Embed Size (px)
Citation preview

Histopathology 1983, 7, 537-548
Possible liver cell differentiation in testicular germ cell tumours
GRETE KRAG JACOBSEN* & MARIANNE JACOB SENT Departments of Pathological Anatomy, *Herlev Hospital and
Hvidovre Hospital, University of Copenhagen, Denmurk
Accepted for publication 19 May 1982
JACOBSEN G. KRAC & JACOBSEN M. (1983) Histopathology 7, 537-548
Possible liver cell differentiation in testicular germ cell tumours
Germ cell tumours may imitate various structures of the developing embryo and foetus. Certain structures have, however, very rarely or never been observed in these tumours. Thus the presence of hepatic tissue in testicular germ cell tumours has not been reported. In a series of 37 non-seminomatous testicular tumours seven tumours contained epithelial structures showing morphological and func- tional resemblance to liver cell trabeculae. These structures were present in tumours with yolk sac turnour (YST) components and most of the tumours also contained teratoid elements. With the immunoperoxidase technique the epithelial structures were heavily stained for alpha-foetoprotein (AFP) and ferritin in all cases, while positive staining for albumin, prealbumin and transferrin was occasionally found. Alpha-I-antitrypsin and haernoglobin F were demonstrated in few scattered cells. Whether or not these epithelial structures should be included among the various patterns of YST or considered to be teratoid components is uncertain. It is suggested that examinations of the heterogeneity of the concomitant serologic AFP may support one or other assumption.
Keywords : testis, germ cell tumours, liver cells, yolk sac tumour, alpha-foeto- protein, ferritin
Introduction
During our studies of testicular germ cell tumours, we observed in some of the non-seininomatous tumours, epithelial structures that showed morphological resem- blance to liver cell trabeculae. Our attention was especially drawn to these epithelial structures in sections stained for alpha-foetoprotein (AFP) by the immunoperoxidase technique. Although the question whether or not patterns of testicular germ cell tuinours can be interpreted as liver or gut precursors has been raised before (Parkinson
Address for correspondence: G.Krag Jacobsen, Department of Pathological Anatomy, Hvidovre Hospital, DK-2650 Hvidovre, Denmark.
0309-0r67/83/0~00-0~37 $02.00 0 1983 Blackwell Scientific Publications 537

538 G.K.Jacobsen and M.Jacobsen
& Beilby 1980), hepatic tissue per se has not been reported to our knowledge. In the following paper we present seven non-seminomatous testicular germ cell tumours which contained epithelial structures with morphological resemblance to liver cell trabeculae. In addition the results of immunoperoxidase staining for a series of proteins that are present in normal foetal liver cells and/or in the foetal yolk sac are given.
Materials and methods
TISSUES
In a 4 year period, 68 orchidectomy specimens with germ cell tumours were received in the Department of Pathological Anatomy, Herlev Hospital.
The specimens were handled according to the recommendations of the Danish Testicular Cancer Project (DATECA Study Group 1978). Thus, a minimum of five blocks was available from each tumour. The tissues were fixed in 10% buffered formalin and processed as previously described (Jacobsen, Jacobsen & Clausen 1981). Sections were stained with haematoxylin and eosin (H & E), periodic acid- Schiff’s reagent (PAS) before and after digestion with diastase, Perls’ Prussian blue method for iron, p-dimethylaminobenzylidenerhodamine for copper, Gordon & Sweets’ method for reticulin and Hall’s method for bilirubin.
Haematoxylin and eosin stained sections were used for classification according to the WHO typing of testis tumours (Mostofi 1977) and to the concept of Teilum regarding the yolk sac tumour (YST) (Teilum 1976). Haematoxylin and eosin stained sections were also used for selection of tumour blocks for immunohistochemical investigations.
Tissue from three foetal livers obtained from abortions or ectopic pregnancies were fixed, processed and stained in the same way.
I M M U N O P E R O X I D A S E S T A I N I N G PROCEDURE
Serial sections cut at 5 pm were stained for alpha-foetoprotein (AEP),. prealbumin (PA), @bumin (ALB), alpha-I-antitrypsin (AlAT), ferritin (FER), transferrin (TR), immunoglobulin G (IgG), carcinoembryonic antigen (CEA) and haemoglobin F (HbF), using the indirect immunoperoxidase technique (Clausen & Thomsen I 978).
For control staining the specific antiserum was always replaced by the immuno- globulin fraction of serum from unimmunized rabbit as well as by phosphate- buffered saline (PBS). Furthermore, in the sections from foetal liver the specific antiserum was replaced by the appropriate absorbed antiserum, when available. All control stainings were negative. The specimens were counterstained with haematoxylin and mounted in Aquamount @.
ANTISERA
Specific rabbit antisera against human AFP, PA, ALB, AlAT, FER, TR, IgG and

Liver cells in testicular turnours 539
CEA were obtained from DAKO, Denmark, and against HbF from Behringwerke, Hoechst, Denmark. The specificity of the antisera was verified by performance testing on formalin-fixed and paraffin-embedded material with known positive reaction for the various antigens.
The working dilutions were found by titration. The following dilutions were used : anti-AFP 1/40 (lot. 109 C, protein concentration 9.8 g/l), anti-PA 1/40 (lot. 108 C, protein concentration I 1.6 g/l), anti-ALB 1/2000 (lot. 048 C, protein concentration 15.6 g/l), anti-AIAT 1/40 (lot. 029 C, protein concentration 10.6 g/l), anti-FER 1/150 (lot. 038 A, protein concentration 8.6 g/l), anti-TR 1/150 (lot. 118 E, protein concen- tration 10.6 g/l), anti-IgG 1/30 (lot. 020 D, protein concentration 10.2 g/l), anti- CEA xi100 (lot. 036, protein concentration 17.8 g/l) and anti-HbF 1/80(lot. 2456AM).
Peroxidase swine-antirabbit immunoglobulin (DAKO, lot. 090) was used in dilution 1/20 determined by chessboard titration. For control stainings the immuno- globulin fraction from unimmunized rabbit (DAKO, lot. 109 B, protein concentration 20.0 g/l) was used in dilution 1/50. For absorption human AFP-standard (foetal chord serum), crystallized ALB (Behringwerke, Hoechst, Denmark) and human liver- FER (provided by N. Strandberg Pedersen, Statens Seruminstitut, Denmark) were used.
MICROSCOP[C E V A L U A T I O N
The staining reactions were read as positive or negative. Only characteristic granular, pronounced red-brown staining in which there was no disagreement in evaluation was registered as positive.
Results
Thirty-seven of the 68 testicular germ cell tumours were non-seminomatous tumours of various types. In seven of these, epithelial structures showing resemblance to liver cell trabeculae were found. Table I shows the numbers of the various tumour types and the distribution of the various combinations of the mixed tumours. Yolk sac tumour (YST) components were present in all of the seven tumours. Eleven tumours of the remaining 30 non-seminomatous tumours contained YST components without the presence of structures resembling liver cell trabeculae.
M O R P H O L O G Y
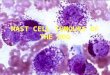
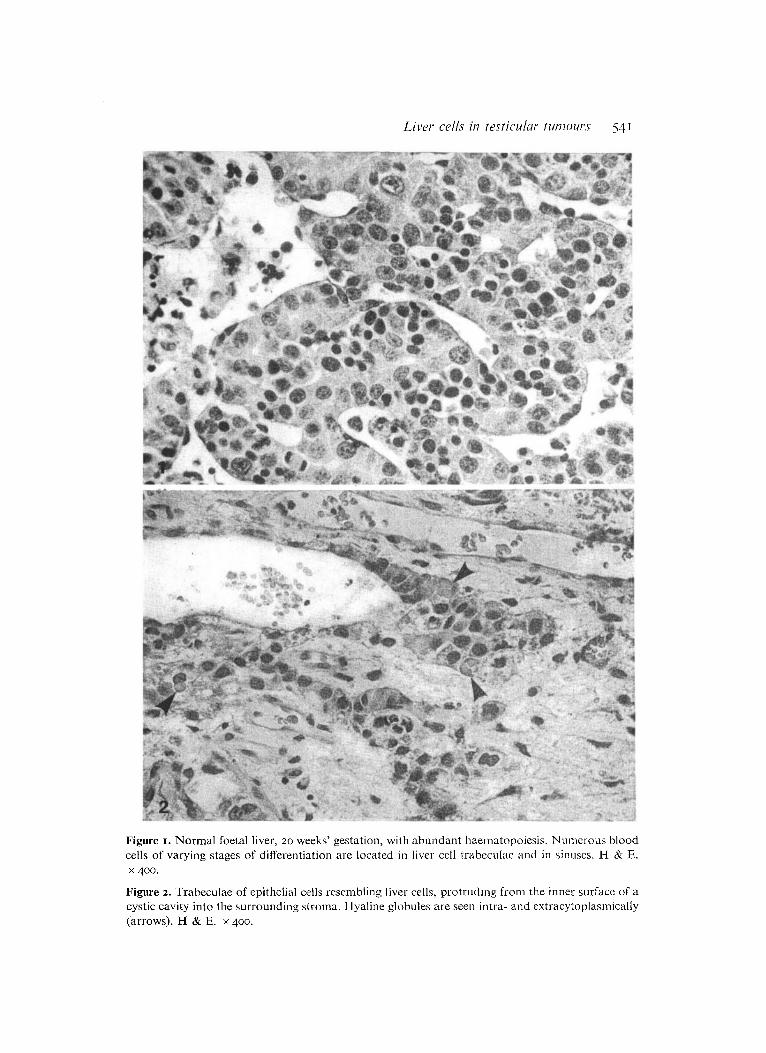
In seven tumours we observed trabeculae or irregular groups of epithelial cells resembling liver cell trabeculae (Figures 2 & 3). These structures were present in various numbers, unevenly distributed i n the tumour tissue. The trabeculae were usually two to four cells thick. The largest of the cell groups in the individual tumours comprised from 10 to 150 cells. The cells were characteristically polyhedral with distinct cell borders. The nuclei were fairly large with heavy nuclear membrane and conspicuous nucleoli. The cytoplasm was eosinophilic and finely granular. In some

540 G.K. Jacobsen and M . Jacobsen
Table I. Number of turnours and distribution of turnour components in seven non-semino- matous germ cell tumours with possible liver cell differentiation
No. of tumours Turnour type
I YST+T 2 EC+ YSTS T I 2
I
EC + YST + CC EC + YST + T + CC S + EC+ YST+ T + CC
Abbreviations : YST= yolk sac tumour. T =teratoma. EC =embryonal carcinoma. CC = choriocarcinoma. S =seminoma.
cells brown granular material or eosinophilic globules of various sizes were seen (Figures 2 & 3). Bile was not observed. Occasionally erythroblasts were present and numerous erythrocytes were often found in the vicinity.
The liver cell structures never occurred as tumour formation per se. They often appeared as isolated islands in a loose mesenchymal stroma with myxoid appearance or they protruded into the surrounding stroma from the inner surface of cystic cavities (Figure 2). I n the epithelial structures small acini or tubular formations sometimes occurred (Figures 5 & 8).
The epithelial structures were usually closely associated with Y ST elements. Various patterns of the YST were present. No single pattern was especially associated with the epithelial structures.
HISTOCHEMICAL S T A I N I N G
Some of the epithelial cells in the trabeculae also contained PAS-positive granules in the cytoplasm after digestion with diastase. Iron, 'copper, reticulin fibres and bile were not demonstrated in these structure. In the foetal livers, PAS-positive diastase resistant material was also present. In addition sparse iron granules were seen in a few cells, but copper was not found.
IMMUNOHISTOCHEMICAL S T A I N I N G
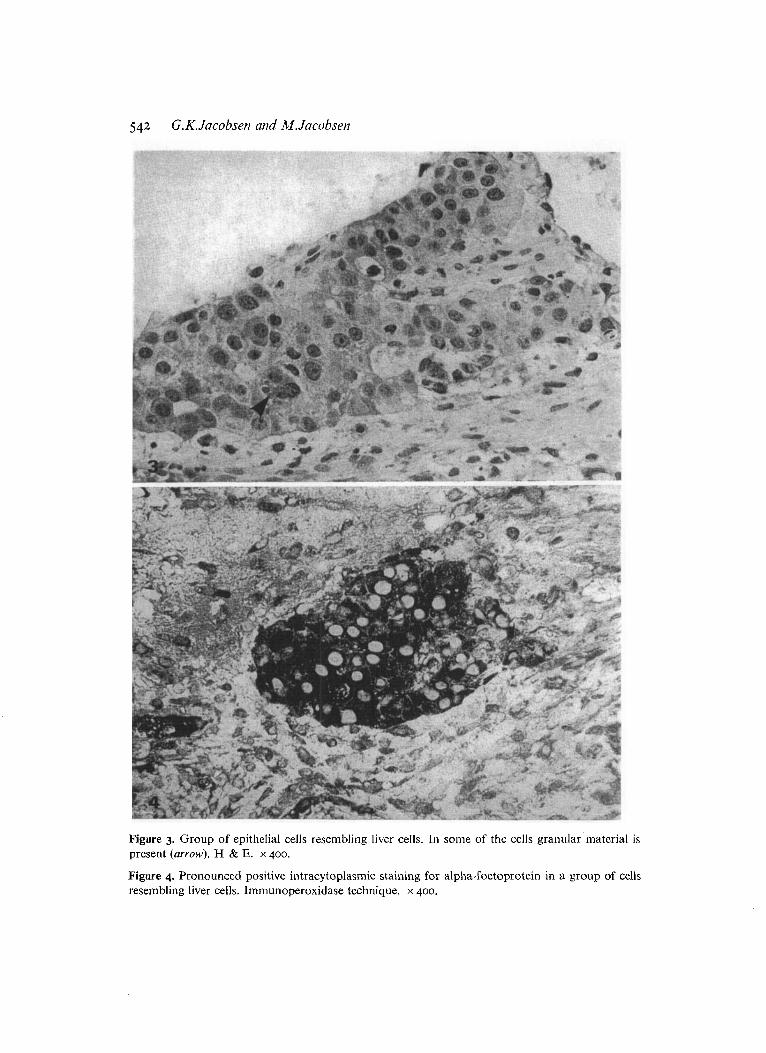
All the epithelial structures were heavily stained for AFP and FER (Figures 4 & 5) . In the majority of the cells a pronounced red-brown staining of the cytoplasin was found. Staining reactions for ALB, PA and T R were occasionally positive (Figures 6 & 7). This staining was usually less pronounced than the staining for AFP and FER
Liver cells ill testiculLlr turll0ur.r 54 1
Figure I. Normal foetal liver, 20 weeks’ gestation, with abundant haematopoiesis. Numerous blood cells of varying stages of differentiation are located in liver cell trabeculae and in sinuses. H & E. x 400.
Figure 2. Trabeculae of epithelial cells rcsenibling liver cells, protruding from the inner surface of a cystic cavity into the surrounding stroma. Hyaline globules are seen intra- and extracytoplasmically (arrows). H & E. x 400.
542 G.K..Jacobsen and M.Jacobsen
Figure 3. Group of epithelial cells resembling liver cells. In some of the cells granular material is present (arrow). H & E. x 400.
Figure 4. Pronounced positive intracytoplasmic staining for alpha-foetoprotein in a group of cells resembling liver cells. Immunoperoxidase technique. x 400.
Liver cells in testicular. tumours 543
Figure 5. A group of cells positively stained for ferritin. The histiocytes in the surrounding stronia are also positively stained. lmmunoperoxidase technique. x 400.
Figure 6. Positive staining reaction for prealbumin is seen in a few cells in a group of cells resembling liver cells. Immunoperoxidase technique. x 400.
.
544 G.K.Jacobsen and M.Jacobsen
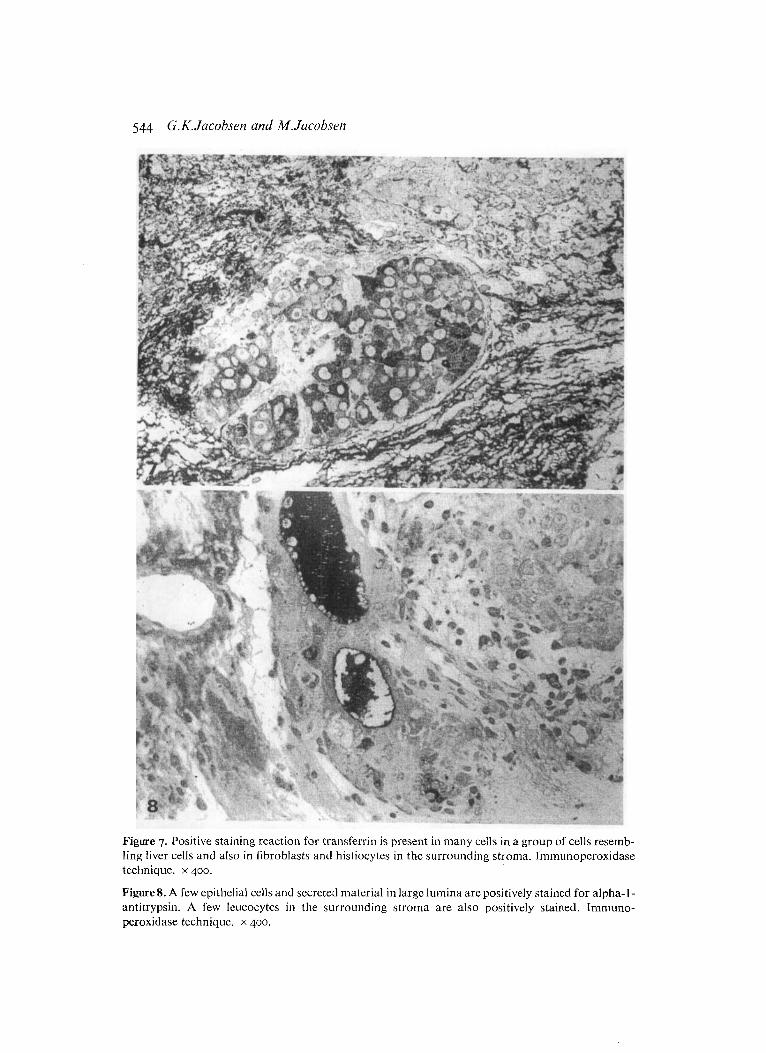
Figure 7. Positive staining reaction for transferrin is present in many cells in a group of cells resemb- ling liver cells and also in fibroblasts and histiocytes in the surrounding stroma. Inimunoperoxidase technique. x 400.
Figures. A few epithelial cells and secreted material in large lumina are positively stained for alpha-l- antitrypsin. A few leucocytes in the surrounding stroina are also positively stained. Immuno- peroxidase technique. x 400.

Liver cells in testiculav turnours 545
and appeared in fewer cells. AlAT was demonstrated in scattered cells and in the lumina of some of the tubular structures (Figure 8). HbF was shown i n the cytoplasm in a few of the epithelial cells and often in erythroblasts in tubular or vesicular lumina. No positive staining for CEA and IgG was found.
The various YST patterns were positively stained for AFP, AlAT, FER, TR, CEA and IgG.
In the hepatocytes of the foetal livers AFP, AlAT, ALB, FER and TR were demonstrated. In the haematopoietic tissue between the liver trabeculae positive staining reaction for HbF was found and the bile canaliculi were positively stained for CEA.
Discussion
The normal yolk sac during development undergoes phases of variegated appearance and function. These have been divided into periods of evolution, function and regression (Hesseldahl & Falck Larsen 1969). In the first period the yolk sac is a two-layered epithelial sac. In the second solid cords and tubules grow from the endodermal layer into the wall of the yolk sac in which also islands of haeinatopoietic cells and capillaries appear. The endodermal tubules are especially prominent in the sixth week of gestation. In the last period progressive degeneration takes place.
During folding of the embryo a part of the yolk sac endoderm is incorporated into the body cavity as the gut. In the third week the liver primordium or hepatic diverticulum appears as an outgrowth from the epithelium of the foregut (Langman 1975). Thus the epithelial cells of the liver and the endodermal cells of the yolk sac are closely related. This relationship is reflected morphologically as the liver trabeculae and the tubules of the yolk sac in the second period appear very similar at light micro- scopic level.
I t has been shown that AFP is synthesized in the visceral layer of the very early yolk sac in mice (Pziadek & Adamson 1978, Hogan & Taylor 1981). In man it has been shown A i t AFP as well as PA, ALB, AlAT and T R are synthesized in the secondary yolk sac (Gitlin & Perricelli 1970), and the synthesis of the same proteins has been found in liver tissue from an embryo of 32 days gestation (Gitlin & Biasucci 1969). In a previous immunohistochemical study we have demonstrated the presence of AFP, ALB, PA, AlAT, FER, IgG, TR and HbF in the endodermal cells of the inner surface of the yolk sac from the first period as well as in the endodermal cells and in the tubules in the yolk sac from the second period, while CEA was not demon- strated. We have suggested that IgG in the yolk sac might be absorbed from the maternal serum at least in the outer mesothelial layer (Jacobsen, Jacobsen & Henriksen 1981b). Albrechtsen, Wever & Wimberley (1980) have suggested that HbF is synthesized in the yolk sac endoderm. In the foetal liver no positive reaction was found for IgG and HbF, and the positive reaction for CEA was located in bile canaliculi only. The differences in the presence of HbF in yolk sac endoderm, in foetal liver cells and in the epithelial structures resembling liver cell trabeculae may be due to various stages of differentiation.

546 G.K. Jacobsen and M. Jacobsen
Although we have not been able to prove the specificity of all the used antisera by absorption with proper antigen our staining results as to foetal liver, yolk sac and yolk sac tumour components are in agreement with previous studies (Jacobsen et ul. 1981a, 1981bj. Thus in our opinion it is verified that in the epithelial structures resembling liver cell trabeculae in the germ cell tumours of this series, the same proteins as in foetal liver tissue and as in foetal yolk sac have been demonstrated. Since these structures thus show functional relationship and morphological resemb- lance to foetal liver as well as to the tubules of the normal yolk sac they may represent imitations of both.
Teilum (1976), who was the first to observe yolk sac differentiation in germ cell tumours, described eight different patterns of this tumour element which he desig- nated endodermal sinus tumour based on comparative studies with normal rat yolk sac. Epithelial structures resembling liver cell trabeculae as presented in our series are not included among these variants. The presence of YST elements in all the seven tumours with epithelial structures resembling liver cell trabeculae support the assump- tion that they are a variant of yolk sac differentiation. On the other hand all but one tumour contained teratoid elements and the epithelial structures might therefore also be considered an element among these. Recently endodermal sinus tumours of the ovary with a solid pattern resembling that of hepatocellular carcinoma have been described (Prat, Bhan & Dickersin 1980). In our series the epithelial structures resembling liver cell trabeculae never occurred as regular tumour formations. This discrepancy may be due to different histogenesis or it may be due to a variation in appearance of the same histogenetic related structures in the ovary compared to the testis. It should, however, be noticed that the epithelial structures in our series also showed resemblance to hepatocellular carcinoma, albeit of a highly differentiated type. Whether the epithelial structures resembling liver cell trabeculae had a malignant nature or not can only be proved by frank invasion or metastatic properties which we have not been able to demonstrate in our series.
Whether or not the epithelial structures which we have observed are variants of yolk sac differentiations or in fact liver trabeculae cannot be determined with certainty on the basis of the present study. Ultrastructural studies may add information favouring one or the other possibility. In their ultrastructural study of the human yolk sac, Hesseldahl & Falck Larsen (1969), however, stressed the ultrastructural similarities between yolk sac endodermal cells and liver cells.
Recently, it has been shown that the reactivity of AFP with the lectins concana- valin A (Con A) and lens culinaris agglutinin (LCA) varies according to the tissues in which the AFP is synthesized (Ruoslahti & Adamson 1978). Thus it has been shown that the Ieactivity pattern for germ cell tumours resembles that for foetal yolk sac, whereas the reactivity pattern for hepatocellular carcinomas resembles the pattern for foetal liver (Toftager-Larsen, Lund Petersen & N~rgaard-Pedersen 1981 j. Although the clinical significance of this heterogeneity of AFP remains to be clarified in larger series, the findings open up new possibilities. All the patients in our study had elevated serum values of AFP and studies are now in progress in order to correlate lectin binding properties of AFP with the various histological patterns of testicular germ cell tumours.

Liver cells in testicular tunzours 547
Acknowledgements
We are grateful to Susanne Smidt, Ulla Hansen and Aksel Ankerstjerne for skilful technical assistance and to Berit Alstrup for typing the manuscript.
References
ALBRECHTSEN R., WEWER U. & WIMBERLEY P.D. (1980) Immunohistochemical demonstration of a hitherto undescribed localization of hemoglobin A and F in endodermal cells of normal human yolk sac and endodermal sinus tumor. Acta pathologica et microbiologica scandinavica (Sect. A)
CLAUSEN P.P. & THOMSEN P. (1978) Demonstration of hepatitis B surface antigen in liver tissue. A comparative investigation of immunoperoxidase and orcein staining on identical sections of formalin fixed, paraffin embedded tissue. Acta pathologica et microbiologica scandinavica (Sect. A) 86, 383-388
DATECA STUDY GROUP (1978) The Danish Testicular Carcinoma Project. Acta pathologica et microbiologica scandinavica (Sect. A) 8, Suppl. 8, 147-151
DZIADEK M. & ADAMSON E. (1978) Localization and synthesis of alphafoetoprotein in post- implantation mouse embryos. Journal of Embryology and Experimental Morphology 43, 289-313
GITLIN D. & BIASUCCI A. (1969) Development of yG, yA, yM, P I C / / ~ I A , C’I esterase inhibitor, ceruloplasmin, transferrin, hemopexin, haptoglobin, fibrinogen, plasminogen, al-antitrypsin, orosomucoid, P-lipoprotein, az-macroglobulin and prealbumin in the human conceptus. Journal of Clinical Investigation 48, 1433-1446
GITLIN D. & PERRICELLI A. (1970) Synthesis of serum albumin, prealbumin, a-foetoprotein, al-antitrypsin and transferrin by the human yolk sac. Nature 228, 995-997
HESSELDAHL H. & FALCK LARSEN J. (1969) Ultrastructure of human yolk sac: endoderm, mesen- chyme, tubules and mesothelium. American Journal of Anatomy 126, 215-335
HOGAN B.L.M. & TAYLOR A. (1981) Cell interactions modulate embryonal carcinoma cell differen- tiation into parietal or visceral endoderm. Nature 291, 235-237
JACOBSEN G.KRAG, JACOBSEN M. & CLAUSEN P.P. (1981a) Distribution of tumor-associated antigens in the various histologic components of germ cell tumors of the testis. American Journal of Surgical Pathology 5, 257-266
JACOBSEN G.KRAG, JACOBSEN M. & HENRIKSEN O.B. (198rb) An immunohistochemical study of a series of plasma proteins in the early human conceptus. Oncodevelopmental Biology and Medicine 2, 399-410
LANGMAN J. (1975) Medical embryology. Human Development-Normal and Abnormal, 3rd edn. The Williams and Wilkins Company, Baltimore
MOSTOFI F.K. (I 977) International Histological Classification of Tumours, No. I 6. Histological Typing of Testis Tumours. WHO, Geneva
PARKINSON C. & BEILBY J.O.W. (1980) Testicular germ cell tumours. Should current classification be revised? Investigative & Cell Pathology 3, 135-140
PRAT J., BHAN A.K. & DICKERSIN S.J. (1980) Endodermal sinus tumour of ovary with solid pattern resembling that of hepatocellular carcinoma: an immunohistochemical and ultrastructural study. In abstracts, XIIth International Congress International Academy of Pathology Paris, 15-19 September
RUOSLAHTI E. & ADAMSON E. (1978) Alpha-fetoproteins produced by the yolk sac and the liver are glucosylated differently. Biochemical and Biophysical Research Communications 85, I 622-1 630
TEILUM G. (1976) Special tumours of ovary and testis and related extragonadal lesions. Comparative Pathology and Histological Identification, 2nd edn. Munksgaard, Copenhagen, and J.B. Lippincott Co., Philadelphia
88, 175-178

548 C.K. Jacobseii and M. Jacobseiz
TOFTAGER-LARSEN K., LUND PETERSEN P. & N0RGAARD-PEDERSEN B. (1981) Carbohydrate micro- heterogeneity of human alpha-fetoprotein. Oncodevelopmental aspects. In Lectins-Biology, Biochemistry, Clinical Biochemistry, Vol. I , pp. 283-292, ed. T.C.&og-Hansen. Walter de Gruyter, Berlin, New York