Embed Size (px)
Citation preview

Arch Virol (1996) 141:1623-1635
_Archives
Virology © Springer-Verlag 1996 Printed in Austria
Positive and negative host factors for Sendai virus transcription and their organ distribution in rat
T. Takagi ~'z'*, M. Iwama ~, K. S e t a 1'** , T. Kanda 3, T. Tsukamoto 1, S. Tominaga 2, and K. Mizumoto ~
i School of Pharmaceutical Sciences, Kitasato University, Tokyo, 2jichi Medical School, Tochigi,
3 National Institute of Health, Tokyo, Japan
Accepted May 7, 1996
S u m m a r y . In vitro mRNA synthesis by Sendai virus is almost entirely depend- ent on the addition of cellular proteins (positive host factors), one of which could be tubutin. In this study, we investigated the distribution of host factors in various rat organs. Extracts from the brain, thymus, heart, lung, testis, ovary, and uterus all supported in vitro Sendai virus transcription, among which the highest activity was obtained with the brain extract. On the other hand, little or no activity was detected in the liver, spleen, and kidney extracts. An inverse correlation between the apparent host factor activity to stimulate mRNA synthesis and RNase activity that hydrolyzes Sendai virus mRNAs was found, except in the liver extract. However, when a transcription initiation complex was isolated and subjected to RNA chain elongation reaction, all of the extracts including those from liver, spleen and kidney, were active. Immunoblotting showed that tubulin molecules were integrated in these initiation complexes, supporting the notion that tubulin is involved in the initiation complex forma- tion. We also identified a transcription inhibitory activity without any detectable RNase activity in the liver extract. This negative host factor seemed to act on RNA chain elongation. It is likely that Sendai virus transcription is regulated by both positive and negative regulatory factors.
Introduct ion
Sendai virus, a member of the Paramyxoviridae, contains a single, nonsegmented RNA genome of negative polarity which encodes six viral proteins, nucleo- capsid protein (NP), phosphoprotein (P), matrix protein (M), fusion protein (F),
Present addresses: * Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, MA, U.S.A., **National Defense Medical College, Saitama, Japan.

t624 T. Takagi et al.
hemagglutinin-neuraminidase glycoprotein (HN), and large protein (L). The NP, P, and L proteins are associated with the genome of 15.3 kilobases to form a nucleocapsid. The genetic information of this RNA genome is expressed through at least six monocistronic (one of them is functionally polycistronic) mRNA species by a virion-associated RNA-dependent RNA polymerase, which is thought to be composed of the P and L proteins [4].
Studies using an in vitro transcription system with purified virus particles have demonstrated that cellular proteins (host factors) are essential for Sendai virus mRNA synthesis, one of which could be tubulin [10, 13, 18]. A requirement for host factors has also been reported for other paramyxoviruses, including human parainfluenza virus type 3 (HPIV-3) [3, 11], mumps [20], measles [12, 15], and respiratory syncytial viruses (RSV) [2, 5, 6]. Therefore, it is important to elucidate the function of host factors to understand the molecular mechanism of transcription and replication of the paramyxovirus genome. We showed that tubulin molecules, which are essential for Sendal virus transcription, are integrated into the transcription initiation complex [-18].
Here, we studied host factor activity in the extracts from various rat organs and the results indicated that all of the organs examined contained host factor activity that facilitated viral mRNA synthesis, although at different levels. In addition, the presence of an inhibitory factor(s) was suggested. It is likely that Sendai virus mRNA synthesis is regulated by both accelerating and inhibiting host factors.
Materials and methods
Virus
Sendai virus strain Z [t6], propagated in the allantoic cavity of 10-day-old embryonated chicken eggs, was purified as described [17].
Preparation of extracts from rat organs
Sprague-Dawly rats (body weight 100-150 g) were decapitated, and the organs were removed immediately. All further operations proceeded at 0-4 °C. The organs were soaked in 2 volumes of a buffer solution containing 50raM Tris-HC1 (pH 7.9 at 20 °C)/100mM KC1/5mM MgClz/0.35M sucrose/5mM 2-mercaptoethanol, minced with scissors and homogenized with a motor-driven Potter-Elvejhem homogenizer (15-20 strokes). The homogenates were centrifuged at 13 000 x g for 30rain, then the supernatants were subse- quently centrifuged at 100000 x g for 120rain. The pellets were discarded, and the super- natants were dialyzed overnight against 200 volumes of 20raM Tris-HC1 (pH 7.9 at 20°C)/100mM KC1/lmM MgClz/0.1mM EDTA/2mM DTT/20% (v/v) glycerol. The dialysates were stored at -80 °C in small aliquots.
Preparation of extracts (S-l OO fraction) from Sendal virus-infected or unitfected HeLa cells
Extracts were prepared from 1.5 x 1 0 9 HeLa $3 cells spinner cultured in 3 L of Eagle's minimal essential medium (MEM) (Nissui Pharmaceutical Co., Ltd., Japan) supplemented with 5 % fetal calf serum (FCS) (Gibco) at 37 °C. Sendal virus infection was initiated at a m.o.i.

Positive and negative host factors for Sendai virus transcription 1625
of 30, and the extract was prepared at 16 h post-infection. Extraction was performed at 0-4°C. Cells harvested and washed with PBS ( - ) were resuspended in two packed-cell volumes of 10mM Tris-HC1 (pH 7.9 at 20 °C)/1.5 mM MgC12/10 mM KC1/0.5 mM DTT. After 20 min, the cells were homogenized by 15-20 strokes in a Dounce homogenizer with a "B" pestle. The lysates were centrifuged at 800 x g for 10min to remove nuclei and cell debris. The supernatants were combined and adjusted to 30mM Tris-HC1 (pH 7.9 at 20°C)/100mM KC1/lmM MgClz/0.5mM DTT/20% (v/v) glycerol. After removing the mitochondrial fraction by centrifugation at 12000 x g for 20min, the supernatants were centrifuged at 100000 x g for 120rain. The pellets were discarded, and the supernatants (S-100) were dialyzed overnight against 200 volumes of 20mM Tris-HC1 (pH 7.9 at 20°C)/100mM KC1/lmM MgC12/0.1mM EDTA/2mM DTT/20% (v/v) glycerol. The dialysates were centrifuged at 12000 x g for 40 rain to remove precipitates, and stored at - 8 0 °C in small aliquots.
In vitro RNA synthesis with purified Sendai virus particles
In vitro transcription with purified Sendai virus particles was carried out as described [10]. To assay RNase, 32p-labeled Sendai virus mRNAs were synthesized in vitro under the standard transcription conditions [10] with 30 gCi of [a_32p] UTP. After an incubation for 120rain at 30 °C, the :reaction mixtures were treated with proteinase K (200 ~tg/ml) in the presence of 0.5% SDS for 20min at 30°C. The RNA transcripts were extracted with phenol-chloroform (1:1), precipitated with ethanol, and dissolved in water.
Isolation of transcription initiation complex and RNA synthesis by the complex
The transcription initiation complex was isolated as described [20]. Purified Sendai virus particles (10 gg protein) were incubated for 10 rain at 30 °C with appropriate amounts of cell extracts in 65 gl, and the reaction mixture was centrifuged in 0.8ml ultraclear tubes (Beckman) containing a 200 gl 30% (v/v) glycerol/20 mM Tris-HC1 (pH 7.9 at 20 °C)/70 mM KC1/I0 mM MgC12/2 m M DTT/0.16% NP-40 on a 200 gl cushion of 50% (v/v) glycerol/ 40 mM Tris-HC1 (pH 7.9 at 20 °C)/70 mM KC1/10 mM MgC12/2 mM DTT at 100 000 x g for 120min at 4°C in a Beckman SW 55 Ti rotor with adaptors. The pelleted RNP was resuspended in 30 ~tl of the 50% glycerol cushion. The elongation reaction proceeded for 120 rain at 30 °C in 25 ~tl mixture [ 18], containing 50 gM [~-32P] UTP (5 000-10 000 cpm/pmol) and 4 ~tl of RNP fraction. The transcripts were analyzed as described [10, 18].
Immunoblottin9
SDS-PAGE was performed essentially as described [8], and protein bands were immuno- blotted as described [19], using a mouse monoclonal anti-squid optic lobe a-tubulin antibody (E3B8) [1].
Results
Stimulation of in vitro mRNA synthesis of Sendai virus by various organ extracts
In vitro t ranscr ipt ion of the Sendai virus genome almost entirely depends on hos t factors [10]. To s tudy the organ dis t r ibut ion of host factor activity, we p repared crude extracts f rom rat organs including the brain, thymus, heart, lung, liver, spleen, kidney, testis, ovary and uterus, and s tudied their abili ty to suppor t
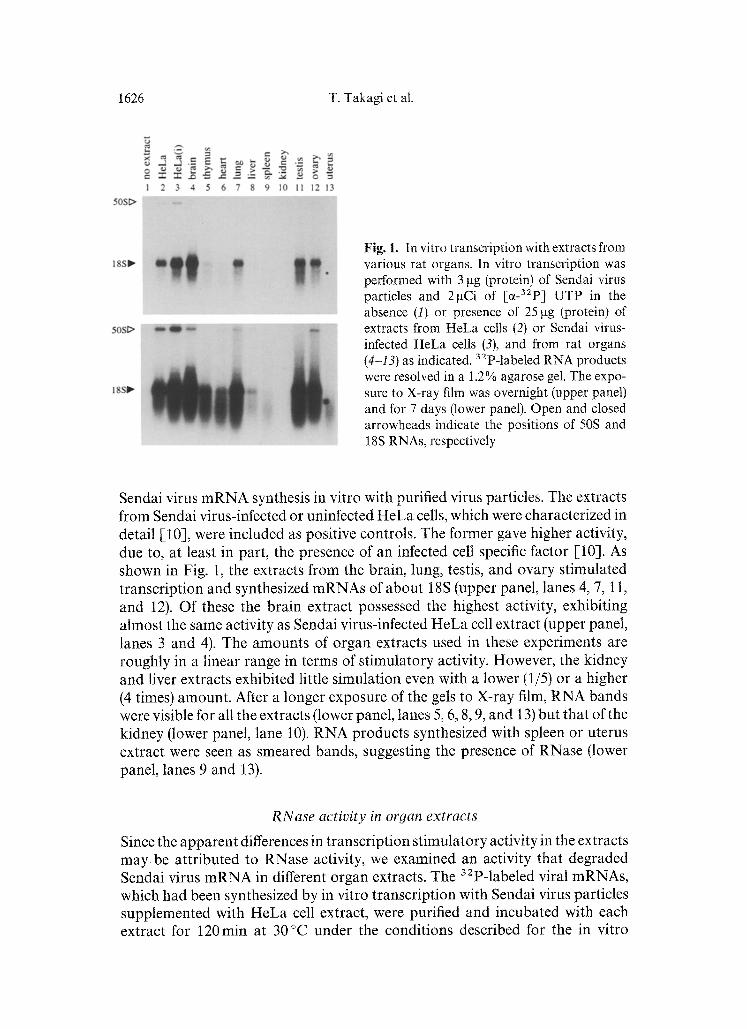
1626 T. Takagi et al.
Fig. 1. In vitro transcription with extracts from various rat organs. In vitro transcription was performed with 3 gg (protein) of Sendai virus particles and 2gCi of [a_32p] UTP in the absence (I) or presence of 25 gg (protein) of extracts from HeLa cells (2) or Sendai virus- infected HeLa cells (3), and from rat organs (4-13) as indicated. 32p-labeled RNA products were resolved in a 1.2% agarose gel. The expo- sure to X-ray film was overnight (upper panel) and for 7 days (lower panel). Open and closed arrowheads indicate the positions of 50S and 18S RNAs, respectively
Sendai virus mRNA synthesis in vitro with purified virus particles. The extracts from Sendai virus-infected or uninfected HeLa cells, which were characterized in detail [-10], were included as positive controls. The former gave higher activity, due to, at least in part, the presence of an infected cell specific factor [10]. As shown in Fig. 1, the extracts from the brain, lung, testis, and ovary stimulated transcription and synthesized mRNAs of about 18S (upper panel, lanes 4, 7, 11, and 12). Of these the brain extract possessed the highest activity, exhibiting almost the same activity as Sendai virus-infected HeLa cell extract (upper panel, lanes 3 and 4). The amounts of organ extracts used in these experiments are roughly in a linear range in terms of stimulatory activity. However, the kidney and liver extracts exhibited little simulation even with a lower (1/5) or a higher (4 times) amount. After a longer exposure of the gels to X-ray film, RNA bands were visible for all the extracts (lower panel, lanes 5, 6, 8, 9, and 13) but that of the kidney (lower panel, lane 10). RNA products synthesized with spleen or uterus extract were seen as smeared bands, suggesting the presence of RNase (lower panel, lanes 9 and 13).
RNase activity in organ extracts
Since the apparent differences in transcription stimulatory activity in the extracts may be attributed to RNase activity, we examined an activity that degraded Sendai virus mRNA in different organ extracts. The 32p-labeled viral mRNAs, which had been synthesized by in vitro transcription with Sendai virus particles supplemented with HeLa cell extract, were purified and incubated with each extract for 120min at 30°C under the conditions described for the in vitro
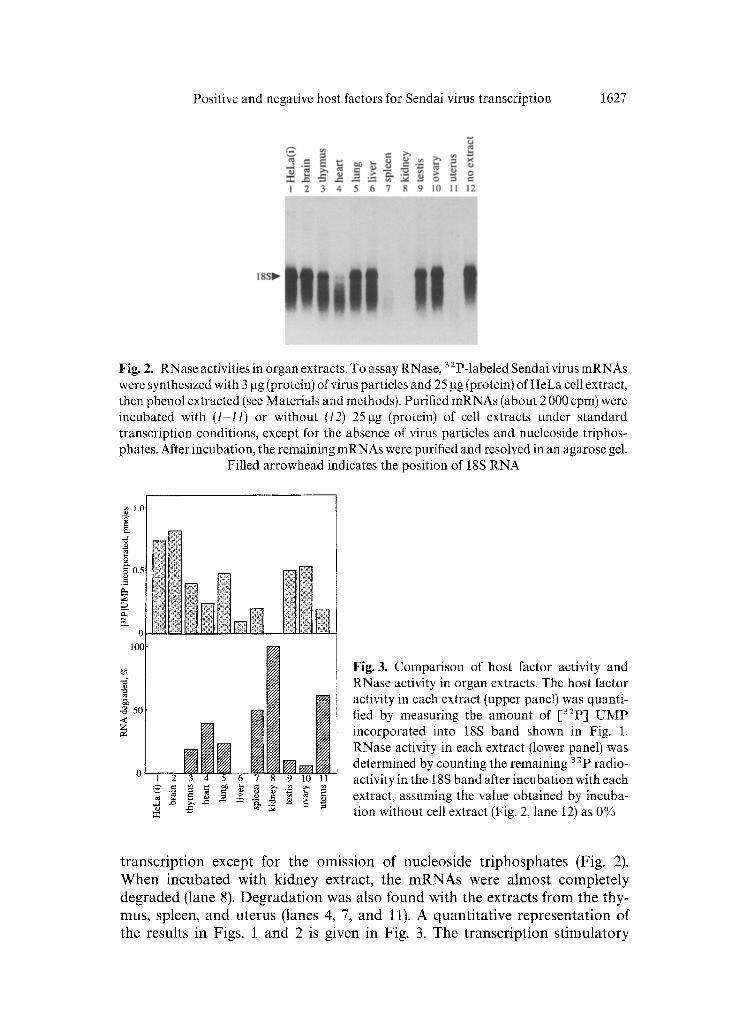
Positive and negative host factors for Sendai virus transcription 1627
Fig. 2. RNase activities in organ extracts. To assay RNase, 32p-labeled Sendai virus mRNAs were synthesized with 3 gg (protein) of virus particles and 25 gg (protein) of HeLa cell extract, then phenol extracted (see Materials and methods). Purified mRNAs (about 2 000 cpm) were incubated with (1-11) or without (t2) 25gg (protein) of cell extracts under standard transcription conditions, except for the absence of virus particles and nucleoside triphos- phates. After incubation, the remaining mRNAs were purified and resolved in an agarose gel.
Filled arrowhead indicates the position of 18S RNA
1.0
o ~ 0.5 .=_ o _
m 0
100
-~ 50 <
I ~ ! 1 i
0 1 2
v
3 4 5 6 7 8 9 10 11
Fig. 3. Comparison of host factor activity and RNase activity in organ extracts. The host factor activity in each extract (upper panel) was quanti- fied by measuring the amount of [32p] UMP incorporated into 18S band shown in Fig. 1. RNase activity in each extract (lower panel) was determined by counting the remaining 32p radio- activity in the 18S band after incubation with each extract, assuming the value obtained by incuba- tion without cell extract (Fig. 2, lane 12) as 0%
t ranscr ip t ion except for the omiss ion of nucleos ide t r iphospha tes (Fig. 2). W h e n incuba ted with k idney extract, the m R N A s were a lmos t comple te ly deg raded (lane 8). D e g r a d a t i o n was also found with the extracts f rom the thy- mus, spleen, and u te rus (lanes 4, 7, and 11). A quant i ta t ive represen ta t ion of the results in Figs. 1 and 2 is given in Fig. 3. The t ranscr ip t ion s t imula to ry

1628 T. TakaN et at.
activity and the mRNA-degrading activity was inversely correlated in most of the extracts. This finding suggested that the apparently lower host factor activities in the thymus, spleen, and uterus extracts are due to higher RNase activity. A notable exception was the liver extract, which did not show detect- able RNase activity, despite its nearly complete lack of stimulating ability (lane 6).
Isolation of transcription initiation complex
To minimize the deleterious effect of RNase activity and to estimate net transcription stimulatory activity, the transcription initiation complexes were prepared and subjected to the chain-elongation reaction [18]. Virus particles were incubated with each organ extract in the absence of 4NTPs, followed by ultracentrifugation to obtain viral RNPs (initiation complexes) as the pellets. Four NTPs were then added to the pellets to determine their RNA synthesizing (chain elongation) activity (Fig. 4A). Extracts from the liver, spleen, and kidney, all of which had exhibited almost no activity in the overall transcription (Fig. 1), supported the initiation complex formation, and generated RNA products (lanes 8 and 10). These results indicated that a considerable portion of the RNases and/or the inhibitory activity was eliminated during the isolation of initiation complexes, and that the positive host factors required at initiation as well as elongation are present in all of these extracts, although at different levels.
We showed that the transcription initiation complex formed with bovine brain extract contains tubulin, an essential factor required for Sendai virus transcription [18]. Therefore, initiation complexes isolated with various extracts were analyzed by immunoblotting with anti-tubulin antibody (Fig. 4B). When only virus particles (lane 2) or cell extracts (lanes 3 through 25, odd numbers) were incubated separately, tubulin was not detected in any pellet fraction obtained by uttracentrifugation. However, all the active complexes, which had been obtained after incubation of virus particles with each organ extract, contained tubulin (lanes 4 through 26, even numbers). The complexes formed with liver and kidney extracts apparently showed transcription activity, al- though to a lesser extent than other complexes (Fig. 4A, lanes 8 and 10), and they contained less tubulin than the others (Fig. 4B, lanes 16 and 20). These results confirmed the notion that cellular tubulin plays an important role in the initiation complex formation [18].
Presence of a transcription inhibitory activity
It appeared that the inability of the liver extract to stimulate transcription was not due to an RNase activity (Fig. 3). On the other hand, the liver extract did support the formation of an active initiation complex (Fig. 4). This suggested that there is an inhibitory component(s) in the liver extract. To examine this possibil- ity, we tested the effect of each extract on the standard transcription supported by
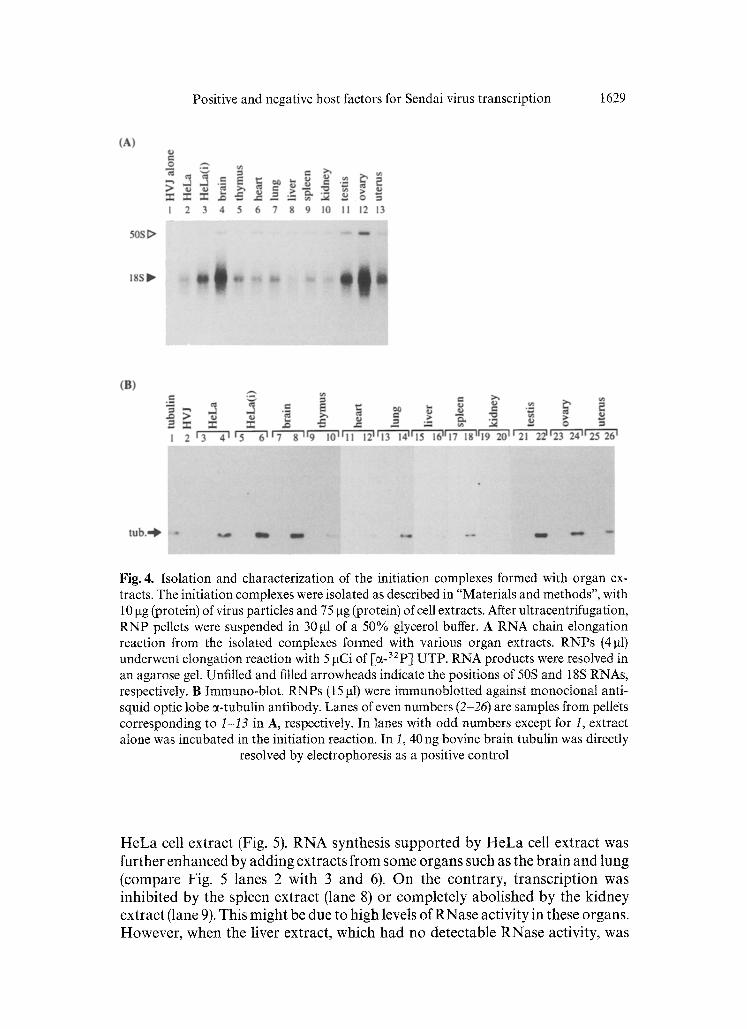
Positive and negative host factors for Sendai virus transcription 1629
Fig. 4. Isolation and characterization of the initiation complexes formed with organ ex- tracts. The initiation complexes were isolated as described in "Materials and methods", with 10 gg (protein) of virus particles and 75 gg (protein) of cell extracts. After ultracentrifugation, RNP pellets were suspended in 30 gl of a 50% glycerol buffer. A RNA chain elongation reaction from the isolated complexes formed with various organ extracts. RNPs (4 gt) underwent elongation reaction with 5 gCi of [~-32P l UTP. RNA products were resolved in an agarose gel. Unfilled and filled arrowheads indicate the positions of 50S and 18S RNAs, respectively. B Immuno-blot. RNPs (15 gl) were immunoblotted against monoclonal anti- squid optic lobe ~-tubutin antibody. Lanes of even numbers (2-26) are samples from pellets corresponding to t-13 in A, respectively. In lanes with odd numbers except for 1, extract alone was incubated in the initiation reaction. In I, 40 ng bovine brain tubulin was directly
resolved by electrophoresis as a positive control
H e L a cell extract (Fig. 5). R N A synthesis s u p p o r t e d by H e L a cell extract was fur ther enhanced by add ing extracts f rom some organs such as the brain and lung (compare Fig. 5 lanes 2 wi th 3 and 6). O n the cont rary , t r anscr ip t ion was inhibi ted by the spleen extract (lane 8) or comple te ly abol i shed by the k idney extract (lane 9). This migh t be due to h igh levels of RNase activity in these organs. However , when the liver extract, which had no detec table RNase activity, was
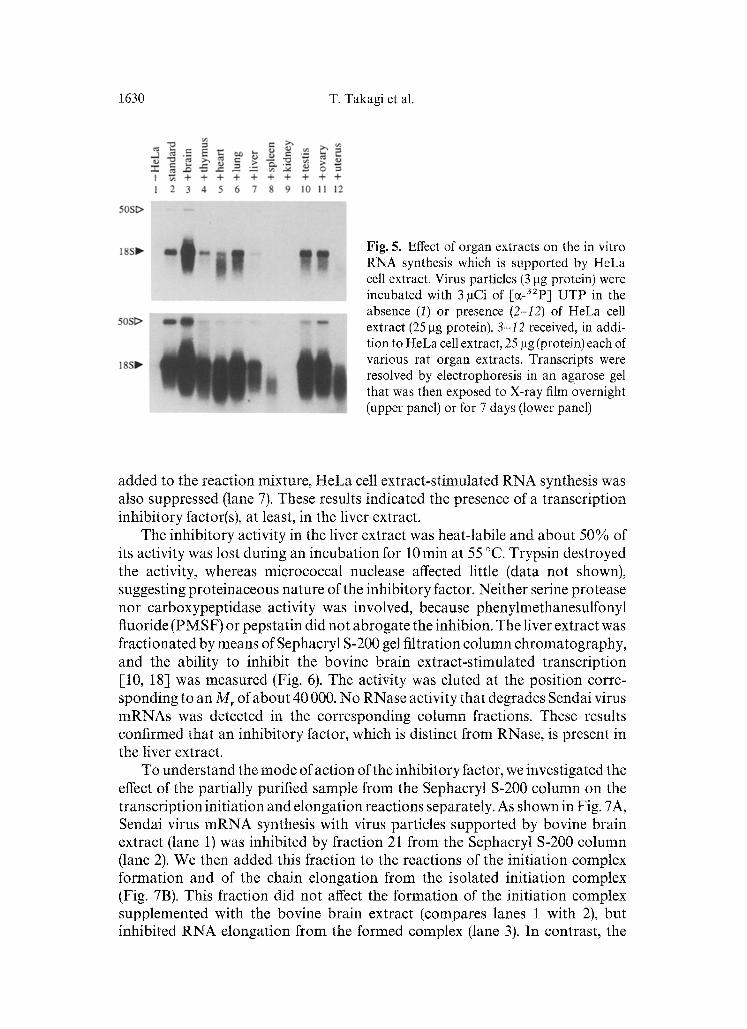
1630 T. Takagi et al.
Fig. 5. Effect of organ extracts on the in vitro RNA synthesis which is supported by HeLa cell extract. Virus particles (3 gg protein) were incubated with 3 laCi of [C~-32P] UTP in the absence (1) or presence (2-I2) of HeLa cell extract (25 l-tg protein). 3-12 received, in addi- tion to HeLa cell extract, 25 ~tg (protein) each of various rat organ extracts. Transcripts were resolved by electrophoresis in an agarose gel that was then exposed to X-ray film overnight (upper panel) or for 7 days (lower panel)
added to the reaction mixture, HeLa cell extract-stimulated RNA synthesis was also suppressed (lane 7). These results indicated the presence of a transcription inhibitory factor(s), at least, in the liver extract.
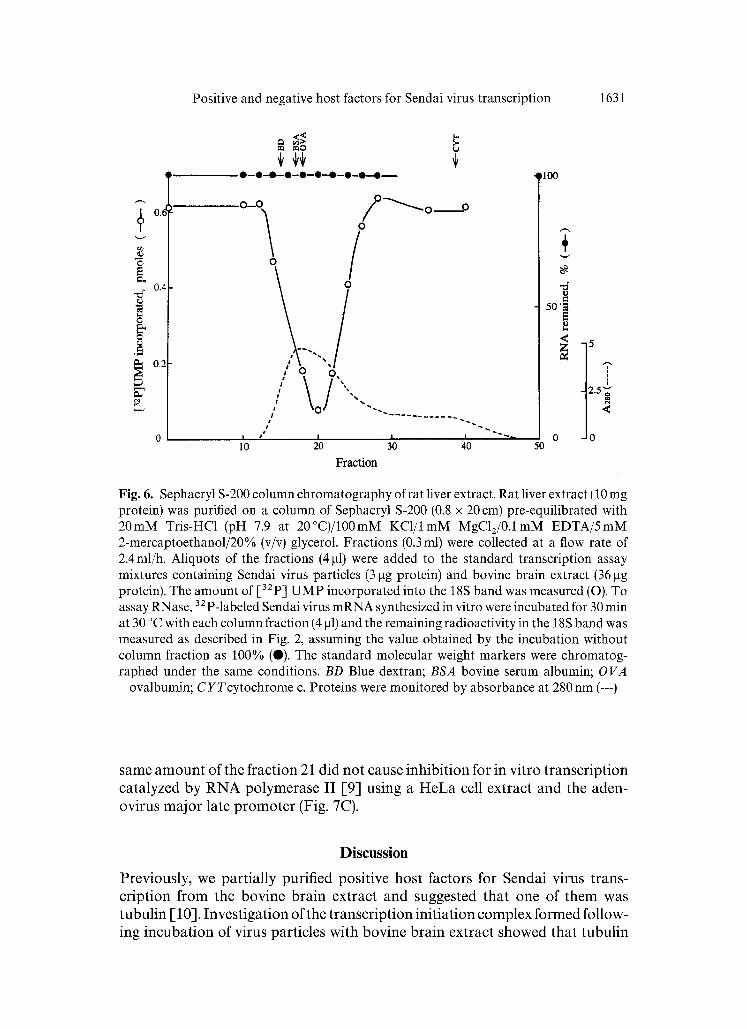
The inhibitory activity in the liver extract was heat-labile and about 50% of its activity was lost during an incubation for 10 rain at 55 °C. Trypsin destroyed the activity, whereas micrococcal nuclease affected little (data not shown), suggesting proteinaceous nature of the inhibitory factor. Neither serine protease nor carboxypeptidase activity was involved, because phenylmethanesulfonyl fluoride (PMSF) or pepstatin did not abrogate the inhibion. The liver extract was fractionated by means of Sephacryl S-200 gel filtration column chromatography, and the ability to inhibit the bovine brain extract-stimulated transcription [10, 18] was measured (Fig. 6). The activity was eluted at the position corre- sponding to an M r of about 40 000. No RNase activity that degrades Sendai virus mRNAs was detected in the corresponding column fractions. These results confirmed that an inhibitory factor, which is distinct from RNase, is present in the liver extract.
To understand the mode of action of the inhibitory factor, we investigated the effect of the partially purified sample from the Sephacryl S-200 column on the transcription initiation and elongation reactions separately. As shown in Fig. 7A, Sendai virus mRNA synthesis with virus particles supported by bovine brain extract (lane 1) was inhibited by fraction 21 from the Sephacryl S-200 column (lane 2). We then added this fraction to the reactions of the initiation complex formation and of the chain elongation from the isolated initiation complex (Fig. 7B). This fraction did not affect the formation of the initiation complex supplemented with the bovine brain extract (compares lanes 1 with 2), but inhibited RNA elongation from the formed complex (lane 3). In contrast, the
Positive and negative host factors for Sendai virus transcription 1631
0.q
O
E 0.4
0.2
<<
i,i, + O--Q--O--O--O--O--O--O-O--O~
o_k S o o /
i I I %
I sa I I I " ~ 10 20 30 40
Fraction
+
Q.I
50 '~
~100
",5 I
? i
-hs"~. <
0 - 0 50
Fig. 6. Sephacryl S-200 column chromatography of rat liver extract. Rat liver extract (10 mg protein) was purified on a column of Sephacryl S-200 (0.8 x 20 cm) pre-equilibrated with 20mM Tris-HC1 (pH 7.9 at 20°C)/100mM KC1/lmM MgC12/0.1mM EDTA/5mM 2-mercaptoethanol/20% (v/v) glycerol. Fractions (0.3 ml) were collected at a flow rate of 2.4ml/h. Aliquots of the fractions (4 gl) were added to the standard transcription assay mixtures containing Sendai virus particles (3 gg protein) and bovine brain extract (36 gg protein). The amount of [32p] UMP incorporated into the 18S band was measured (O). To assay RNase, 32p_labele d Sendai virus mRNA synthesized in vitro were incubated for 30 min at 30 °C with each column fraction (4 gl) and the remaining radioactivity in the 18S band was measured as described in Fig. 2, assuming the value obtained by the incubation without column fraction as 100% (0). The standard molecular weight markers were chromatog- raphed under the same conditions. BD Blue dextran; BSA bovine serum albumin; OVA
ovalbumin; CYTcytochrome c. Proteins were monitored by absorbance at 280 nm (---)
same a m o u n t of the fraction 21 did not cause inhibit ion for in vitro t ranscr ipt ion catalyzed by R N A polymerase II [9] using a H e L a cell extract and the aden- ovirus major late p r o m o t e r (Fig. 7C).
Discussion
Previously, we part ial ly purified positive host factors for Sendai virus trans- cription f rom the bovine brain extract and suggested that one of them was tubul in [-10]. Invest igat ion of the t ranscr ipt ion init iat ion complex fo rmed follow- ing incubat ion of virus particles with bovine brain extract showed that tubul in

1632 T. Takagi et al.
Fig. 7. Effect of partially purified transcription inhibitory factor on the initiation and elongation of Sendai virus transcription. A Overall transcription reaction. The standard transcription reaction was performed using 3 gg of virus particles, 3 ~tCi of [~_32p] UTP and 36 gg of bovine brain extract, in the presence (2) or absence (1) of 3 gl of fraction 21 from the Sephacryl S-200 column (Fig. 6). B Initiation complex formation and elongation reaction. The initiation reaction was performed as described in Materials and methods with some modifications. The first incubation was for 30min in 125 lat, with 30gg (protein) of virus particles and 360 fag (protein) of bovine brain extract in the absence (I, 3) or presence (2) of 30 lal of the fraction 21. After ultracentrifugation, RNP pellets were suspended in 40 gl of a 50% glycerol cushion buffer. RNPs (8 lal) underwent chain elongation with 5 laCi [~ 32p] UTP in the absence (1, 2) or presence (3) of 3 gl of fraction 21. C Effect of fraction 21 on transcription by RNA polymerase II. The in vitro transcription was performed essentially as described [9], with 1 lag of Sinai digested pSmaF DNA [-9] as the template and 40 lag (protein) of the HeLa cell extract, in the absence (I) or the presence (2) of 3 lat of fraction 21 from the Sephacryl S-200 column. The filled arrowhead (536 nt) indicates the run-off transcript from
the above template
was in tegra ted in to the complex [18]. The invo lvemen t of tubul in , which is ub iqu i tous in eukaryo t ic cells, in Sendai virus gene express ion was also suggested by the inh ib i t ion of R N A synthesis by a m o n o c l o n a l anti-13-tubulin an t ibody [13].
Here, we examined the o rgan d is t r ibu t ion of the posi t ive hos t factor activity us ing crude extracts of var ious ra t o rgans and showed tha t the a p p a r e n t difference in the activity a m o n g t h e m is chiefly due to the relative level of RNase activity (Fig. 3) except for the liver. F u r t h e r m o r e , all of the extracts examined s t imula ted the fo rma t ion of t ranscr ip t ion in i t ia t ion complexes and e longa ted chains f rom t h e m (Fig. 4A). Tash i ro et al. [21, 22] isolated the pro tease sensitive m u t a n t , F1-R, which has a m i n o acid subs t i tu t ions near the cleavage site of

Positive and negative host factors for Sendai virus transcription 1633
F protein and which causes a generalized infection in mice. Thus, the mutant virus can proliferate not only in cells in the respiratory tract, but also in other cell types, supporting our observation that the positive host factors are ubiquitous among organs. The presence of tubulin in all of the initiation complexes obtained (Fig. 4B) supports the notion that tubulin is involved in Sendai virus mRNA synthesis [10, 13, 18]. However, it should be noted that there were differences in the tubulin content among complexes (Fig. 4B). Since the amounts of tubulin in the liver and kidney extracts were found comparable to other organ extracts as measured by immunoblot analysis (data not shown), it seems unlikely that tubulin contents are insufficient for the active complex formation in these two extracts. We reported that a positive host factors activity, which is complementary to tubulin, is required for the overall reaction in addition to tubulin [10]. It is possible that the function of the other factor is to stimulate the initiation complex formation, and that its level is low in the liver or kidney.
To our knowledge, this is the first report of an activity in the cell extract (Figs. 5-7) that inhibits the transcription of paramyxovirus. Since the nega- tive host factor partially purified through a Sephacryl S-200 column did not show any inhibitory effect on the in vitro transcription by RNA polymerase II with the adenovirus major late promoter (Fig. 7C), it is unlikely that this inhibitory component generally affects RNA synthesis. It seems that the nega- tive factor in the liver extract inhibits specifically the step(s) after the initi- ation step, e.g., the chain elongation itself, or the recycling of the RNA polymerase and/or the positive host factor(s) (Fig. 7B). It is of considerable interest to purify and identify this inhibitory activity, and to reveal its mode of action.
It was reported that the organ tropism of Sendai virus is chiefly determined by the presence of a protease(s) on the cell surface of respiratory tract, which cleaves viral F protein at specific position [-7, 14, 21, 22]. The widespread distribution of host factor activities does not seem to account directly for the pneumotropic nature of Sendai virus. The results of this study, however, leave the possibility that both positive and negative factors contribute to the fine regulation of Sendai virus transcription after the entry of viral genome into the cell, which depends upon proteolytically activated state of F protein. The internal cellular environment, in other words, the availability or the abundance of these positive and negative host factors, should provide favor- able or restrictive conditions for viral proliferation. Further studies on these factors will be important for understanding the molecular basis underlying the regulation of replication of not only Sendal virus, but also of other para- myxoviruses.
Acknowledgement This study was supported by Grants-in-Aid from the Ministry of Education, Science, and Culture of Japan.

1634 T. Takagi et al.
References
1. Arai T, Matsumoto G (1988) Subcellular localization of functionally differentiated microtubules in squid neurons: regional distribution of microtubule-associated proteins and ]3-tubulin isotypes. J Neurochem 51:1 825-1 838
2. Barik S (1992) Transcription of human respiratory syncytial virus genome RNA in vitro: requirement of cellular factor(s). J Virol 66:6 813-6 818
3. De BP, Galinski MS, Banerjee AK (1990) Characterization of an in vitro system for the synthesis of mRNA from human parainfluenza virus type 3. J Virol 64:1 135-1 142
4. Galinski MS, Wechsler SL (1991) The molecular biology of the Paramyxovirus genus, In: Kingsbury DW (ed) The paramyxoviruses. Plenum Press, New York, pp 41-82
5. Herman RC (1989) Synthesis of respiratory syncytial virus RNA in cell-free extracts. J Gen Virol 70:755-761
6. Huang YT, Romito RR, De BP, Banerjee AK (1993) Characterization of the in vitro system for the synthesis of mRNA from human respiratory syncytial virus. Virology 193: 862-867
7. Ishida N, Homma M (t978) Sendai virus. Adv Virus Res 23:349-383 8. Laemmli UK (1971) Cleavage of structural proteins during the assembly of the head of
bacterophage T4. Nature 227:680-685 9. Manley JL, Fire A, Cano A, Sharp PA, Gefler ML (1980) DNA-dependent transcription
of adenovirus genes in a soluble whole cell extract. Proc Natl Acad Sci USA 83: 5 405-5 409
10. Mizumoto K, Muroya K, Takagi T, Omata-Yamada T, Shibuta H, Iwasaki K (1995) Protein factors required for in vitro transcription of Sendal virus genome. J Biochem 117: 527-534
11. Moscona A, Peluso RW (1991) Properties of human parainfluenza virus type 3 RNA polymerase/replicase activity in vitro: consensus with other negative-stranded RNA viruses. J Virol 65:4470-4474
12. Moyer SA, Baker SC, Horikami SM (1990) Host cell proteins required for measles virus production. J Gen Virol 71:775-783
13. Moyer SA, Baker SC, Lessard JL (1986) Tubulin: a factor necessary for the synthesis of both Sendai virus and vesicular stomatitis virus RNAs. Proc Natl Acad Sci USA 83: 5 405-5 409
14. Nagai Y (1993) Protease-dependent virus tropism and pathogenicity. Trends Microbiol t: 81-87
15. Ray J, Fujinami RS (1987) Characterization of in vitro transcription and transcriptional products of measles virus. J Virol 61:3 381-3 387
16. Shibuta H, Akami M, Matumoto M (1971) Plaque formation by Sendai virus of parainfluenza group, type 1 on monkey, calf kidney and chick embryo cell monolayers. Jpn J Microbiol 15:175-183
17. Shibuta H, Kanda T, Adachi A, Yogo Y (1979) Characterization of bovine parainfluenza virus type 3. Microbiol Immunol 23:617-628
18. Takagi T, Muroya K, Iwama M, Shioda T, Tsukamoto T, Mizumoto K (1995) In vitro mRNA synthesis of Sendai virus: isolation and characterization of transcription initi- ation complex. J Biochem 118:390-396
19. Takagi T, Yanagisawa K, Tsukamoto T, Tetsuka T, Nagata S, Tominaga S (1993) Identification of the product of the murine ST2 gene. Biochem Biophys Acta 117: 194-200
20. Takeuchi K, Tanabayashi K, Okazaki K, Hishiyama M, Yamada A (1993) In vitro transcription and replication of the mumps virus genome. Arch Virol 128:177-183

Positive and negative host factors for Sendai virus transcription 1635
21. Tashiro M, Pritzer E, Khoshnan MA, Yamakawa M, Kuroda K, Ktenk H-D, Rott R, Seto JT (1988) Characterization of a pantropic variant of Sendai virus derived from a host-range mutant. Virology 165:577-583
22. Tashiro M, Yamakawa M, Tobita K, Klenk H-D, Rott R, Seto JT (1990) Organ tropism of Sendai virus in mice: proteolytic activation of the fusion glycoprotein in mouse organs and budding site at the bronchial epithelium. J Virol 64:3 627-3 634
Authors' address: Dr. K. Mizumoto, School of Pharmaceutical Sciences, Kitasato University, 5-9-1 Shirokane, Minato-ku, Tokyo 108, Japan.
Received March 12, 1996




























