Embed Size (px)
Citation preview

Posttranslational modifications of Rab proteins causeeffective displacement of GDP dissociation inhibitorLena K. Oesterlina, Roger S. Goodya, and Aymelt Itzena,b,1
aDepartment of Physical Biochemistry, Max Planck Institute of Molecular Physiology, Otto-Hahn-Strasse 11, 44227 Dortmund, Germany; and bCenter forIntegrated Protein Science Munich (CIPSM), Chemistry Department, Technische Universität München, Lichtenbergstrasse 4, 85747 Garching, Germany
Edited by James A. Spudich, Stanford University School of Medicine, Stanford, CA, and approved January 28, 2012 (received for review December 21, 2011)
Intracellular vesicular trafficking is regulated by approximately60 members of the Rab subfamily of small Ras-like GDP/GTP bind-ing proteins. Rab proteins cycle between inactive and active statesas well as between cytosolic and membrane bound forms. Mem-brane extraction/delivery and cytosolic distribution of Rabs ismediated by interactionwith the protein GDP dissociation inhibitor(GDI) that binds to prenylated inactive (GDP-bound) Rab proteins.Because the Rab:GDP:GDI complex is of high affinity, the questionarises of how GDI can be displaced efficiently from Rab protein inorder to allow the necessary recruitment of the Rab to its specifictarget membrane. While there is strong evidence that DrrA, as abacterially encoded GDP/GTP exchange factor, contributes to thisevent, we show here that posttranslational modifications of Rabscan also modulate the affinity for GDI and thus cause effective dis-placement of GDI from Rab:GDI complexes. These activities havebeen found associated with the phosphocholination and adenyly-lation activities of the enzymes AnkX and DrrA/SidM, respectively,from the pathogenic bacterium Legionella pneumophila. Bothmodifications occur after spontaneous dissociation of Rab:GDIcomplexes within their natural equilibrium. Therefore, the effec-tive GDI displacement that is observed is caused by inhibition ofreformation of Rab:GDI complexes. Interestingly, in contrast toadenylylation by DrrA, AnkX can covalently modify inactive Rabswith high catalytic efficiency even when GDP is bound to theGTPase and hence can inhibit binding of GDI to Rab:GDP com-plexes. We therefore speculate that human cells could employ si-milar mechanisms in the absence of infection to effectively displaceRabs from GDI.
Legionnaires’ disease ∣ Rab recruitment ∣ RabGDI
The intracellular activity of most signaling proteins is regulatedat the temporal and spatial level. In the regulation of vesicular
trafficking, small Ras-like GTPases of the Rab subfamily are con-trolled in their distribution between the cytosol and intracellularmembranes, which is in turn coupled to the binding to eitherGDP or GTP. Cycling between the cytosol and membranes is de-pendent on the activation state of the Rab protein. In the activeGTP-bound form, Rabs are associated with membranes, wherethey interact with effector proteins that promote diverse stepsin vesicular trafficking. When bound to GDP, Rabs are definedas inactive and are predominantly distributed in the cytosolbound to GDI. In order to associate with the cytosolic surfaceof intracellular membranes, most Rab proteins possess two hy-drophobic prenyl (geranylgeranyl) moieties at their structurallyflexible C terminus that are posttranslationally attached via theconcerted action of Rab geranylgeranyl transferase (RabGG-Tase) and Rab escort protein utilizing geranylgeranyl pyropho-sphate as a cosubstrate. These geranylgeranyl groups mediatestable peripheral binding of Rabs to a cognate membrane by in-serting the hydrophobic anchors into the lipid bilayer (1). Activa-tion of Rab proteins is catalyzed by GDP/GTP exchange factors(GEFs) that displace the tightly bound GDP by the excess ofGTP from the cellular environment. Because most active Rabproteins do not hydrolyze GTP on a physiologically relevant timescale, GTPase activating proteins assist in deactivating Rabs by
stimulating their intrinsic GTPase activity and thereby generatingGDP bound Rab.
Because Rab proteins cycle not only between the GDP andGTP states but also between the cytosolic and membrane asso-ciated forms, another regulatory factor is necessary to competewith the high binding affinity between prenylated Rabs and lipidbilayers. In mammalian cells, two isoforms of the GDP dissocia-tion inhibitor are capable of binding ca. 60 mammalian Rabs intheir prenylated form with high affinity, thereby extracting theprenylated Rabs from membranes and solubilizing them in theform of Rab:GDP:GDI complexes. The stability of these com-plexes is only high (Kd in the nanomolar range) if the Rab proteinis both prenylated and in its inactive state. When active and/orunprenylated, this affinity is decreased by 3–4 orders of magni-tude to a high micromolar Kd (2, 3). The recruitment of Rabsfrom the pool of soluble cytosolic Rab:GDP:GDI complexes toa specific membrane thus requires the dissociation of the tightlybound GDI prior to the initiation of a certain vesicular transportstep via generation of active, GTP-bound Rabs on the membrane.Two models have been put forward to explain how the effectivedissociation of Rab:GDI complexes can be achieved.
Membrane association of the two endosomal Rab proteinsRab5 and Rab9 is accompanied by nucleotide exchange that oc-curs shortly after membrane association (4, 5). These results ledto the assumption that a GDI displacement factor (GDF) couldinfluence the release of prenylated Rab proteins from GDI (6).The only identified GDI displacement factor to date is Yip3/PRA1 (7, 8). Yip3/PraI can inhibit the extraction of Rab3 frommembranes (7) and furthermore has been shown to influence thedissociation of a Rab9:GDI complex and to facilitate the mem-brane recruitment of Rab9 from its complex with GDI in in vitroextraction assays (8). In vivo, Yip3/PRA1 depletion by RNAileads to reduced membrane association of Rab9 (8). Based onthese data a GDF would be defined as a protein that can influ-ence the equilibrium between GDI bound and free Rab proteins.
Recently, we have demonstrated that the GEF domain ofDrrA is sufficient to effectively displace GDI from Rab:GDP:GDI complexes by generating the GTP state of Rab proteins aftercomplex dissociation, which has only low affinity for GDI (3, 9).It has been shown that the protein DrrA/SidM secreted by theintracellular bacterial pathogen Legionella pneumophila can effi-ciently recruit Rab1 to the Legionella containing vacuoles inwhich the bacterium resides and replicates intracellularly (10, 11).Rab1 recruitment and GDI displacement is exerted by the GEF-activity of DrrA (9), suggesting that mammalian GEFs may alsobe capable of activating and thereby recruiting Rab proteins from
Author contributions: R.S.G. and A.I. designed research; L.K.O. conducted experiments;L.K.O., R.S.G., and A.I. analyzed the data; and L.K.O., R.S.G., and A.I. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
See Commentary on page 5555.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1121161109/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1121161109 PNAS ∣ April 10, 2012 ∣ vol. 109 ∣ no. 15 ∣ 5621–5626
BIOCH
EMISTR
YSE
ECO
MMEN
TARY
their complexes with GDI to membranes where specific GEFs arelocalized.
This model requires that the Rab protein dissociates fromGDIbefore activation by GEFs. However, the intrinsic rate of GDIdissociation is low due to the high affinity between prenylatedRab:GDP and GDI (3, 9, 12) and hence limits the rate at whichactive Rab can be generated. Thus, GDI dissociation can poten-tially become rate limiting for the initiation of vesicular traffick-ing processes. In addition to the possibility that bona fide GDFmolecules exist that can accelerate this process, we wonderedwhether inactive Rab bound to membranes could be generatedby different mechanisms and that this pool of Rab:GDP couldbe mobilized rapidly. Here we show that two posttranslationalmodifications (by AnkX and DrrA) can drastically reduce theaffinity of different Rab proteins for GDI. Furthermore we showthat AnkX can dissociate Rab:GDI complexes efficiently, be-cause it modifies even inactive, GDP-bound Rab proteins thatspontaneously dissociate from the Rab:GDP:GDI complex withhigh catalytic activity and inhibits their rebinding to GDI via thismodification.
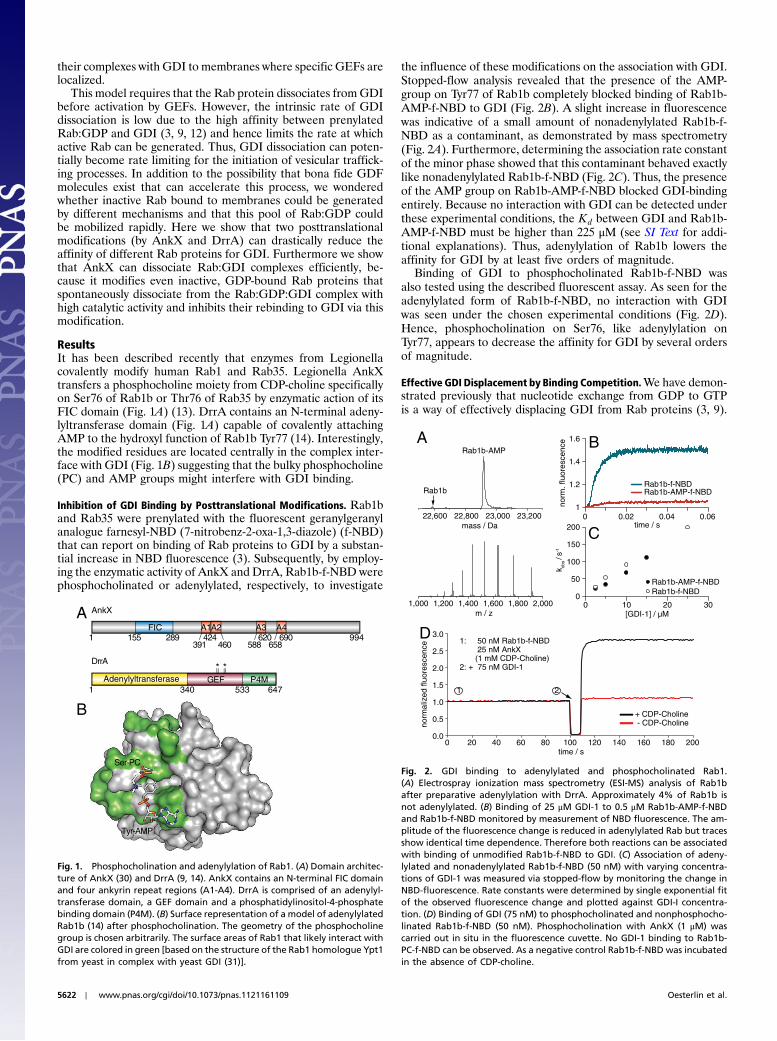
ResultsIt has been described recently that enzymes from Legionellacovalently modify human Rab1 and Rab35. Legionella AnkXtransfers a phosphocholine moiety from CDP-choline specificallyon Ser76 of Rab1b or Thr76 of Rab35 by enzymatic action of itsFIC domain (Fig. 1A) (13). DrrA contains an N-terminal adeny-lyltransferase domain (Fig. 1A) capable of covalently attachingAMP to the hydroxyl function of Rab1b Tyr77 (14). Interestingly,the modified residues are located centrally in the complex inter-face with GDI (Fig. 1B) suggesting that the bulky phosphocholine(PC) and AMP groups might interfere with GDI binding.
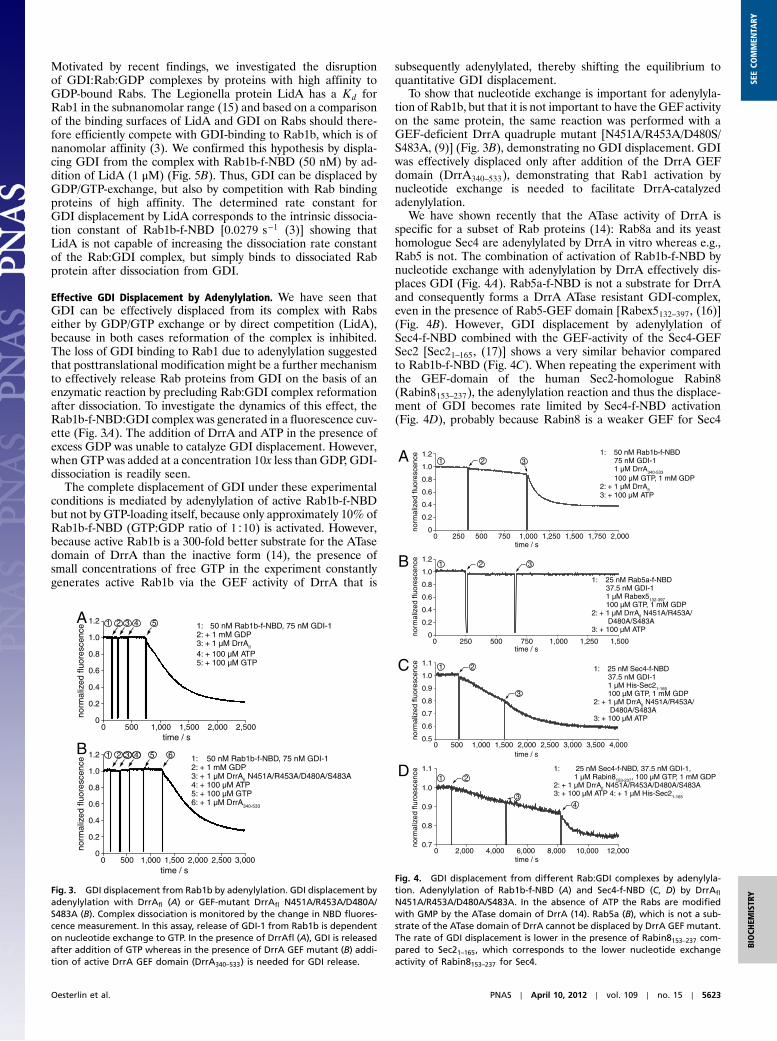
Inhibition of GDI Binding by Posttranslational Modifications. Rab1band Rab35 were prenylated with the fluorescent geranylgeranylanalogue farnesyl-NBD (7-nitrobenz-2-oxa-1,3-diazole) (f-NBD)that can report on binding of Rab proteins to GDI by a substan-tial increase in NBD fluorescence (3). Subsequently, by employ-ing the enzymatic activity of AnkX and DrrA, Rab1b-f-NBD werephosphocholinated or adenylylated, respectively, to investigate
the influence of these modifications on the association with GDI.Stopped-flow analysis revealed that the presence of the AMP-group on Tyr77 of Rab1b completely blocked binding of Rab1b-AMP-f-NBD to GDI (Fig. 2B). A slight increase in fluorescencewas indicative of a small amount of nonadenylylated Rab1b-f-NBD as a contaminant, as demonstrated by mass spectrometry(Fig. 2A). Furthermore, determining the association rate constantof the minor phase showed that this contaminant behaved exactlylike nonadenylylated Rab1b-f-NBD (Fig. 2C). Thus, the presenceof the AMP group on Rab1b-AMP-f-NBD blocked GDI-bindingentirely. Because no interaction with GDI can be detected underthese experimental conditions, the Kd between GDI and Rab1b-AMP-f-NBD must be higher than 225 μM (see SI Text for addi-tional explanations). Thus, adenylylation of Rab1b lowers theaffinity for GDI by at least five orders of magnitude.
Binding of GDI to phosphocholinated Rab1b-f-NBD wasalso tested using the described fluorescent assay. As seen for theadenylylated form of Rab1b-f-NBD, no interaction with GDIwas seen under the chosen experimental conditions (Fig. 2D).Hence, phosphocholination on Ser76, like adenylylation onTyr77, appears to decrease the affinity for GDI by several ordersof magnitude.
Effective GDI Displacement by Binding Competition.We have demon-strated previously that nucleotide exchange from GDP to GTPis a way of effectively displacing GDI from Rab proteins (3, 9).
*
Ser-PC
FIC9941
A
6471GEF P4MAdenylyltransferase
*
B
Tyr-AMP
A1A2 A3 A4155 289
391424
460 588620 690
658
340 533
DrrA
AnkX
Fig. 1. Phosphocholination and adenylylation of Rab1. (A) Domain architec-ture of AnkX (30) and DrrA (9, 14). AnkX contains an N-terminal FIC domainand four ankyrin repeat regions (A1-A4). DrrA is comprised of an adenylyl-transferase domain, a GEF domain and a phosphatidylinositol-4-phosphatebinding domain (P4M). (B) Surface representation of a model of adenylylatedRab1b (14) after phosphocholination. The geometry of the phosphocholinegroup is chosen arbitrarily. The surface areas of Rab1 that likely interact withGDI are colored in green [based on the structure of the Rab1 homologue Ypt1from yeast in complex with yeast GDI (31)].
A
1,000 1,6001,400m / z
2,0001,200 1,800
22,600 23,20023,000mass / Da
22,800
Rab1b
Rab1b-AMP
D3.0
0.5
1.5
1.0
2.5
2.0
0.020018016040200 60 80 120 140
21
1: 50 nM Rab1b-f-NBD 25 nM AnkX (1 mM CDP-Choline)2: + 75 nM GDI-1
+ CDP-Choline - CDP-Choline
1.6
1
Rab1b-f-NBDRab1b-AMP-f-NBD
1.2
1.4
norm
. flu
ores
cenc
e B
0.060 0.040.02time / s
0 2010[GDI-1] / µM
30
Rab1b-AMP-f-NBDRab1b-f-NBD
50
100
150
k obs /
s-1
200
0
C
100time / s
norm
aliz
ed fl
uore
scen
ce
Fig. 2. GDI binding to adenylylated and phosphocholinated Rab1.(A) Electrospray ionization mass spectrometry (ESI-MS) analysis of Rab1bafter preparative adenylylation with DrrA. Approximately 4% of Rab1b isnot adenylylated. (B) Binding of 25 μM GDI-1 to 0.5 μM Rab1b-AMP-f-NBDand Rab1b-f-NBD monitored by measurement of NBD fluorescence. The am-plitude of the fluorescence change is reduced in adenylylated Rab but tracesshow identical time dependence. Therefore both reactions can be associatedwith binding of unmodified Rab1b-f-NBD to GDI. (C) Association of adeny-lylated and nonadenylylated Rab1b-f-NBD (50 nM) with varying concentra-tions of GDI-1 was measured via stopped-flow by monitoring the change inNBD-fluorescence. Rate constants were determined by single exponential fitof the observed fluorescence change and plotted against GDI-I concentra-tion. (D) Binding of GDI (75 nM) to phosphocholinated and nonphosphocho-linated Rab1b-f-NBD (50 nM). Phosphocholination with AnkX (1 μM) wascarried out in situ in the fluorescence cuvette. No GDI-1 binding to Rab1b-PC-f-NBD can be observed. As a negative control Rab1b-f-NBD was incubatedin the absence of CDP-choline.
5622 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1121161109 Oesterlin et al.
Motivated by recent findings, we investigated the disruptionof GDI:Rab:GDP complexes by proteins with high affinity toGDP-bound Rabs. The Legionella protein LidA has a Kd forRab1 in the subnanomolar range (15) and based on a comparisonof the binding surfaces of LidA and GDI on Rabs should there-fore efficiently compete with GDI-binding to Rab1b, which is ofnanomolar affinity (3). We confirmed this hypothesis by displa-cing GDI from the complex with Rab1b-f-NBD (50 nM) by ad-dition of LidA (1 μM) (Fig. 5B). Thus, GDI can be displaced byGDP/GTP-exchange, but also by competition with Rab bindingproteins of high affinity. The determined rate constant forGDI displacement by LidA corresponds to the intrinsic dissocia-tion constant of Rab1b-f-NBD [0.0279 s−1 (3)] showing thatLidA is not capable of increasing the dissociation rate constantof the Rab:GDI complex, but simply binds to dissociated Rabprotein after dissociation from GDI.
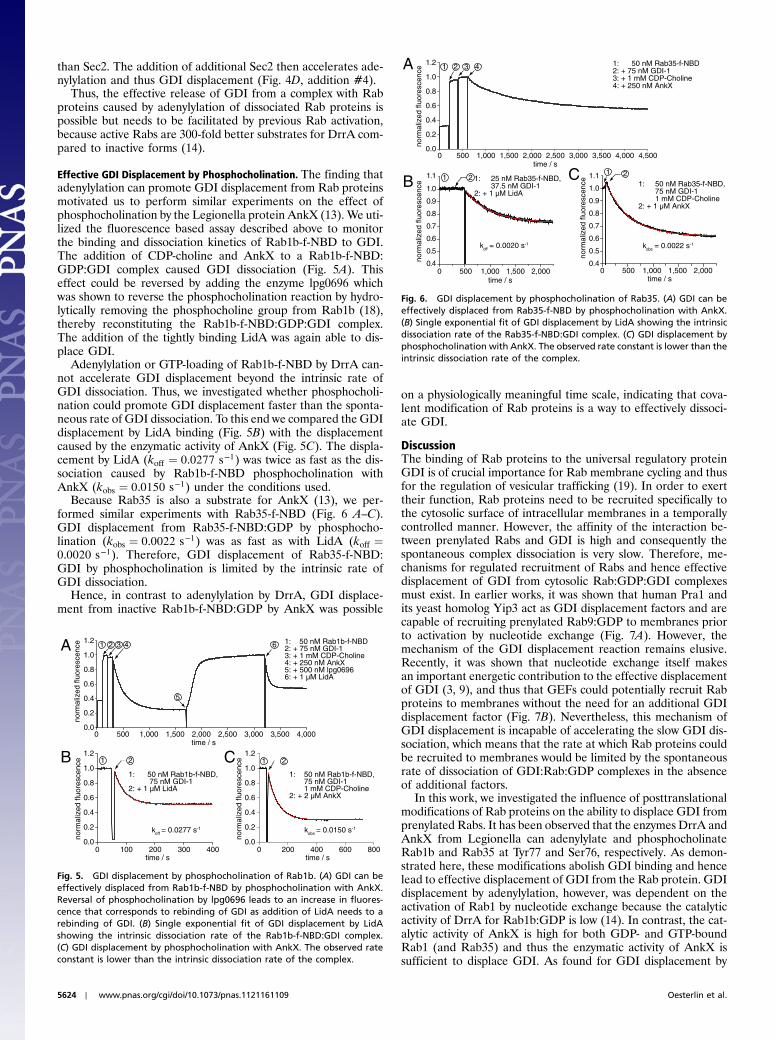
Effective GDI Displacement by Adenylylation. We have seen thatGDI can be effectively displaced from its complex with Rabseither by GDP/GTP exchange or by direct competition (LidA),because in both cases reformation of the complex is inhibited.The loss of GDI binding to Rab1 due to adenylylation suggestedthat posttranslational modification might be a further mechanismto effectively release Rab proteins from GDI on the basis of anenzymatic reaction by precluding Rab:GDI complex reformationafter dissociation. To investigate the dynamics of this effect, theRab1b-f-NBD:GDI complex was generated in a fluorescence cuv-ette (Fig. 3A). The addition of DrrA and ATP in the presence ofexcess GDP was unable to catalyze GDI displacement. However,when GTP was added at a concentration 10x less than GDP, GDI-dissociation is readily seen.
The complete displacement of GDI under these experimentalconditions is mediated by adenylylation of active Rab1b-f-NBDbut not by GTP-loading itself, because only approximately 10% ofRab1b-f-NBD (GTP:GDP ratio of 1∶10) is activated. However,because active Rab1b is a 300-fold better substrate for the ATasedomain of DrrA than the inactive form (14), the presence ofsmall concentrations of free GTP in the experiment constantlygenerates active Rab1b via the GEF activity of DrrA that is
subsequently adenylylated, thereby shifting the equilibrium toquantitative GDI displacement.
To show that nucleotide exchange is important for adenylyla-tion of Rab1b, but that it is not important to have the GEFactivityon the same protein, the same reaction was performed with aGEF-deficient DrrA quadruple mutant [N451A/R453A/D480S/S483A, (9)] (Fig. 3B), demonstrating no GDI displacement. GDIwas effectively displaced only after addition of the DrrA GEFdomain (DrrA340–533), demonstrating that Rab1 activation bynucleotide exchange is needed to facilitate DrrA-catalyzedadenylylation.
We have shown recently that the ATase activity of DrrA isspecific for a subset of Rab proteins (14): Rab8a and its yeasthomologue Sec4 are adenylylated by DrrA in vitro whereas e.g.,Rab5 is not. The combination of activation of Rab1b-f-NBD bynucleotide exchange with adenylylation by DrrA effectively dis-places GDI (Fig. 4A). Rab5a-f-NBD is not a substrate for DrrAand consequently forms a DrrA ATase resistant GDI-complex,even in the presence of Rab5-GEF domain [Rabex5132–397, (16)](Fig. 4B). However, GDI displacement by adenylylation ofSec4-f-NBD combined with the GEF-activity of the Sec4-GEFSec2 [Sec21–165, (17)] shows a very similar behavior comparedto Rab1b-f-NBD (Fig. 4C). When repeating the experiment withthe GEF-domain of the human Sec2-homologue Rabin8(Rabin8153–237), the adenylylation reaction and thus the displace-ment of GDI becomes rate limited by Sec4-f-NBD activation(Fig. 4D), probably because Rabin8 is a weaker GEF for Sec4
A
B 51.2 6 1: + 50 nM Rab1b-f-NBD, 75 nM GDI-12: + 1 mM GDP3: + 1 µM DrrAfl N451A/R453A/D480A/S483A4: + 100 µM ATP 5: + 100 µM GTP6: + 1 µM DrrA340-533
0 1,000 2,000 2,500500 1,500time / s
3,000
time / s
1 2 3 4
5 1:+ 50 nM Rab1b-f-NBD, 75 nM GDI-12: + 1 mM GDP3: + 1 µM DrrAfl
4: + 100 µM ATP 5: + 100 µM GTP
500
1 2
0 1,000 1,500 2,000 2,500
0
0.2
0.4
0.6
0.8
1.0
norm
aliz
ed fl
uore
scen
ce
3 4
0
0.2
0.4
0.6
0.8
1.0
1.2
norm
aliz
ed fl
uore
scen
ce
Fig. 3. GDI displacement from Rab1b by adenylylation. GDI displacement byadenylylation with DrrAfl (A) or GEF-mutant DrrAfl N451A/R453A/D480A/S483A (B). Complex dissociation is monitored by the change in NBD fluores-cence measurement. In this assay, release of GDI-1 from Rab1b is dependenton nucleotide exchange to GTP. In the presence of DrrAfl (A), GDI is releasedafter addition of GTP whereas in the presence of DrrA GEF mutant (B) addi-tion of active DrrA GEF domain (DrrA340–533) is needed for GDI release.
3
1: 25 nM Rab5a-f-NBD 37.5 nM GDI-1 1 µM Rabex5132-397 100 µM GTP, 1 mM GDP2: + 1 µM DrrAfl N451A/R453A/ D480A/S483A3: + 100 µM ATP
1,2501,000time / s
750
A
0
0.2
0.4
0.6
0.8
1.0
1.2
1,5000 500250
B 1 2
3
1: 25 nM Sec4-f-NBD, 37.5 nM GDI-1, 1 µM Rabin8153-237, 100 µM GTP, 1 mM GDP2: + 1 µM DrrAfl N451A/R453A/D480A/S483A3: + 100 µM ATP 4: + 1 µM His-Sec21-165
10,0008,0006,000
4
norm
aliz
ed fl
uore
scen
ce
1.1
0 2,000
1 2
norm
aliz
ed fl
uroe
scen
ce
0.7
0.8
0.9
1.0
12,0004,000
1,7501,5001,2501,000time / s
750
1: 50 nM Rab1b-f-NBD 75 nM GDI-1 1 µM DrrA340-533
100 µM GTP, 1 mM GDP2: + 1 µM DrrAfl3: + 100 µM ATP
31 2
norm
aliz
ed fl
uore
scen
ce
0
0.2
0.4
0.6
0.8
1.0
1.2
2,0000 500250
3
1: 25 nM Sec4-f-NBD 37.5 nM GDI-1 1 µM His-Sec21-165 100 µM GTP, 1 mM GDP2: + 1 µM DrrAfl N451A/R453A/ D480A/S483A3: + 100 µM ATP
3,5003,000time / s
2,500
1.1
4,0000 2,0001,500
1 2C
D
time / s
norm
aliz
ed fl
uore
scen
ce
0.5
0.6
0.7
0.8
0.9
1.0
1,000500
Fig. 4. GDI displacement from different Rab:GDI complexes by adenylyla-tion. Adenylylation of Rab1b-f-NBD (A) and Sec4-f-NBD (C, D) by DrrAfl
N451A/R453A/D480A/S483A. In the absence of ATP the Rabs are modifiedwith GMP by the ATase domain of DrrA (14). Rab5a (B), which is not a sub-strate of the ATase domain of DrrA cannot be displaced by DrrA GEF mutant.The rate of GDI displacement is lower in the presence of Rabin8153–237 com-pared to Sec21–165, which corresponds to the lower nucleotide exchangeactivity of Rabin8153–237 for Sec4.
Oesterlin et al. PNAS ∣ April 10, 2012 ∣ vol. 109 ∣ no. 15 ∣ 5623
BIOCH
EMISTR
YSE
ECO
MMEN
TARY
than Sec2. The addition of additional Sec2 then accelerates ade-nylylation and thus GDI displacement (Fig. 4D, addition #4).
Thus, the effective release of GDI from a complex with Rabproteins caused by adenylylation of dissociated Rab proteins ispossible but needs to be facilitated by previous Rab activation,because active Rabs are 300-fold better substrates for DrrA com-pared to inactive forms (14).
Effective GDI Displacement by Phosphocholination. The finding thatadenylylation can promote GDI displacement from Rab proteinsmotivated us to perform similar experiments on the effect ofphosphocholination by the Legionella protein AnkX (13). We uti-lized the fluorescence based assay described above to monitorthe binding and dissociation kinetics of Rab1b-f-NBD to GDI.The addition of CDP-choline and AnkX to a Rab1b-f-NBD:GDP:GDI complex caused GDI dissociation (Fig. 5A). Thiseffect could be reversed by adding the enzyme lpg0696 whichwas shown to reverse the phosphocholination reaction by hydro-lytically removing the phosphocholine group from Rab1b (18),thereby reconstituting the Rab1b-f-NBD:GDP:GDI complex.The addition of the tightly binding LidA was again able to dis-place GDI.
Adenylylation or GTP-loading of Rab1b-f-NBD by DrrA can-not accelerate GDI displacement beyond the intrinsic rate ofGDI dissociation. Thus, we investigated whether phosphocholi-nation could promote GDI displacement faster than the sponta-neous rate of GDI dissociation. To this end we compared the GDIdisplacement by LidA binding (Fig. 5B) with the displacementcaused by the enzymatic activity of AnkX (Fig. 5C). The displa-cement by LidA (koff ¼ 0.0277 s−1) was twice as fast as the dis-sociation caused by Rab1b-f-NBD phosphocholination withAnkX (kobs ¼ 0.0150 s−1) under the conditions used.
Because Rab35 is also a substrate for AnkX (13), we per-formed similar experiments with Rab35-f-NBD (Fig. 6 A–C).GDI displacement from Rab35-f-NBD:GDP by phosphocho-lination (kobs ¼ 0.0022 s−1) was as fast as with LidA (koff ¼0.0020 s−1). Therefore, GDI displacement of Rab35-f-NBD:GDI by phosphocholination is limited by the intrinsic rate ofGDI dissociation.
Hence, in contrast to adenylylation by DrrA, GDI displace-ment from inactive Rab1b-f-NBD:GDP by AnkX was possible
on a physiologically meaningful time scale, indicating that cova-lent modification of Rab proteins is a way to effectively dissoci-ate GDI.
DiscussionThe binding of Rab proteins to the universal regulatory proteinGDI is of crucial importance for Rab membrane cycling and thusfor the regulation of vesicular trafficking (19). In order to exerttheir function, Rab proteins need to be recruited specifically tothe cytosolic surface of intracellular membranes in a temporallycontrolled manner. However, the affinity of the interaction be-tween prenylated Rabs and GDI is high and consequently thespontaneous complex dissociation is very slow. Therefore, me-chanisms for regulated recruitment of Rabs and hence effectivedisplacement of GDI from cytosolic Rab:GDP:GDI complexesmust exist. In earlier works, it was shown that human Pra1 andits yeast homolog Yip3 act as GDI displacement factors and arecapable of recruiting prenylated Rab9:GDP to membranes priorto activation by nucleotide exchange (Fig. 7A). However, themechanism of the GDI displacement reaction remains elusive.Recently, it was shown that nucleotide exchange itself makesan important energetic contribution to the effective displacementof GDI (3, 9), and thus that GEFs could potentially recruit Rabproteins to membranes without the need for an additional GDIdisplacement factor (Fig. 7B). Nevertheless, this mechanism ofGDI displacement is incapable of accelerating the slow GDI dis-sociation, which means that the rate at which Rab proteins couldbe recruited to membranes would be limited by the spontaneousrate of dissociation of GDI:Rab:GDP complexes in the absenceof additional factors.
In this work, we investigated the influence of posttranslationalmodifications of Rab proteins on the ability to displace GDI fromprenylated Rabs. It has been observed that the enzymes DrrA andAnkX from Legionella can adenylylate and phosphocholinateRab1b and Rab35 at Tyr77 and Ser76, respectively. As demon-strated here, these modifications abolish GDI binding and hencelead to effective displacement of GDI from the Rab protein. GDIdisplacement by adenylylation, however, was dependent on theactivation of Rab1 by nucleotide exchange because the catalyticactivity of DrrA for Rab1b:GDP is low (14). In contrast, the cat-alytic activity of AnkX is high for both GDP- and GTP-boundRab1 (and Rab35) and thus the enzymatic activity of AnkX issufficient to displace GDI. As found for GDI displacement by
1,000500
B C
643
0.0
0.2
0.4
0.6
0.8
1.0
1.2
4003000time / s200100
koff = 0.0277 s-1
1 2
3,5003,000time / s
2,500
2
5
1
norm
aliz
ed fl
uore
scen
ce
0.0
0.2
0.4
0.6
0.8
1.0
1.2
4,0000 2,0001,500
A
norm
aliz
ed fl
uore
scen
ce
norm
aliz
ed fl
uore
scen
ce
0.0
0.2
0.4
0.6
0.8
1.0
1.2
8006000 200
kobs = 0.0150 s-1
400time / s
1: 50 nM Rab1b-f-NBD2: + 75 nM GDI-13: + 1 mM CDP-Choline4: + 250 nM AnkX 5: + 500 nM lpg06966: + 1 µM LidA
1: 50 nM Rab1b-f-NBD, 75 nM GDI-1 1 mM CDP-Choline2: + 2 µM AnkX
1: 50 nM Rab1b-f-NBD, 75 nM GDI-12: + 1 µM LidA
21
Fig. 5. GDI displacement by phosphocholination of Rab1b. (A) GDI can beeffectively displaced from Rab1b-f-NBD by phosphocholination with AnkX.Reversal of phosphocholination by lpg0696 leads to an increase in fluores-cence that corresponds to rebinding of GDI as addition of LidA needs to arebinding of GDI. (B) Single exponential fit of GDI displacement by LidAshowing the intrinsic dissociation rate of the Rab1b-f-NBD:GDI complex.(C) GDI displacement by phosphocholination with AnkX. The observed rateconstant is lower than the intrinsic dissociation rate of the complex.
norm
aliz
ed fl
uore
scen
ce
0 1,500 2,0000.4
0.6
0.7
0.8
0.9
1.0
1.1 1 2 1: 25 nM Rab35-f-NBD, 37.5 nM GDI-12: + 1 µM LidA
time / s1,000500
koff = 0.0020 s-1
0.5
B
norm
aliz
ed fl
uore
scen
ce
0.0
0.2
0.4
0.6
0.8
1.0
1.2
4,5000 3,0002,500
A 1 2 1: 50 nM Rab35-f-NBD2: + 75 nM GDI-13: + 1 mM CDP-Choline4: + 250 nM AnkX
4,000time / s
3,500
3 4
2,0001,000
C
norm
aliz
ed fl
uore
scen
ce
0 1,500 2,000time / s
1,0005000.4
0.6
0.7
0.8
0.9
1.0
1.1
0.5
1 2
kobs = 0.0022 s-1
1: 50 nM Rab35-f-NBD, 75 nM GDI-1 1 mM CDP-Choline2: + 1 µM AnkX
1,500500
Fig. 6. GDI displacement by phosphocholination of Rab35. (A) GDI can beeffectively displaced from Rab35-f-NBD by phosphocholination with AnkX.(B) Single exponential fit of GDI displacement by LidA showing the intrinsicdissociation rate of the Rab35-f-NBD:GDI complex. (C) GDI displacement byphosphocholination with AnkX. The observed rate constant is lower than theintrinsic dissociation rate of the complex.
5624 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1121161109 Oesterlin et al.

nucleotide exchange (3, 9), the rate of complex disruption causedby phosphocholination is limited by the intrinsic dissociationrate of Rab:GDI complexes. Hence, neither phosphocholinationnor nucleotide exchange actively displace GDI from Rab becausethey do not accelerate the rates of dissociation of the complexes.Nevertheless, disruption of the Rab:GDP:GDI complex forma-tion by covalent modifications might solve the thermodynamicproblem of slow complex dissociation even if they cannot accel-erate the process. For example, phosphocholination of Rab1 byAnkX will disrupt GDI binding and thus lead to incorporation ofRab1 into membranes where AnkX resides (Fig. 7C). In thisstate, Rab1 is membrane bound in the inactive state but is resis-tant to activation by GEFs (13). This scenario would lead to astable membrane pool of GDP-bound Rab, with serious conse-quences for transport mechanisms. The action of a demodifyingenzyme, lpg0696 in this case, renders Rab1 accessible to GEFsagain so that vesicular trafficking could proceed. The rate of gen-eration of active Rabs would then become dependent on the de-modification reaction instead of the dissociation of Rab:GDIcomplexes. Therefore, posttranslational modifications of Rabproteins could potentially generate a stable pool of GDP-boundRabs that can be quickly utilized for subsequent steps. Posttran-slational modifications might represent a general mechanism ofcontrolling GDI binding and the cytosolic or membrane distribu-tion of Rab proteins. In agreement with this argument, it has beenreported that several phosphorylation reactions can modify themembrane-cytoplasm partitioning of Rab proteins. Rab1a andRab4 are modified during mitosis by Cdk1 (20). The cytoplasmicconcentration of Rab4 is increased upon phosphorylation (20, 21)whereas phosphorylation of Rab1 leads to a decrease in cytoplas-mic protein (20). Phosphorylation of Rab6 by Protein Kinase Calso decreases the cytoplasmic protein concentration (22).Furthermore, it could be shown that Rab4 and Rab5 associatedGDI molecules are phosphorylated in vivo (23), but it remainsunclear which residues are phosphorylated. Serine phosphoryla-tion of GDI-1 by serum- and glucocorticoid-inducible kinase 1(24) as well as tyrosine phosphorylation on GDI-2 increases theamount of cytoplasmic Rab4:GDI complex (25). These differ-ences in membrane-cytoplasm partitioning could be caused bya modification in Rab:GDI affinity similar to the mechanismsof phosphocholination and adenylylation described here. The
existence of eukaryotic enzymes that modify Rab proteins in or-der to suppress GDI binding and to localize Rabs to intracellularmembranes prior to activation by GEFs is conceivable but nonehave been reported thus far.
We have shown that adenylylation and phosphocholination ofRab proteins are able to modulate the affinity for the regulatorprotein GDI. The enzyme AnkX can cause effective GDI displa-cement from Rab1 and Rab35 by phosphocholinating these Rabs.GDI displacement is limited by the rate of spontaneous Rab GDIcomplex dissociation only, therefore it is similar to the GDI dis-placement by GEFs in vitro (3, 9). Consequently, posttransla-tional modifications of Rab proteins could represent a way torecruit Rabs to a particular membrane prior to their activationby nucleotide exchange.
Materials and MethodsProtein Isolation. Sec4, Rab5a, Rab35, and codon-optimized Rab1b (Gene,Regensburg, Germany) were C-terminally modified to contain the GGTaseI recognition amino acid sequence CVIL using the QuikChange mutagenesiskit (Stratagene). Protein expression was performed in BL21(DE3) (Rab1b,Rab5a, DrrA) or BL21-CodonPlus(DE3)-RIL cells (Rab35, Sec4, AnkX, lpg0696,GGTase I) at 20 °C over night or at 37 °C for 5 h (Rab5a). Protein expressionwas induced with 0.2 mM, 0.3 mM (Sec4, LidA) or 0.5 mM IPTG (GGTase I).Rab1b, Rab5a, Rab35 were expressed as cleavable His6-MBP-fusion con-structs, Sec4 and Rabin8 were expressed as cleavable His6-fusion constructs.GGTase I α and β were coexpressed. GGTase I α was expressed as a His6-GSTfusion construct and GGTase I β as a His6-fusion construct. Bovine GDI-Iwas expressed in Sf21 insect cells using the Bac-to-Bac Expression System(Invitrogen) with a cleavable His-tag.
Protein purification was performed analogous to the isolation proceduredescribed for Rab8 (but without ATP) (26). Isolation of Rabin8, Rab5a, andRab35 was performed at pH 7.5 and Rabin8, DrrA, AnkX, and lpg0696 wereisolated without GDP and MgCl2. To stabilize the proteins for Rab5a, AnkX,and lpg0696 the NaCl concentration was increased to 200 mM. AdditionallyRab5a was isolated in the presence of 5% glycerol. Residual MBP-tag con-taminants were removed via an amylose resin. GGTase I was isolated as a fu-sion protein; therefore the protease cleavage and the second Ni-IDAchromatography steps were omitted. The first Ni-IDA chromatography withGDI-I was performed with 50 mM NaPi pH 8, 300 mM NaCl and 2 mM β-Mer-captoethanol and 5 mM β-Mercaptoethanol was used instead of DTE(dithioerythritol) during the other purification steps. His6-Sec21–165 and LidAwere expressed and isolated as described (17, 27). Rabex5132–397 was clonedby the Dortmund Protein Facility into a pOPINF-vector (N-terminal His6-tagfollowed by a PreScission protease cleavage sequence) by the in-fusion
Fig. 7. Conceivable GDI displacement mechanisms. Forced or spontaneous GDI-dissociation are prerequisites for membrane association of Rab proteins. Inthe classical mechanism (A), PraI recruits Rab proteins to the membrane; but it is unclear whether PraI interacts with GDI-free or GDI-bound Rab. Anotherrecruitment mechanism is based on RabGEFs that displace GDI by exploiting its low affinity for active, GTP-bound Rabs (B). Whether this process operates oncytosolic or membrane-bound Rabs is unclear. Here it has been found that the displacement of GDI from Rab proteins by posttranslational modifications isanother possibility for Rab membrane recruiting (C). Similar to GEFs, phosphocholination by AnkX could happen on cytosolic or membrane-bound Rabs. By thismechanism, a membrane-bound pool of inactive Rabs can be generated that can be utilized by another enzyme (here lpg0696) for downstream processes suchas activation by GEFs. In this manner, a slow Rab GDI-dissociation step can be replaced by a faster enzymatic process; i.e., dephosphocholination, when fastmobilization and activation of a Rab protein is needed.
Oesterlin et al. PNAS ∣ April 10, 2012 ∣ vol. 109 ∣ no. 15 ∣ 5625
BIOCH
EMISTR
YSE
ECO
MMEN
TARY

cloning method (28). Rabex5132–397 was expressed in Escherichia coli BL21(DE3) RIL 4 h at 37 °C after induction with 1 mM IPTG and purified by a com-bination of Ni-IDA and size exclusion chromatography in a final buffer con-sisting of 25 mM Hepes, pH 7.5, 40 mM NaCl and 1 mM TCEP.
Prenylation. NBD-farnesyl pyrophosphate was produced as described else-where (29). Prenyltransferase, Rab protein and substrate were mixed in a0.5∶1 : 5 ratio and incubated at room temperature for 1.5 h. GGTase I andfarnesyl-NBD were preincubated for 30 min before addition of Rab protein.Prenylated Rab1b was separated from GGTase I by several rounds of GSH andNi-IDA affinity chromatography in the presence of 5% CHAPS with a finalbuffer containing 20 mM Hepes pH 7.2, 50 mM NaCl, 2 mM β-Mercaptoetha-nol, 2 mM MgCl2 and 10 μM GDP. Rab5a, Rab35, and Sec4 were separatedfrom GGTase I with Gel filtration chromatography using a Superdex 20010∕30 column with 20 mM Hepes pH 7.2, 500 mM NaCl, 2 mM MgCl2,2 mM DTE and 10 μM GDP. The NaCl concentration in the reaction mixturewas increased to 500 mM and 5% CHAPS (wt∕vol) was added to the reactionbefore loading.
Kinetic Measurements. Fluorescence measurements were performed with aFluoroMax-3 (Horiba Jobin Yvon Inc.) or a stopped-flow apparatus (AppliedPhotophysics). NBD fluorescence was excited at 479 nm and emission was de-tected at 525 nm. For stopped-flow measurements fluorescence was excitedat 437 nm and emission was detected through a 530 nm cut off filter. Mea-surements were performed in 25 mM Hepes pH 8, 50 mM NaCl, 5 mM MgCl2and 5 mM DTE at 25 °C.
Preparative Adenylylation. Rab1b, DrrAfl, and ATP were mixed in a 10∶1∶25ratio and incubated for 1.5 h at room temperature. DrrAfl was removed bygel filtration chromatography using a Superdex 75 column equilibrated in20 mM Hepes pH 7.5, 50 mM NaCl, 1 mM MgCl2, 2 mM DTE and 10 μM GDP.
ACKNOWLEDGMENTS. Nathalie Bleimling is acknowledged for invaluabletechnical assistance. L.K.O. was supported by the International Max PlanckResearch School in Chemical Biology (Dortmund). This work was supportedby a grant from the Deutsche Froschungsgemeinschaft SFB642, project A4.
1. Shahinian S, Silvius JR (1995) Doubly-lipid-modified protein-sequence motifs exhibitlong-lived anchorage to lipid bilayer-membranes. Biochemistry 34:3813–3822.
2. Ignatev A, Kravchenko S, Rak A, Goody RS, Pylypenko O (2008) A structural model ofthe GDP dissociation inhibitor rab membrane extraction mechanism. J Biol Chem283:18377–18384.
3. Wu YW, et al. (2010) Membrane targeting mechanism of Rab GTPases elucidated bysemisynthetic protein probes. Nat Chem Biol 6:534–540.
4. Ullrich O, Horiuchi H, Bucci C, Zerial M (1994) Membrane association of Rab5 mediatedby GDP-dissociation inhibitor and accompanied by GDP/GTP exchange. Nature368:157–160.
5. Soldati T, Shapiro AD, Svejstrup AB, Pfeffer SR (1994) Membrane targeting of the smallGTPase Rab9 is accompanied by nucleotide exchange. Nature 369:76–78.
6. Novick P, Garrett MD (1994) Vesicular transport—no exchangewithout receipt.Nature369:18–19.
7. Hutt DM, Da-Silva LF, Chang LH, Prosser DC, Ngsee JK (2000) PRA1 inhibits the extrac-tion of membrane-bound rab GTPase by GDI1. J Biol Chem 275:18511–18519.
8. Sivars U, Aivazian D, Pfeffer SR (2003) Yip3 catalyses the dissociation of endosomalRab-GDI complexes. Nature 425:856–859.
9. Schoebel S, Oesterlin LK, Blankenfeldt W, Goody RS, Itzen A (2009) RabGDI displace-ment by DrrA from Legionella is a consequence of its guanine nucleotide exchangeactivity. Mol Cell 36:1060–1072.
10. Machner MP, Isberg RR (2007) A bifunctional bacterial protein links GDI displacementto Rab1 activation. Science 318:974–977.
11. IngmundsonA, Delprato A, Lambright DG, Roy CR (2007) Legionella pneumophila pro-teins that regulate Rab1 membrane cycling. Nature 450:365–369.
12. Wu YW, Tan KT, Waldmann H, Goody RS, Alexandrov K (2007) Interaction analysis ofprenylated Rab GTPasewith Rab escort protein andGDP dissociation inhibitor explainsthe need for both regulators. Proc Natl Acad Sci USA 104:12294–12299.
13. Mukherjee S, et al. (2011) Modulation of Rab GTPase function by a protein phospho-choline transferase. Nature 477:103–106.
14. MüllerMP, et al. (2010) The Legionella effector protein DrrA AMPylates themembranetraffic regulator Rab1b. Science 329:946–949.
15. Schoebel S, Cichy AL, Goody RS, Itzen A (2011) Protein LidA from Legionella is a RabGTPase supereffector. Proc Natl Acad Sci USA 108:17945–17950.
16. Delprato A, Lambright DG (2007) Structural basis for Rab GTPase activation by Vps9domain exchange factors. Nat Struct Mol Biol 14:406–412.
17. Itzen A, Rak A, Goody RS (2007) Sec2 is a highly efficient exchange factor for the Rabprotein Sec4. J Mol Biol 365:1359–1367.
18. Tan Y, Arnold RJ, Luo ZQ (2011) Legionella pneumophila regulates the small GTPaseRab1 activity by reversible phosphorylcholination. Proc Natl Acad Sci USA108:21212–21217.
19. Garrett MD, Zahner JE, Cheney CM, Novick PJ (1994) GDI1 encodes a GDP dissociationinhibitor that plays an essential role in the yeast secretory pathway. EMBO J13:1718–1728.
20. Bailly E, et al. (1991) Phosphorylation of two small GTP-binding proteins of the Rabfamily by p34cdc2. Nature 350:715–718.
21. van der Sluijs P, et al. (1992) Reversible phosphorylation dephosphorylation deter-mines the localization of Rab4 during the cell-cycle. EMBO J 11:4379–4389.
22. Fitzgerald ML, Reed GL (1999) Rab6 is phosphorylated in thrombin-activated plateletsby a protein kinase C-dependent mechanism: effects on GTP/GDP binding and cellulardistribution. Biochem J 342:353–360.
23. Steele-Mortimer O, Gruenberg J, Clague MJ (1993) Phosphorylation of GDI and mem-brane cycling of Rab proteins. Febs Letts 329:313–318.
24. Liu WH, Yuen EY, Yan Z (2010) The stress hormone corticosterone increases synapticalpha-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic Acid (AMPA) receptors viaSerum- and Glucocorticoid-inducible Kinase (SGK) regulation of the GDI-Rab4 com-plex. J Biol Chem 285:6101–6108.
25. Shisheva A, Chinni SR, DeMarco C (1999) General role of GDP dissociation inhibitor 2 inmembrane release of Rab proteins: modulations of its functional interactions by invitro and in vivo structural modifications. Biochemistry 38:11711–11721.
26. Bleimling N, Alexandrov K, Goody R, Itzen A (2009) Chaperone-assisted production ofactive human Rab8A GTPase in Escherichia coli. Protein Expres Purif 65:190–195.
27. Machner MP, Isberg RR (2006) Targeting of host Rab GTPase function by the intrava-cuolar pathogen Legionella pneumophila. Dev Cell 11:47–56.
28. Berrow NS, et al. (2007) A versatile ligation-independent cloning method suitable forhigh-throughput expression screening applications. Nucleic Acids Res 35(e45).
29. Dursina B, et al. (2006) Identification and specificity profiling of protein prenyltrans-ferase inhibitors using new fluorescent phosphoisoprenoids. J Am Chem Soc128:2822–2835.
30. de Castro E, et al. (2006) ScanProsite: detection of PROSITE signature matches andProRule-associated functional and structural residues in proteins. Nucleic Acids Res34:W362–W365.
31. Rak A, et al. (2003) Structure of Rab GDP-dissociation inhibitor in complex withprenylated Ypt1 GTPase. Science 302:646–650.
5626 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1121161109 Oesterlin et al.





























