for workers exposed to vaporsand aerosols of bitumen and were not occu-pationally exposed to these workplace-relatedhazards (German Human Bitumen Study, Raulf-Heimsoth et al. 2011b). Biomarker levels werecompared in blood and samples collectedfrom the lower and upper airways pre- versuspostshift by smoking status.
MATERIALS AND METHODS
Study DesignThis analysis was performed in
59 nonsmoking and 59 smoking male roadsideconstruction workers in Germany recruitedas a reference group in the German HumanBitumen Study. Details on the study designwere described in detail elsewhere (Raulf-Heimsoth et al. 2011b). In brief, these workerswere not exposed to vapors and aerosols ofbitumen or other occupational carcinogensand served as a reference group for workersexposed to vapors and aerosols of bitumenor polycyclic aromatic hydrocarbons (PAH).From each worker, one sample each of spoturine, blood, IS, and NALF was collected pre-(6–10 a.m.) and postshift (4–7 p.m.) at mid-week. Smoking status was assessed as current ornoncurrent by self-report using a questionnaireand corrected using the postshift concentra-tion of urinary cotinine with a cutoff of 100μg/L (Haufroid and Lison 1998). All workersprovided written informed consent prior toinvestigation. The German Human BitumenStudy was approved by the Ethics Committeeof the Ruhr-Universität Bochum and conductedin accordance with the Declaration of Helsinki.
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014

CROSS-SHIFT VARIATION OF BIOMARKERS IN WORKERS 535
Collection and Analysis of InducedSputum and Nasal Lavage FluidInduced sputum samples were collected
according to the German Human BitumenStudy protocol (Raulf-Heimsoth et al. 2011b).In brief, isotonic (0.9%) saline aerosol wasinhaled, and the workers were motivated toactively cough, clear their throat, and expecto-rate sputum. Sputum samples were processedwith 0.1% sputolysin (Calbiochem, Darmstadt,Germany). After centrifugation, aliquots of thecell-free supernatants were stored at –80◦Cuntil further analysis of soluble markers. Totalcell number was determined in a Neubauercell chamber, and differential cell counts ofsputum cells were performed using May–Grünwald–Giemsa-stained cytospins. Total pro-tein content was measured according toBradford (1976). Interleukin-8 (IL-8) and othercytokines were determined from thawed cell-free sputum supernatants using commercialmonoclonal “sandwich” enzyme immunoassays(Raulf-Heimsoth et al. 2007; Raulf-Heimsothet al. 2011a). A corresponding procedure wasapplied to NALF samples that were collected byrepeated washing of left nasal cavities with 7ml0.9% saline (Raulf-Heimsoth et al. 2007; Raulf-Heimsoth et al. 2011a). For a few subjects,the recovery of the NALF was not sufficient tomeasure various cellular and soluble markers.
Blood Collection and Determination ofDNA Damage in White Blood CellsThe sample processing time was similar for
both pre- and postshift samples (Marczynskiet al. 2006). Genotoxic effects were assessedby measuring 8-oxodGuo DNA adducts inWBC. In addition, DNA strand breaks weremeasured by assessing percent DNA in tailand Olive tail moment (OTM) in lymphocytesusing the Comet assay (alkaline single-cell gelelectrophoresis assay) as previously reported(Marczynski et al. 2011). In brief, 8-oxodGuoadducts/dGuo were determined according tothe method recommended by the EuropeanStandards Committee on Oxidative DNADamage (Gedik et al. 2002; Gedik and Collins2005). The Comet assay was performed using
a protocol modified by Östling and Johanson(1984) and Singh et al. (1988). This method fol-lows guidelines from the In Vivo Comet AssayWorkgroup of the International Conference onHarmonisation (Burlinson et al. 2007; Tice et al.2000). OTM and percent DNA in tail were usedto quantify DNA strand breaks.
Annexin V PE Apoptosis Detection KitAn Annexin V-Phycoerythrin (Annexin
V-PE) apoptosis detection kit (BD BiosciencesPharmingen, Heidelberg, Germany) was usedto quantify the percent of cells that are activelyundergoing apoptosis. Freshly isolated periph-eral blood mononuclear cells (PBMC) (1 × 106
cells/tube; separated by Ficoll gradient cen-trifugation) were used, and staining for AnnexinV-PE was combined with the dye 7-amino-actinomycin (7-AAD). Cell analysis was per-formed by flow cytometry (FACScalibur, BDBiosciences, Heidelberg, Germany). In total,20,000 cells were counted for each sam-ple, and apoptosis was expressed as percentapoptotic cells in the relevant quadrant. Thefollowing staining results were discriminated:Annexin V-PE negative/7-AAD negative: viablecells (no measurable apoptosis), Annexin V-PEpositive/7-AAD negative: early apoptotic cells(membrane integrity present), and AnnexinV-PE and 7-AAD double positive: late apoptoticcells (dead cells). This new marker was detectedin the blood of 61 subjects enrolled pre- andpostshift after establishing the assay.
Data AnalysisStatistical analyses were performed with
SAS v. 9.2 (SAS Institute, Inc., Cary, NC). Valuesbelow the limits of quantification (LOQ) wereset to two-thirds of the LOQ. Distributionsof values were presented by time point (pre-and postshift) with medians and interquartileranges (IQR) stratified by smoking. To deter-mine the pre- and postshift differences ofpaired observations in the study population orin subgroups, the Wilcoxon signed-rank testwas applied. Bland–Altman plots (Bland andAltman 1986) were constructed to determine
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014

536 B. KENDZIA ET AL.
whether biomarker concentration differencesbetween pre- and postshift (y-axis) were depen-dent on the average values of the two concen-trations (x-axis) separate from smoking status.
RESULTS
The characteristics of the 59 nonsmokingand 59 smoking construction workers are sum-marized in Table 1. In nonsmokers, the medianage was 44 yr and numerically higher as com-pared to smokers (median age 40 yr). The frac-tion of Germans was similar among nonsmokersand smokers (81.4% and 83.1%). In the currentsmoker group, more workers displayed a posi-tive atopy test (specific immunoglobulin [Ig] Eagainst environmental allergens) compared tononsmokers.
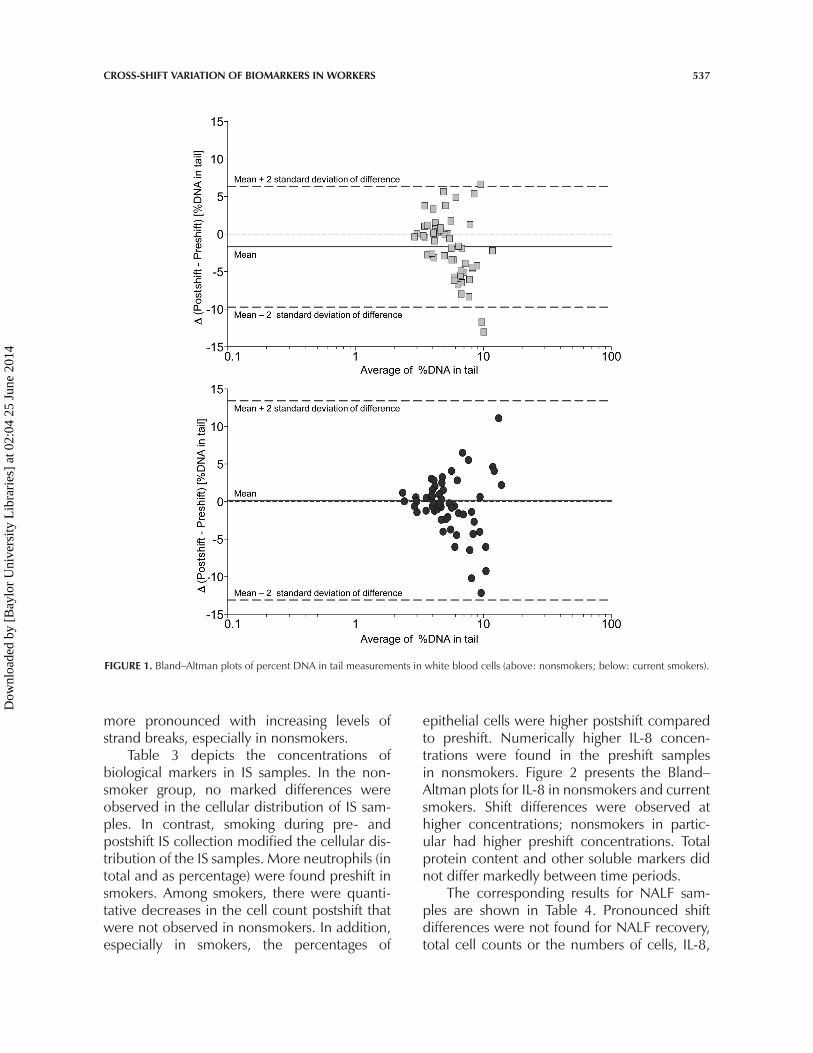
Table 2 presents the biological mark-ers determined in blood samples that wereobtained pre- and postshift. Pre- or postshiftdifferences were not observed for leukocytes,neutrophils, lymphocytes, and monocytesin the nonsmoker and smoker groups.Numerically higher preshift concentrationswere found only for eosinophils in nonsmokersbut not in smokers. Early apoptosis was higherin postshift samples compared to preshiftsamples in both nonsmokers and smokers.In nonsmokers, DNA adducts as evidenced by8-oxodGuo/106 dGuo, were markedly higherin the afternoon, and more DNA strand breakswere observed in preshift than in postshift sam-ples. Figure 1 shows the Bland–Altman plots forDNA strand breaks separately for nonsmokersand current smokers. The shift differences were
TABLE 1. Characteristics of the 59 Nonsmoking and 59 Smoking Construction Workers
Characteristics Nonsmokers, n = 59 Current smokers, n = 59
Age (yr) Median (range) 44 (19–65) 40 (19–59)Body mass index (kg/m2) Median (range) 27.6 (19.6–39.4) 25.5 (21.1–36.1)German nationality N (%) 48 (81.4) 49 (83.1)Cotinine (μg/L) Median (interquartile range) <5 (<5–7.4)a 1666 (966–2248)Positive atopic status sx1 N (%) 16 (27.1) 24 (40.7)
aMedian and 25th percentile are below the limit of detection.
TABLE 2. Distribution (Median, Interquartile Range in Parentheses) of Biological Markers in Blood Samples Pre- and Postshift
Nonsmokers Current smokers
Biological markers Time n Median (P25–P75) pa n Median (P25–P75) pa
Cell distribution Leukocytes (104/μl) 6–10 a.m. 59 5450 (4700–6550) .939 59 6650 (5550–7300) .3544–7 p.m. 59 5750 (4700–6800) 59 6250 (5250–7400)
Neutrophils (abs/μl) 6–10 a.m. 59 3123 (2567–4050) .331 59 3774 (3010–4501) .6444–7 p.m. 59 3317 (2550–4061) 59 3500 (2827–4521)
Lymphocytes (abs/μl) 6–10 a.m. 59 2170 (1870–2635) .366 59 2382 (1811–2849) .2244–7 p.m. 59 2156 (1517–2718) 59 2262 (1888–2727)
Monocytes (abs/μl) 6–10 a.m. 59 35 (14–78) .933 59 43 (17–69) .9244–7 p.m. 59 27 (12–77) 59 22 (0–81)
Eosinophils (abs/μl) 6–10 a.m. 59 105 (60–172) .029 59 133 (88–192) .1144–7 p.m. 59 86 (48–135) 59 124 (58–169)
Genotoxic markers Early apoptosis (%) 6–10 a.m. 25 35.7 (25.9–39.4) .009 36 23.3 (17.4–32.6) <.0014–7 p.m. 25 39.8 (35.2–45.8) 36 34.6 (30.0–40.8)
DNA adducts 6–10 a.m. 58 2.89 (2.47–3.42) <.001 58 3.13 (2.61–3.78) .1148–oxodGuo/106 dGuo 4–7 p.m. 58 3.28 (2.86–4.14) 58 3.27 (2.75–4.38)Percent DNA in tail 6–10 a.m. 58 5.70 (3.99–9.10) .005 58 4.86 (3.59–8.42) .511
4–7 p.m. 58 4.47 (3.49–5.54) 58 4.99 (3.82–7.09)Olive tail moment 6–10 a.m. 58 1.46 (0.96–2.44) .001 58 1.29 (0.98–2.30) .250
4–7 p.m. 58 1.13 (0.95–1.36) 58 1.23 (0.98–1.62)
Note. P25: 25th percentile; P75: 75th percentile.ap Value, for pairwise comparisons between pre- and postshift data with Wilcoxon signed-rank test.
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014
CROSS-SHIFT VARIATION OF BIOMARKERS IN WORKERS 537
FIGURE 1. Bland–Altman plots of percent DNA in tail measurements in white blood cells (above: nonsmokers; below: current smokers).
more pronounced with increasing levels ofstrand breaks, especially in nonsmokers.
Table 3 depicts the concentrations ofbiological markers in IS samples. In the non-smoker group, no marked differences wereobserved in the cellular distribution of IS sam-ples. In contrast, smoking during pre- andpostshift IS collection modified the cellular dis-tribution of the IS samples. More neutrophils (intotal and as percentage) were found preshift insmokers. Among smokers, there were quanti-tative decreases in the cell count postshift thatwere not observed in nonsmokers. In addition,especially in smokers, the percentages of
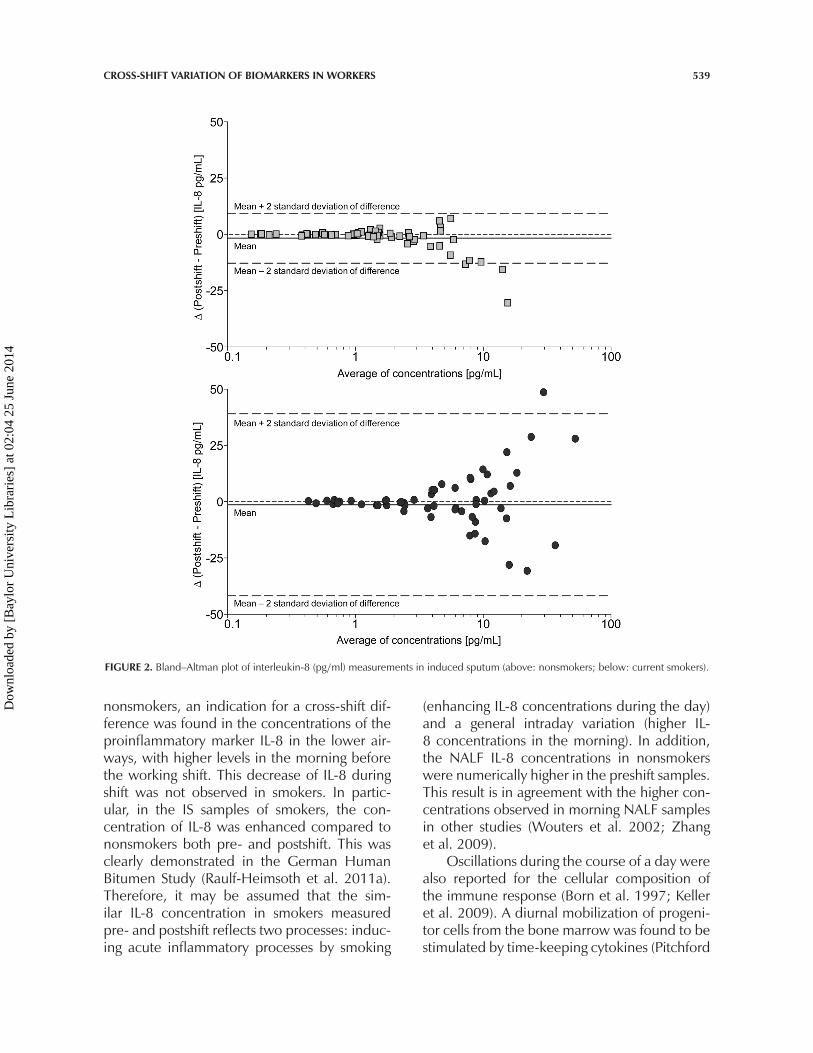
epithelial cells were higher postshift comparedto preshift. Numerically higher IL-8 concen-trations were found in the preshift samplesin nonsmokers. Figure 2 presents the Bland–Altman plots for IL-8 in nonsmokers and currentsmokers. Shift differences were observed athigher concentrations; nonsmokers in partic-ular had higher preshift concentrations. Totalprotein content and other soluble markers didnot differ markedly between time periods.
The corresponding results for NALF sam-ples are shown in Table 4. Pronounced shiftdifferences were not found for NALF recovery,total cell counts or the numbers of cells, IL-8,
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014
538 B. KENDZIA ET AL.
TABLE 3. Distribution (Median, Interquartile Range in Parentheses) of Inflammatory Markers in Induced Sputum Pre- and Postshift
Nonsmokers Current smokers
Inflammatory markers Time n n <LOQ Median (P25-P75) pa n n <LOQ Median (P25-P75) pa
Sputum (g) 6–10 a.m. 59 — 2.5 (1.9–3.5) .875 59 — 2.2 (1.6–3.0) .5264–7 p.m. 59 — 2.6 (1.8–3.5) 59 — 2.4 (1.7–3.1)
Cell count (×105) 6–10 a.m. 59 — 20 (15–34) .906 59 — 30 (19–52) .0354–7 p.m. 59 — 22 (14–42) 59 — 22 (14–39)
Neutrophils (%) 6–10 a.m. 59 — 3.0 (0–12.7) .591 59 — 12.3 (5.0–32.0) <.0014–7 p.m. 59 — 1.3 (0–13.7) 59 — 5.3 (1.0–17.0)
Neutrophils (×104) 6–10 a.m. 59 — 7.1 (0–25.6) .313 59 — 33.9 (7.4–95.7) <.0014–7 p.m. 59 — 3.5 (0–32.8) 59 — 9.2 (2.1–31.4)
Macrophages (%) 6–10 a.m. 59 — 0.7 (0–4.3) .174 59 — 13.7 (2.7–25.7) .2694–7 p.m. 59 — 0.3 (0–2.3) 59 — 4.3 (0.3–22.0)
Macrophages (×104) 6–10 a.m. 59 — 1.4 (0–12.6) .221 59 — 30.8 (5.4–92.8) .0044–7 p.m. 59 — 0.3 (0–7.3) 59 — 9.9 (1.4–39.6)
Epithelial cells (%) 6–10 a.m. 59 — 94.0 (79.4–99.7) .723 59 — 67.0 (36.6–86.3) .0014–7 p.m. 59 — 96.7 (83.0–99.7) 59 — 84.0 (49.7–96.7)
Epithelial cells (×105) 6–10 a.m. 59 — 16.9 (13.0–27.4) .666 59 — 17.2 (11.7–28.1) .8824–7 p.m. 59 — 20.6 (10.0–33.9) 59 — 17.0 (11.0–28.9)
Protein content (μg/ml) 6–10 a.m. 59 0 323 (166–568) .609 59 0 500 (252–766) .0994–7 p.m. 59 0 329 (190–462) 59 0 575 (252–984)
IL-8 (pg/ml) 6–10 a.m. 59 0 1509 (640–3178) .018 58 0 4385 (1709–10041) .7224–7 p.m. 59 1 1085 (513–1966) 58 0 4228 (1074–12276)
IL-1β (pg/ml) 6–10 a.m. 59 0 22.6 (11.7–34.1) .751 58 0 19.2 (12.0–34.8) .8674–7 p.m. 59 0 19.0 (8.4–32.9) 58 0 23.8 (8.7–37.9)
LTB4 (pg/ml) 6–10 a.m. 57 0 2084 (1601–2553) .828 57 0 2025 (1633–2819) .8484–7 p.m. 57 0 1956 (1398–2784) 57 0 2450 (1519–2985)
Note. LOQ: Limit of quantitation. P25: 25th percentile; P75: 75th percentile.ap Value, for pairwise comparisons between pre- and postshift data with Wilcoxon signed-rank test.
IL-1β, and leukotriene (LT) B4 in NALF samplesof nonsmokers. In smokers, the percentages ofneutrophils were higher preshift and the relatedpercentages of epithelial cells were lower.
Adjusting for age and atopy status didnot significantly affect these results (data notshown).
DISCUSSION
A cross-shift study was conducted onirritative, genotoxic, and apoptotic effectsin construction workers with and withoutexposure to vapors and aerosols of bitumen.In order to explore the potential influence ofcircadian variation on pre- or postshift con-centrations, this analysis was restricted to theworkers who served as a reference group in theGerman Human Bitumen Study without expo-sure to workplace-related airborne hazards.The shift difference might by affected by thecircadian variation of inflammatory biomarkersand DNA repair. However, the collection of
blood and other time-consuming material likeNALF and IS more than two times at theworkplace were not feasible in the GermanHuman Bitumen Study. Smoking is a knownconfounder that may exert severe inflamma-tion (Lee et al. 2011; van der Vaart et al.2005) and this parameter was included in thestratified analysis. Further, a relatively large setof biomarkers was investigated. Therefore, astatistical measure of the shift difference wasconsidered not as a dichotomous indicator (sig-nificant or nonsignificant) but as an indicator ofa potentially underlying diurnal variation thatneeds to be confirmed in future studies.
There is growing evidence that immunefunction is under the control of circadian pace-makers to utilize the rest acquired duringsleep for defense mechanisms in peripheraltissues, with cytokines as one of the majortimekeepers (Coogan and Wyse 2008). Mostpublished evidence is derived from nocturnalrodents, a fact that hampers the translationof results to humans. Restricting analyses to
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014
CROSS-SHIFT VARIATION OF BIOMARKERS IN WORKERS 539
FIGURE 2. Bland–Altman plot of interleukin-8 (pg/ml) measurements in induced sputum (above: nonsmokers; below: current smokers).
nonsmokers, an indication for a cross-shift dif-ference was found in the concentrations of theproinflammatory marker IL-8 in the lower air-ways, with higher levels in the morning beforethe working shift. This decrease of IL-8 duringshift was not observed in smokers. In partic-ular, in the IS samples of smokers, the con-centration of IL-8 was enhanced compared tononsmokers both pre- and postshift. This wasclearly demonstrated in the German HumanBitumen Study (Raulf-Heimsoth et al. 2011a).Therefore, it may be assumed that the sim-ilar IL-8 concentration in smokers measuredpre- and postshift reflects two processes: induc-ing acute inflammatory processes by smoking
(enhancing IL-8 concentrations during the day)and a general intraday variation (higher IL-8 concentrations in the morning). In addition,the NALF IL-8 concentrations in nonsmokerswere numerically higher in the preshift samples.This result is in agreement with the higher con-centrations observed in morning NALF samplesin other studies (Wouters et al. 2002; Zhanget al. 2009).
Oscillations during the course of a day werealso reported for the cellular composition ofthe immune response (Born et al. 1997; Kelleret al. 2009). A diurnal mobilization of progeni-tor cells from the bone marrow was found to bestimulated by time-keeping cytokines (Pitchford
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014
540 B. KENDZIA ET AL.
TABLE 4. Distribution (Median, Interquartile Range in Parentheses) of Inflammatory Markers in Nasal Lavage Fluid Pre- and Postshift
Nonsmokers Current smokers
Inflammatory markers Time n n <LOQ Median (P25–P75) pa n n <LOQ Median (P25–P75) pa
Nasal lavage (ml) 6–10 a.m. 59 — 5 (4–6) .367 59 — 5 (4–6) .8414–7 p.m. 59 — 5 (4–6) 59 — 5 (5–6)
Cell count (×104) 6–10 a.m. 59 — 4 (4–8) .725 59 — 8 (4–8) .5844–7 p.m. 59 — 8 (4–8) 59 — 8 (4–8)
Neutrophils (%) 6–10 a.m. 44 — 36.3 (11.2–70.9) .162 49 — 29.3 (2.7–59.4) .0014–7 p.m. 44 — 25.0 (9.2–65.2) 49 — 19.3 (3.3–39.4)
Neutrophils (×103) 6–10 a.m. 44 — 24 (7–46) .091 49 — 22 (1–46) .0794–7 p.m. 44 — 16 (5–31) 49 — 11 (2–26)
Macrophages (%) 6–10 a.m. 44 — 0 (0–1.2) .683 49 — 0 (0–0.3) .1824–7 p.m. 44 — 0 (0–1.0) 49 — 0
Macrophages (×103) 6–10 a.m. 44 — 0 (0–1.2) .565 49 — 0 (0–0.2) .1634–7 p.m. 44 — 0 (0–0.3) 49 — 0
Epithelial cells (%) 6–10 a.m. 44 — 55.5 (25.5–83.9) .189 49 — 64.7 (32.3–97.3) .0014–7 p.m. 44 — 65.8 (32.5–88.5) 49 — 79.3 (57.0–96.0)
Epithelial cells (×103) 6–10 a.m. 44 — 35 (21–48) .337 49 — 39 (26–50) .0354–7 p.m. 44 — 36 (15–61) 49 — 46 (29–76)
Protein content (μg/ml) 6–10 a.m. 59 8 35.8 (17.6–79.6) .212 59 6 30.6 (17.0–63.0) .0024–7 p.m. 59 2 54.3 (27.0–87.1) 59 1 55.0 (31.0–76.0)
IL-8 (pg/ml) 6–10 a.m. 59 2 156.7 (73.6–290.6) .127 59 0 171.1 (72.0–392.9) .1844–7 p.m. 59 0 143.5 (76.0–269.7) 59 0 144.6 (84.0–341.7)
IL-1β (pg/ml) 6–10 a.m. 59 12 1.5 (0.7–2.9) .127 59 19 1.2 (0.3–2.6) .2674–7 p.m. 59 12 1.2 (0.6–2.0) 59 15 1.1 (0.4–2.3)
LTB4 (pg/ml) 6–10 a.m. 53 3 38.6 (25.2–98.1) .112 56 6 28.6 (16.0–65.3) .7814–7 p.m. 53 3 35.4 (23.3–82.4) 56 1 28.0 (16.3–55.6)
Note. LOQ: Limit of quantitation. P25: 25th percentile; P75: 75th percentile.ap Value, for pairwise comparisons between pre- and postshift data with Wilcoxon signed-rank test.
et al. 2009). In our study, the numbers ofcells, especially neutrophils in IS and NALFwere not markedly different between pre- andpostshift among nonsmokers. This finding is inagreement with a study in nonsmoking aller-gic asthmatic subjects where IS was sampled inthe morning and afternoon (Panzer et al. 2003).However, large individual variations in cellu-lar and soluble marker compositions betweenworkers impaired the detection of cross-shiftvariations. In addition, smokers were presentedwith higher preshift cell counts and in particularneutrophils in the IS.
Genotoxic markers in WBC were also mea-sured and intraday differences in nonsmokerswere noted. Two different processes mightinterfere: the diurnal variation in DNA repair,and the accumulation of damage during theday. The separation of these effects mightbe impaired by chance with regard to mul-tiple testing. For example, more DNA strandbreaks were found in the morning, whereasthe percent of cells in the early apoptosis
stage and DNA adducts increased during theshift. The role of smoking on genotoxic effectsis less clear (Pesch et al. 2007). No inter-ference was noted with smoking for adductsand DNA strand breaks. The higher levels ofDNA strand breaks in WBC in morning sam-ples raise the question of whether DNA damageand repair processes undergo circadian vari-ation. Emerging links were reported betweenDNA damage and circadian oscillation (Chenand McKnight 2007). For example, circadiangenes were found to be associated with Cometassay results in experimental animals and invitro (Hoffman et al. 2008; 2010), although lessis known in humans. In contrast, 8-oxodGuolevels were higher postshift (Marczynski et al.2011). This is in agreement with a study of sub-jects with multiple sclerosis (Kanabrocki et al.2006), where urinary 8-oxodGuo was higherin the afternoon. Daytime activity may leadto a permanent production of ROS (Halliwell1998), which are continuously produced afterDNA repair as an endogenous “waste product.”
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014

CROSS-SHIFT VARIATION OF BIOMARKERS IN WORKERS 541
Although a correlation between cellular DNAdamage response and circadian rhythm systemhas long been recognized, its molecular basisis still not clear. Cells have developed a net-work of checkpoint pathways, allowing them toinitiate either cell cycle arrest and DNA repairor apoptosis. Our data showed an increase ofPBMC in the early apoptotic stage in postshiftsamples that was more pronounced in smokers.Moreover, circadian clocks control these path-ways and malignant growth in peripheral tissues(Gery et al. 2006). More studies in humans areneeded to explore the coordination of blood-cell turnover and the aging of blood-cell sub-populations in the context of circadian variationand daily activities.
Noninvasive methods can be applied tocollect samples from peripheral tissues suchas saliva, IS, and NALF. Changes of biomarkerconcentrations in these media during a shiftare considered to be due to occupationalor environmental exposure. Only a few, rela-tively small studies have employed a cross-shiftdesign to investigate biomarkers thus far (Langeet al. 2010; Wouters et al. 2002). A limita-tion of epidemiological studies on chronobi-ology in workers is that samples cannot betaken in narrow time intervals during a shift.Further, the time of processing biological sam-ples from different time points varies accordingto the laboratory work schedule. A poten-tial alteration of biomarkers during transportand storage is another methodological con-cern of biomarker studies. In this study, thesepotential confounding factors were well con-trolled because samples taken in the morn-ing and in the afternoon were immediatelytransported to the laboratory. An impact ofweekday, season, or distance to the labora-tory in the German Human Bitumen Study wasnot detected (Marczynski et al. 2011; Raulf-Heimsoth et al. 2011a).
Our results indicate that in addition to envi-ronmental influences by smoking a cross-shiftdifference in the concentrations of biomarkersof inflammation or DNA damage may alsoreflect an intraday variation of these mark-ers. When selecting the cross-shift design, thepotential interference of circadian rhythms with
exposure during a working shift needs to betaken into account. The extent of such an influ-ence may vary by biomarker or tissue. Soundknowledge of diurnal variations in biomarkerscommonly investigated in occupational epi-demiology is important when assessing shifteffects.
REFERENCES
Bland, J. M., and Altman, D. G. 1986.Statistical-methods for assessing agreementbetween 2 methods of clinical measurement.Lancet 1: 307–10.
Born, J., Lange, T., Hansen, K., Molle, M.,and Fehm, H. L. 1997. Effects of sleepand circadian rhythm on human circulatingimmune cells. J. Immunol. 158: 4454–64.
Bradford, N. M. 1976. A rapid and sensitivemethod for the quantification of microgramquantities of protein utilizing the principleof protein–dye binding. Anal. Biochem. 72:248–54.
Bryant, P. A., Trinder, J., and Curtis, N. 2004.Sick and tired: Does sleep have a vital role inthe immune system? Nat. Rev. Immunol. 4:457–67.
Burlinson, B., Tice, R. R., Speit, G., Agurell,E., Brendler-Schwaab, S. Y., Collins, A. R.,Escobar, P., Honma, M., Kumaravel, T. S.,Nakajima, M., Sasaki, Y. F., Thybaud, V.,Uno, Y., Vasquez, M., and Hartmann, A.2007. Fourth International Workgroup onGenotoxicity testing: Results of the in vivoComet assay workgroup. Mutat. Res. 627:31–35.
Caldecott, K. W. 2003. XRCC1 and DNAstrand break repair. DNA Repair (Amst.) 2:955–69.
Chen, Z., and McKnight, S. L. 2007. Aconserved DNA damage response pathwayresponsible for coupling the cell divisioncycle to the circadian and metabolic cycles.Cell Cycle 6: 2906–12.
Collins, A. R., Oscoz, A. A., Brunborg, G.,Gaivao, I., Giovannelli, L., Kruszewski, M.,Smith, C. C., and Stetina, R. 2008. Thecomet assay: Topical issues. Mutagenesis 23:143–51.
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014

542 B. KENDZIA ET AL.
Coogan, A. N., and Wyse, C. A. 2008.Neuroimmunology of the circadian clock.Brain Res. 1232: 104–12.
Ellingsen, D. G., Ulvestad, B., Andersson, L.,and Barregard, L. 2010. Pneumoproteins andinflammatory biomarkers in asphalt pavers.Biomarkers 15: 498–507.
Gedik, C. M., Boyle, S. P., Wood, S. G.,Vaughan, N. J., and Collins, A. R. 2002.Oxidative stress in humans: validation ofbiomarkers of DNA damage. Carcinogenesis23: 1441–46.
Gedik, C. M., and Collins, A. 2005. Establishingthe background level of base oxidation inhuman lymphocyte DNA: results of an inter-laboratory validation study. FASEB J. 19:82–84.
Gery, S., Komatsu, N., Baldjyan, L., Yu, A., Koo,D., and Koeffler, H. P. 2006. The circadiangene per1 plays an important role in cellgrowth and DNA damage control in humancancer cells. Mol. Cell 22: 375–82.
Gunga, H. C., Kirsch, K. A., Roecker, L.,Kohlberg, E., Tiedemann, J., Steinach, M.,and Schobersberger, W. 2007. Erythropoietinregulations in humans under different envi-ronmental and experimental conditions.Respir. Physiol. Neurobiol. 158: 287–97.
Halliwell, B. 1998. Can oxidative DNA dam-age be used as a biomarker of cancer risk inhumans? Problems, resolutions and prelimi-nary results from nutritional supplementationstudies. Free Radical Res. 29: 469–86.
Haufroid, V., and Lison, D. 1998. Urinarycotinine as a tobacco-smoke exposure index:A minireview. Int. Arch. Occup. Environ.Health 71: 162–68.
Hoffman, A. E., Zheng, T. Z., Ba, Y., Stevens, R.G., Yi, C. H., Leaderer, D., and Zhu, Y. 2010.Phenotypic effects of the circadian genecryptochrome 2 on cancer-related pathways.BMC Cancer 10.110.
Hoffman, A. E., Zheng, T. Z., Ba, Y., and Zhu, Y.2008. The circadian gene NPAS2, a putativetumor suppressor, is involved in DNA dam-age response. Mol Cancer Res 6: 1461–68.
Kanabrocki, E. L., Ryan, M. D., Murray,D., Jacobs, R. W., Wang, J., Hurder, A.,Friedman, N. C., Siegel, G., Eladasari, B.,
Nemchausky, B. A., Cornelissen, G., andHalberg, F. 2006. Circadian variation in mul-tiple sclerosis of oxidative stress marker ofDNA damage. A potential cancer marker?Clin. Ther. 157: 117–22.
Kapsimalis, F., Richardson, G., Opp, M. R.,and Kryger, M. 2005. Cytokines and normalsleep. Curr. Opin. Pulmon. Med. 11: 481–84.
Kasai, H. 1997. Analysis of a form ofoxidative DNA damage, 8-hydroxy-2′-deoxyguanosine, as a marker of cellularoxidative stress during carcinogenesis. Mutat.Res. Rev. Mutat. Res. 387: 147–63.
Keller, M., Mazuch, J., Abraham, U., Eom, G.D., Herzog, E. D., Volk, H. D., Kramer, A.,and Maier, B. 2009. A circadian clock inmacrophages controls inflammatory immuneresponses. Proc. Natl. Acad. Sci. USA 106:21407–12.
Kusanagi, H., Hida, A., Satoh, K., Echizenya,M., Shimizu, T., Pendergast, J. S., Yamazaki,S., and Mishima, K. 2008. Expression profilesof 10 circadian clock genes in human periph-eral blood mononuclear cells. Neurosci. Res.61: 136–42.
Kusanagi, H., Mishima, K., Satoh, K.,Echizenya, M., Katoh, T., and Shimizu, T.2004. Similar profiles in human period1 geneexpression in peripheral mononuclear andpolymorphonuclear cells. Neurosci. Lett.365: 124–27.
Lange, T., Dimitrov, S., and Born, J. 2010.Effects of sleep and circadian rhythm on thehuman immune system. Ann. NY Acad. Sci.1193: 48–59.
Lange, T., Dimitrov, S., Fehm, H. L.,Westermann, J., and Born, J. 2006. Shift ofmonocyte function toward cellular immu-nity during sleep. Arch. Intern. Med. 166:1695–700.
Lee, J., Taneja, V., and Vassallo, R. 2011.Cigarette Smoking and inflammation: cellu-lar and molecular mechanisms. J. Dent. Res.91: 142–149.
Lindahl, T. 1993. Instability and decay of the pri-mary structure of DNA. Nature 362: 709–15.
Marczynski, B., Raulf-Heimsoth, M., Preuss, R.,Kappler, M., Schott, K., Pesch, B., Zoubek,G., Hahn, J. U., Mensing, T., Angerer, J.,
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014

CROSS-SHIFT VARIATION OF BIOMARKERS IN WORKERS 543
Kafferlein, H. U., and Bruning, T. 2006.Assessment of DNA damage in WBCs ofworkers occupationally exposed to fumesand aerosols of bitumen. Cancer Epidemiol.Biomarkers Prev. 15: 645–51.
Marczynski, B., Raulf-Heimsoth, M.,Spickenheuer, A., Pesch, B., Kendzia,B., Mensing, T., Engelhardt, B., Lee, E.H., Schindler, B. K., Heinze, E., Welge,P., Bramer, R., Angerer, J., Breuer, D.,Kafferlein, H. U., and Bruning, T. 2011.DNA adducts and strand breaks in workersexposed to vapours and aerosols of bitumen:Associations between exposure and effect.Arch. Toxicol. 85 Suppl 1: S53–S64.
Mendez-Ferrer, S., Lucas, D., Battista, M., andFrenette, P. S. 2008. Haematopoietic stemcell release is regulated by circadian oscilla-tions. Nature 452: 442–47.
Opp, M. R. 2005. Cytokines and sleep. SleepMed. Rev. 9: 355–64.
Östling, O., and Johanson, K. J. 1984.Microelectrophoretic study of radiation-induced DNA damages in individualmammalian cells. Biochem. Biophys. Res.Commun. 123: 291–98.
Panzer, S. E., Dodge, A. M., Kelly, E. A., andJarjour, N. N. 2003. Circadian variation ofsputum inflammatory cells in mild asthma. J.Allergy Clin. Immunol. 111: 308–12.
Pesch, B., Kappler, M., Straif, K., Marczynski,B., Preuss, R., Rossbach, B., Rihs, H. P.,Weiss, T., Rabstein, S., Pierl, C., Scherenberg,M., Adams, A., Kafferlein, H. U., Angerer,J., Wilhelm, M., Seidel, A., and Bruning, T.2007. Dose-response modeling of occupa-tional exposure to polycyclic aromatic hydro-carbons with biomarkers of exposure andeffect. Cancer Epidemiol. Biomarkers Prev.16: 1863-–73.
Raulf-Heimsoth, M., Pesch, B., Scott, K.,Kappleer, M., Preuss, R., Marczynski, B.,Angerer, J., Rihs, H.P., Hahn, J.U., Merget,R., and Bruning, T. 2007. Irritative effectsof fumes and aerosols of bitumen on theairways: Results of a cross-shift study. Arch.Toxicol. 81: 35–44.
Pitchford, S. C., Furze, R. C., Jones, C. P.,Wengner, A. M., and Rankin, S. M. 2009.Differential mobilization of subsets of pro-genitor cells from the bone marrow. Cell StemCell 4: 62–72.
Raulf-Heimsoth, M., Pesch, B., Kendzia, B.,Spickenheuer, A., Bramer, R., Marczynski,B., Merget, R., and Bruning, T. 2011a.Irritative effects of vapours and aerosols ofbitumen on the airways assessed by non-invasive methods. Arch. Toxicol. 85(suppl. 1):S41–52.
Raulf-Heimsoth, M., Pesch, B., Ruhl, R., andBruning, T. 2011b. The Human BitumenStudy: Executive summary. Arch. Toxicol.85(suppl. 1): 3–9.
Singh, N. P., McCoy, M. T., Tice, R. R., andSchneider, E. L. 1988. A simple techniquefor quantitation of low levels of DNA dam-age in individual cells. Exp. Cell. Res. 175:184–91.
Tice, R. R., Agurell, E., Anderson, D., Burlinson,B., Hartmann, A., Kobayashi, H., Miyamae,Y., Rojas, E., Ryu, J. C., and Sasaki, Y. F. 2000.Single cell gel/comet assay: Guidelines for invitro and in vivo genetic toxicology testing.Environ. Mol. Mutagen. 35: 206–21.
Van der Vaart, H., Postma, D. S., Timens, W.,Hylkema, M. N., Willemse, B. W., Boezen,H. M., Vonk, J. M., De Reus, D. M.,Kauffman, H. F., and Ten Hacken, N. H.2005. Acute effects of cigarette smoking oninflammation in healthy intermittent smok-ers. Respir. Res. 6: 22.
Wouters, I. M., Hilhorst, S. K., Kleppe, P.,Doekes, G., Douwes, J., Peretz, C., andHeederik, D. 2002. Upper airway inflamma-tion and respiratory symptoms in domesticwaste collectors. Occup. Environ. Med. 59:106–12.
Zhang, Y. J., Luroe, S., Schieber, F., Kelsey,J., Nabbie, F., Rizzi, G., Richards, P.,Weiner, R., and Rhyne, P. W. 2009.Immunoassay-based measurement of clini-cal biomarkers for monitoring changes innasal cavity. J. Pharm. Biomed. Anal. 50:823–30.
Dow
nloa
ded
by [
Bay
lor
Uni
vers
ity L
ibra
ries
] at
02:
04 2
5 Ju
ne 2
014




























