Embed Size (px)
Citation preview

Pre- and post-developmental neurogenesis in primates
Pasko Rakic*
Department of Neurobiology, Yale University School of Medicine, 333 Cedar Street, New Haven, CT 06510, USA
Abstract
The [3H]TdR or bromodeoxyuridine (BrdU) methods for marking DNA replication were used to determine time of origin of over 125
classes of neurons in the 34 structures of the macaque monkey central nervous system. The analysis revealed that each neuronal class is
generated in a precisely defined developmental period, the duration of which did not depend on the size and final number of neurons in a
given structure. An exception was the granule cell class of the olfactory bulb and dentate gyrus that continues genesis at a low rate even after
sexual maturity. Thus, most neurons of the primate brain, including cerebral cortex last the entire life span and are normally not renewable.
Overcoming resistance of the adult primate’s brain to acquisition of functionally competent neurons as a form of therapy would benefit from
insight into why neurogenesis normally ceases at a specific developmental period as well as why there are species-specific and regional
differences in the capacity for neurogenesis. q 2002 Elsevier Science B.V. All rights reserved.
Keywords: Neuronal stem cell; Neuronal regeneration; Cerebral cortex
1. Introduction
The enormous interest in research on the embryonic and
neural stem cells is to a large degree inspired by the hope
that they can be used to substitute for lost or damaged
neurons that are normally not being replaced or replaceable.
It is well established that many organs in the human body,
such as the skin or liver are adept at regenerating them-
selves, while other tissues such as the brain and heart are
not [1]. The apparent resistance of the human brain to
renewal of its neuronal constituents stands in contrast to
the higher level of neurogenesis observed in non-mamma-
lian vertebrates such as fish, amphibians, reptiles and birds
(e.g. [2–6]). Adult neurogenesis in these species has been
associated with ether continuous growth and/or turnover of
neurons, and is usually, but not always correlated with the
power for regeneration. Thus, species with a higher capacity
for cell turnover are indispensable model systems for unra-
veling the basic mechanisms essential for restoration of
tissues in an adult organism with lesser or no capacity [7].
Most of what we know about the time of neuron origin in
mammals has been learned from autoradiographic studies
done in mice and rats using radioactive thymidine ([3H]dT)
(e.g. [8–10]). Studies in rodents have provided the basis for
our present understanding of neuronal production in the
mammalian brain in general. Because adult neurogenesis
in mammals is more restricted, it has received renewed
attention only after the introduction of bromodeoxyuridine
(BrdU) labeling as a method for detection of possible neuro-
nal production [11]. In terms of adult neurogenesis in adult
rodents, so far unambiguous evidence has been obtained
only for the interneurons of the olfactory bulb and dentate
gyrus (e.g. [12,13]). Since considerable difference in the
magnitude of neurogenesis has been observed within the
same rodent species depending on their genetic background
[14,15], it is important to examine the time of neuronal
origin in an Old World nonhuman primate that is phylogen-
etically, genetically and anatomically closer to human.
2. Prenatal neurogenesis
In order to determine the timing and sequence of neuron
origin in nonhuman primates, we have initiated a compre-
hensive research program designed to examine the brain of
developing and adult macaque monkeys exposed to [3H]dT,
and more recently to BrdU. Pregnancy in macaque monkey
species lasts 165 days (determined by the time of conception
at the middle of the 28-day menstrual cycle); and, the post-
natal periods of infancy, adolescence and puberty are also
well established [16]. Since [3H]dT is a DNA-specific
nucleotide that is incorporated into nuclear DNA during
the S phase of the cell cycle, the amount of its incorporation
in a given cell is directly proportional to the number of
reduced silver grains in the photosensitive emulsion over-
laying its nucleus. Although the total duration of the cell
cycle in the macaque is 3–4 times longer than in mice, the
phase of DNA synthesis lasts about 8 h in both species [17].
Clinical Neuroscience Research 2 (2002) 29–39
1566-2772/02/$ - see front matter q 2002 Elsevier Science B.V. All rights reserved.
PII: S1566-2772(02)00005-1
www.elsevier.com/locate/clires
* Tel.: 11-203-785-4330; fax: 11-203-785-5263.
E-mail address: [email protected] (P. Rakic).
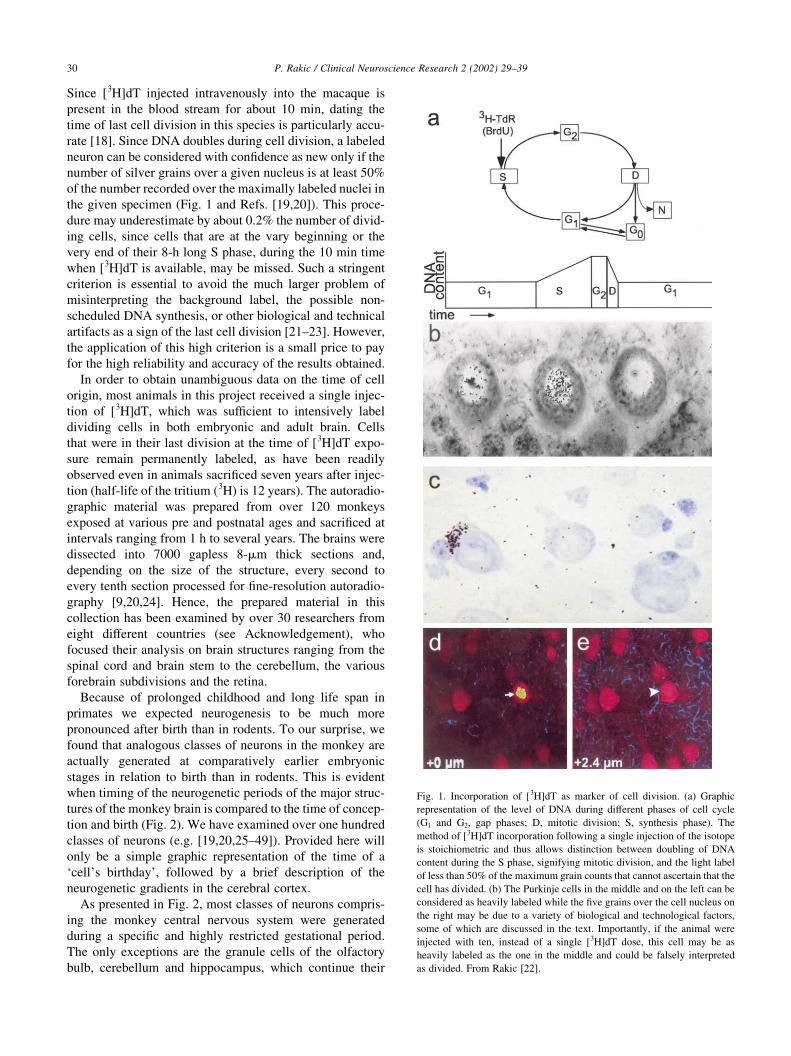
Since [3H]dT injected intravenously into the macaque is
present in the blood stream for about 10 min, dating the
time of last cell division in this species is particularly accu-
rate [18]. Since DNA doubles during cell division, a labeled
neuron can be considered with confidence as new only if the
number of silver grains over a given nucleus is at least 50%
of the number recorded over the maximally labeled nuclei in
the given specimen (Fig. 1 and Refs. [19,20]). This proce-
dure may underestimate by about 0.2% the number of divid-
ing cells, since cells that are at the vary beginning or the
very end of their 8-h long S phase, during the 10 min time
when [3H]dT is available, may be missed. Such a stringent
criterion is essential to avoid the much larger problem of
misinterpreting the background label, the possible non-
scheduled DNA synthesis, or other biological and technical
artifacts as a sign of the last cell division [21–23]. However,
the application of this high criterion is a small price to pay
for the high reliability and accuracy of the results obtained.
In order to obtain unambiguous data on the time of cell
origin, most animals in this project received a single injec-
tion of [3H]dT, which was sufficient to intensively label
dividing cells in both embryonic and adult brain. Cells
that were in their last division at the time of [3H]dT expo-
sure remain permanently labeled, as have been readily
observed even in animals sacrificed seven years after injec-
tion (half-life of the tritium (3H) is 12 years). The autoradio-
graphic material was prepared from over 120 monkeys
exposed at various pre and postnatal ages and sacrificed at
intervals ranging from 1 h to several years. The brains were
dissected into 7000 gapless 8-mm thick sections and,
depending on the size of the structure, every second to
every tenth section processed for fine-resolution autoradio-
graphy [9,20,24]. Hence, the prepared material in this
collection has been examined by over 30 researchers from
eight different countries (see Acknowledgement), who
focused their analysis on brain structures ranging from the
spinal cord and brain stem to the cerebellum, the various
forebrain subdivisions and the retina.
Because of prolonged childhood and long life span in
primates we expected neurogenesis to be much more
pronounced after birth than in rodents. To our surprise, we
found that analogous classes of neurons in the monkey are
actually generated at comparatively earlier embryonic
stages in relation to birth than in rodents. This is evident
when timing of the neurogenetic periods of the major struc-
tures of the monkey brain is compared to the time of concep-
tion and birth (Fig. 2). We have examined over one hundred
classes of neurons (e.g. [19,20,25–49]). Provided here will
only be a simple graphic representation of the time of a
‘cell’s birthday’, followed by a brief description of the
neurogenetic gradients in the cerebral cortex.
As presented in Fig. 2, most classes of neurons compris-
ing the monkey central nervous system were generated
during a specific and highly restricted gestational period.
The only exceptions are the granule cells of the olfactory
bulb, cerebellum and hippocampus, which continue their
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–3930
Fig. 1. Incorporation of [3H]dT as marker of cell division. (a) Graphic
representation of the level of DNA during different phases of cell cycle
(G1 and G2, gap phases; D, mitotic division; S, synthesis phase). The
method of [3H]dT incorporation following a single injection of the isotope
is stoichiometric and thus allows distinction between doubling of DNA
content during the S phase, signifying mitotic division, and the light label
of less than 50% of the maximum grain counts that cannot ascertain that the
cell has divided. (b) The Purkinje cells in the middle and on the left can be
considered as heavily labeled while the five grains over the cell nucleus on
the right may be due to a variety of biological and technological factors,
some of which are discussed in the text. Importantly, if the animal were
injected with ten, instead of a single [3H]dT dose, this cell may be as
heavily labeled as the one in the middle and could be falsely interpreted
as divided. From Rakic [22].

genesis after birth. The duration of the neurogenetic period
of a given structure did not depend on its size and final
number of centers. This point can be illustrated by the
period of neurogenesis in the visual system (Fig. 3). For
example, 1.2 million ganglion cells in the retina are gener-
ated within 40 days (E30–70 [41]) whereas 1.5 million
neurons in their synaptic targets, the lateral geniculate
nucleus, are produced during a period five times shorter
(E36–43 [20]). Even after taking into account that these
two structures display a somewhat different rate of
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–39 31
Fig. 2. Time of neuron origin of the major classes of neurons in the macaque central nervous system. The age of animals in post-conceptual (E ¼ embryonic)
days is represented on the horizontal line, ending at term (E165 ¼ birth) and thereafter in postnatal (P) days. The horizontal lines corresponding to each
neuronal class indicate the animal’s age when an injection of [3H]thymidine results in a heavy label in the neuronal nuclei when the monkeys are killed
postnatally. The dotted line corresponding to the dentate gyrus indicates continued neurogenesis based on the data obtained by BrdU/NeuN double labeling in
adult monkeys. See text for further explanation.
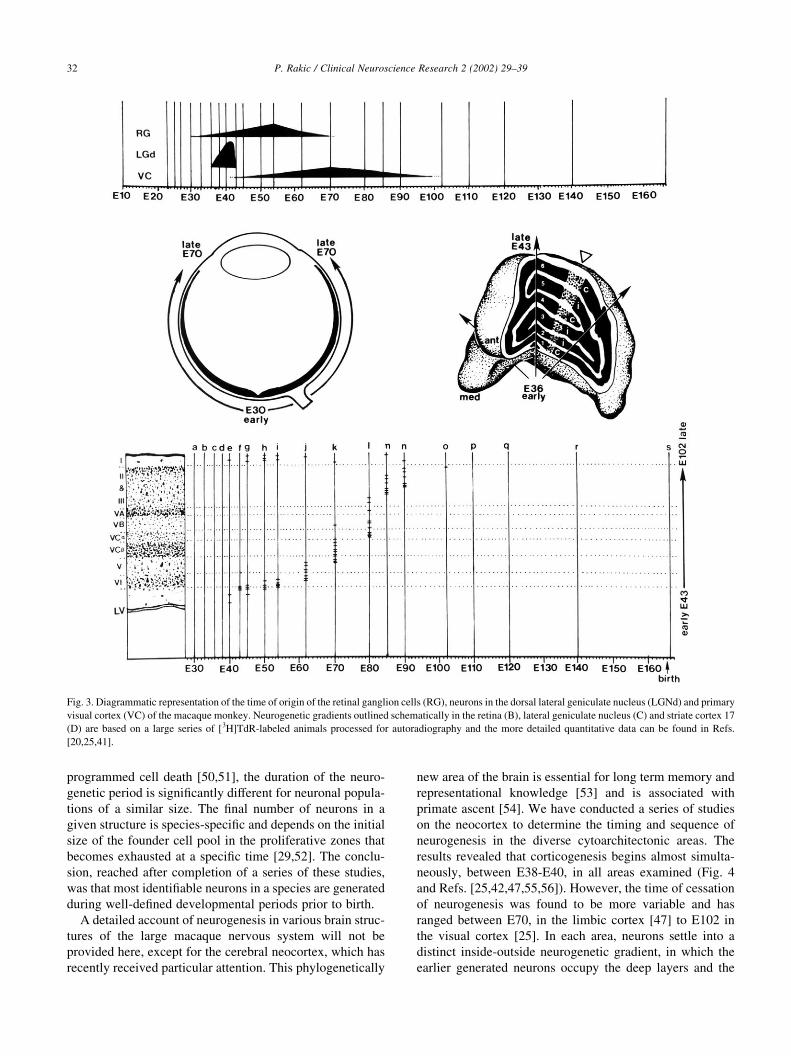
programmed cell death [50,51], the duration of the neuro-
genetic period is significantly different for neuronal popula-
tions of a similar size. The final number of neurons in a
given structure is species-specific and depends on the initial
size of the founder cell pool in the proliferative zones that
becomes exhausted at a specific time [29,52]. The conclu-
sion, reached after completion of a series of these studies,
was that most identifiable neurons in a species are generated
during well-defined developmental periods prior to birth.
A detailed account of neurogenesis in various brain struc-
tures of the large macaque nervous system will not be
provided here, except for the cerebral neocortex, which has
recently received particular attention. This phylogenetically
new area of the brain is essential for long term memory and
representational knowledge [53] and is associated with
primate ascent [54]. We have conducted a series of studies
on the neocortex to determine the timing and sequence of
neurogenesis in the diverse cytoarchitectonic areas. The
results revealed that corticogenesis begins almost simulta-
neously, between E38-E40, in all areas examined (Fig. 4
and Refs. [25,42,47,55,56]). However, the time of cessation
of neurogenesis was found to be more variable and has
ranged between E70, in the limbic cortex [47] to E102 in
the visual cortex [25]. In each area, neurons settle into a
distinct inside-outside neurogenetic gradient, in which the
earlier generated neurons occupy the deep layers and the
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–3932
Fig. 3. Diagrammatic representation of the time of origin of the retinal ganglion cells (RG), neurons in the dorsal lateral geniculate nucleus (LGNd) and primary
visual cortex (VC) of the macaque monkey. Neurogenetic gradients outlined schematically in the retina (B), lateral geniculate nucleus (C) and striate cortex 17
(D) are based on a large series of [3H]TdR-labeled animals processed for autoradiography and the more detailed quantitative data can be found in Refs.
[20,25,41].
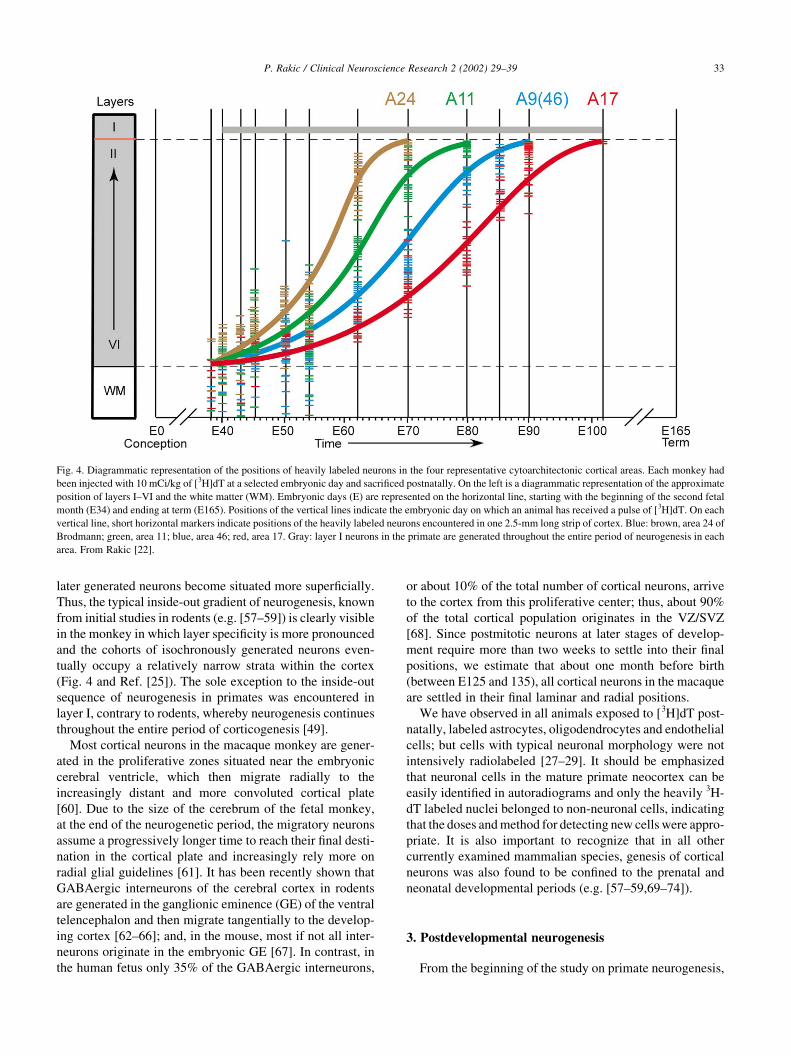
later generated neurons become situated more superficially.
Thus, the typical inside-out gradient of neurogenesis, known
from initial studies in rodents (e.g. [57–59]) is clearly visible
in the monkey in which layer specificity is more pronounced
and the cohorts of isochronously generated neurons even-
tually occupy a relatively narrow strata within the cortex
(Fig. 4 and Ref. [25]). The sole exception to the inside-out
sequence of neurogenesis in primates was encountered in
layer I, contrary to rodents, whereby neurogenesis continues
throughout the entire period of corticogenesis [49].
Most cortical neurons in the macaque monkey are gener-
ated in the proliferative zones situated near the embryonic
cerebral ventricle, which then migrate radially to the
increasingly distant and more convoluted cortical plate
[60]. Due to the size of the cerebrum of the fetal monkey,
at the end of the neurogenetic period, the migratory neurons
assume a progressively longer time to reach their final desti-
nation in the cortical plate and increasingly rely more on
radial glial guidelines [61]. It has been recently shown that
GABAergic interneurons of the cerebral cortex in rodents
are generated in the ganglionic eminence (GE) of the ventral
telencephalon and then migrate tangentially to the develop-
ing cortex [62–66]; and, in the mouse, most if not all inter-
neurons originate in the embryonic GE [67]. In contrast, in
the human fetus only 35% of the GABAergic interneurons,
or about 10% of the total number of cortical neurons, arrive
to the cortex from this proliferative center; thus, about 90%
of the total cortical population originates in the VZ/SVZ
[68]. Since postmitotic neurons at later stages of develop-
ment require more than two weeks to settle into their final
positions, we estimate that about one month before birth
(between E125 and 135), all cortical neurons in the macaque
are settled in their final laminar and radial positions.
We have observed in all animals exposed to [3H]dT post-
natally, labeled astrocytes, oligodendrocytes and endothelial
cells; but cells with typical neuronal morphology were not
intensively radiolabeled [27–29]. It should be emphasized
that neuronal cells in the mature primate neocortex can be
easily identified in autoradiograms and only the heavily 3H-
dT labeled nuclei belonged to non-neuronal cells, indicating
that the doses and method for detecting new cells were appro-
priate. It is also important to recognize that in all other
currently examined mammalian species, genesis of cortical
neurons was also found to be confined to the prenatal and
neonatal developmental periods (e.g. [57–59,69–74]).
3. Postdevelopmental neurogenesis
From the beginning of the study on primate neurogenesis,
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–39 33
Fig. 4. Diagrammatic representation of the positions of heavily labeled neurons in the four representative cytoarchitectonic cortical areas. Each monkey had
been injected with 10 mCi/kg of [3H]dT at a selected embryonic day and sacrificed postnatally. On the left is a diagrammatic representation of the approximate
position of layers I–VI and the white matter (WM). Embryonic days (E) are represented on the horizontal line, starting with the beginning of the second fetal
month (E34) and ending at term (E165). Positions of the vertical lines indicate the embryonic day on which an animal has received a pulse of [3H]dT. On each
vertical line, short horizontal markers indicate positions of the heavily labeled neurons encountered in one 2.5-mm long strip of cortex. Blue: brown, area 24 of
Brodmann; green, area 11; blue, area 46; red, area 17. Gray: layer I neurons in the primate are generated throughout the entire period of neurogenesis in each
area. From Rakic [22].

it was clear that granule cells of the hippocampus and cere-
bellum are generated to a considerable extent postnatally
[19,27–29,36]. However, we were even more impressed
that among over the hundreds of classes of neurons exam-
ined, only the class of small interneurons (granule cells) and
few rod photoreceptors at the edge of the retina are gener-
ated after birth in the macaque monkey. While production of
the rods halts a few days after birth [41], the production of
the cerebellar granule cells continues during infancy and the
juvenile period, but could not be observed in animals
injected with [3H]dT after the third postnatal month [27].
While we noticed that proliferation of cells in the subgra-
nular zone of the dentate gyrus continues after sexual matur-
ity (third year of life) the methods for determining the nature
of these small cells at the time did not allow conclusive
distinction between granule cells (neurons) and astrocytes
(glia). Application of autoradiography at the electron micro-
scopic level revealed heavily labeled cells with only the
ultrastructural characteristics of astrocytes and oligodendro-
cytes [39]. Thus, we concluded that “if neurogenesis has
been missed as a result of methodological limitations, the
number of newly generated neurons must be very small and
thus hardly comparable to massive levels of neurogenesis
reported in birds and rodents “ [39]. At the time, the olfac-
tory bulb had not been dissected and examined with the
[3H]dT autoradiographic method.
After introduction of immunohistochemistry for BrdU and
the immunomarkers for neurons, we re-examined this issue
and found double labeled cells in the dentate gyrus of the
adult monkey [75]. To prove that these were indeed new
cells, we followed their developmental history from their
last division in the subgranular zone, through the migration
phase of bipolar cells, and finally to their differentiation in the
dentate gyrus [75]. No new neurons were formed by division
of the pre-existing hippocampal neurons. Rather, they origi-
nated from a remnant of the pluripotential stem cells situated
in the subgranular zone of the dentate gyrus in the hippocam-
pus. We estimated that in the dentate gyrus of the adult
monkey (5.5–16.5 years of age) one new granule cell is
generated each day per 24 000 existing cells, or 0.004% of
the granule cell population [75], which is about one order of
magnitude less than in mice [15]. It is not known whether
these cells in the primate incorporate into appropriate synap-
tic circuits, or how long they survive. It they are destined to
replace existing neurons, it would take over 65 years or about
three average life spans of the macaque to turn over all gran-
ule cells in the dentate gyrus. However, since the majority of
newly generated neurons die shortly after their genesis [76],
in reality it would take much longer. Since there is a consid-
erable genetic difference in the rate of neurogenesis in the
dentate gyrus even among different strains of mice [14,15], it
is not surprising that the difference may exist between species
millions of years apart in the evolutionary scale. At this point
it is not known whether new granule cells in the primate
dentate gyrus become incorporated into proper synaptic
circuits and what is their function.
We have also found that the subependymal zone (SEZ) in
the adult monkey telencephalon, similar to other mammals,
generates glial cells as well as prospective neurons that
migrate tangentially, via the rostral migratory stream
(RMS) and olfactory nerve, to the olfactory bulb [77].
These SEZ/RMS cells in adult monkeys are morphologi-
cally and biochemically similar to migrating cells described
in adult rodents (e.g. [12,28]) and form chains that are
immunopositive for surface markers, TuJ1 and PSA-
NCAM that characterize this migratory stream [77,78].
The molecular mechanisms that channel newly generated
neurons to the olfactory bulb via the RMS and repels
them from entering into other neighboring structures has
been elucidated in the mouse [79]. However, in the monkey
it takes about 3 months for cells to complete their migration
via the RMS to become mature neurons, probably reflecting
a lower rate of genesis as well as migration and a protracted
tempo of differentiation. The relatively smaller daily rate of
neurogenesis in the olfactory system in the adult primate
compared to that observed in rodents can be explained by
the reduced size of the olfactory bulb in relation to the large
expansion of the primate cerebral cortex and other structures
engaged in cognitive functions [80,81].
Although all classes of neurons in the large macaque
nervous system have not been examined with the new meth-
ods, the overall assessment of the presently available data is
that neurogenesis in adult primates is more limited than in
rodents [22,75,77,82]. A similar limit appears to exist in the
human forebrain (e.g. [83]). Thus, the report of a large daily
influx of new neurons to the association neocortex in maca-
que monkey, particularly the principal sulcus of the prefron-
tal area [84,85] came as a surprise and has necessitated re-
examination using the same method. Thus we performed a
study on additional 10 macaque monkeys of mixed gender,
some of which were raised in a group and exposed to enriched
environment [82]. Examination of the cerebral cortex follow-
ing BrdU injections to these animals revealed labeled cells in
the subependymal zone of the lateral cerebral ventricle [82]
as previously observed by other investigators (e.g. [86,87]).
However, we also found that cells doubly-stained with TuJ1
(a marker for immature neurons) were entering the rostral
migratory stream on way to the olfactory bulb, where they
differentiate into granule cells [77]. Although BrdU-labeled
nuclei were also observed in the neocortex, examination of
several thousand cells using triple-labeled immunofluores-
cent histochemistry for BrdU, NeuN (a marker for mature
neurons) and GFAP (a marker for astrocytes) yielded none
that met the basic criteria for a new neuron [77]. Likewise, the
massive migratory stream to the principal sulcus that was
suggested to be a conduit for new neurons to the prefrontal
cortex [84] could not be identified.
A more detailed evaluation of the available evidence about
adult neurogenesis and the problems of the methods used for
detecting new cells can be found elsewhere [21–23,82,88]. In
short, the review of the caveats of the present state of the art
shows that BrdU incorporation alone can not be taken as a
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–3934

definitive marker for cell division, but rather a marker for
DNA synthesis, which can also occur in nondividing cells
and can actually be enhanced by a variety of factors and
conditions including the process of programmed cell death
(e.g. [89]). It is often overlooked that BrdU is toxic and may
act as a mutagen or stimulator of DNA synthesis if given in
high and repeated doses (e.g. [90]). Finally, one has to
exclude the possibility that BrdU labeling or polyploidy is
not due to the fusion of the newly generated blood born cells
with the nuclei of existing, damaged or aged neurons [91,92].
Thus, BrdU incorporation in the cell’s nucleus is essential,
but not a sufficient indication that a given cell has actually
divided [21,22,23]. Before concluding that the occasional
BrdU labeled cell found in the adult or aging brain tissue is
a newly generated neuron, it is essential to exclude BrdU
labeling as not resulting from DNA synthesis in response to
cell damage, abortive mitoses, polyploidy, or the initial steps
in naturally occurring cell death.
It is noteworthy that, although cortical neurons in the
monkey are produced prenatally, the volume of the cerebral
cortex continues to grow at a considerable degree postna-
tally. Detailed examination of the cytological maturation of
the primate neocortex reveals that its postnatal expansion is
due to multiple aspects of cellular growth and maturation
and a large addition of non-neuronal cells. First, the size of
neurons in the primate cortex doubles during the postnatal
period in both monkey and human [93–95]. Second, after
cessation of neurogenesis and completion of neuronal
migration to the cortical plate, the formation of axonal
connections and synaptogenesis continues at an exponential
pace during the postnatal period in both monkeys and
human [96–98]. Third, postnatal development of the
primate cerebrum is also associated with a massive invasion
of various types of glial and endothelial cells. Although
formation of astrocytes in the macaque monkey begins
before birth [99–101], the major wave of gliogenesis that
includes oligodendrocytes occurs during the neonatal and
postnatal period when these cells are produced in both the
SEZ as well as in the GE [86,87,95]. The BrdU labeled
oligodendrocytes can be seen throughout the white matter
of the forebrain including the corpus callosum. Small ellip-
soid nuclei and bipolar shape resemble migrating neurons,
but their size, distributions along myelinated tracts, their
tinctorial properties and absence of reactivity to neuronal
markers indicate their glial nature [22,23,28]. The majority
of oligodendroglial cells, which in the primate cerebrum
outnumber neurons by severalfold, are generated until
puberty. However, gliogenesis continues during adulthood
in both the SEZ, as well as in situ within the white matter
and cerebral cortex itself throughout the prolonged primate
life span in both monkeys and human [28,82,95,102,103].
4. Evolutionary and medical perspective
With the notable exception of the granule cells in the
dentate gyrus and olfactory bulb, most neurons in the
primate brain including our own, appear to be generated
before birth or the early neonatal period and then persist
during many decades of the primate life span. In contrast,
cells in many other organs of the body are being recycled
every few months [104]. In a more primitive organism such
as hydra and even in lower vertebrates such as amphibians
and reptiles, the capacity for self renewal is much higher, as
their individual organs or appendages can be restored
following amputation [105]. Newts and salamanders are,
for example, capable of a spectacular degree of entire
limb regeneration [106]. This capacity, also called
epimorphic restoration, is not possible in most birds and
mammals. The general decline in capacity for cell renewal
and epimorphic restoration during the course of evolution
has puzzled evolutionary biologists for years. Why should
such a useful ability be lost? One explanation offered was
that, in warm-blooded animals, it has lost survival value.
While cold-blooded animals can survive in their ecological
niche or a sufficient length of time without movement and
food to allow re-grow of a new limb, warm-blooded animals
die from hunger or are killed by predators before their limbs
or organs can be restored (e.g. [105,107]).
The gradual diminishing capacity of neuronal turnover in
the adult brain during vertebrate evolution is equally
puzzling. The heart is the only other organ that is not
capable of renewing its contractile cells under ether normal
or pathological conditions. [108]. Thus, the argument of
‘loss of survival value’ made for the other organs (see
above and Ref. [107]) could also apply to the heart or
brain. In the latter case, the process of neuronal prolifera-
tion, migration and the formation of elongated axons that
form precise synaptic connections takes much more time
than most mammals, in their natural environment, survive.
In spite of these negative evolutionary pressures, it seems
reasonable to explore whether there may be some positive
survival value for keeping the same set of neurons for an entire
life span. The need for a permanent population of neurons may
be a biological necessity for storage of learned behavior in all
species. Even in organisms which have unusual evolutionary
adaptation, like the male canary, that undergoes seasonal turn-
over of particular neuronal types, have a subpopulation of
neurons that are nonrenewable and apparently important in
memory function (e.g. [109]). Could increasing the proportion
of the nondividing neurons that last during the entire life span
in the brains of the mammals help their survival? Could such
neurons have a particular functional significance in primates
including human? I have hypothesized that during vertebrate
evolution the ability for neuronal renewal and regeneration
may have been traded for the ability to retain them throughout
the entire life span in order to enhance retention of long-term
memory and use the learned behavior for their survival [28].
Preservation of acquired information within a permanent
population of neurons in these species may be more valuable
for the survival of an organism than the introduction of ‘naive’
neurons. Is the active population of stem cells found in the SEZ
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–39 35

and subgranular zone which produce granule cells for the
olfactory bulb or dentate gyrus in the adult primate, a rudiment
left from our predecessors, similar to the appendix in the diges-
tive system, which has lost survival value? Or alternatively, is
it a small but important aspect of the adult function in this
particular structure? Irrespective of what the final answer on
these questions may eventually be, it is essential to continue
research on these systems in order to learn how newly formed
neurons are incorporated into the circuitry and what may be
their function in species where this is a more common event.
Understanding the basis of the difference between species may
help to elucidate the significance of the newly generated gran-
ule cells as well as to design a method on how to enhance this
process if deemed functionally useful.
While longevity of neurons may be useful for storage of
long-term memory and life-long experience in the cerebral
neocortex, the decline in capacity for neurogenesis may be
an impediment for replacement of lost neurons in the motor
and sensory systems following trauma, in neurodegenera-
tive disorders or natural aging [110]. With the notable
exception of the granule cells of the dentate gyrus [111],
the neurons in the human central nervous system shows no
sign of natural turnover [83]. The lack of frank regeneration
of neural tissues of the adult human brain is due partly to its
inability to generate new neurons, a limitation that has
severely hindered the development of therapies for neuro-
logical injury or degeneration (e.g. [112]). Although stem
cells in the SEZ of the human brain appears to cease to
produce cortical neurons under normal condition in vivo,
its cells can be induced to produce neurons in vitro indicat-
ing that the suppression may be exerted by local cellular
milieu [114]. Resistance of differentiated neurons to divi-
sion in vivo seems to be so powerful that no malignancy
derived from mature neurons has yet been recorded in the
adult human cerebrum, whereas gliomas derived from glial
cells are very frequent [113]. Thus, identification of the
inhibitory mechanism preventing neuronal proliferation
could be equally as important for cancer research as it is
for restorative neuroscience. The powerful molecular
mechanisms that prevent neurons in adult mammalian
brain to divide are a complete mystery. Therefore, in an
effort to enhance neurogenesis as a form of therapy in the
brain, we must also to increase our knowledge of how and
why capacity for cell division has declined during evolution.
It is our hope that modern biomedical science can overcome
this naturally evolving impediment while simultaneously
preventing uncontrolled neurogenesis formation of tumors
and preservation of an individual’s identity.
Lack of neuronal turnover in the major structures of the
human brain is not a reason for a decline in our efforts to find
a way to replace cells lost due to disease and aging. To the
contrary, it calls for more intense, sophisticated research
aimed at finding how to overcome the natural resistance
of the human brain to spontaneous regeneration than
hitherto has been possible [1]. Indeed, due to development
of ever more sophisticated methods, there are some good
reasons for hope. For example, there is a growing body of
evidence that embryonic and even adult neuronal stem cells
in the SEZ are highly plastic [115–117]. However, to gener-
ate functionally meaningful substitution therapy it is essen-
tial to reproduce sequentially restriction steps that occur
normally during formation of specific neuronal phenotypes
and their point-to-point connectivity. At present, our ability
to direct either endogenous or exogenous stem cells to
migrate, acquire desired neuronal phenotype and finally
incorporate appropriately into an already established adult
neuro- and synaptoarchitecture in order to restore lost func-
tion in a predictable manner is still rather limited [118,119].
While cells in organs such as the liver (where function does
not depend on positional information of the cells) can be
successfully replaced, the projection neurons in large and
stereotypically organized structures such as the cerebral
cortex, acquire their final site and form long distance synap-
tic connections through precise developmental sequences
[120]. So far, these neurons have been refractory to natural
replacement and regeneration in response to injury.
However, if we discover the genes and factors that inhibit
neurogenesis in the adult human brain, we might eventually
find a way to induce neurons in individuals affected with
specific neurodegenerative disorders or following stroke
and trauma, either by stimulation or blocking inhibition of
mitotic activity or by transplanting neuronal stem cells. As a
first step, we need to learn more how to regulate stem cell
differentiation in an unresponsive environment along speci-
fic neuronal and glial cell fates, and also how to make
appropriate synaptic connections [110,112,121–123]. To
generate such connections in the large adult human brain
may not be a trivial task but, as I have argued elsewhere, the
word impossible is not in the vocabulary of contemporary
neuroscience [110].
Acknowledgements
This work was supported in part by the National Institute of
Heath of the U.S. Public Health Service. I am grateful to my
colleagues J.-P. Bourgeois, S. Brand, T. Bump, M. Cooper, B.
Csillik, J. Dekker, M. Eckenhoff, P. Goldman-Rakic, B. Gran-
ger, B. Gould, N. Halfon, E. Knyhar-Csilik, J. Kordower, D.
Kornack, I. Kostovic, M. LaVail, N. Lenn, P. Levitt, M.
Lidow, R. Nowakowski, M. Ogren, P. Piecinski, D. Rapaport,
D. Schmechel, M. Schwartz, R. Sidman, F. Van Eereburg, G,
Wikmark, N. Zecevic and D. Yasamura who, over the years,
have used the collection of autoradiographs in my laboratory
to make crucial observations as well as engender insightful
discussions about the role of timing in brain development. I
also thank Dr. E. Markakis and J. Breunig for helpful
comments on the manuscript.
References
[1] National Research Council. National Research Council Report. Stem
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–3936

cells and the future of regenerative medicine. Washington, DC:
National Academy Press, 2001.
[2] Kirsche W. Ueber postembryonale matrixzonen im Gehirn
verschiedener Vertebraten und deren Beziehung zur Hirnbauplan-
lehre. Z Forsch 1957;77:313–406.
[3] Jacobson M. Developmental neurobiology. 1st ed. New York:
Plenum, 1970.
[4] Meyer RL. Evidence from thymidine labeling for continuing growth
of retina and tectum in juvenile goldfish. Exp Neurol 1978;59:99–
111.
[5] Goldman SA, Nottebohm F. Neuronal production, migration and
differentiation in a vocal control nucleus of the adult female canary
brain. Proc Natl Acad Sci USA 1983;80:2390–2394.
[6] Perez-Canellas MM, Garcia-Verdugo JM. Adult neurogenesis in the
telencephalon of a lizard: a [3H]thymidine autoradiographic and
bromodeoxyuridine immunocytochemical study. Dev Brain Res
1996;93:49–61.
[7] Horner PJ, Gage FH. Regenerating the damaged central nervous
system. Nature 2000;407:963–970.
[8] Angevine Jr JB. Time of neuron origin in the hippocampal region. An
autoradiographic study in the mouse. Exp Neurol 1965;2(1):Suppl–
70.
[9] Sidman RL. Autoradiographic methods and principles for study of
the nervous system with thymidine-H3. In: Nauta WJH, Ebbeson
SOE, editors. Contemporary research methods in neuroanatomy,
Berlin: Springer, 1970. pp. 252–274.
[10] Bayer SA, Altman J. Neurogenesis and neuronal migration. In: Paxi-
nos G, editor. The rat nervous system, Orlando, FL: Academic Press,
1995. pp. 1041–1078.
[11] Nowakowski RS, Lewin SB, Miller MW. Bromodeoxyuridine
immunohistochemical determination of the lengths of the cell
cycle and the DNA-synthetic phase for an anatomically defined
population. J Neurocytol 1989;18:311–318.
[12] Lois C, Alvarez-Buylla A. Long-distance neuronal migration in the
adult mammalian brain. Science 1994;264:1145–1148.
[13] van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage
FH. Functional neurogenesis in the adult hippocampus. Nature
2002;415:1030–1034.
[14] Kempermann G, Kuhn HG, Gage FH. Genetic influence on neuro-
genesis in the dentate gyrus of adult mice. Proc Natl Acad Sci USA
1997;9:10409–10414.
[15] Kempermann G, Gage FH. Genetic influence on phenotypic differ-
entiation in adult hippocampal neurogenesis. Dev Brain Res
2002;134:1–12.
[16] Napier JR, Napier H. A handbook of living primates. New York:
Academic Press, 1967.
[17] Kornack DR, Rakic P. Changes in cell cycle kinetics during the
development and evolution of primate neocortex. Proc Natl Acad
Sci USA 1998;95:1242–1246.
[18] Nowakowski RS, Rakic P. Decrease of exogenous thymidine-H3 in
the plasma of pregnant rhesus monkeys. Cell Tissue Kinet
1974;7:189–194.
[19] Rakic P. Kinetics of proliferation and latency between final cell
division and onset of differentiation of cerebellar stellate and basket
neurons. J Comp Neurol 1973;147:523–546.
[20] Rakic P. Genesis of the dorsal lateral geniculate nucleus in the rhesus
monkey: site and time of origin, kinetics of proliferation, routes of
migration and pattern of distribution of neurons. J Comp Neurol
1977;176:23–52.
[21] Nowakowski RS, Hayes NL. Stem cells: the promises and pitfalls.
Neuropsychopharmacology 2001;25:799–804.
[22] Rakic P. Adult corticogenesis: an evaluation of the evidence. Nat
Rev Neurosci 2002;3:65–71.
[23] Rakic P. Adult neurogenesis in mammals: an identity crisis. J
Neurosci 2002;22:614–618.
[24] Rakic P, Sidman RL. Subcommissural organ and adjacent epen-
dyma: autoradiographic study of their origin in the mouse brain.
Am J Anat 1968;122:317–335.
[25] Rakic P. Neurons in the monkey visual cortex: systematic relation
between time of origin and eventual disposition. Science
1974;183:425–427.
[26] Rakic P. Early developmental events: cell lineages, acquisitions of
neuronal positions, and areal and laminar development. Neurosci
Res Prog Bull 1982;20:439–451.
[27] Rakic P. Mechanisms of neuronal migration in developing cerebellar
cortex. In: Edelman GE, Cowan WM, Gall E, editors. Molecular
basis of neural development, New York: Wiley, 1985. pp. 139–160.
[28] Rakic P. Limits of neurogenesis in primates. Science 1985;227:154–
156.
[29] Rakic P. DNA synthesis and cell division in the adult primate brain.
Ann NY Acad Sci 1985;457:193–211.
[30] Brand S, Rakic P. Genesis of the primate neostriatum: 3H-thymidine
autoradiographic analysis of the time of neuron origin in the rhesus
monkey. Neuroscience 1979;4:767–778.
[31] Brand S, Rakic P. Neurogenesis of the nucleus accumbens septi and
neighboring septal nuclei in the rhesus monkey: a combined 3H-
thymidine and electron microscopic study. Neuroscience 1980;5:
2125–2138.
[32] Schmechel DE, Rakic P. Arrested proliferation of radial glial cells
during midgestation in rhesus monkey. Nature 1979;227:303–305.
[33] Lenn JN, Halfon N, Rakic P. Development of the interpeduncular
nucleus in the midbrain of rhesus monkey and human. Anat Embryol
1978;152:273–289.
[34] Ogren MP, Rakic P. The ontogenetic development of the pulvinar in
the monkey: morphometric and 3H-thymidine autoradiographic
analyses. Anat Embryol 1981;162:1–20.
[35] Cooper ML, Rakic P. Neurogenetic gradients in the superior and
inferior colliculi of the rhesus monkey. J Comp Neurol 1981;202:
309–334.
[36] Gould BB, Rakic P. The total number, time of origin and kinetics of
proliferation of neurons comprising the deep cerebellar nuclei in the
rhesus monkey. Exp Brain Res 1981;44:195–206.
[37] Rakic P, Nowakowski RS. Time of origin of neurons in the hippo-
campal region of the rhesus monkey. J Comp Neurol 1981;196:99–
124.
[38] Levitt P, Rakic P. The time of genesis, embryonic origin and differ-
entiation of the brainstem monoamine neurons in the rhesus monkey.
Dev Brain Res 1982;4:35–57.
[39] Eckenhoff ME, Rakic P. Nature of the proliferative cells in the
hippocampal dentate gyrus during the life span of the rhesus
monkey. J Neurosci 1988;8:2729–2747.
[40] Kordower JH, Rakic P. Neurogenesis of the magnocellular basal
nuclei in the rhesus monkey. J Comp Neurol 1990;291:637–653.
[41] LaVail MM, Rapaport DH, Rakic P. Cytogenesis in the monkey
retina. J Comp Neurol 1991;309:86–114.
[42] Schwartz ML, Rakic P, Goldman-Rakic PS. Early phenotype expres-
sion of cortical neurons: Evidence that a subclass of migrating
neurons have callosal axons. Proc Natl Acad Sci USA 1991;88:
1354–1358.
[43] Rapaport DH, Fletcher J, LaVail MM, Rakic P. Genesis of
subclasses of neurons in the retinal ganglion cell layer of the
monkey. J Comp Neurol 1992;322:577–588.
[44] Rapaport DH, Rakic P, Yasamura D, LaVail MM. Genesis of the
retinal pigment epithelium in the macaque monkey. J Comp Neurol
1995;363:359–376.
[45] Kordower JH, Piecinski P, Rakic P. Neurogenesis of the amygdalar
complex in the rhesus monkey. Dev Brain Res 1992;68:9–15.
[46] Van Eerdenburg FJCM, Rakic P. Early neurogenesis in the anterior
hypothalamus of the rhesus monkey. Dev Brain Res 1994;79:290–
296.
[47] Granger B, LeSurd AM, Rakic P, Bourgeois J-P. Tempo of neuro-
genesis and synaptogenesis in the primate cingulate mesocortex:
comparison with the neocortex. J Comp Neurol 1995;360:363–376.
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–39 37

[48] Knyhar-Csillik E, Rakic P, Csillik B. Development of glomerular
complexes and immunocytochemical differentiation in the superfi-
cial dorsal horn of the embryonic primate spinal cord. Anat Embryol
1999;199:125–148.
[49] Zecevic N, Rakic P. Development of layer I neurons in the primate
cerebral cortex. J Neurosci 2001;21:5607–5619.
[50] Rakic P, Riley KP. Overproduction and elimination of retinal axons
in the fetal rhesus monkey. Science 1983;209:1441–1444.
[51] Williams RW, Rakic P. Elimination of neurons in the rhesus
monkey’s lateral geniculate nucleus during development. J Comp
Neurol 1988;272:424–436.
[52] Rakic P. A small step for the cell – a giant leap for mankind: a
hypothesis of neocortical expansion during evolution. Trends
Neurosci 1995;18:383–388.
[53] Goldman-Rakic PS. Circuitry of primate prefrontal cortex and regu-
lation of behavior by representational memory. In: Plum F, editor.
Handbook of physiology, The nervous system, higher functions of
the brain, vol. V section I part 1. Bethesda, MD: American Physio-
logical Society, 1987. pp. 373–417.
[54] Mountcastle V. The evolution of ideas concerning the function of the
neocortex. Cereb Cortex 1995;5:289–295.
[55] Rakic P. Differences in the time of origin and in eventual distribution
of neurons in areas 17 and 18 of the visual cortex in the rhesus
monkey. Exp Brain Res Suppl 1976;1:244–248.
[56] Dekker JJ, Rakic P. Genesis of the motor cortex and VA-VL Thala-
mic complex in rhesus monkey. Abstr Soc Neurosci 1980;6:205.
[57] Angevine Jr JB, Sidman RL. Autoradiographic study of cell migra-
tion during histogenesis of cerebral cortex in the mouse. Nature
1961;192:766–768.
[58] Caviness Jr V, Sidman RL. Time of origin of corresponding cell
classes in the cerebral cortex of normal and reeler mutant mice: an
autoradiographic analysis. J Comp Neurol 1973;148:141–152.
[59] Smart IM, Smart M. Growth patterns in the lateral wall of the mouse
telencephalon. I. Autoradiographic studies of the histogenesis of the
isocortex and adjacent areas. J Anat (Lond) 1982;134:273–298.
[60] Rakic P. Mode of cell migration to the superficial layers of fetal
monkey neocortex. J Comp Neurol 1972;145:61–84.
[61] Rakic P. Timing of major ontogenetic events in the visual cortex of
the rhesus monkey. In: Buchwald NA, Brazier M, editors. Brain
mechanisms in mental retardation, New York: Academic Press,
1975. pp. 3–40.
[62] de Carlos JA, Lopez-Mascaraque L, Valverde F. Dynamics of cell
migration from the lateral ganglionic eminence in the rat. J Neurosci
1996;16:6146–6156.
[63] Powel EM, Mara WM, Levitt P. Hepatocyte growth factor/scatter
factor is mitogen for interneuron migrating from the ventral to dorsal
telencephalon. Neuron 2001;30:1–20.
[64] Anderson S, Mione M, Yun K, Rubenstein JLR. Differential origins
of neocortical projection and local circuit neurons: role of Dlx genes
in neocortical interneuronogenesis. Cereb Cortex 1999;9:646–654.
[65] Lavdas AA, Grigoriou M, Pachnis V, Parnavelas JG. The medial
ganglionic eminence gives rise to a population of early neurons in
the developing cerebral cortex. J Neurosci 1999;19:7881–7888.
[66] Wichterle H, Garcia-Verdugo JM, Herrera DG, Alvarez-Buylla A.
Young neurons from medial ganglionic eminence disperse in adult
and embryonic brain. Nat Neurosci 1999;2:461–466.
[67] Marin O, Rubenstein JL. A long, remarkable journey: tangential
migration in the telencephalon. Nat Rev Neurosci 2001;2:780–790.
[68] Letinic K, Zoncu R, Rakic P. Origin of GABAergic neurons in the
human neocortex. Nature 2002 in press.
[69] Berry M, Rogers AW. The migration of neuroblasts in the develop-
ing cortex. J Anat 1965;99:691–709.
[70] Hicks SP, D’Amato CJ. Cell migrations to the isocortex in the rat.
Anat Rec 1968;160:619–634.
[71] Luskin MB, Shatz CJ. Neurogenesis of the cat’s primary visual
cortex. J Comp Neurol 1985;242:611–631.
[72] Jackson CA, Peduzzi JD, Hickey TL. Visual-cortex development in
the ferret. 1. Genesis and migration of visual cortical-neurons. J
Neurosci 1989;9:1242–1253.
[73] Takahashi T, Nowakowski RS, Caviness VS. The leaving or Q frac-
tion of the murine cerebral proliferative epithelium: a general model
of neocortical neuronogenesis. J Neurosci 1996;16:6183–6196.
[74] Magavi SS, Leavitt BR, Macklis JD. Induction of neurogenesis in
the neocortex of adult mice. Nature 2000;405:951–955.
[75] Kornack DR, Rakic P. Continuation of neurogenesis in the hippo-
campus of the adult macaque monkey. Proc Natl Acad Sci USA
1999;96:5768–5773.
[76] Kuhn HG, Palmer TD, Fuchs E. Adult neurogenesis: a compensatory
mechanism for neuronal damage. Eur Arch Psy Clin N 2001;251:152–
158.
[77] Kornack D, Rakic P. Generation and migration of new olfactory
neurons in adult primates. Proc Natl Acad Sci USA 2001;98:
4752–4757.
[78] Luskin MB. Restricted proliferation and migration of postnatally
generated neurons derived from the forebrain subventricular zone.
Neuron 1993;1:173–189.
[79] Wu W, Wong K, Jin-hui Chen J, Jiang Z, Dupuis S, Wu SJ, Yi Rao
Y. Directional guidance of neuronal migration in the olfactory
system by the protein Slit. Nature 1999;400:331–336.
[80] Kappers AU, Huber GC, Crosby EK. The comparative anatomy of
the nervous system of vertebrates, including man. New York:
Hafner, 1965.
[81] Nieuwenhuys R, ten Donkelaar HJ, Nicholson C. The central
nervous system of vertebrates. Berlin: Springer, 1998.
[82] Kornack RD, Rakic P. Cell proliferation without neurogenesis in
adult primate neocortex. Science 2001;294:2127–2130.
[83] Seress L, Abraham H, Tornoczky T, Kosztolanyi G. Cell formation
in the human hippocampal formation from mid-gestation to the late
postnatal period. Neuroscience 2001;105:831–843.
[84] Gould E, Reeves AJ, Graziano MS, Gross CG. Neurogenesis in the
neocortex of adult primates. Science 1999;286:548–552.
[85] Gould E, Vail N, Wagers M, Gross C. Adult-generated hippocampal
and neocortical neurons in macaques have a transient existence. Proc
Natl Acad Sci USA 2001;98:10910–10917.
[86] McDermott KWG, Lantos PL. Cell proliferation in the subependy-
mal layer of the postnatal marmoset, Callithrix jacchus. Dev Brain
Res 1990;57:269–277.
[87] Lewis PD. Mitotic activity in the primate subependymal layer and
the genesis of the gliomas. Nature 1968;217:974–975.
[88] Nowakowski RS, Hayes NL. New neurons: extraordinary evidence
or extraordinary conclusion. Science 2000;288:771a see details in
Technical Comment on-line: http//www.sciencemag.org/cgi/
content/ful/288/5467/771a?.
[89] Yang Y, Geldmacher DS, Herrup K. DNA replication precedes neuro-
nal cell death in Alzheimer’s disease. J Neurosci 2001;15:2661–2668.
[90] Morris SM. The genetic toxicology of 5-bromodeoxyuridine in
mammalian-cells. Mutat Res 1991;258:161–188.
[91] Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz DM, Nakano Y,
et al. Bone marrow cells adopt the phenotype of other cells by
spontaneous cell fusion. Nature 2002;416:542–545.
[92] Ying Q-L, Nichols J, Evans EP, Smith AG. Changing potency by
spontaneous fusion. Nature 2002;416:545–548.
[93] Kretschmann HJ. Brain growth, Basel: Karger, 1986. pp. 1–140.
[94] Poliakov GI. Development of the child’s brain. Leningrad: Medi-
cina, 1965 in Russian.
[95] Sidman RL, Rakic P. Development of the human central nervous
system. In: Haymaker W, Adams RD, editors. Histology and histo-
pathology of the nervous system, Springfield, IL: Thomas, 1982. pp.
3–145.
[96] Bourgeois J-P, Rakic P. Changing of synaptic density in the primary
visual cortex of the rhesus monkey from fetal to adult stage. J
Neurosci 1993;13:2801–2820.
[97] Bourgeois J-P, Goldman-Rakic PS, Rakic P. Synaptogenesis in the
prefrontal cortex of rhesus monkey. Cereb Cortex 1994;4:78–96.
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–3938

[98] Huttenlocher PR, Dabholkar AS. Regional differences in synapto-
genesis in human cerebral cortex. . J. Comp Neurol 1997;387:167–
178.
[99] Levitt P, Rakic P. Immunoperoxidase localization of glial fibrillary
acid protein in radial glial cells and astrocytes of the developing
rhesus monkey brain. J Comp Neurol 1980;193:815–840.
[100] Levitt P, Cooper ML, Rakic P. Coexistence of neuronal and glial
precursor cells in the cerebral ventricular zone of the fetal monkey:
an ultrastructural immunoperoxidase analysis. J Neurosci
1981;1:27–39.
[101] Schmechel DE, Rakic P. A Golgi study of radial glial cells in devel-
oping monkey telencephalon. Anat Embryol 1979;156:1115–1152.
[102] Yakovlev PI, Lecours AR. The myelogenetic cycles of regional
maturation of the brain. In: Minkowski A, editor. Regional devel-
opment of the brain in early life, Oxford: Blackwell, 1967. pp. 3–70.
[103] Sidman RL, Rakic P. Neuronal migration with special reference to
developing human brain: a review. Brain Res 1973;62:1–35.
[104] Leblond CP. Classification of cell populations on the basis of their
proliferative behavior. Natl Cancer Inst Monogr 1964;14:119–150.
[105] Goss RJ. Principles of regeneration. New York: Academic Press,
1969.
[106] Wallace H. Vertebrate limb regeneration. New York: Wiley, 1981.
[107] Goss RJ. Modes of growth and regeneration. In: Falkner F, Tanner
JM, editors. Human growth, New York: Plenum, 1980. pp. 3–51.
[108] Yannas IV. Tissue and organ regeneration in adults. Berlin:
Springer, 2001.
[109] Scharff C, Kirn JR, Grossman M, Macklis JD, Nottebohm F.
Targeted neuronal death affects neuronal replacement and vocal
behavior in adult songbirds. Neuron 2000;25:481–492.
[110] Rakic P. Young neurons for old brains? Nat Neurosci 1998;1:643–
645.
[111] Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg
C, Peterson DA, et al. Neurogenesis in the adult human hippocam-
pus. Nat Med 1998;1:1313–1317.
[112] Pincus DW, Goodman RR, Fraser RAR, Nedergaard M, Goldman
SA. Neural stem and progenitor cells: a strategy for gene therapy and
brain repair. Neurosurgery 1998;42:858–867.
[113] Kirschenbaum B, Nedergaard M, Preuss A, Barami IK, Fraser RAR,
Goldman SA. In-vitro neuronal production and differentiation by
precursor cells derived from the adult human forebrain. Cereb
Cortex 1994;4:576–589.
[114] Russell DS, Rubinstein LJ. Pathology of tumours of the nervous
system. Baltimore, MD: Williams and Wilkins, 1963.
[115] Anderson JD, Gage FH, Weissman IL. Can stem cells cross lineage
boundaries. Nat Med 2001;7:393–395.
[116] Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla
A. Subventricular zone astrocytes are neural stem cells in adult
mammalian brain. Cell 1999;97:703–716.
[117] Laywell ED, Rakic P, Kukekov VG, Holland EC, Steindler DA. An
identification of a multipotent astrocytic stem cell in the immature
and adult mouse brain. Proc Natl Acad Sci USA 2000;97:13883–
13888.
[118] Morrison SJ. Neuronal differentiation: proneural genes inhibit glio-
genesis. Curr Biol 2001;11:R349–R351.
[119] Rossi F, Cattaneo E. Neural stem cell therapy for neurological
disease: dreams and reality. Nat Rev Neurosci 2002;3:401–409.
[120] Rakic P. Specification of cerebral cortical areas. Science
1988;241:170–176.
[121] Bjorklund A, Lindvall O. Cell replacement therapies for central
nervous system disorders. Nat Neurosci 2000;3:537–544.
[122] Gage FH. Mammalian neural stem cells. Science 2000;287:1433–
1438.
[123] McKay R. Stem cells – hype and hope. Nature 1988;406:361–364.
P. Rakic / Clinical Neuroscience Research 2 (2002) 29–39 39





























