Embed Size (px)
Citation preview

Developmental Brain Research, 47 (1989) 197-207 197 Elsevier
BRD 50914
Postnatal development of the efferent innervation of the rat cochlea
Donald Robertson 1, Alan R. Harvey 2 and K. Stewart Cole 1
1Department of Physiology and 2Department of Anatomy and Human Biology, The University of Western Australia, Nedlands, Western Australia (Australia)
(Accepted 10 January 1989)
Key words: Brainstem; Efferent; Rat; Developmental plasticity; Auditory pathway; Cell death
The postnatal development of the.efferent innervation of the rat cochlea was studied by intracochlear injection of the fluorescent retrograde neuronal tracers Diamidino yellow and Fast blue. Injections were performed on adult rats and on neonatal rats ranging from 0 to 8 postnatal days. It was found that the total number of neurones labelled in the brainstem after intracochlear injection was not significantly different in the newborn rat, compared to the adult. On the basis of cell body location and laterality of projections, there was a clear separation into lateral and medial efferent systems at the earliest postnatal age studied (P0). Evidence was also found in the newborn for a tonotopicity in the lateral system projection similar to that in the adult. Differences between the newborn and adult were a slight but significantly greater number of bilaterally-projecting cells in the newborn, and the presence in the newborn of a small number of cells located in the lateral superior olivary nucleus contralateral to their target cochlea. These were extremely rare in the adult brainstem. Evidence was found for the occurrence of postnatal neuronal death in nuclei of origin of both efferent systems. It is suggested that although the overall extent and general organization of the efferent projection to the cochlea in the rat appears to be established at birth, regressive changes are occurring during the postnatal shaping and maturation of this brainstem-to-cochlea pathway.
INTRODUCTION
The efferent innervat ion of the mammal ian co-
chlea would seem to provide a rich field for the
invest igat ion of the factors that de te rmine the
deve lopmen t of neural project ions. Two distinct
systems pro jec t ing from the bra ins tem to the cochlea
have been descr ibed in a number of species 2" 3.18,19.24,25. These efferent systems differ in their site
of origin, neuronal morphology and cellular targets
within the cochlea. In the adult rat , the small
unmyel ina ted axons of the lateral ol ivocochlear
system arise in the lateral super ior olivary nucleus
(LSO) , ascend towards the midline and turn sharply
near the facial genua to innervate exclusively the
ipsi lateral cochlea. This system has been shown in
the cat to te rmina te pr imari ly on the distal dendri tes
of the eighth nerve afferent neurones. The larger
neurones of the medial o l ivocochlear system give
rise to myel ina ted axons which ascend to the floor of
the IVth ventricle and then turn to innervate e i ther
the ipsi lateral or contra la tera l cochlea. Thei r targets
in the cochlea are p robab ly the ou te r hair cells.
Based on e lec t ronmicroscopic examinat ion , it has
been repor ted that in the rat, the first efferent
synapses on the outer hair cells appea r at about
postnata l day 6 (P6) and gradual ly displace a
t ransient afferent innervat ion of these recep tor
cells 12. This evidence, which accords with results in
o ther mammals 16, is suggestive of a dynamic shaping
of the media l system efferent innervat ion in the first
10 days of pos tnata l life in this species. This not ion
is re inforced by a recent immunohis tochemica l s tudy
which shows that nerve growth factor ( N G F ) is
t ransient ly expressed by the ou te r hair cells be tween
birth and P8 (ref. 11).
Correspondence: D. Robertson, Department of Physiology, University of Western Australia, Nedlands, Western Australia 6009,' Australia.
0165-3806/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

198
In this study, we were concerned to answer the
following questions. Firstly, at what age is it possi- ble, after intracochlear injection of tracers, to retrogradely label rat olivocochlear neurones of the lateral and medial systems, and how do the relative numbers of cells compare between the neonate and the adult? Secondly, is the period when the efferent innervation of the outer hair cells is reportedly being established associated with significant cell death
within the brainstem? This might be expected from the known mechanisms of shaping of innervation in other neuronal systems 7"1°'j4. Thirdly, is there evi-
dence for an exuberant bilateral projection from brainstem to cochlea early in development? We have suggested previously ~9 that bilaterally projecting cells, found in small numbers in the adult guinea
pig 1~, might represent the remnants of an extensive bilateral projection early in development which is later retracted or eliminated to form the adult
pattern.
MATERIALS AND METHODS
Surgery Experiments were performed on pigmented
(PVG/c) rats free of middle ear infection. All intracochlear injections were performed under gen- eral anaesthesia, the wounds were closed with surgical silk and the animals were allowed to recover for periods ranging from 24 h to 5 days before being sacrificed.
Adult rats weighing 180-270 g were anaesthetized with Hypnorm (Janssen; fentanyl citrate, 0.315 mg/ml and fluanisone, 10 mg/ml) and Hypnovel (Roche; midazolam, 5 mg/ml). The initial doses were 0.2 ml intraperitoneal and intramuscular re- spectively, supplemented ad libitum, and incision points were infiltrated with Lignocaine. A small hole was drilled in the cochlear apex and in the basal turn scala tympani. Fluorescent dyes were pressure- injected via a glass pipette through the hole in the basal turn until dye was seen emerging from the apex. In all animals, one cochlea was injected with a 2% aqueous suspension of Diamidino yellow (DY) and in a number of animals the other cochlea was injected with a 4% aqueous solution of Fast blue (FB). Quantitative data are reported from 5 adult animals in which fully successful bilateral injections
were carried out. In a number of other animals, partially successful injections were performed, and only qualitative results from these are included in the present report.
Similar intracochlear injections were performed on litters of neonatal rats ranging from 0 to 8 days postnatal age (P0-P8; day of birth = P0). They were anaesthetized by intraperitoneal injection with 2, 2,2-tribromoethanol. In these animals, the intra- cochlear injections were performed by insertion of the pipette through the round window, in successful injections the dyes were visible through the cartilag-
inous otic capsule as they coursed through the scala tympani. The apex of the cochlea was not opened. The rat pups were left with their mothers until being sacrificed 24 h postinjection. A small number of animals was allowed to survive for 48 h postinjection and no significant difference was found in total cell numbers compared to the 24-h groups. Successful bilateral injections were performed on 4 P0 animals, and 4 P1 animals, and a number of unilateral DY
injections were successfully carried out at these and other postnatal ages.
Cell counting After the survival periods, all animals were fixed
by intracardiac perfusion of 8% paraformaidehyde in 0.1 M phosphate buffer, pH 7.4. The brains were removed and soaked for 24 h in 30% sucrose in 0.1 M phosphate buffer, pH 7.4 (with the inclusion of 4% paraformaldehyde for neonatal brains). Serial 40-~m frozen sections were cut in the coronal plane and mounted on subbed slides. All sections were examined by fluorescence microscopy (peak excita- tion wavelength 405 nm, barrier filter cut-off 455 nm) and all labelled cells were counted and their locations noted.
In the case of FB-labelled cells only perikarya were counted. For DY-labelled cells, in which only the nucleus was brightly labelled, cells were only scored as labelled if a clearly ovoid profile was seen. Thin slivers through nuclear caps and clearly patho- logical structures were not scored. Double-labelled cells were identified by their FB-labelled cytoplasm and DY-stained nuclei, as reported previously ~8"19.
The cell counts presented are the total numbers of labelled cells scored in all serial sections. We have not applied any correction to these counts for the

199
following reasons. We have estimated the size of cell
nuclei of DY-labelled cells and applied a modifica-
tion of Abercrombie ' s correction for double counting 1. This led to a maximum correction of only
12%, which is considerably less than the variance in
the data. This is in any case an overestimate of the
double-counting error, since it ignores the fact that we did not count nuclear caps or small fragments of
cells. Furthermore, the numbers of FB- and DY-
labelled cells in any given double-injected animal
were in good agreement, although different features
of the cells are stained by the two dyes. Finally, in two animals, we cut alternate 40-/~m and 60-pm
sections and separately estimated the total cell
counts from these two sets of sections. The estimated
total numbers agreed within 10% of each other. We
Fig. 1. Photomicrographs of fluorescent-labelled cells in the left brainstem of rats after injection of retrograde tracer into left cochlea. A: adult LSO ipsilateral to DY-injected cochlea; B: adult rostral periolivary region; C,D: animal injected at P0 and sacrificed at P1 showing ipsilateral LSO (C) and rostral periolivary region (D). Arrow in C indicates labelled medial system cells ventral to LSO. Scale bars for adult and neonatal brains = 100/zm. Arrows at bottom right of D indicate dorsal and lateral coordinates.

20O
therefore feel confident that although a number of
corrections could be applied, some to compensate for over-counting and some for under-counting, their application in the present case is not strongly indicated.
After counting of labelled cells, every third section was stained with Neutral red and camera lucida drawings were made of the brainstem. Nuclei were classified according to the general scheme of Paxinos and Watson is, the one modification being
that we have defined the ventral nucleus of trapezoid
body (VNTB). In a further series of 5 animals (P1, P4, P8, P16 and adult) in which intracochlear injections were not carried out, serial 40-/zm frozen
sections were stained with Cresyi fast violet. The injected cochleas were examined with a
dissecting microscope and the location of the dye in
the scala tympani was verified. In a number of animals, dye was found to be located in the scala media and scala vestibuli. In these animals, the brainstem, particularly the cochlear nuclei and trap- ezoid body, was contaminated by a large amount of diffuse, non-specific staining of neurones and neu- roglia. We have no explanation for this except to suggest that leakage into cerebrospinal fluid may occur from scala media. Results from these animals were not included in the analysis.
RESULTS
Adult distribution
The appearance of labelled efferent neurones and their typical distribution within the brainstem in adult animals is shown in Figs. 1A,B and 2A. The
A
ADULT
LSO 27
t I m r n

201
pattern of labelling is in qualitative agreement with that obtained in other adult rat strains 2'3"25.
Small neurones of the lateral system were located
within the LSO ipsilateral to the injected cochlea. In
animals in which the injections did not completely
fill the scala tympani in all cochlear turns, the
intensity of labelling was greatest in the medioven-
tral lobe of the LSO, most probably corresponding
to the mid-to-high frequency parts of the ascending
pathway. The number of labelled neurones found in
the contralateral LSO was extremely small (mean
number = 0.2, S.D. = 0.42, in 5 animals).
Medial system neurones were found on both sides
of the brainstem in the lateral and ventral nuclei of
the trapezoid body and in the rostral peri-olivary
zone. We did not find the 2:1 preponderance of
contralateral medial cells reported by some authors
using horseradish peroxidase 25. The cause of this
discrepancy is unclear since in a previous study in the guinea pig, fluorescent tracers were shown to yield
greater numbers of labelled neurones than HRP, but
without any significant change in the relative num-
bers of ipsilaterally and contralaterally located me-
dial neurones ~9. In our material, the mean number
of ipsilateral medial neurones was 220.6 (S.D. =
43.89) and the mean number of contralateral medial
neurones was 253 (S.D. = 38.04). Quantitatively,
these results are in good agreement with recent
reports in the rat also using FB and DY 3. In the adult
rat, the number of double-labelled cells was small
o , , o
n,... MSO ~ ( ( J ( / ~ 30
i i 1 r a m
Fig. 2. Camera lucida drawings showing the typical distribution of DY- and FB-labelled cells after bilateral intracochlear injections in adult (A) and P0 rats (B) in which the left cochlea was injected with DY and the right with FB. Open circles, FB-labeUed cells; asterisks, D Y-labelled cells; solid circles, double-labelled cells. MSO, medial superior olive; LSO, lateral superior olive; Mo5, motor nucleus of nerve 5; N5, sensory nucleus of nerve 5; CN, cochlear nucleus; $5, sensory root of nerve 5; 7, facial nerve root; Py, pyramidal tract; MTz, medial nucleus of trapezoid body; VLL, ventral nucleus of trapezoid body; SPO, superior olivary region; lfp, longitudinal fasciculus pons; PN, pontine nuclei; VNTB, ventral nucleus of trapezoid body. Classification according to Paxinos and Watson 15. Numbers beside each drawing indicate section number increasing rostrally.

202
when care was taken to exclude those animals with
FB contaminat ion of the cochlear nucleus. The mean
number of double-labelled cells in 5 successful
bilaterally-injected animals was 8.8 (S.D. = 4.65),
which represented only 0.8% of the total efferent
projection to one cochlea (mean total number =
1055, S.D. = 131.7). These double-labelled cells
were found entirely in zones of origin of the medial
system. Table I summarizes the results for all adult
animals in which successful injections were carried
out. To be included in the analysis there had to be
an absence of brainstem contaminat ion, and brightly
labelled cells had to be present throughout all lobes
of the ipsilateral LSO, the latter being indicative of
adequate perfusion of the entire length of the cochlea 17.
Neonata l distribution
The typical distribution of labelled cells in a
neonatal rat injected at P0 and sacrificed 24 h later
is shown in Figs. 1C,D and 2B. Both lateral and
medial olivocochlear systems could be identified.
There was a clear differentiation of the LSO at this
stage of development , although it was considerably
smaller than in the adult. Large numbers of small
labelled cells were seen located within this nucleus in
a manner strikingly similar to the picture in the adult
brainstem. As in the adult brainstem, labelling was
often found to be brightest for cells located in the
medial aspect of the LSO, particularly in animals in
which the dye injection did not fill all turns of the
cochlea (Fig. 3). In all except one P0 animal, the
FB-labelled cells in the LSO were very closely
packed together and it was not possible to count
their number with confidence; due to the lack of
TABLE 1
Distribution of labelled efferents in adult brainstem
Mean total number/cochlea = 1055, S.D. = 131.7.
Ipsi- Contra- lpsi- Contra- Double LSO LSO medial medial label
Number of cells 572.7 0.2 220.6 253.0 8.8
+ S.D. 52.2 0.4 43.9 38.1 4.7
% Total 54.3 0.0 20.9 24.0 0.8
A D U L T
O 0 0 ©
~ e v v - I f I
Media l O. 1 mm q
• °O,,o o- o o
t I
0 .1 mm
Fig. 3. Camera lucida drawings of typical distribution of labelled cells in LSO of adult and neonate after intracochlear injections restricted to basal and second cochlear turns. In both cases, the cochlea ipsilateral to the LSO shown was injected with DY and the contralateral cochlea was injected with FB. Medial is to the left in both cases. DY-labeUing is confined to medial and ventral regions of the nucleus. Solid circles, brightly-stained cells; solid triangles, cells with clear but not strong labelling; open circles, just detectable labelling; solid squares, FB-labelled cells.
1 5 0 0 - - w
m
1 0 0 0 -
W 0
0 500-- $
0 0
n:5 n:5 n:5
T T I
PO P1 ADULT
n:3
L n:4
P4 P 7 - 8
Fig. 4. Histogram comparing mean total numbers of labelled ceils projecting to one cochlea at various postnatal ages. Error bars indicate one standard deviation. There were no statisti- cally significant differences between adult brains and any of the neonatal age groups studied; n indicates numbers of animals in each group.
203
cytoplasmic label it was easier to count the number
of DY-labelled cells and the less densely packed FB-labelled cells in other brainstem regions could also be readily quantified. Larger cells were found in regions ventral, lateral and rostral to the LSO in zones clearly corresponding to the medial system zones in the adult (Figs. 1 and 2). This overall
adult-like pattern of distribution of labelled neu- rones was found at all ages investigated, which included P0, P1, P4 and P7/P8.
Differences between adult and neonatal organization The total numbers of labelled cells projecting to
one cochlea are compared for P0, P1, P4, P7/8 and adult rats in the histogram of Fig. 4. These data include both bilaterally and unilaterally injected animals. There were no significant differences be- tween any of the groups (P > 0.05 or more in all cases) and thus there is no evidence for any peak in total cell numbers at any of the postnatal ages studied. Fig. 5 shows a comparison between P0 and adult animals for the single-labelled cells in ipsilat- eral LSO, and ipsilateral and contralateral medial zones. Again there were no significant differences between P0 and adult animals for these major cell groups.
However, significant differences were found be- tween the adult and neonatal rats in the number of double-labelled cells and the number of single- labelled cells located within the contralateral LSO.
Q t t l
i I l u
_1
. I u l
0
ipsl LSO Ipsl reed. contra m~l.
Fig. 5. Histogram comparing mean numbers of ipsilateral LSO and ipsilateral and contralaterai medial system ceils projecting to one cochlea in P0 and adult animals. Open bars, P0 animals; solid bars, adult animals. Error bars indicate one standard deviation. There were no significant differences between P0 and adult groups in any of the regions of origin.
50 - -
-I, 4 0 - - W 0 a W ..J
30-
< .J
I W ~ 2 0 -
0 a
-J 1 0 - -
I,-- 0 I I
Z
DOUBLE-LABELLED CELLS
T
PO P1
Iii ADULT
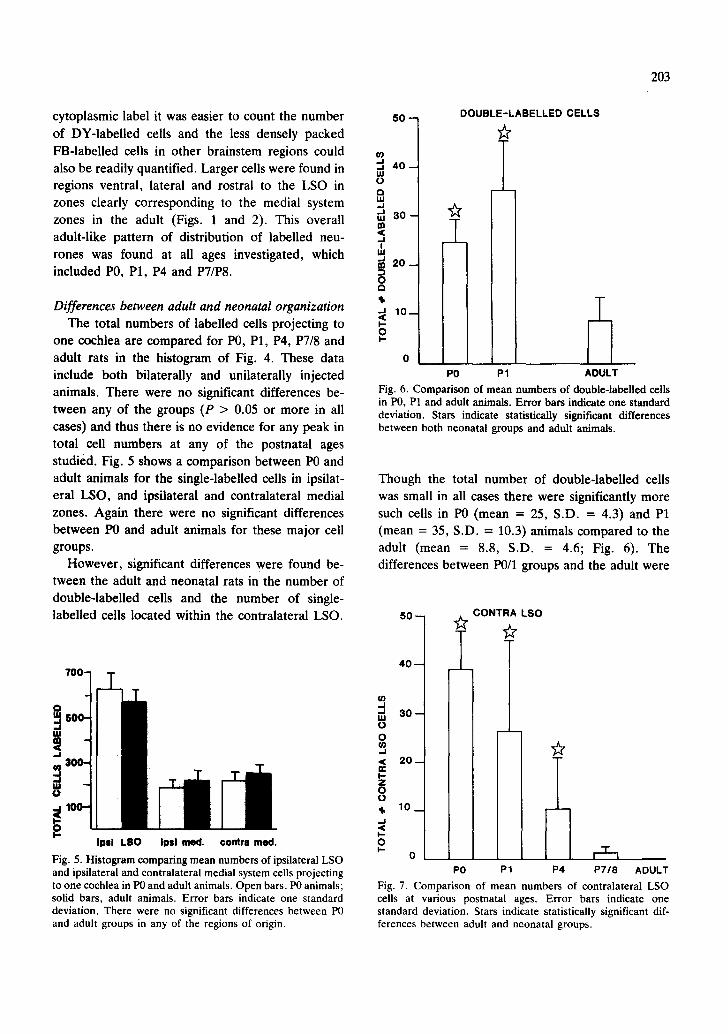
Fig. 6. Comparison of mean numbers of double-labelled cells in P0, P1 and adult animals. Error bars indicate one standard deviation. Stars indicate statistically significant differences between both neonatal groups and adult animals.
Though the total number of double-labelled cells was small in all cases there were significantly more such cells in P0 (mean = 25, S.D. = 4.3) and P1 (mean = 35, S.D. = 10.3) animals compared to the adult (mean = 8.8, S.D. = 4.6; Fig. 6). The differences between P0/1 groups and the adult were
-1 -1 W 0
0
<
I-- Z 0 0
I , .J .< I.- o I.-
5 0 ~
4 0 ~
3 0 -
2 0 - -
l O m
C O N T R A L S O
_I PO P1 P4 P7/8 ADULT
Fig. 7. Comparison of mean numbers of contralateral LSO cells at various postnatal ages. Error bars indicate one standard deviation. Stars indicate statistically significant dif- ferences between adult and neonatal groups.
204
statistically significant (P < 0.01). There was no significant difference (P > 0.1) when Pt/ and PI
groups were compared. Successful bilateral injec-
tions were not achieved in P4 and P7/8 animals, and
the number of double-labelled cells could therefore
not be quantified in these groups. Most of the difference between PO/1 animals and adults in the
number of double-labelled cells was in the medial
nuclei of origin, but a small number (7) of double-
labelled cells was also found in the ipsilateral LSO in
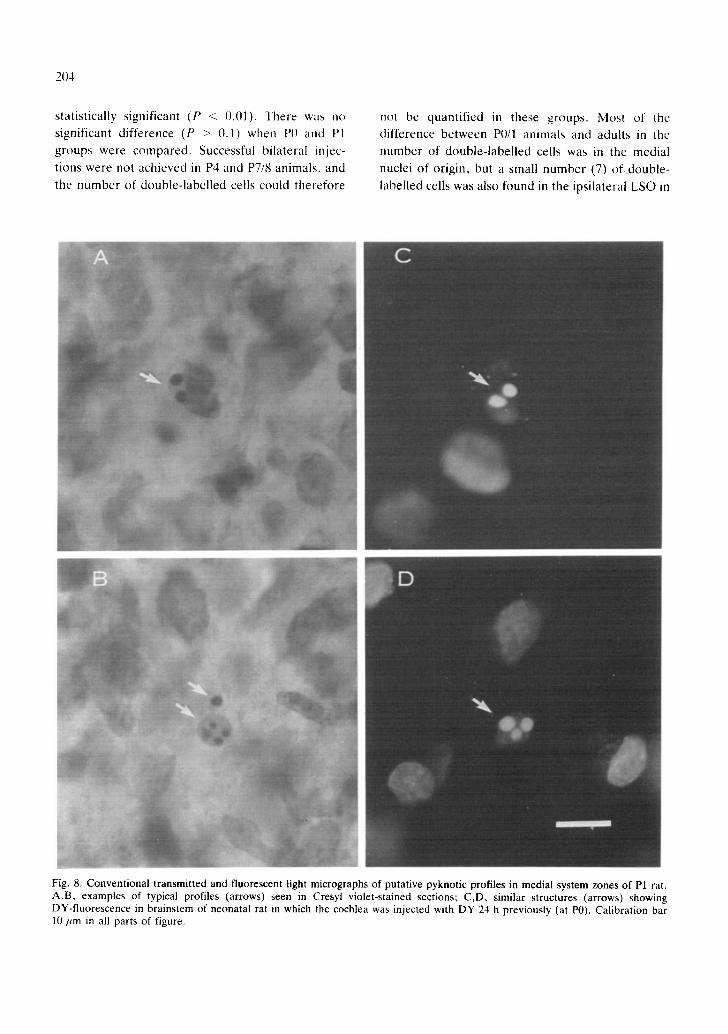
Fig. 8. Conventional transmitted and fluorescent light micrographs of putative pyknotic profiles in medial system zones of P1 rat. A,B, examples of typical profiles (arrows) seen in Cresyl violet-stained sections; C,D, similar structures (arrows) showing DY-fluorescence in brainstem of neonatal rat in which the cochlea was injected with DY 24 h previously (at P0). Calibration bar 10 um in all parts of figure.

3 out of 4 P0 animals. Three double-labelled LSO cells were also found in 4 P1 animals. Such cells were never found in the adult brainstem when diffuse contamination of the brainstem by FB was absent.
The number of single-labelled neurones found within the contralateral LSO at various postnatal ages is shown in Fig. 7. These data include both bilaterally and unilaterally-injected animals. There was a systematic decline in numbers of these cells from P0 (mean = 39, S.D. = 7.8) to adult (mean = 0.2, S.D. = 0.42). The mean number of these contralateral LSO cells was significantly higher than the adult in all postnatal groups except P7/8 (P > 0.05).
Evidence for neuronal death We obtained some evidence that cell death occurs
postnatally in all the regions of origin of the olivocochlear projection. Examination of Cresyl fast violet-stained sections revealed numbers of pyknotic profiles in both the LSO and rostral, and the ventral peri-olivary regions of the brainstem, indicative of neuronal degeneration 4'9"1°. Typical examples of
pyknotic cells in the medial system regions in a P1 animal are shown in Fig. 8A,B. These pyknotic profiles were also seen in lateral and medial regions of origin in Neutral red-stained sections. They were seen frequently in P0, P1 and P4 specimens. They were rare in P8 and P16 animals and absent in the adult. Furthermore, in fluorescence material, strik- ingly similar profiles could be observed stained intensely with DY in animals injected intracoch- learly with this tracer at P0-P4. Typical examples are shown in Fig. 8C,D. The striking similarity between these labelled structures and the pyknotic profiles seen with conventional staining clearly suggests that at least some olivocochlear neurones degenerate postnatally.
DISCUSSION
The present results show that in terms of the total numbers of efferent neurones projecting to the cochlea and the distribution of their cell bodies within the brainstem, there appears to be little difference between rats in which the cochlea is injected with retrograde tracer at P0 and at maturity. We obtained no evidence for any peak in the total
205
number of efferent neurones labelled after intra- cochlear injection at any of the postnatal ages investigated (Figs. 4, 5).
As far as the lateral efferent system is concerned, this result is consistent with previous observations at the electronmicroscopical level which report that axo-axonic synapses are present at birth in the target region of this system beneath the inner hair cells 12A6. Thus, intracochlear synaptic connections are proba- bly formed in utero by the lateral system. Moreover, certain details of the labelling are consistent with this early projection being tonotopically organized pre- natally. The brighter labelling of cells in the medial aspect of the ipsilateral LSO, which was found in cases of incomplete basal region injections in both the adult and P0 animals (Fig. 3), is consistent with these cells projecting to the high frequency basal turn of the cochlea, nearer the site of dye injection 17.
As far as the medial system is concerned, our findings show that axons of this system enter the cochlea in adult numbers at least 6 days before the adult pattern of synaptic organization of this sub- system, namely innervation of the outer hair cells, is thought to become established 12. This result in the rat is consistent with a recent report using light microscopy and histochemical staining 27, which de- scribed numbers of efferent-like fibres in the region of the outer hair cells in the newborn mouse cochlea.
However, further interpretation of these results is not simple, and our findings could be consistent with either one of two rather different views of develop- ment of the olivocochlear projection.
In utero maturation? One possible explanation for our results is that the
adult numbers of efferents of both systems have entered the cochlea in utero and that there is no major neuronal death or axon retraction during the postnatal maturation of the system. This interpreta- tion would require that the terminal processes of the medial system neurones can survive within the cochlea for at least 6 days before they establish contact with their final targets. Either these neu- rones can survive during this period without making synaptic connections with the outer hair cells, or they are sustained by synaptic contacts formed on some other target. In this respect, it is noteworthy that in the adult rat and cat, 70% or more of medial

206
efferent neurones have been reported to send col- laterals to the cochlear nucleus 5'~3, although another possibility is that the medial efferents could make transient synaptic contact with other intracochlear
elements before synapsing with the outer hair cells. This interpretation further implies that the adult
pattern of projection of the efferents to the periph- eral hearing organ is specified without any afferent electrical input to the brainstem nuclei where the efferent cell bodies reside. The first high threshold, broadly tuned afferent responses from the cochlea, and the first behavioral responses, occur in the rat at about P7-8, and the full adult pattern of sensitivity and frequency selectivity is established only by about the 16th postnatal day 6829.
Postnatal maturation? According to this alternative interpretation, neu-
ronal death and/or pruning of exuberant early projections 7'9'14"26'2~ does occur postnatally in this
system and the picture obtained with retrograde labelling is partly misleading. The total number of cells labelled by intracochlear injection early in postnatal development may appear to be close to the adult numbers because of a sequential pattern of arrival of efferent processes which keeps pace with a loss of nerve fibres due to neuronal death and/or collateral pruning. Interestingly, the present results provide some evidence in support of this second interpretation of a dynamic shaping of the system postnatatly.
Firstly, there were significantly higher numbers of double-labelled cells in the medial system in the early neonatal animals compared to the adult (Fig. 6). In addition, a small number of double-labelled cells were found in the neonatal LSO which were never seen in the adult. These bilaterally projecting cells may either die or undergo collateral pruning postnatally. Secondly there were cells located in the LSO contralaterai to the injected cochlea in early postnatal animals and not in adults (Fig. 7). These cells may also die during postnatal development or they may change their projection pattern in some way. Though the numbers involved are small, these differences between neonatal and adult animals were significant and they are suggestive of a more diffuse, less precise projection pattern in utero and in the early postnatal period, which is modified during
maturation. Lastly, we obtained clear evidence from pyknotic profiles that there is postnatal cell death occurring in all the nuclei of origin of both medial
and lateral olivocochlear systems. This does not of course mean that this death necessarily involves olivocochlear efferents, since the majority of neu-
rones in these nuclei belong to the ascending auditory pathways. However, the striking similarity between the pyknotic profiles seen in conventionally stained material and the abnormal DY-stained cells in neonatal brains (Fig. 8) strongly suggests that at least some of this cell death does indeed involve the olivocochlear efferents.
Taken together, these data suggest the possibility that there may be a significant amount of cell death of olivocochlear neurones and perhaps the retraction of axonal processes during the postnatal period. The apparently constant numbers of the major efferent
systems from P0 to adult may simply mask a dynamic situation and there may indeed be a surplus of efferent neurones in the brainstem at birth which is gradually reduced by cell death to reach the final adult numbers. Only a fraction of this early popu- lation may project to the cochlea at any one time and be able to be detected by intracochlear injection of retrograde label. This interpretation is consistent with what is known to occur in other systems and
may accord with the transient postnatal expression of NGF by intracochlear structures ~. In this regard, the role of NGF in cochlear innervation and in the induction of cholinergic enzymes in efferent neu- rones deserves further investigation.
If the postnatal development of the auditory brainstem-to-cochlea pathway is indeed a dynamic process, it is possible that this has functional signif- icance. Plasticity of the olivocochlear projection during a period coincident with the emergence of cochlear function and maturation of the ascending pathways might permit physiological modification of the efferent system in the first few days after birth (cf. ref. 23). This may be important in the estab- lishment of specific inputs to olivocochlear efferents and in determining their laterality of projection. It is known, for example, that medial olivocochlear ef- ferent neurones in adult mammals are sharply tuned to acoustic stimulation and project back to the receptor epithelium in a more or less precise tono- topic manner 13'2°'21. Furthermore, the laterality of

projection of at least some medial efferents appears
to be related to the cochlea from which they receive
their dominant input 22. A process of selective effer-
ent cell death and/or axonal el imination within the
brainstem during the period when connections with
afferent input are becoming established may well
provide a mechanism for ensuring that the appro-
priate neural circuitry is established between the
brainstem and the periphery.
REFERENCES
1 Abercrombie, M., Estimation of nuclear populations from microtome sections, Anat. Rec., 94 (1946) 239-247.
2 Aschoff, A. and Ostwald, J., Distribution of cochlear efferents and olivo-collicular neurons in the brainstem of rat and guinea pig, Exp. Brain Res., 71 (1988) 241-251.
3 Aschoff, A. and Ostwald, J., Different origin of cochlear efferents in some bats, rat and guinea pig, J. Comp. Neurol., 264 (1987) 56-72.
4 Beazley, L.D., Darby, J.E. and Perry, V.H., Cell death in the retinal ganglion cell layer during optic nerve regener- ation for the frog Rana pipiens, Vision Res., 26 (1986) 543-556.
5 Brown, M.C., Liberman, M.C. and Ryugo, D.K., La- belled collaterals of medial olivocochlear axons in the cochlear nucleus of cats and rodents, Abstracts of 11th Midwinter Meeting, Assoc. Res. Otolaryngol., (1988) 49.
6 Carlier, E., Lenoir, M. and Pujol, R., Development of cochlear frequency selectivity tested by compound action potential tuning curves, Hearing Res., 1 (1979) 197-201.
7 Cowan, W.M., Fawcett, J.W., O'Leary, D.D.M. and Stanfield, B.B., Regressive events in neurogenesis, Sci- ence, 225 (1984) 1258-1265.
8 Crowley, D.E. and Hepp-Reymond, M.-C., Development of cochlear function in the ear of the infant rat, J. Comp. Physiol. Psychol., 62 (1966) 427-432.
9 Cunningham, T.J., Naturally occurring neuron death and its regulation by developing neural pathways, Int. Rev. Cytol., 74 (1982) 163-186.
10 Cunningham, T.J., Mohler, I.M. and Giordano, D.L., Naturally occurring neuron death in the ganglion cell layer of the neonatal rat: morphology and evidence for regional correspondence with neuron death in superior colliculus, Dev. Brain Res., 2 (1981) 203-215.
11 Despres, G., Giry, N. and Romand, R., Immunohistoche- mical localisation of nerve growth factor-like protein in the organ of Corti of the developing rat, Neurosci. Lea., 85 (1988) 5-8.
12 Lenoir, M., Shnerson, A. and Pujol, R., Cochlear receptor development in the rat with emphasis on synaptogenesis, Anat. Embryol., 160 (1980) 253-262.
13 Liberman, M.C. and Brown, M.C., Physiology and anat- omy of single olivocochlear neurons in the cat, Hearing Res., 24 (1986) 17-36.
14 Oppenheim, R.W., Naturally occurring cell death during neural development, TINS, 8 (1985) 487-493.
15 Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Coordinates, 2rid edn., Academic, Australia, 1986.
207
ACKNOWLEDGEMENTS
This research was supported by separate grants to
B.M. Johnstone and A .R .H . from the National
Health and Medical Research Council (Australia)
and the University of Western Australia. The au-
thors are indebted to L.D. Beazley, B.M. John-
stone, W.F.C. Blumer, G.K. Yates and R.A. Rush
for helpful discussions, and K. Corbett and K.
Duffal for care of experimental animals.
16 Pujol, R., Carlier, E. and Devigne, C., Different patterns of cochlear innervation during the development of the kitten, J. Comp. Neurol., 177 (1978) 529-535.
17 Robertson, D., Anderson, C.-J. and Cole, K.S., Segrega- tion of efferent projections to different turns of the guinea pig cochlea, Hearing Res., 25 (1987) 69-76.
18 Robertson, D., Cole, K.S. and Corbett, K., Quantitative estimate of bilaterally projecting medial olivocochlear neurones in the guinea pig brainstem, Hearing Res., 27 (1987) 177-181.
19 Robertson, D., Cole, K.S. and Harvey, A.R., Brainstem organization of efferent projections to the guinea pig cochlea studied using the fluorescent tracers fast blue and diamidino yellow, Exp. Brain Res., 66 (1987) 449-457.
20 Robertson, D. and Gummer, M., Physiology of cochlear efferents in the mammal. In J. Syka and R.B. Masterton (Eds.), Auditory Pathway, Plenum, New York, 1988, pp. 269-278.
21 Robertson, D. and Gummer, M., Physiological and mor- phological characterization of efferent neurones in the guinea pig cochlea, Hearing Res., 20 (1985) 63-78.
22 Robertson, D. and Winter, I.M., Cochlear nucleus inputs to olivocochlear neurones revealed by combined antero- grade and retrograde labelling in the guinea pig, Brain Res., 462 (1988) 47-55.
23 Walsh, E.J. and McGee, J., Rhythmic discharge properties of caudal cochlear nucleus neurons during postnatal devel- opment in cats, Hearing Res., 36 (1988) 233-248.
24 Warr, W.B. and Guinan, J.J., Efferent innervation of the organ of Corti: two separate systems, Brain Res., 173 (1979) 152-155.
25 White, J.S. and Warr, W.B., The dual origins of the olivocochlear bundle in the albino rat, J. Comp. Neurol., 219 (1983) 203-214.
26 O'Leary, D.D.M., Stanfield, B.B. and Cowan, W.W.M., Evidence that the early postnatal restriction of the cells of origin of the callosal projection is due to the elimination of axonal collaterals rather than to the death of neurones, Dev. Brain Res., 1 (1981) 607-617.
27 Sobkowicz, H.M., Emmerling, M.R. and Whitlon, D.S., Development of cochlear efferents in the postnatal mouse, Abstracts 11th Midwinter Meeting, Assoc. Res. Otolaryn- gol., (1988) 131.
28 Tolbert, D.L., Intrinsically directed pruning as a mecha- nism regulating the elimination of transient collateral pathways, Dev. Brain Res., 33 (1987) 11-21.
29 Uziel, A., Romand, R. and Marot, M., Development of cochlear potentials in rats, Audiology, 20 (1981) 89-100.



![Automatic Cochlea Multi-modal Images Segmentation · 2018-04-03 · Automatic Cochlea Multi-modal Images Segmentation Al-Dhamari, CI2018 Methods: Cochlea Model 9 [5] Gerber et al,](https://img.pdfslide.us/doc/110x75/5f8e42f1fe0c2a0180250f2a/automatic-cochlea-multi-modal-images-segmentation-2018-04-03-automatic-cochlea.jpg)
























