Embed Size (px)
Citation preview

Research in Microbiology 164 (2013) 327e334www.elsevier.com/locate/resmic
Post-translational regulation of a developmental catalase, CatB, involvesa metalloprotease, SmpA and contributes to proper differentiation and
osmoprotection of Streptomyces coelicolor
Hyo-Sub Kim a, Eun-Jin Lee a,1, You-Hee Cho b,*, Jung-Hye Roe a
a School of Biological Sciences, Institute of Microbiology, Seoul National University, Seoul 151-742, Republic of KoreabDepartment of Pharmacy, College of Pharmacy, CHA University, Gyeonggi-do 463-840, Republic of Korea
Received 1 November 2012; accepted 17 December 2012
Available online 31 December 2012
Abstract
Streptomyces coelicolor produces at least three different catalases (catalases A, B, and C) under different physiological conditions. Catalase B(CatB) is a developmentally regulated catalase required for proper differentiation and osmoprotection of S. coelicolor. We previously observedthat the N-terminal 752 amino acids (aa) of CatB are cleaved off, with the remaining 75-kDa processed CatB detectable in the extracellularfraction during sporulation. We here report that either the deletion of the N-terminal 75 aa or the arginine-to-alanine substitution (R75A) at thecleavage site, but not the histidine-to-alanine substitution (H131A) at the catalytic site, impaired both the secretion of CatB proteins and theproper differentiation of S. coelicolor cells. The proteolytic activity responsible for the cleavage of CatB was purified and then identified asa metalloprotease, which was named as SmpA (Streptomyces metalloprotease A). The SmpA protein was newly detected after sporulation,coincident with the intracellular appearance of 75-kDa CatB, which was not detected in the smpA null mutant, confirming that SmpA indeedprocesses CatB in vivo. The smpA mutant was osmosensitive as catB mutant, but it displayed delayed sporulation, with the 75-kDa CatB stilldetectable in the extracellular milieu. Based on these results, we propose that the post-translational regulation of CatB, which cleaves the N-terminal 75 aa residues through SmpA is crucial for proper differentiation and osmoprotection of S. coelicolor. In the absence of SmpA, analternative route for CatB processing may function to allow delayed sporulation.� 2012 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved.
Keywords: Differentiation; Osmotic stress; Catalase; Metalloprotease; Processing; Streptomyces
1. Introduction
Most living organisms that are exposed to molecular oxy-gen (O2) produce common enzymes that include catalase.Catalase prevents the formation of highly reactive hydroxylradical from hydrogen peroxide (H2O2) by decomposing H2O2
into water and dioxygen. Most of the known catalases areclassified into one of three types, based on the enzymatic
* Corresponding author. Tel.: þ82 31 8017 9463; fax: þ82 31 8017 9420.
E-mail address: [email protected] (Y.-H. Cho).1 Present address: Section of Microbial Pathogenesis, Yale Microbial
Diversity Institute, Yale School of Medicine, New Haven, CT 06536, USA.2 The 75 aa was once designated as 95 aa in our previous study (Cho et al.,
2000), due to a mistaken duplication of 20 aa at the N-terminal region.
0923-2508/$ - see front matter � 2012 Institut Pasteur. Published by Elsevier Ma
http://dx.doi.org/10.1016/j.resmic.2012.12.009
properties: heme-containing monofunctional catalases, heme-containing bifunctional catalase-peroxidases, and non-hemepseudocatalases. Almost all bacterial species employ morethan one catalase. The role of the bacterial multiple catalaseshas not been comprehensively understood, whereas distinctphysiological role of each catalase has been suggested inStreptomyces coelicolor (Cho et al., 2000).
S. coelicolor producing at least three different catalases, isa Gram-positive spore-forming soil bacterium that exploits anintricate program of morphological and physiological differen-tiation resembling that of filamentous fungi. When S. coelicolorspores are germinated on solidmedia, the developmental processbegins to form complex vegetative mycelium by hyphalbranching. External cues such as nutrient limitation triggercells to escape from the vegetative growth into erecting aerial
sson SAS. All rights reserved.

328 H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
mycelia as the onset of differentiation. Concomitantly, manygenes in S. coelicolor are newly expressed for differentiationleading to morphological andmetabolic changes (Chater, 1993),which include a developmentally regulated catalase, CatB.
We previously reported that CatB is required for osmo-protection and proper differentiation of S. coelicolor J1501(Cho et al., 2000). The transcription of the catB promoter wasinduced by osmotic shock, being dependent on a stress-responsive sigma factor, sB (Cho et al., 2001). Besides thetranscriptional control by sB, post-translational control wasexerted on CatB expression during the development, whichincludes proteolytic cleavage and extracellular secretion ofCatB: at least three isoforms of 92, 87, and 75 kDa weredetected during prolonged growth on solid media and the 75-kDa CatB was predominantly detected in the extracellularfraction after sporulation (Cho et al., 2000). These two pro-cesses were coincident with morphological differentiation.However, the mechanism by which both processes are tem-porally and spatially coordinated in response to osmotic stressand developmental signals of S. coelicolor needs to be furtherinvestigated, which will elucidate a key event in the regulatorynetworks governing the developmental cycle of this bacterium.
In this study, we present the evidence that the processing ofCatB to cleave off the N-terminal 75-aa residues, but not itscatalase enzyme activity, lies behind the requirement of CatBin proper differentiation and osmoprotection of S. coelicolor.To further examine the role of the N-terminal region in theprocessing and secretion of CatB, we identified a metal-loprotease (SmpA) encoded by SCO2529 from the extrac-ellular fraction, which was predominantly required for thedevelopmental processing of CatB.
2. Materials and methods
2.1. Bacterial strains and culture conditions
Growth and maintenance of S. coelicolor A3(2) strains(J1501, smpA mutant, catB mutant, and their derivatives) weredone as described previously (Cho et al., 2000; Hopwoodet al., 1985) For plate surface culture, 107 pre-germinatedspores or patches of mycelia were streaked on R2YE, NA,or minimal agar media with 1% glucose as the sole carbonsource. To facilitate harvesting of mycelia from the platecultures, inocula were spread on cellophane discs on themedia. The growth rates and phases were determined asdescribed elsewhere (Cho and Roe, 1997).
2.2. DNA manipulations
Restriction and modifying enzymes were used accordingto the manufacturer’s recommendations (POSCOCHEM,Boeringer-Manheim, and NEB), using the standard recombi-nant DNA methods. DNA fragments were purified fromagarose gels using GeneClean Kit II (BIO101) or freeze-squeeze method. Escherichia coli DH5a, methylation-negative E. coli ET12567 (MacNeil, 1988) and J1501 cellswere used as hosts for various recombinant DNAs.
2.3. Complementation with catB derivatives andcreation of smpA mutant
The 3.5 kb wild-type copy of the catB gene (CatB-WT) wascloned into pSET152 and introduced into the catB mutant byconjugal transfer (MacNeil, 1988). CatB-DN was constructedby the deletion of the N-terminal 225 bp fragment of the CatB-WT, and CatB-H131A and CatB-R75A were constructed bysite directed mutagenesis, based on splicing by overlappingextension (SOEing) PCR using the primer pairs (50-CGG AGAAAG GCATAT GGC GCA GTC C-30 and 50-GGA CTG CGCCAT ATG CCT TTC TCC G-30 for DN; 50-GCG CGT GGTCGC CGC GAA GGG CGC-30 and 50-GC GCC CTT CGCGGC GAC CAC GCG C-30 for H131A and 50-GCC GACCAG GCC CGG GCC GCG CAG TCC-30 and 50-GGA CTGCGC GGC CCG GGC CTG GTC GGC-30 for R75A; codonsubstitution positions underlined). These derivatives werecloned into pSET152 and then introduced into E. coliET12567 containing the conjugation driver plasmid,pUZ8002. Conjugal transfer of each clone from E. coli to S.coelicolor was performed (Mazodier et al., 1989). Exconju-gants were selected by single colony isolation the expectedgene structure was verified by PCR and Southern blot ana-lyses. To create a null mutant for SCO2529 (smpA), REDI-RECT PCR-targeting method was used with the followingprimers (50-ATG ACG CCA CAC GGG GGC TTC CAG CCCGTC TTC TGC ACG ATT CCG GGG ATC CGT CGA CC-30
and 50-CAC CTG AAT CCG CAT GGA CCC GTG TCTAGTACG GAA CTA TGT AGG CTG GAG CTG CTT C-30) asdescribed elsewhere (Gust et al., 2003; Lee et al., 2005).
2.4. Catalase profiling
To confirm the absence of CatB activity in the catB mutantexpressing CatB-H131A, the crude extracts of the catB mutantcells expressing either CatB-WTor CatB-H131A that had beengrown to the stationary phase in YEME liquid media wereprepared and subjected to Q-Sepharose ion exchange columnchromatography (Amersham Pharmacia). The bound proteinswere washed with TGED buffer (10 mM Tris, 20% Glycerol,1 mM EDTA, and 0.1 mM DTT) containing 50 mM NaCl, andthen eluted using a gradient of 300e500 mM NaCl in TGED at0.5ml/min into 15 fractions, each of which was dialyzed againststorage buffer (25 mM KCl, 10 mM Tris, 50% Glycerol, 1 mMEDTA, and 0.1 mM DTT) and tested for catalase activity basedon spectrophotometry at 280 nm (Beers and Sizer, 1952).
2.5. Purification and identification of SmpA
J1501 cells were grown for 10 days on 100 R2YE platescovered with cellophane disc, and the plate agar was chopped forelution of extracellular proteins using 50 mM NaCl-TGEDbuffer. After centrifugation at 12,000 rpm for 30 min at 4 �C,the supernatant was subjected to precipitation with 2 volume of100% saturated (NH4)2SO4 solution (pH 7.9). After precipita-tion, the pellet was dissolved in TED (TGED minus glycerol)and then applied to a Q-Sepharose column (Amersham

329H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
Pharmacia). After washing with 50 mM NaCl-TGED, proteinswere eluted using a gradient of 50e800 mM NaCl in TGEDbuffer at 0.5 ml/min. Ten microliter of each fraction was sub-jected to in vitro CatB processing assay using C-terminal trun-cated CatB derivative (CatB-42). Hexahistidine (His)-taggedCatB-42 was overproduced and purified from E. coli BL21containing the truncated catB gene cloned in pET21-c, whichhad been obtained from NdeI and SalI digestion of pET3965(Cho et al., 2000). CatB-42 processing was resolved on 13%SDS-PAGE. CatB-processing activity was detected in the frac-tions eluted between 500 and 800mMNaCl, which were pooledand subjected to (NH4)2SO4 precipitation (60% w/v). Theresulting precipitate was dissolved in 500 ml of 50 mM NaCl-TGED buffer and then applied to a Superdex G-75 column(Amersham Pharmacia) and eluted with 50 mM NaCl-TGEDbuffer. Each fraction (10 ml) eluted by 50 mM NaCl-TGEDbuffer was subjected to in vitro CatB-processing assay fol-lowed by SDS-PAGE. The fractions with CatB-processing ac-tivity eluted between 83 and 93 ml were pooled and dialyzedagainst storage buffer.
Partially purified CatB-processing activity was resolved on13% SDS-PAGE. After staining by Coomassie brilliant blue(CBB) R-250 staining (BioRad), the protein bands were sliced,washed using water, and then treated with 200 mM ammoniumbicarbonate for 20 min. The gel slices were chopped, washedwith water, dehydrated using acetonitrile, and then subjected totrypsin (5e10 ng/ml) digestion for over 12 h. The digestedpeptides were eluted with 10e20 ml of 0.1% of trifluoroaceticacid in 50% acetonitrile and then concentrated for mass spec-trometry (Voyager-DETM STR Biospectrometry Workstation,Applied Biosystems Inc.). Protein identification based on themeasured peptide masses was performed using the MASSCOTprogram (www.matrixscience.com/search_form_select.html).
2.6. Preparation of anti-SmpA antibody
To prepare C-terminal His-tagged SmpA proteins, themutagenic forward (50-GCG AGA GTG CCG CATATG ACGCCA CAC-30, NdeI site underlined) and reverse (50-GTG GTGGTG GTG GTG GTG CAG CGT CCG CAC CCC GAC-30,His-tag sequences underlined) primers were used to amplifya 1101 bp fragment containing the entire smpA gene. The PCRproduct containing the smpA gene was cloned into pGEM-TEasy vector (Promega), recovered with NdeI and BamHIdigestion, and then cloned into pET-21c (Novagen). His-tagged proteins were overproduced in E. coli BL21 and pu-rified according to the manufacturer’s instruction (Novagen).Polyclonal antibodies against SmpA were prepared as descri-bed previously (Cho et al., 2000).
2.7. Western blot analysis
Total proteins from surface-grown mycelial cultures wereresolved on 10% (for CatB) and 12% (for SmpA) SDS-PAGE.After SDS-PAGE, the gel was soaked in transfer buffer(25 mM Tris, 192 mM glycine, 20% (v/v) methanol) for10 min, and then electrotransferred to nitrocellulose
membrane BA79 (Schleicher & Schuell) at 160 mA for 60 minin Trans-Blot Cell (Bio-Rad). Membranes were blocked inTris-buffered saline buffer containing 0.1% Triton X-100(TBST) supplemented with 0.5% BSA, for more than 1 h. Theblocked membrane was incubated with primary antisera for1 h, and then washed twice with TBST for 10 min. The washedmembrane was incubated with goat anti-mouse IgGAM(Cappell) conjugated with horseradish peroxidase and washedtwice with TBST for 10 min. Detection was done usingWestern ECL detection system (Amersham Pharmacia) andLAS-3000 imaging device (FUJI film).
For extracellular protein analysis, bacteria were grown onR2YE solid medium implemented with cellophane discs. Atthe designated time points after inoculation, cellophane discsand bacterial cells were removed and the agar blocks wereobtained, chopped into small pieces, and then soaked into50 mM potassium phosphate (KP) buffer containing 1 mMPMSF (phenylmethylsulfonyl fluoride) for 1 h at 4 �C. Aftercentrifugation at 11,000 rpm for 30 min, agar debris werediscarded and the supernatants were concentrated using(NH4)2SO4 (60% w/v). Precipitates were dissolved in50 mM kP buffer, further concentrated using TCA (20% v/v),and then dissolved in TriseHCl buffer (pH 8.0), prior to SDS-PAGE followed by Western blot analyses.
2.8. Plate-lift immunoassay
For plate-lift immunoassay, a nitrocellulose membraneBA79 (Schleicher & Schuell) was carefully blotted onto cul-ture plates of bacterial cells on R2YE solid media for at least10 min, and treated as described elsewhere (Viollier et al.,2003; Willey et al., 1991). The membrane was blockedusing TBS-T containing 1.5% BSA and then subjected toimmunodetection as described above.
2.9. Zymography
J1501 and smpA mutant cells were grown on R2YE platescovered with cellophane disc, and the agar blocks were used forassay the extracellular metalloprotease activity. Extracellularproteins were obtained by soaking with 50 mM potassiumphosphate (KP) buffer for 1 h at 4 �C and the supernatant wassubjected to (NH4)2SO4 precipitation (60% w/v). Precipitateswere dialyzed against 50 mM kP buffer, and separated on 12%SDS-PAGE containing gelatin (1/10 volume). The gel waswashed using 2.5% Triton X-100 at room temperature for 1.5 h,incubated in developing buffer (50mMTriseHCl, 1.15%TritonX-100, 200 mM NaCl, 5 mM CaCl2, and 1 mM ZnCl2) for 48 hand then stained with coomassie brilliant blue (CBB).
3. Results
3.1. Construction of CatB derivatives
The detection of the N-terminally processed CatB of75 kDa (CatB-75) in sporulated mycelia led us to evaluate thesignificance of the processing event in differentiation. We
330 H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
constructed truncated forms of the wild type CatB that aredevoid of the N-terminal processed region (CatB-DN). Wealso created two codon substitution mutants for the 75tharginine residue at the cleavage site to alanine (CatB-R75A),or for the 131st histidine residue at the catalytic site (Gouetet al., 1995) to alanine (CatB-H131A). We cloned the wildtype (CatB-WT) and the mutated catB genes into the pSET152vector and conjugally transferred them to the catB null mutantstrain (YD9147) which does not form aerial mycelium (i.e.bald phenotype) and exhibits osmosensitivity (Cho et al.,2000), to examine whether the CatB derivatives were pro-cessed and/or secreted and complemented the bald andosmosensitive phenotype of catB mutant.
3.2. The processing mutants do not support secretion,differentiation, and osmoprotection
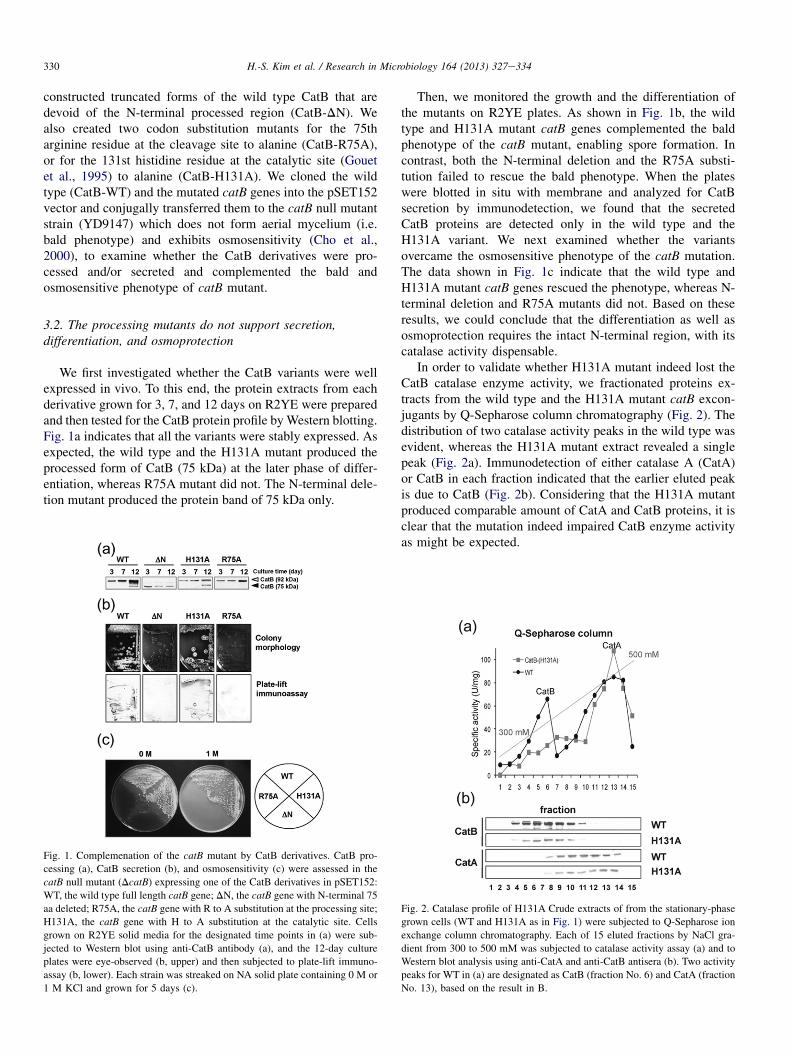
We first investigated whether the CatB variants were wellexpressed in vivo. To this end, the protein extracts from eachderivative grown for 3, 7, and 12 days on R2YE were preparedand then tested for the CatB protein profile byWestern blotting.Fig. 1a indicates that all the variants were stably expressed. Asexpected, the wild type and the H131A mutant produced theprocessed form of CatB (75 kDa) at the later phase of differ-entiation, whereas R75A mutant did not. The N-terminal dele-tion mutant produced the protein band of 75 kDa only.
Fig. 1. Complemenation of the catB mutant by CatB derivatives. CatB pro-
cessing (a), CatB secretion (b), and osmosensitivity (c) were assessed in the
catB null mutant (DcatB) expressing one of the CatB derivatives in pSET152:
WT, the wild type full length catB gene; DN, the catB gene with N-terminal 75
aa deleted; R75A, the catB gene with R to A substitution at the processing site;
H131A, the catB gene with H to A substitution at the catalytic site. Cells
grown on R2YE solid media for the designated time points in (a) were sub-
jected to Western blot using anti-CatB antibody (a), and the 12-day culture
plates were eye-observed (b, upper) and then subjected to plate-lift immuno-
assay (b, lower). Each strain was streaked on NA solid plate containing 0 M or
1 M KCl and grown for 5 days (c).
Then, we monitored the growth and the differentiation ofthe mutants on R2YE plates. As shown in Fig. 1b, the wildtype and H131A mutant catB genes complemented the baldphenotype of the catB mutant, enabling spore formation. Incontrast, both the N-terminal deletion and the R75A substi-tution failed to rescue the bald phenotype. When the plateswere blotted in situ with membrane and analyzed for CatBsecretion by immunodetection, we found that the secretedCatB proteins are detected only in the wild type and theH131A variant. We next examined whether the variantsovercame the osmosensitive phenotype of the catB mutation.The data shown in Fig. 1c indicate that the wild type andH131A mutant catB genes rescued the phenotype, whereas N-terminal deletion and R75A mutants did not. Based on theseresults, we could conclude that the differentiation as well asosmoprotection requires the intact N-terminal region, with itscatalase activity dispensable.
In order to validate whether H131A mutant indeed lost theCatB catalase enzyme activity, we fractionated proteins ex-tracts from the wild type and the H131A mutant catB excon-jugants by Q-Sepharose column chromatography (Fig. 2). Thedistribution of two catalase activity peaks in the wild type wasevident, whereas the H131A mutant extract revealed a singlepeak (Fig. 2a). Immunodetection of either catalase A (CatA)or CatB in each fraction indicated that the earlier eluted peakis due to CatB (Fig. 2b). Considering that the H131A mutantproduced comparable amount of CatA and CatB proteins, it isclear that the mutation indeed impaired CatB enzyme activityas might be expected.
Fig. 2. Catalase profile of H131A Crude extracts of from the stationary-phase
grown cells (WT and H131A as in Fig. 1) were subjected to Q-Sepharose ion
exchange column chromatography. Each of 15 eluted fractions by NaCl gra-
dient from 300 to 500 mM was subjected to catalase activity assay (a) and to
Western blot analysis using anti-CatA and anti-CatB antisera (b). Two activity
peaks for WT in (a) are designated as CatB (fraction No. 6) and CatA (fraction
No. 13), based on the result in B.
331H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
3.3. Isolation of a CatB-processing protease, SmpA
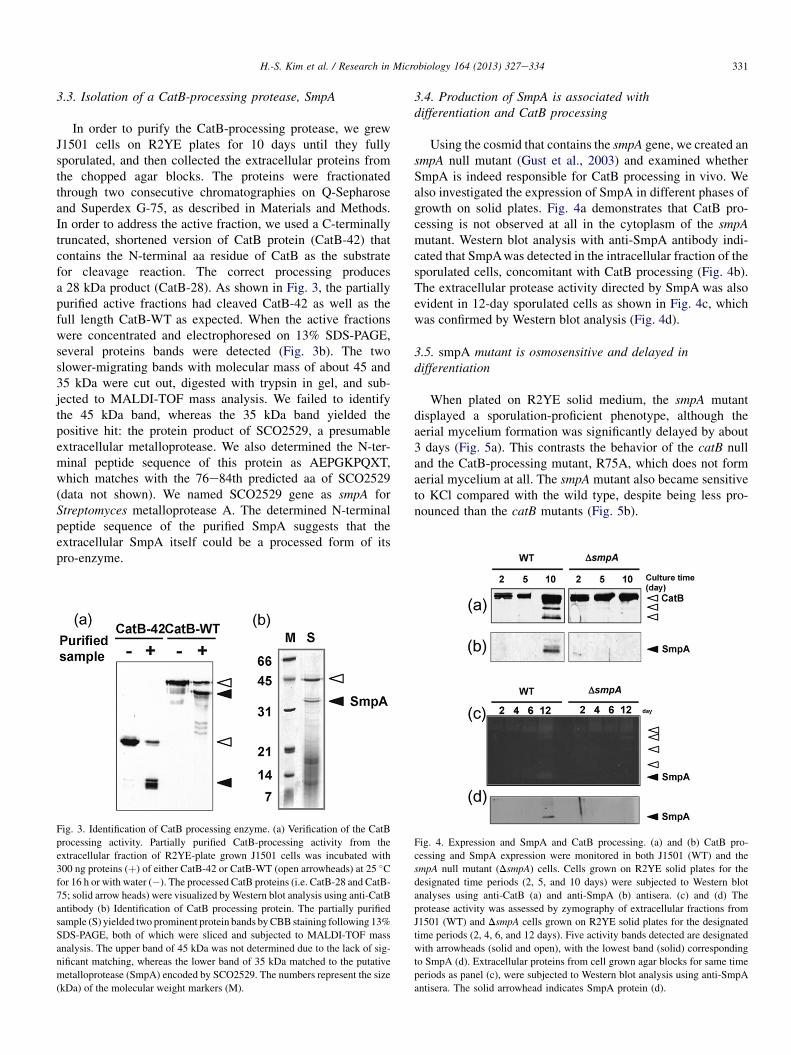
In order to purify the CatB-processing protease, we grewJ1501 cells on R2YE plates for 10 days until they fullysporulated, and then collected the extracellular proteins fromthe chopped agar blocks. The proteins were fractionatedthrough two consecutive chromatographies on Q-Sepharoseand Superdex G-75, as described in Materials and Methods.In order to address the active fraction, we used a C-terminallytruncated, shortened version of CatB protein (CatB-42) thatcontains the N-terminal aa residue of CatB as the substratefor cleavage reaction. The correct processing producesa 28 kDa product (CatB-28). As shown in Fig. 3, the partiallypurified active fractions had cleaved CatB-42 as well as thefull length CatB-WT as expected. When the active fractionswere concentrated and electrophoresed on 13% SDS-PAGE,several proteins bands were detected (Fig. 3b). The twoslower-migrating bands with molecular mass of about 45 and35 kDa were cut out, digested with trypsin in gel, and sub-jected to MALDI-TOF mass analysis. We failed to identifythe 45 kDa band, whereas the 35 kDa band yielded thepositive hit: the protein product of SCO2529, a presumableextracellular metalloprotease. We also determined the N-ter-minal peptide sequence of this protein as AEPGKPQXT,which matches with the 76e84th predicted aa of SCO2529(data not shown). We named SCO2529 gene as smpA forStreptomyces metalloprotease A. The determined N-terminalpeptide sequence of the purified SmpA suggests that theextracellular SmpA itself could be a processed form of itspro-enzyme.
Fig. 3. Identification of CatB processing enzyme. (a) Verification of the CatB
processing activity. Partially purified CatB-processing activity from the
extracellular fraction of R2YE-plate grown J1501 cells was incubated with
300 ng proteins (þ) of either CatB-42 or CatB-WT (open arrowheads) at 25 �Cfor 16 h or with water (�). The processed CatB proteins (i.e. CatB-28 and CatB-
75; solid arrow heads) were visualized byWestern blot analysis using anti-CatB
antibody (b) Identification of CatB processing protein. The partially purified
sample (S) yielded two prominent protein bands by CBB staining following 13%
SDS-PAGE, both of which were sliced and subjected to MALDI-TOF mass
analysis. The upper band of 45 kDa was not determined due to the lack of sig-
nificant matching, whereas the lower band of 35 kDa matched to the putative
metalloprotease (SmpA) encoded by SCO2529. The numbers represent the size
(kDa) of the molecular weight markers (M).
3.4. Production of SmpA is associated withdifferentiation and CatB processing
Using the cosmid that contains the smpA gene, we created ansmpA null mutant (Gust et al., 2003) and examined whetherSmpA is indeed responsible for CatB processing in vivo. Wealso investigated the expression of SmpA in different phases ofgrowth on solid plates. Fig. 4a demonstrates that CatB pro-cessing is not observed at all in the cytoplasm of the smpAmutant. Western blot analysis with anti-SmpA antibody indi-cated that SmpAwas detected in the intracellular fraction of thesporulated cells, concomitant with CatB processing (Fig. 4b).The extracellular protease activity directed by SmpA was alsoevident in 12-day sporulated cells as shown in Fig. 4c, whichwas confirmed by Western blot analysis (Fig. 4d).
3.5. smpA mutant is osmosensitive and delayed indifferentiation
When plated on R2YE solid medium, the smpA mutantdisplayed a sporulation-proficient phenotype, although theaerial mycelium formation was significantly delayed by about3 days (Fig. 5a). This contrasts the behavior of the catB nulland the CatB-processing mutant, R75A, which does not formaerial mycelium at all. The smpA mutant also became sensitiveto KCl compared with the wild type, despite being less pro-nounced than the catB mutants (Fig. 5b).
Fig. 4. Expression and SmpA and CatB processing. (a) and (b) CatB pro-
cessing and SmpA expression were monitored in both J1501 (WT) and the
smpA null mutant (DsmpA) cells. Cells grown on R2YE solid plates for the
designated time periods (2, 5, and 10 days) were subjected to Western blot
analyses using anti-CatB (a) and anti-SmpA (b) antisera. (c) and (d) The
protease activity was assessed by zymography of extracellular fractions from
J1501 (WT) and DsmpA cells grown on R2YE solid plates for the designated
time periods (2, 4, 6, and 12 days). Five activity bands detected are designated
with arrowheads (solid and open), with the lowest band (solid) corresponding
to SmpA (d). Extracellular proteins from cell grown agar blocks for same time
periods as panel (c), were subjected to Western blot analysis using anti-SmpA
antisera. The solid arrowhead indicates SmpA protein (d).
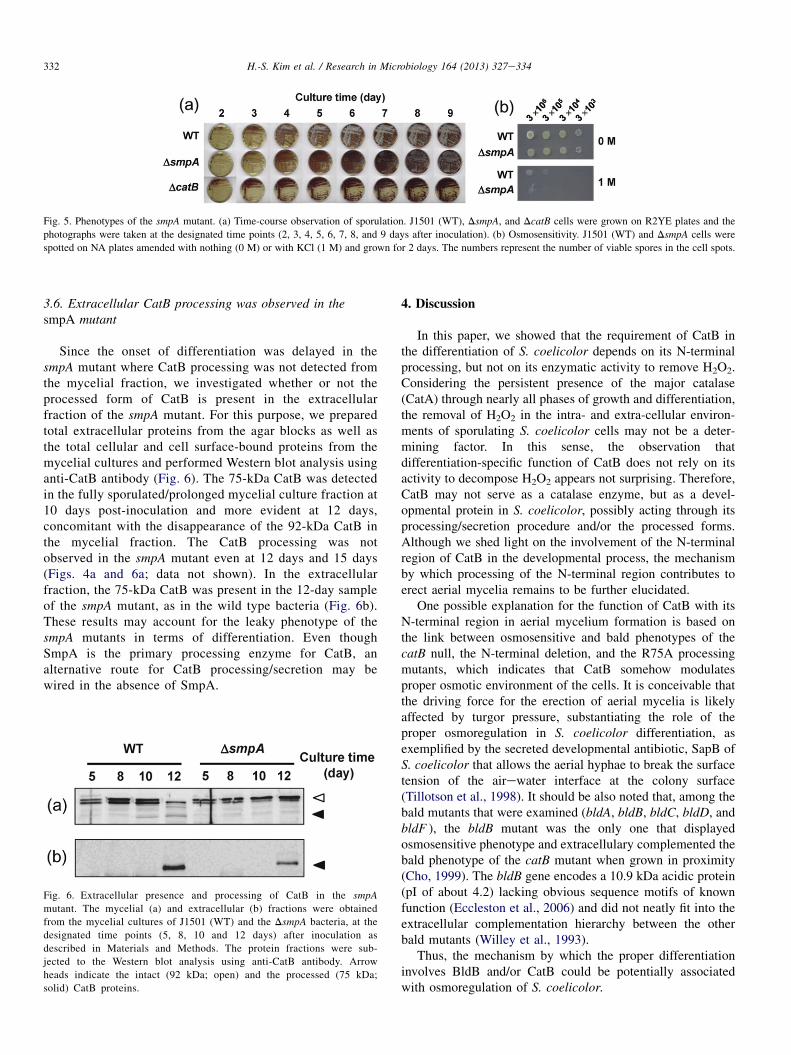
Fig. 5. Phenotypes of the smpA mutant. (a) Time-course observation of sporulation. J1501 (WT), DsmpA, and DcatB cells were grown on R2YE plates and the
photographs were taken at the designated time points (2, 3, 4, 5, 6, 7, 8, and 9 days after inoculation). (b) Osmosensitivity. J1501 (WT) and DsmpA cells were
spotted on NA plates amended with nothing (0 M) or with KCl (1 M) and grown for 2 days. The numbers represent the number of viable spores in the cell spots.
332 H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
3.6. Extracellular CatB processing was observed in thesmpA mutant
Since the onset of differentiation was delayed in thesmpA mutant where CatB processing was not detected fromthe mycelial fraction, we investigated whether or not theprocessed form of CatB is present in the extracellularfraction of the smpA mutant. For this purpose, we preparedtotal extracellular proteins from the agar blocks as well asthe total cellular and cell surface-bound proteins from themycelial cultures and performed Western blot analysis usinganti-CatB antibody (Fig. 6). The 75-kDa CatB was detectedin the fully sporulated/prolonged mycelial culture fraction at10 days post-inoculation and more evident at 12 days,concomitant with the disappearance of the 92-kDa CatB inthe mycelial fraction. The CatB processing was notobserved in the smpA mutant even at 12 days and 15 days(Figs. 4a and 6a; data not shown). In the extracellularfraction, the 75-kDa CatB was present in the 12-day sampleof the smpA mutant, as in the wild type bacteria (Fig. 6b).These results may account for the leaky phenotype of thesmpA mutants in terms of differentiation. Even thoughSmpA is the primary processing enzyme for CatB, analternative route for CatB processing/secretion may bewired in the absence of SmpA.
Fig. 6. Extracellular presence and processing of CatB in the smpA
mutant. The mycelial (a) and extracellular (b) fractions were obtained
from the mycelial cultures of J1501 (WT) and the DsmpA bacteria, at the
designated time points (5, 8, 10 and 12 days) after inoculation as
described in Materials and Methods. The protein fractions were sub-
jected to the Western blot analysis using anti-CatB antibody. Arrow
heads indicate the intact (92 kDa; open) and the processed (75 kDa;
solid) CatB proteins.
4. Discussion
In this paper, we showed that the requirement of CatB inthe differentiation of S. coelicolor depends on its N-terminalprocessing, but not on its enzymatic activity to remove H2O2.Considering the persistent presence of the major catalase(CatA) through nearly all phases of growth and differentiation,the removal of H2O2 in the intra- and extra-cellular environ-ments of sporulating S. coelicolor cells may not be a deter-mining factor. In this sense, the observation thatdifferentiation-specific function of CatB does not rely on itsactivity to decompose H2O2 appears not surprising. Therefore,CatB may not serve as a catalase enzyme, but as a devel-opmental protein in S. coelicolor, possibly acting through itsprocessing/secretion procedure and/or the processed forms.Although we shed light on the involvement of the N-terminalregion of CatB in the developmental process, the mechanismby which processing of the N-terminal region contributes toerect aerial mycelia remains to be further elucidated.
One possible explanation for the function of CatB with itsN-terminal region in aerial mycelium formation is based onthe link between osmosensitive and bald phenotypes of thecatB null, the N-terminal deletion, and the R75A processingmutants, which indicates that CatB somehow modulatesproper osmotic environment of the cells. It is conceivable thatthe driving force for the erection of aerial mycelia is likelyaffected by turgor pressure, substantiating the role of theproper osmoregulation in S. coelicolor differentiation, asexemplified by the secreted developmental antibiotic, SapB ofS. coelicolor that allows the aerial hyphae to break the surfacetension of the airewater interface at the colony surface(Tillotson et al., 1998). It should be also noted that, among thebald mutants that were examined (bldA, bldB, bldC, bldD, andbldF ), the bldB mutant was the only one that displayedosmosensitive phenotype and extracellulary complemented thebald phenotype of the catB mutant when grown in proximity(Cho, 1999). The bldB gene encodes a 10.9 kDa acidic protein(pI of about 4.2) lacking obvious sequence motifs of knownfunction (Eccleston et al., 2006) and did not neatly fit into theextracellular complementation hierarchy between the otherbald mutants (Willey et al., 1993).
Thus, the mechanism by which the proper differentiationinvolves BldB and/or CatB could be potentially associatedwith osmoregulation of S. coelicolor.

333H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
It was quite unexpected that the R75A processing mutantwas undetectable in the extracellular milieu. This secretionfailure could be attributed to the block in developmentalprocessing of the mutant. Many successive events during thebacterial sporulation especially those involving different celltypes (for example, fore spores and mother cells in Bacillussubtilis) can be temporally coordinated in each cell type, ina manner to cross-check the completion of the events thatoccur in the other cell type (Shank and Kolter, 2011). We havenot yet delved into the secretion/processing of CatB in dif-ferent cell types (i.e. substrate mycelia and aerial mycelia),which may enable us to dismantle the precise time and spacethat involve the post-translational regulation of CatB and/orcan be affected by R75A substitution as well. It is also feasiblethat the R75A mutation could hamper the function of the N-terminal region and/or affect the interaction between the CatBproteins and the secretion machinery. More systematic inves-tigation of the role of R75 and other residues in the N-terminalregion of CatB in either of the two presumably linked post-translational steps (e.g., secretion and processing) is in need.
The observation that SmpA is the major processing enzymefor CatB is quite clear, which is consistent with the fact thatSmpA proteins are detected when cells are sporulating, con-comitant with the timing of CatB processing. Due to diffi-culties in isolating intact mRNA samples, we were unable toexamine the smpA transcripts in the plate culture grown formore than a week. However, when grown in liquid culture, theproduction of smpA mRNA occurs when cells enter the sta-tionary growth phase as assessed by S1 mapping (data notshown). This observation agrees with its production duringdifferentiation on solid medium observed by immunoblotanalysis. The delay in aerial mycelium formation of smpAmutant may agree with the observation that processing of CatBwas impaired, but not completely, in the absence of SmpA.Compared with the bald and severe osmosensitive phenotypesof catB null and the R75A mutants, the milder phenotypes ofthe smpA mutant suggest that the smpA phenotypes wereattributed to the improper CatB processing, with regard totime, space and/or amount, may have been compensated byother back-up or alternative protease(s) in the mutant. It is alsolikely that the absence of SmpA protease may affect the cellphysiology in a broader sense, considering that smpA mutantdisplayed partial growth defect on minimal agar media, whichis not a phenotype of catB mutant (data not shown). Recently,SmpA has been identified as a member of the BldD regulonthat include over 20 well-known genes that have been previ-ously known to play important roles in developmental pro-cesses (den Hengst et al., 2010). SmpA is also suggested to beunder the direct control by AfsQ1, which is important inantibiotic biosynthesis and metabolic homeostasis (Wanget al., 2013). Thus, the function of SmpA involving proteinturnover is to support prototropic growth as well as timelydifferentiation of S. coelicolor.
Based on the facts that protein processing is widelyemployed in bacterial differentiation (Haldenwang, 1995;Rudner et al., 1999; Stragier et al., 1988; Viollier et al., 2003)and that the smpA and the catB genes are well conserved in
other Streptomyces species such as Streptomyces avermitilis,Streptomyces griseus, Streptomyces clavuligerus and Strepto-myces scabies (http://strepdb.streptomyces.org.uk/), an inter-esting avenue for future investigation would be to elucidate thekey residues and interacting partners of the CatB N-terminalregion that enable the post-translational control of CatB inregards to differentiation and osmoprotection. Further exami-nation of the expression, processing, and secretion of SmpA andother related proteases, as well as the identification of theirtarget proteins, promises to provide major insights into theconserved role of the bacterial protease cascade in the devel-opmental process of Streptomyces species (Chater et al., 2010).
Acknowledgment
This works was supported by a grant for NRL of MolecularMicrobiology to J.-H. Roe. (NRF-2009-0079278). H.-S. Kimwas supported by the second-stage of BK21 Program for LifeScience at SNU.
References
Beers Jr., R.F., Sizer, I.W., 1952. A spectrophotometric method for measuring
the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 195,
133e140.Chater, K.F., 1993. Genetics of differentiation in Streptomyces. Annu. Rev.
Microbiol. 47, 685e713.
Chater, K.F., Biro, S., Lee, K.-J., Palmer, T., Schrempf, H., 2010. The complex
extracellular biology of Streptomyces. FEMS Microbiol. Rev. 34,
171e198.
Cho, Y.-H., 1999. Gene Expression and the Role of Catalases in Streptomces
coelicolor A3(2). Seoul National University.
Cho, Y.-.H., Lee, E.-J., Ahn, B.-E., Roe, J.-H., 2001. SigB, an RNA poly-
merase sigma factor required for osmoprotection and proper differentiation
of Streptomyces coelicolor. Mol. Microbiol. 42, 205e214.
Cho, Y.-H., Lee, E.-J., Roe, J.-H., 2000. A developmentally regulated catalase
required for proper differentiation and osmoprotection of Streptomyces
coelicolor. Mol. Microbiol. 35, 150e160.
Cho, Y.-H., Roe, J.-H., 1997. Isolation and expression of the catA gene
encoding the major vegetative catalase in Streptomyces coelicolor Muller.
J. Bacteriol. 179, 4049e4052.
den Hengst, C.D., Tran, N.T., Bibb, M.J., Chandra, G., Leskiw, B.K.,
Buttner, M.J., 2010. Genes essential for morphological development and
antibiotic production in Streptomyces coelicolor are targets of BldD during
vegetative growth. Mol. Microbiol. 78, 361e379.
Eccleston, M., Willems, A., Beveridge, A., Nodwell, J.R., 2006. Critical res-
idues and novel effects of overexpression of the Streptomyces coelicolor
developmental protein BldB: evidence for a critical interacting partner. J.
Bacteriol. 188, 8189e8195.
Gouet, P., Jouve, H.M., Dideberg, O., 1995. Crystal structure of Proteus
mirabilis PR catalase with and without bound NADPH. J. Mol. Biol. 249,
933e954.
Gust, B., Challis, G.L., Fowler, K., Kieser, T., Chater, K.F., 2003. PCR-tar-
geted Streptomyces gene replacement identifies a protein domain needed
for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad.
Sci. U S A 100, 1541e1546.
Haldenwang, W.G., 1995. The sigma factors of Bacillus subtilis. Microbiol.
Rev. 59, 1e30.
Hopwood, D.A., Bibb, M.J., Chater, K.F., Kieser, T., Bruton, C.J.,
Kieser, H.M., 1985. Genetic Manipulation of Streptomyces, a Laboratory
Manual. The John Innes Foundation, Norwich.
Lee, E.-J., Karoonuthaisiri, N., Kim, H.-S., Park, J.-H., Cha, C.-J., Kao, C.M.,
Roe, J.-H., 2005. A master regulator sigmaB governs osmotic and

334 H.-S. Kim et al. / Research in Microbiology 164 (2013) 327e334
oxidative response as well as differentiation via a network of sigma factors
in Streptomyces coelicolor. Mol. Microbiol. 57, 1252e1264.
MacNeil, D.J., 1988. Characterization of a unique methyl-specific restriction
system in Streptomyces avermitilis. J. Bacteriol. 170, 5607e5612.
Mazodier, P., Petter, R., Thompson, C., 1989. Intergeneric conjugation between
Escherichia coli and Streptomyces species. J. Bacteriol. 171, 3583e3585.
Rudner, D.Z., Fawcett, P., Losick, R., 1999. A family of membrane-embedded
metalloproteases involved in regulated proteolysis of membrane-associated
transcription factors. Proc. Natl. Acad. Sci. U S A 96, 14765e14770.
Shank, E.A., Kolter, R., 2011. Extracellular signaling and multicellularity in
Bacillus subtilis. Curr. Opin. Microbiol. 14, 741e747.
Stragier, P., Bonamy, C., Karmazyn-Campelli, C., 1988. Processing of
a sporulation sigma factor in Bacillus subtilis: how morphological struc-
ture could control gene expression. Cell 52, 697e704.
Tillotson, R.D., Wosten, H.A., Richter, M., Willey, J.M., 1998. A surface
active protein involved in aerial hyphae formation in the filamentous
fungus Schizophillum commune restores the capacity of a bald mutant of
the filamentous bacterium Streptomyces coelicolor to erect aerial struc-
tures. Mol. Microbiol. 30, 595e602.
Viollier, P.H., Weihofen, A., Folcher, M., Thompson, C.J., 2003. Post-tran-
scriptional regulation of the Streptomyces coelicolor stress responsive
sigma factor, SigH, involves translational control, proteolytic processing,
and an anti-sigma factor homolog. J. Mol. Biol. 325, 637e649.
Wang,R.,Mast,Y.,Wang, J., Zhang,W., Zhao,G.,Wohlleben,W., Lu,Y., Jiang,W.,
2013. Identification of two-component system AfsQ1/Q2 regulon and its
cross-regulation with GlnR in Streptomyces coelicolor. Mol. Microbiol. 87,
30e48.
Willey, J., Santamaria, R., Guijarro, J., Geistlich, M., Losick, R., 1991.
Extracellular complementation of a developmental mutation implicates
a small sporulation protein in aerial mycelium formation by S. coelicolor.
Cell 65, 641e650.
Willey, J., Schwedock, J., Losick, R., 1993. Multiple extracellular signals
govern the production of a morphogenetic protein involved in aerial
mycelium formation by Streptomyces coelicolor. Genes. Dev. 7, 895e903.





























