Embed Size (px)
Citation preview

Population dynamics of Hydropsychidae
(Insecta ; Trichoptera)
in the N’Zi River (Ivory Coast),
a temporary stream partly treated
with the insecticide Chlorphoxim ~1
Bernhard STATZNER (2)
111 the temporary river N’Zi -an uffluent of the Bandama (Ivory Coast)- the population dynamics of four species of hydropsychid caddis flics, Cheumatopsyche spp., and Aethaloptera dispar, were folloured from January 1977 until March 1978. Prom June 1977 until December 1977 parts of the N’Zi were treated rvith fhe insecticide Chlorphoxim (organophosphate compound) in order to evaluate the effects of such treatment on Simulium damno- sum-lhe vecfor of human onchocerciasis in Wesf Africa-and on the non-large1 fauna.
The hydropsychid larvae and pupae were obtained by using a Surber sampler, a tore sampler, and from quali- tative samples, yielding a tolal of about 17,000 specimens. These tuere identified 10 the instar level.
Densities ut the surface of the subsiraium as me11 as in fhe hyporheic habitat appear 10 be influenced by the dischar!e pattern. Decreasing discharge and consequently a reduction of areas with sufficient currenb produce increaszng larval and pupal densities per surface unit. It is suggested that zones with sufficient current, mhich are controlled by the morphology of the rapid in relation to a certain discharge and fo some exteni by the densify of luron1 tubes of A. dispar, deiermine the level of intra- and interspecific aggression betrveen the Cheumatopsyche spp. and thereby leacls 10 changes in relative abundances of the species and probably their instars.
Tt’hile A. dispar is no1 able to survive the periods mithout flow in ils aquatic stage, Cheumatopsyc.he spp. cari,, probably in the remaining stagnant pools. The instar disiribution as lue11 as the changes in the mean lveighf of the Cheumatopsyche spp. srrggesi thai a period of increased reproduciion occurs after reswmhg flolv and ut the end of the spate of the main rainy season. Undoubtedly the larval life of fhe species skrdied here lasis longer than is generally assumed for insects of tropical rivers, and probably only trvo or three generations per year occur.
The resulis are discussed in relation to the secomlary producfivity of African streams. Since a11 four species are seriously affected by fhe chlorphoxim treatment, their disappearance is briefly discussed
in relation to the consequences for the ecosystem, and especially fo the decrease in the predator pressure 011
Simulium damnosum.
KEY WORDS : Africa - Running Water - Benthos - Ecology - Development - Pesticides - Ivory Coast.
RÉSUMÉ
DYNAMIQUE DES POPULATIONS D’?krYDROPSYCHIDAE (INSECTA ; TRICHOPTERA) DANS LE N’ZI (CÔTE D’IVOIRE), RrvrÈnE TEMPORAIRE PARTIELLEMENT TRAITÉE A L’INSECTICIDE CHL~R~HOXIM
La dynamique des populations de quatre Hydropsychidae (Trichoptera), trois Cheumatopsyc.he spp. et Aet,haloptera dispar, a étk étudiée de janvier 1977 à mars 1,978 ~III* le N’Zi, affluant du Bandama (C;üfe d’ivoire), COUPS d’eau temporaire. De juin 7977 & dtkembre 1978 certaines parties clu N’Zi furent fruitées avec un insecticide
(1) The field studies and parts of the laboratory studies were carried out durin g a period of World Hcalth Organization consnltantship in the Onchocerciasis Control Programme (O.C.P.).
(2) Zooloqisches Insfilut I der Unioersifüt (T.H.), Postfach 6350, Kornblumensir. 13, D - 7500 Karlsruhe 1, PR Germnny.
Reo. Hfydrobiol. trop. 15 (2): 157-176 (1982).

1t,rj B. STATZNER
(Chlorphosim, composé orqanophosphoré) dans le but d’étudier les effets sur la fume cible et non cible, de tels épan- dages destinés ci détruire iimulium damnosum, vecteur de l’onchocercose humaine en Afrique de l’ouest.
Dix-sepf mille spécimens d’Hydropsychidae, déterminés au niveau du stade larvaire furent récoltés par pros- peefions qualiiatives et quantitatives à l’échanfillonneur de Surber et au caroitier.
Les densifés tanf hyporhéiques qu’à la surface du substrat varient en fonction du dkbif. Un débit décroissanf lié Cr une diminuiion des zones présentanf un courant suffisant augmente les densités larvaires rf nymphales par unité de surface. Il est vraisemblable que le niveau d’agression infer- et intraspècifique entre les Cheumatopsyche spp. modifie les abondances relnfives des espèce s et probablement des sfades larvaires. Il est dt!terminé par l’esistence de zones présentant un conranl suffisant, ces zones étant elles-mêmes, sur chaque rapide, fonction de la morphologie du cours I d’eau, du dt?bit et de la densité des tubes larvaires d’A. dispar.
Conkrairement à A. dispar, les Cheumatopsyche sont capables de survivre & l’ftat larvaire pendant la période d’Ptiage, probablement dans les mares stagnanfes qui subxistenf. Les réparfitions par stades et les changements de poids moyen suggèrent qu’une période de reproducfion accrue survien! aprés le retour de 1’Pcoulement ainsi qu’à la fin de la période de crue de la principale saison des pluies. La durèe du développemeni larvaire des espèces étudiées ici est plus longue que celle généralement avancée pour les insectes des rivières tropicales et il n’y a probablemenf que &uæ ou trois générafions par an. Ce résultat esb discuté dans le cadre de la producbinn secondaire des fleuves africains.
Les quaire. espéces étudiées ici éfanf sérieusemenf affectées par les traitements au chlorphoxim, leur disparition est bridvement discutée dans l’opfique de ses conséquences sur l’éçosystème et en particulier de la diminution de pression prEdatrice sur Simulium damnosum.
h!crrs-cLÉs : Afrirple - Eaux Courantes - Benthos - Écologie - Développement~ - Pesticides - COte d’ivoire.
1. INTRODUCTION
Seasnnal trop& or c.onst-ant trop&? Considering benthic invertebrates of permanent st,reams near the ecjuator in Africa, their populations are obviously flur.Luat-ing (BOTTGER, 1975; CRIDLAND, 1958; Ko- PELKE, 1981; LEHMANN, 1979; MALAISSE and RIPERT, unpubl., cit. ex ibfALAIo§E, 1974; RAMZANAN- KASINA, 1973; STATZNER, 1976; ZWICK, 19%).
8uc.h fluct.uations in the benthic invertebrate fauna bave becn suggested and confined also for teniporary or nearly temporary streams in African savannah areas (CRIS, 1956; HARRISON, 1966; ,J. D. HYNEY, 1975a,b; LE BERRE,~~~~;PETR, 1970). However, probably due to taxonomie. difficulties of suah studios, det.aded investigations on the popu- lation dynarniw of sinEle species are few, and, in thrsr African areas, to -the best, of my knowledge, restricted to t 1112 Simulirzm damnosum complex (LE BERRE, 1966j, the vector of humnn oncho- eerciasis in West. Africa.
The Lrmporary river N’Zi (Ivory Coast) was chosen for a large-scale field invest,igation int,o the efl’ects ou the aquatic target and non-target fauna of regular applicat.ions of the insecticide chlorphoxim, directeci against Simulium dnmnosum.
Ii was hoped that observations on the rec.overy of the fauna afi-er larviciding as compared with data from the untreatecl çontrol section of the N’Zi would giva indical-ions about the life cycles of the
four abundant hydropsychid species studied in this paper as well as t-heir contribution to secondary production. The usc of insecticides in suc.h kinds of studies was introduced by H. B. N. HYNES and WILLIAMS (1962).
This paper also considers the effect of t.he inse& icide on the four hydropsychid species it. deals wit,h. C)t,ber invert,ebrat,e species from different, streams in the Ivory Coast will be studied in papers short.ly to be published hi; the hydrobiological team of the O.R.S.T.O.M. in Bouaké (Ivory Coast), with which 1 was associat.ed during a period as consultant of the W.H.O.
2. METHODS
Five sampling places were visited once a mont11 between January 1977 and March 1978. At low and medium discharges qualitative samples were taken from a11 subst,rata colonized by the four hrdropsyc.hid species. This was carried out accordmg t,o the approximate relative abundance of the hydraulic conditions (combination of the fac,tors: velocity, subst,rate Lype, wet.er depth) on a rapid, a method w1lic.h is thought t.o be necessary in studies on life cycles of running wat,er species (STATZNER, 1981a). Threa additional samples (June 1977) came from Butter experiments, a method described in detail hy DEJOUX (1975) and TROURAT (1981). The samples

HYDROPSYCHIDAE IN THE NfZI
were brushed into a bucket, the content of the bucket then was sieved through a 200 pm me&.
At one rapid (Ouokoukro, see 3.) Surber samples were taken from a grave1 and pebble substratum. The sampler used (see fig. 3 in LÉVEQUE et al., 1977) was almost square sampling an area of 16 X 17 cm, with the mean mesh size of 320 ~200 prn. Since the net was made from curt.ain mat.erial, variations in the meshes reached up to 600 x2.80 pm.
At the same rapid where Surber samples were taken, t,he colonization of the hyporheic habitat (gravelipebble substratum) was investigated through one year; at other rapids this aspect was studied only occasionally. This was done by using a tore sampler (for details see STATZNER, 1979). During the dry season in 1975, a hole was dug in the dry part of the rapid until the subterranean water appeared. The wat,er was then sieved through a 200 p.m mesh.
In the field the material was conserved by using ca. SO y/0 alcohol, and lateron sort,ed out in the laboratory with the aid of stereomicroscopes. Instar separat.ion and dry weight det,erminat,ion were c.arried out as described in QTATZNER (1979).
3. AREA STUT>IED -THE N’ZI RIVER
3.1. General description, chlorphoxim application, geomorphology
The N’Zi River in the Ivory Coasl is a tributary of the Bandama River (fig. 1). Similarly to most of the large rivers in that count,ry, the N’Zi flows from the nort!h to the south. In the rainy season the N’Zi originates south of Ferkéssédougou. It enters the Bandama north of Tiassalé, thus extending between 60 and 100 North.
Among the numerous rapids, five rapids accessible by car were chosen for a regular survey. Since the eit’ect of a regular insecticide application on the non- target fauna of that river was the primary aim of the investigation, out of five stations, three were situated in the treat,ed area: Tinbé, Kolomikro, and Bocanda (fig. 1).
Tinbé was the only St#ation situated in the opera- Gona area of the Onchocerciasis Control Programme (O.C.P.) at that time, thus it was treated before with Temephos (Abate), the operational larvicide used during the first phase of t.he area treatment since 1974. Abate applications stopped on the 1st Decem- ber, 1976.
The application of Chlorphoxim, t.he larvicide t.ested during the period of this investigation, started on 14th of June 1977 at Tinbé and on 28th of June 1977 at Kolomikro and Bocanda.
Reu. Hydrobiol. frop. 78 (2): 167-176 (1953).
0 ,-.-..5!? "?'J km
FIG. 1. - The awa studicd with the monthly sampled rapids of the N’Zi River (indicatrd by a hlnck triangle) considered
in t.his study. Dnshed: area of larvicide npplicat.ion
The N’Zi and, occasionally, some affluent,s of the N’Zi were treated wit.h Chlorphoxim sprayed from a helicopter once a week. During the dry season each rapid was t,reated separately, during the rainy season (when the insec.ticide carries better) the insecticide was applied in general every 20 km. Normally 0.025 ppm per 10 minutes of Chlorphoxim was applied, but sometime overdosage occurred, especially in the north during low discharge conditions. The last Chlorphoxim application was on the 14th December, 1977.
The middle section of t.he studied stretch of the N’Zi was chosen as control and it was not treated with larvicide. Here two sampling stations were established at Fetekro and Ouokoukro (fig. 1). Fetekro was treated previously with Abate (from Oct. 1976 until 1st Dec. 1976). At Ouokoukro, an unknown poison was probably applied between Nov. and Dec. 1977 (local fishermen use this method sometimes for fishing).

The investigated stretch of the N’Zi exhibits only a weak slope (L)E MEKONA, 1981). The five rapids’ characteristics are:
Tinbt?: a natural rapid; mostly smooth roc.ks and s ,ones. t
Fetekro: an artificial rapid, remains of an old bridge; mostly large stunes with a smooth surface.
Ouokoukro: a natural rapid; mostly graveljpebble, few rocks with a smooth surface.
Kolomikro: a natural rapid; mostly rocks and large stones with an uneven surface.
R»canda: a natural rapid; mostly rocks, large st.ones with an uneven surface; gravel/pebble.
The main dif’ference between these five rapids was in t.he c,ross-section (fig. 2), which, in periods of diminishing discharge resulted in a different speed of the decrease of the area of habitats with current. For esample in Kolomikro the surface area and Lhe number of habitats in the current decreased more rapitily with a reduced discharge, than in Bocanda, where a smell decrease in discharge resulted in a more gradua1 reductiun of such habitats, until t,he whole rapid dried out except. a small v-shaped channel.
The part of t.he N’Zi investigated flows t,hrough savarrnah. The banks of the N’Zi, covered by a gallcry forest, were mostly undist.urbed, except for small areas of cult,ivation. There were only two regions in which large plantations were situated, viz. aronnd M’Bahiakro (rice) and around Kolomikro (coi.ton).
3.2. Physical and chemical properties
Data on the hydrology of t.he N’Zi came from different sources. The hydrological team of O.R.S.- T.O.M. monitored four water gauges in the experi- mental area of the N’Zi on a more or less regular basis. Where necessary, the records of O.R.S.T.O.M. were complernented with data from ILTIS (pers. ~~J~~lIll~lIl~~~~~o~) and O.C.P. (measurements hy helicopter pilote). Where data were available, with the curves from CAMUS (1971) and preliminary curves for Tinb6 the discharge was calculated for four point,s (fig. 3). It should be not.ed that the Bocanda gauge was sit.uated about 10 km downstream from tfie princxipa1 sampling place.
The discharge pat,tern was similar at a11 stations. ai, the beginninp of 1977 discharge decreased to zero first in the north and later also in the south. The river start-ed flowing again in the last, week of April, reaching a peak in June, and dropping afterwards. At thc end of August tbe main rainy season start#ed. The annual maximum of discharge was recorded in September and CMober. With the beginning of the dry season at tbr Pnd of October discharge decreased
\-_i \;i;“i
Bocanda u FIG. 2. - Scheme of the cross section of thc five monthly
s:unplrd rapids
again and in the nort-hrrn station reac.hed zero in February 1978.
The 16 years’ discharge data for Lhe N’Zi (CAMUS, 1971) dernonst,rate a high variation. The mont.hly means of these 16 years (fig. 3) differ us a lot. of the single years from t-he pattern of the study period by
t,he lack of the decrease in discharge from June to August.. Considering t.hese means, disrharge steadily increased from Marc.h until the rainy season. Cessation of slow was not observed during Camus’ investigations in Fetekro, M’Bahiakro, and Roc,anda. Some’cimes the discharge decreased t,o a few litres/sec., the minimum reported WBY 1 litre/sec in February 1957 in Bocanda. From the hydrological dal.a «ne cari conclude that the N’Zi was running over the whole stretch only in the rainy season. As the disc.harge decreased the river became a series of interconnected pools divided by rapids. These rapids cari be characterized as short, lake outflows and inflows. After the river stopped flowing the pools remained. This had an effect on water temperature and t,ransparency (fig. 3).
Wat,er temperature showed a relatively large range at the bepinning of the year (low discharge) as compared to t.he narrow range in the rainy season (maximum disc.harge).
Transparenc.y (measured by a Secchi disc) in- c.reased with decreasing discharge and vice versa. At Bocanda, where the transparency was measured most freyuently, it. was signifieantly related t.o the discharge by : f.ransparency (cm)=45.7-14.4 log discharge (m3/sec] (n=ll; r=-0,65).
Although rela tionships between disc,harge and the transport of particulat,e matter are complex (&IAN-
GELSDORF and SCHEURMANN, 19811), the above formula implies that transparency was mainly influenced by soi1 wash-in and sedimentation.

HYDROPSYCHIDAE IN THE N'ZI 161
rn. (10s
cm 70
50
30
10
3/ I(r
sec I+I)I
discharge
ml M’Bahiakto: mean 1954-70
77 114177 117177 IllO/ w7a date
. . Tinbé -,-._ Fetekro ____ M’Bahiak’ro - Bocanda
diurnal range of water temperature in the current x
Tinbéi 2<
Ouokoukro>l<
transparency in the current
A4 : 9
B0cand0i
mean of ‘ail rapids - studied
F M month
FIG. 3. - Discharge, transparency, and wa trr lemperaruro of t.he N’Zi Riwr
Planktonic. primary production, which in the pools is expected to be highest, at lowest discharge, did hardly affect, the transparency. Renthic primary production was related to transparency, since ayuatic macrophyt.es covered large st.ones and rocks only in periods of high transparency.
values measured for samples c.ollect.ed from running water only, are ranging as follows:
CO,H- : 23.8-35.6 mg/1 ; Ca++ : 2.245 mg/1 ; Mg++ : Ni +
1 - 1.9 mg/l; K+ : 2 -2.8 mg/1 ; : 1.2- 5.8 mg/1 ;
pH-values, obtained by ILTIS (pers. communi- Conduct,ivit.y: 37-51 qhos/cm. cation) are available for Tmbé and FeLekro over one The N’Zi is essentially a typical white water river, year and range between 7.0 and 7.5. due to the character of soil, vegetation and rainfall
Chemical analyses of the N’Zi at Tinbé were (SIOLI 1965, 1975). Its chemistry lies within the presented by the hydrobiological team of O.R.S.- range of many ot.her West African rivers such a list,ed T.O.i\I. in O.R.S.T.O.M.-O.M.S. (1976). The annual by VISSER and VILLENEUVE (I.F)75).
Rerr. Hydrobiol. irop. 15 (2): 157-116 (1982).

lii2 II. STATZNER
TABLE 1
Ht:acl c*aps~& midth (pm; in brackets: standard daviation) RIld dry weight (mg; cf. test for deterrnination) of instars of the species st.udied
Instar Larva 1 Larva 2 Larva 3 Lai-va 4 Larva 5 Pupa
Cheumtzfopsycke diyifafa !, “::::::“’ :E”i;7’ 278 (Y) a5 (16) 660 ($4) 1005 (40)
. . 0.0079 0.0379 0.193 0.917 1.261
Chezzmafopsyche sp. III 1 ng: l
1: : : 1: 1: F:il:* 220
(11) 283
0.0094 0.0233 (II)) 403 (14) 590 (17) 0.08.35 0.329 0.522
Ckeumafopsyche sp. VII pl. . . . . 231 (8) 344 (10) 553 (12) SO3 (23)
~lefhalopfera dispar l [un. . . . 238
(16) 342 (19) 4-M (20) 769 (36) 1253 (60)
mg. 2.594 . . . . . . . 0.0053 0.0182 0.0456 0.287 1.507
l The quotient headcapsule length: width of the larva one is 1.03 in Ckezzmafopsycke SP. III and 1.13 in Ckeumafopsycke sp. VII.
4. SPECIES STLJDIED
Hydropsychid larvae are usually regarded as a dificult. group t.o identify, concerning especially tropical rnaterial and individual instars. The charac+ f..eriqt.icp useful for the det.erminat,ion of species are briefly described by STAT~NER (1987 b).
One of tbe four species investigated is Aebhaloptera dislwr Brauer. This species has the code number T32 in an O.R.S.T.O.M.-calatogue on ayuatic inverte- bratee fmm the Ivory Coast (under publication).The other three speçies, which are briefly described in STATZNER (1981 h), are placed in the genus Cheum.afo- pyche Wallengren (sensu KIMMINS 1963). One is C. diyiftrfu (Mosely) (O.R.S.T.O.M.-catalogue: TlO). Thc seco11c1 one, CRertmatopsyche sp. VIZ, is expected to be a new species (near C. copiosa Kimmins) (O.R.S.T.O.bi.-catalogue: T5) and the third, Clzeczrna- topsyche “11. III (O.R.S.T.O.M.-catalogue: Tl) pro- hahly is c. f’ulcifiirn (Ulnier). .A definite determinat)ion of I~hezzn~crfopsyche sp. III and VII cannot yet. be given, since the knowledge about these and several othrr hytlropsyc.hid species obtained from the IvorJ; Coast indicat,es, that, correlations between d$, !& and larvae given in the literature are oft,en incorrect.. For the width of t,he head capsule see Tab. 1. The dry weight. of each instar was calculated from formulae obt.ained by regression analyses. The formulae are (x=headcapsule width in mm):
C:. digifafu : mg = 0.0~3. in = 3 ; r = 0.986). Ckezzmtzfopsycke sp. III : mg = 2.2x3.0 (n = 3 ; r = Cl.!)%). A. tlispar : mg = 0.7x3.' (n = 2j.
Pupal weight,s are from direct determinations.
5. RENTHIC AND HYPORHEIC ABUNDANCES
The abundanre of t.he benthic. fauna per area of the substrate increased in periods with decreasing
Hezr. Hydrcrfriol. trop. 15 (2): I.iT-176 (1982).
discharge and vice versa. This was best demonstrated by the results from Surber samples at Ouokoukro (fig. 4). This trenti was shown by the benthic. fauna as a whole as well as by Chironomidae and Tricho- ptera. The Ephemeroptera, nearly a11 Caenidae and Baetidae, did net. follow the same pattern at the beginning of 1977 nor 1978, when abundanc.e de.- creased.
This decrease is assumed to be related t,o the very high abundance of Cheumafopsyche spp. in t,he same periods (fig. 4). Cheumafopsyche is a known predator of Ephemeroptera (H. E. N. HYNES and WILLIAMS, 1962) and it is the most common predator of t,he N’Zi in unt.reated rapids. It is not yet, clear what caused the low abundance of Cher~rnatopsyche in August. 1977 in spite of a very low discharge at that time. It. was probably t.he absence of A. dispar, which was missing aft.er the period in which this rapid was dry. Larvae of A.. dispar construct a tube similar to t.hat of Nacronema (SATTLER, 1963, 1968), with the inflow opening protruding into the c,urrent. Between these tubes Cheumatopsyche spp. c.onstruct t.heir living tubes and their nets (fig. 5), oft,en several larval chambers on t.op of one anot,her. It is thought t.hat the tubes of A. dispar ameliorat,e the habitat for Cheumafopsyche by alt.ering the microcurrents above t,he substratum as well as the substratum itself.
The abundance of hyclropsychids in the hyporheic habitat, which was checked at Ouokoukro from February 1977 until January 1978, showed variations similar to the abundances at the surface. They were highest in February 1977, when many larvae were found in the çore sample. Less hyporheic larvae were found in August 1977 and January 1975, when the surface densities were also higher. In the other mont.hs no Hydropsychidae were found in the tore samples. Tab. II demonst.rat,es that. the hyporheic abundances reac.hed the level of the surface abun-
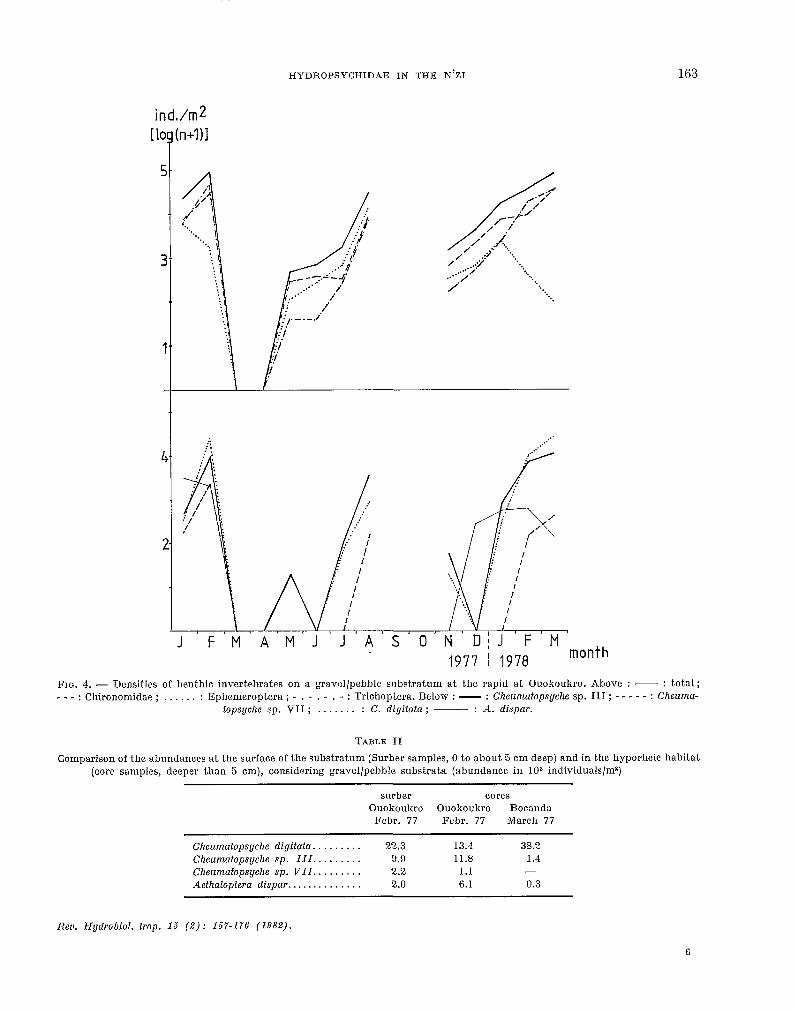
HYDROPSYCHIDAE IN THE N’ZI 163
ind./m2 n+l)l
,H C’ /
///
.y /-.Y-
y/’ .A... / / . ‘... .,/ ,. /.:/’ ‘X., / .% ‘...
Xv
FIG. 4. - Donsitics of benthic invertebrates on a gravcl/pebble substratum at tho rapid at Ouokoukro. Above : - : t.otal; - - - : Chironomidae ; . . . . . : Ephemeroptera ; - . - . - . - : Trichoptera. Below : - : Cheumatopsyche sp. III ; - - - - - : Cheuma-
topsyche sp. VII ; . . . . . . : C. digifata; - : A. dispar.
TABLE II
Comparison of the abundances at the surface of the substratum (Surber samples, 0 t.o about 5 cm deep) and in t.he hyporheic habitat (tore samples, deeper than 5 cm), considering gravel/pebble substrata (abundnnce in 10y individuals/ms)
surber cores Ouokoukro Ouokoukro Bocanda
Febr. 77 Fobr. 77 March 77
Cheumatopsyche digitata. ........ 22.3 13.4 38.2 Cheumatopsyche sp. III ......... 9.9 11.8 1.4 Cheumatopsyche sp. VII ......... 2.2 1.1 - Aethaloptera dispar .............. 2.0 6.1 il.3
Rerr. Hydrobiol. trop. 15 (2): 157-170 (1982).

dances. In t,lie tore most larvae wrre found in the upper 5-15 cm section, few were found in the section 15-25 cm and they rarely occurred in deeper layers.
Core samples from the dry rapid as well as t.he filtration of a large volume of hyporheîc wat,er obtained from a hole yielded no hydropsychid larvae. A single Ckwrnatopsyche larva was captured in a cor? sample from sand in a stagnant pool by Tinbé in a period without flow.
The relative abundance of the various species was approximately equal in surface and tore samples (Tab. II). The relative abundances in surface samples are shown in fig. 6 for a11 five rapids. This figure demonstrat.es, that A. rlisp~~r was not able to sur- vive the period without flow and therefore was probably restricted to the southern part of the N’Zi,
100
L Y 2 50
2
2 100
7 Y s 50
0
J FMAMJ J ASONOJ FM month 1971 19 78
q Cheumatopsyche digitata
El Cheumatopsyche sp. III
n Cheumatopsyche sp VI’
Aethaloptera dispar
FIG. 6. - Relative abundances of t.he four Hydropsychidae on the five rapids studied (Fetekro and Ouokoukro : control stations). The numbers above ihe columns indicate the number of specimens identifled. Dry indicates, that there was no flow on the rapid, ns. indicates, that no samples could be taken,
duo to the spate
where an inlerruption of flow is assumed to be much less frequent than in the north. During Chlorphoxim treatment it was not able to recolonize the southern part of the N’Zi, but it did occur at Ouokoukro after the flood from September ancl October. This would infer that females are able to fly a distance of about 80 km. probably from the N’Zi south of Bocanda or

HYDROPSYCHIDAE IN THE N’ZI 165
1911
Jan. Febr. March M~Y
4 ; 10 ; lb , n-203 f nz0
1
t n=3b
I
Jfb lb
1 Ii=15 I
f : I * :.. , :::.
t 15
, n=ZlO
i
dry
lb n=O
dry
: 3 1'
I ë . 1 , :: 8 H
15 t
11 1 0 0 1 9 I 1 10 13 11 1 9 nz0 Il.128 ! I
12135 n=30 1 n=73 ' ' / 10 f n-1 ; 1 n=2
t FI
, w77 i dl1 1 n=1409 Z
l n. 8. ' I I I
m L I I I L I I l c
dry ; , I I
I I I ! ë
I 1 ! f : . 1.. 15 ilO ; 7 12 ! 14 f 9
June JUIY Aus.
1978
%Dt NOV. Dec. Jan
2 13 20 ' f 11 f 17 t 12 1~707 ix470 n=931 n.277 nn=341', Il=37
-324 I n=455 ; n:325 1 na
n ! I ! samples
Febr
i;l’i A ~~~ 0 / 0 : 13 17
n=5 I w21 ! n=55 f w353 in:
March
9 n=O
2: s c
dry ë z
' nz94 n=79
f
ti
I
1 14 ' 9 1 n=535 tn=31[
I 1 I I I
II 12345~ 12345~ 12345~ 12345~
n=158 1 7 i IO ; ' 9 Il5 t n=24 ; wl7 1 n-0 t n=2
in= ' n:O : n=8 ;
n fn
n=139 2 Ln
n 1’ i. : f . . ; I I ' .i.i i
?6 27- 11 ! 15 ;n=321 nz393n f 6 f 11 ! ! 16 f 14 f ',n=83 f n=63 1 n=576 Ln I 1 ,
1 ?=b ; n=349 f ! n=O 1 I
- ,n.s. n=O f l 5
I - I I I 1 l -
1
c I i I I I
1 I I I ë 1
r 1 1 l I I I G I a 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ instar
FIG. 7. -- Cheumatopsyche digitnfa. Percentage instar-distribution at the five rapids. Application of Chlorphoxim is indicated by the blaclr horizontal bar for each treated rapid. Dat.e and number of specimens identified are given for each sample. Blaclr portion of the pupae-column indicates prepupae. If samples contained only few specimens, they are represenkd dircctly by points. Dot.t.ed parts of the columns at Tinbé (Febr. 1078) indicate the proportion of specimens, which mere dead (sample from stagnant water)
from the Comoe, which is the nearest permanent river to that place. After the end of the t.reatment this species appeared again at Bocanda and therefore the failure to recolonize the southern part might be expected to have been the result of larvicide appli- cation.
TO a certain extent the n-values in fig. 6 reflect the difflcu1t.y in finding enough larvae during periods of medium discharge, when the material was yielded by plunging. Furthermore they indicate, that the three Chezzmatopsyche spp. were very abundant on a11 rapids before the start of Chlorphoxim treatment. They were doubtless the most important inverte- brate group in respect of biomass. Cessat.ion of flow seemed to have only minor influence on this genus. After the beginning of larvicide application no serious reduction was observed at Tinbé. At. Kolo- mikro and Bocanda this genus was reduced heavily, only a few specimens surviving the treatment. This situation last,ed in Kolomikro until August, but in Bocanda an earlier recovery from July to August was
Reu. Hytlrobiol. irop. 15 (8): 157-176 (1982).
observed. After the flood, in November and De- cember, Cheamntopsyche u-as afl’ected SO seriously that it was either completely absent. in t,he samples or only a few early instars were found. On the control rapids Cheumatop.syche occurred at a11 times in “normal” abundance with one exception: In Decem- ber 1977 Cheumatop.syche were reduced to some extent at Ouokoukro. This as well as the condition of the whole benthic fauna clearly indicated the application of an unknown poison to this rapid.
From January to March 1978, Cheumatopsyche were well represented in t,he samples from a11 rapids. The change in relative abundance of these four species is relevant to what is discussed above. Evidently A. dispar had an important. position at a11 three southern st.ations of the N’Zi, in January and February 1977. In the period from November 1977 t.o February 1978 it reached a similar position again only at Ouokoukro, t.he control st,ation.
The change of relative abundance of the single Chezzmatopsyche species, which showed an increase

16l; R. STLTZNER
of C. tfi~gifafu at a11 st.at.ions wit.h decreasing discharge after tlir tlootl during Yepternber and October and a decreasr in the period from the start of flow up to the flood CanIHJt br due to chlorphoxim treatment.. Only t.he higher relative abundance of C. tligifnfn at, the t-rrated rnpids compared t.o the control rapids in the bcginning of 1978 cari probably br rxplaint4 l’y the IYrrvious Iarviciding.
Thr reasons which Icd te t-hese changes in the relative abundanccs of I:heurnafopsgcha will be dis- cusstL(I in connrct.ion with the life cycle dat.a below.
‘l’his chapter deals with the dynamics of the MOLLI-
lation structure of thr four species, with respect. to t he inst,ar clist.ribut.ion and the mean weight. Data from two cor? snmple,, s which yielded a considerable numbcr of hydropsychids, are included. Based on tlicse rcault-s the nature and the causes of the life cyçlrs of fhe four hydropsychids are disc.ussed at. thr rnti of this chapter.
6.1. Instar distribution for surface samples
~Vhen the morithly sample was not too fragment- ary, the instar distribution of any one species of a11 five rapids was conipared by ils-tests (nixn-tables). Thrse tests always indicate highly signiftcant tlitfererrces in thr group of thr tive rapids. For reasons for t.his sec later..
(:. tli.yituttr (fig. 7) h~i~rrie older as tlie rapids were (Irying up. Except at Pet-ekro, pupae were most riumercms imniediat~elp after the river start-cd tlowing in May, there was a distinct. increase of yoiinger inst,ars nt t.he otller four stations until the beginning of June. -A-\t Fetekro t.his inrrease of jur-eniles did net take place until July. Ai; t.he two control st.ations, older inst,ars increased from July t.» Xugust. Aft,er decrease in t Ile discharge in Novem- ber C. tligifclfcr was rare. In December a few specimens were found at, Fetekro but only two were c.aptured at C~uokoukro. The very low numbers at Ouokoukro were probably thr rcsult. of poisoning of this rapid as mentinntd alreatly above. At. Fetekro, older instars clominat.ed unt-il Alarch and the instar composition equallrd t bat; of t.lie prerious year at. the same hydroIcr#al period. At Ouokoukro, the situation \\an diffcrent. t’r~~n thr year before and t,he young and eq~ecially Ii& instar larvae were frequent even in hlarc.11. -4t. the t.reated rapids things developed differently. Tn Tinbe the first. Chlorphosim appli- cation did net- change the genrral dominante pattern.
However, in July older stages were missing and youngers grew only slowly up to August.. September data are net. comparable witb the rest as they were obtained near the bank in shallow water. In Novem- ber and Dec.ember, only Young larvae, almost a11 first instar, were present.. However, in January the instar cluantities were similar to those of the control stat.ions. In February, a11 spec.imens were sampled from a very small stagnant. pool remaining on the rapid and in which several specimens were alive and reac t.ed “normülly”. In this sarnple, fiftli instar formed almost 75 O.’ . :o (Jf the pc~pUlaticln.
In Kolomikro and Bocanda, the applicat,ion of Chlorphoxiru immediately drastieally reduced t,he population, leading to the presence of only old instars. In Boeanda, a11 instars were again present. in August, whilst no rec.olonization occurred at Kolornikro. Up to the end of the application t,he species remained rare but., from January to March, the population reached a level sirnilar to t.he year before.
61.2. ~hertInclfof,sflcht~ sp. 111
This species (fig. 8) appeared in the samples us well as C. digifafn with few of the youngest instars at. the beginning of the invest,igat.ion. At Tinbe, Fetekro, and Bocanda only few or no spec.imens of C:heun2utopsgche sp, TII were found in hlay. Until June, the number of young instars increased except at Fetekro where t.his ocçurred until July. Older inst,ars increased U~J to nugust and then decreased again up to November. At. Fetekro, older instars inc.reased again LII) t.o February.
After the suspccted poisoning in Docember at, Ouokoukro, in January young instars were more numerous than older mies. In contrast tn C. tligifafa younger instar numbers decreased until March.
At Tinbt the start. of t.he application seemed to bave the same efrect, on t.his species as on C. digifafa. However, c[;‘henmtrfopsychP sp. IlI was rare in August. Tn November, nearly a11 spccimens were first instar larvae, entl in December the spec.ies was almost eradicated. In January and Februarp, the sit.uat.ion was as in C. cligifnfcr vvith the dif’ference that, no live specimens were found in t.he st.ill Rater.
At. Kolornikro nnd Hocanda, the treatment had nearly t.he same rfft~ct as on C. digitafa with the difference that sevrral older xpecimena were captured et- Kolomikro in July. =It. bath sarnpling st,ations the numbers of specimens collected in samples after the application remained 10~.
6.1.X Cht~urnafopsflche sp. TV 1 I
Cher~mafo~.syche sp. T-il (fig. 9) seemed to be most seriously affect.ed by t.he st.c$ of flow. Only a few

HYDROPSYCHIDAE IN THE N’ZI 167
1911 1978 Jan. Febr. March JUW July Aug. Sepi. NOV Dec. Jan Febr. March
‘661 0
Febn
I ~247
FIG. 8. - Cheumutop~syche sp. III. Cf. fig. 7 for further tiot.nils
1970
March W JUIX July Aug. Sept. NOV. Dec. Jan. Febr. March
, -- ,,, 15116 1 14115 1 9110
50 1
- _ ri=41 1 ~57 1 n=3 1 n=5 1 n=O ! 617 , lu/11 i 9 14’15 ! 1”‘2 ; lW5 ’ Il=0 Il=0 1 n=15 +- a& 1 - ' 1 I 1 n=O 1 n=O ; 9”6 i ’
I
I I
n=O , n=O , I ;Y
1 1 I 1 1
2.E 30. I 1 1 1
1 ,ns. I I
I 1 n c do I I I I I I 80 1
t 1 I I -FI
10. 1 I I f I I I I I :.. alx r I 1 .: I . : . . . ;’ I I 5 I I I l I 1 1.:::::
12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ 12345~ ins+ar
FIG. 9. - Cheurnatopsyche sp. VII. Cf. fig. 7 for furt.her drtails
Rev. Hydrobiol. trop. 1.i (2) : 167-176 (1082).
specimr~ns were fountl in May, a11 older than second instar. In June thc population still seemed t,o br old ad t.hen tlie peroent.age of young instars increased unt-il ,July at, TinbC, Fetekro, and C~uokoukr».
At. t.lie c,ontrol st.at.ions development up to Lhe end of t-lie period st.udied was similar t-o that of Cherzma- fol.~!@~~ .sl~. 1 II. Only the recovery at. Ouokoukro after l3eceniber sermed less pronounced in Cheurnuto-
l~,yrhe sfj. I*II. M Tinb6, t-he situat.ion seemetl quival& to that. of Cherzmafopsyche sp. III, alt.hough onl,y a few specimens were caught durincy the periofl ot larvicide application. Aft.er larviridin& was st.oppec:l, ChP~rnlnftrpsycha SP. T-1 1 was captured only in Marc11 at. Iiolumlkro and Bocanda.
FI~+. 10. - .lcfhalt>Jlfcra dispar, at. the southern rapids of thc N’Zi. I:f. fig. 7 for fwther drtails.
T he fourt-h sepc.ies under study, -4. tZis~a~, was frequent. at. the beyinning of t.he investigat-ion at. t.he southorn aampling places. As already mentioned in srct.ion 5, thi s apecies was net able t.o survive the period viithout. flow and neefled up t.0 November to recolonizr the rnpid nt. Ouokoukro. Fig. 10 de- nlonst-rates t.liat in this SpPcies the numbers of »Id instam also increased with decreasing dischargr. In Decerubrr et- Ouokoukro. the population was net. as much affected by t.he expected poisoning as t.hose of ChaunlcIfc,p.?ycha. However, after that. poisoning in -4. tlispw t.he quant.it-ies of younger instars were al;io highrr than in the previous year.
6.2. Mean weights at the substrate surface 6.3. Instar distribution and mean weight in the
The clynamics of the populat.ions studied here are prtsbably ruost readily illust.rat,etl by the mean weight. changes [fig. Il). Excluding the period of larviciding and thosc data with only few specimens the following cyeneral patt,ern oi*c*urred: The specific, mean weight. b was hi& ai fhr beginning of 1977 and increased, remained constant. or decreased only sliglit21y over the period without. flow in t.he species present. after
hyporheic habitat
When the material in the tore samples was suf’ficient. for analysis, the mean weight of the hypo- rheic specimens was calculated (fig. 11). Mean weight of the surfac.e and hyporheic, specimens were in a comparable range. Differences in the instar distribu- tion were significanf. in three c.ases, but not extreme (Tab. III). Evidently the hyporheic fauna is net.
0.6
02
0.6
02
18
14
1.0
06
0.2
06
0.2
10
06
02 u l m--.
. ..--_._... L_: _ .-
F M A M J J A S 0 N D J F M month 77 1978
J 19
0 - 0 C. digitata 0 . . 0 Chcumatopsycbe sp.Y//
0 - - - -0 Cheumatopsychc 3,~. Ill O---O A. dispar
FTG. 11. - Mean dry wight of C. diyitata, Chezzmatopsyche sp. III, Chenmafopsychc sp. 1'II and A. dispar at the five rapids. Open synbols: n-c 10; solid SymhJk n>l0; large open sym-
bols: mwn weights in the hyporhcic habitat, (n > 10)
this period. Thrn t.he mean weight decreased until June/July. increasing again until August. -ifter the flood mean weigllts were lower than before the flood and increased again until t,he end of the study period.
Rw. Hydrohiol. frop. 1.5 (2): 157-176 (I?SP).

IIYDRC~PSYCHIDAE IN THE N'ZI 169
TABLE III
Percentage distribution of instars at thc surface of the substrat.um (surf.) and in the hyporbeic habitat (hypo. ; considering the 5-15 cm part of the Gore sample, which was the only one with higher abundancrs of hydropsychidsj. %z-analyses (msn-tables) are
bascd on thc original data
Larw
1 2 3 4 5 pupa
Ouolroulcro, Febr. 1977 . Cheumafopsyche diyitafa
surf.. . . hypo . . .
Chcumaiopsyche sp. III surf.... hypo...
Aeihalopfera dispar surf. . . . hypo.. .
Bocanda, hlarch 1977
Cheumatopsyche digitata surf.. . . hypo. . .
. .
. .
. .
. .
. .
. .
. .
. .
2 14 25 23 24 11 2 10 17 24 34 12 %” = 3.35
1 13 28 29 25 6 Cl 3 15 38 44 Cl j(- = 13.85
Cl 0 ‘2 ‘27 1 !f 5% cl 11 5 19 62 24 x” = 12.07
6 9 11 Cl %3 2.g 1 29 26 19 20 ü %” = HI .!I
rest.ricted t.o smallest larvae, a result similar to that. reported from a tropical stream in Malaysia (BIHHOP, 1973). The restric.ted space (see below) at t.he period of low flow probably led to an immigration of large spec,imens into the hyporheic interstitial. In Lempe- rate zones specimens may immigrate into this habitat frequently in streams wit.h an unstable bottom (SCHWOERBEL, 1967).
6.4. Nature and causes of life cycles
It is assumed that the species considered here are not able to develop from the egg t,o Lhe pupa within a period of about one or two weeks-to the best of my knowledge su& a rapid growth is known for African stream insects only from Simuliidae (LE BERRE, 1966). Then, the three Cheumatopsyche spp. must have survived t,he period without flow in the aquat,ic stage, at least in some of t,he rapids studied. This is indicat.ed by t.he old populat,ions found after t.he resumption of flow. These findings are contrary to those of other studies on Cheumatopsyche in inter- mittent. Africnn strexne, where recolonization was assumed to be due to egg-laying. Without checking t,he populat,ion structure, HARRISON (1966) proposed this for a Rhodesian and J. D. HYNES (1975a) for a Ghnnaian stream. Whet,her tAis difference between the N’Zi and the other studies was caused by diffe- renres in the species under study, t,he methods used or the actual climate is net, clear. The stream studied by HARRISON usually dried up for longer periods than the N’Zi, and most, of bis pools dried up completely.
‘In the stream studied by J. D. HYNES, the flow regime was similnr t,o that of the N’Zzi, and stagnant.
Reu. Hydrobiol. trop. 15 (2): 157-176 (lOS2).
pools remained permanently. However, no larvae of Cheumatopsyche were found there within the first period (6 to 8 weeks) of steady flow aft.er the dry period in the beginning of the year, until a mat of 1110~s and algae become est.abhshed. According to HYNES, Gherrmdopsyche larvae require a subst.rate of IIIOSS to use as a 1.1ase for sewing their nets, and therefore larvae occur only when mose is present,. As it cari kJe seen on fig. 5, in the present study hardly any Cherzmnfops~yche built. a net, a fact. well known for other Hydropsychidae from other studies (FEY and SCHUHMACHEH, 1.978; SCHRODER, 1976; SCHUH- MACHER, 1970). Therefore the presence or absence of these animais cannnt. be explained by t.he presençe or absence of suil.ahle net--sewing-places. Moreover HYNES, results are mainly baser1 on Surber samples from Bat. horizontal sandstones. In the N’Zi, such habitat., when stones were net. covered by macro- phytes, was rarely colonized by Cheumafopsyche. However, Chenvvvcxfopsyche could be found in micro- habitats ot.her t,han on the top of the st,ones in periods wit,h and wit.hout. mass on Lhese stones. T~US, in my opinion, tAe la& of Cl~eumafopsycl~e larvae in t-he Ghanaian st,rearu immediately aft,er the re- sumption of flow ctould be due t.o insufflcient sampl- ing.
In the case of the N’Zi, the questions remain as to where the specimens survivad. The possibi1it.y of survival tic:ep in the subst.rat~um as ment.ioned for example hg B~IIVET (1978), IMHOF and HARRISON (19911, WILLIAMS anal H. B. N. HYNES (1977) was checked by coring and di-ging, but: Ilot, anirnals were obtained. The ot.her possibility, the survival in tbe pools is diflicult t.o prove considering t,he

1 xr B. STATZNER
difference in t.he area of a rapid (some dozens of syuarenietersj and that, of a pool (several hectares). : ssuming that. dispersion is random, one bas to 1. sample a large arpa to find few specimens in the pools. ~~lthouplr net. intensively investigated, this possibi1it.y seenircl t;he most probable one, for Cheumatopsz@ze larvae were twice founrl alive in stagnant water: once in a rapid and (.)ncp in t-he pSaInIni31 of a pool.
Uaually C:haomatop.s~lrhe die in st.ill wat.cr and this was also otsserved for the three species of the N’Zi. F’hysiological changes may enable some specimens to survive without. current, in the dry season, a mrchani+rn which has been described for caddis larvae in temporary running waters (BO~JVET, 1978). The observed changes in weight. over t,he period without. flow coultl be due to differential mortality of diffrrent- inst,ars. Then, t.he presence of speciniens on the rapid after slow st.art,ed could be influenced as much by the survival rate as by the t.ime needed for re-immigrat.ion from t.he pools t.o the rapids, thereby afïecting the larval abundances.
These abundanc.es should be further influenced by emergence and rrpruduction. 111 untreat-ed rapids whick had net. dried up, pupae were present in almost a11 samples. Therefore, here, the presence of imagines cari be expect-ed at a11 t,imes of the year. Emergence data on Trichoptera from the Red Volta RiTer and the Nasia River, which were based mainly on Ckivww qtp., inclic.ate that, imagines left water in increasing nunlbers during decreasing discharge (ÇKISP, 1.966). Whrther t,his was the resuit, of a real increase in t,he emergr~nc~e rate within t,he population or wht’ther this was mainly caused by a concent.ration of spcciniens below t Ire emergenc.e trap due to a decrease of areas with suitable current. remains uncertain. Therefore that method as well as ot.hers iinually applied in studyin g su& questions give no indication as t.o when the imagina1 population reachecl its maximal abundance nor- do they indicate t-he period of maximal successful oviposition, bot.11 of whic*h are due t.o the ext,rrmely high dynaniic. of tdlr environnient~.
Moreover, tbe high abundance of first instar larvae could be due to retarded hat-ching (ILLIES, 1959) and low abund;lnce~ trf first. instar larvae could be due t.o high loss through drift, immecliat.ely aft.er hatching (STATZNER, 3978a). Hence the decrease of mean weight- in the period ,June/July and t,he shift in the population structure, interpreted as a peak in repro- duction at, first. glance, probably are caused by other phenomena. Hoivever, sinre a mass reproduction in that period seenls to he most probable in the moment., June and ,July are assunled to be the one major period of reproduct,ion. A lower level of reproduction or het,t.er larval hatching is indicated by the lower
relative abundance of youngest inst,ars t,hroughout the other periods of tire year.
It. is expected, that. t.he successful oviposition depends to a high degree on the discharge regime and the reactions of females to it, since Hydropsychi- dae enter into the water to fix the eggs t,o t.he sub- stratum (RADCOCX, 1953). Due to exarnples of how the larval abundance of cacldis flics is intluenced by imagina1 behaviour (STATZNER, 1975b, TOZER et al., 1981), one coulcl speculat,e t.hat a lower oviposit.ion act,ivity oc.curred at low discharge and at high dis- charge, whilst the period of medium discharge seems t.o favour oviposition. It, might be, also, that the period of highest. discharge was that of maximal oviposit,ion ac.t.ivity and that eggs dried up during discharge fluctuat.ions and t.herefore led to a new founding of the larval populations at Lhe end of the spate, when the data suggest a second peak of reproduction.
At. Ouokoukro, Cheumatopsyche digitata. and Cheu- matnpsyche SP. Ii1 seemed tao have reproduced after the suspected poisoning of the rapid in December, but. net, in the previous year at this rapid nor at Fetekro at t.he same time. This indicat,es, that ovi- posit,ion, hatching, or loss of larvae by drift. was c.ont.rolled kJy the abundanc.e and t.he age structure of the larval ~JopUht~ions 011 the rapid.
It. is well known for Hydrwpsyche (SATTLER, 1958; SCHUHMACHER, 197CI) and ChenTnafopsyche (GLASS and BOVBJERG, 1969) t,hat their larvae show a distinct intra- and int.erspecific nggression. The significance of the larval stridulation (JOHNSTONE, 1964) for this aggressive behaviour was studied by JANSSON and VOUORISTO (1979) who demonst.rat,ed t.hat stridulation in connection with size plays an import,ant role in the competition of larvae for space.
The st.ridulation ridges of t.he Cheunmtopsyche spp. of t,he N’Zi exhibit. distinct, specific dfferences (STATZNER, 19811)). Whether t.his led to different specific successes in competition or not is un- known, but, it. suggests, t,hat stridulation and size may determine the specific hierarchy in competition bet.ween larvae. Such c.ompetitions cari be expected t.o occur frequently during periods of decreasing discharge, i.e. decreasing areas with sufflcient current, when t,he larvae are expected to move from the place they seMed (EDINGTON, 1968). This fac.tor was probably responsible for t.he change in the relative abundanc.e of the spec.ies as well as in the instar distribution at, Ouokoukro in 1978 after the expected poisoning. One could expect that the “competitive pressure” on rec.ently hatched Young instars would be different. at. a densely populated rapid than in a rapid, where the population was previously eradicated.
HYDROPSYCHIDAE IN THE N’ZI 171
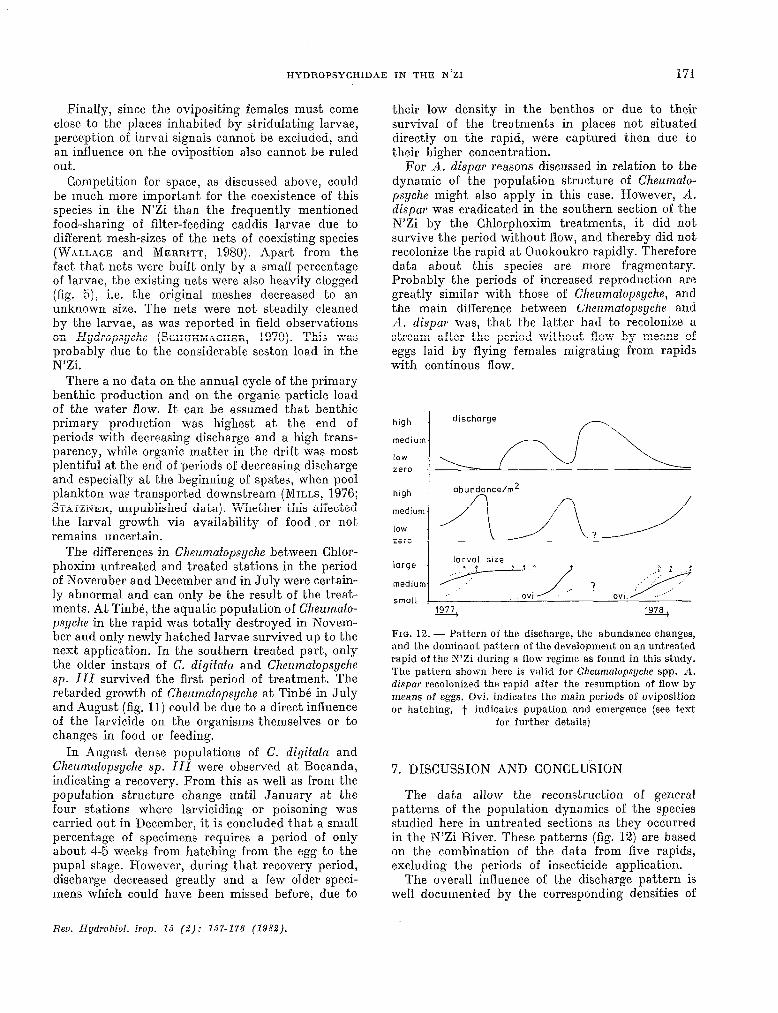
Finally, since the ovipositing females must corne close to the places inhabited by st,ridulating larvae, perception of larval signais cannot be excluded, and an influence on the oviposition also cannot be ruled out.
Competition for space, as discussed above, could be much more important for the coexistence of this species in the N’Zi than the frequently mentioned food-sharing of filter-feeding caddis larvae due to different mesh-sizes of the nets of coexisting species (WALLACE and MERRITT, 1980). Apart from the fact, that nets were built only by a small percentage of larvae, the existing nets were also heavily clogged (fig. 5), i.e. the original meshes decreased to an unknown size. The nets were not st.eadily cleaned by the larvae, as was reported in field observations on Hydropsyclze (SCHUHMACHER, 1970). This was probably due t,o the c.onsiderable seston load in the N’Zi.
There a no data on the annual cycle of the primary benthic production and on the organic particle load of t.he wat,er flow. It. cari be assumed Ihat benthic primary production was highest at the end of periods with decreasing discharge and a bigh trans- parency, while organic matter in the drift was most plentiful at the end of periods of decreasing disc.harge and especially at the beginning of spates, when pool plankton was transport,ed downstream (MILL~, 1976; STATZNER, unpublished data). Whether t.his affected the larval growth via availability of food or net, remains uncertain.
The differences in Chezzmafopsyche between Chlor- phoxim unt.reated and treated stations in the period of November and December and in July were certain- ly abnormal and cari only be ‘rhe result of the t,reat- ments. At. Tinbé, the aqua& population of Cheumato- ps@z~ in the rapid was totally destroyed in Novem- ber and only newly hat,ched larvae survived up to the next application. In the southern treat,ed part, only the older instars of C. digitata and Chezrmatopsyclze SI). III survived the first period of treatment,. The retarded growtb of Cheumatopsyche at Tinbé in July and August (fig. 11) could be due to a direct inlluence of t.he larvicide on the organisms themselves or to changes in food or feeding.
In August dense populations of C. digitata and Chezzmatopsyclze sp. III were observed at Bocanda, indicating a recovery. From this as well as from the population structure change until January at the four stations where larviciding or poisoning was carried out in December, it is conc.luded that a small percentage of spec.imens requires a period of only about 4-5 weeks from hatching from the egg to the pupal stage. However, during that recovery period, discharge decreased greatly and a few older speci- mens which c.ould have been missed before, due to
Rev. Hydrobiol. trop. 15 (2): 157-176 (1982).
their low density in t,he bent.hos or due to their survival of the treatments in places not situated directly on the rapid, were captured then due to their higher concentration.
For A. dispar reasons discussed in relation to the dynamic of the population structure of Cheumato- psyche might also apply in this case. However, A. dispar was eradic,ated in the southern section of the N’Zi by the Chlorphoxim treatments, it did not survive the period without flow, and tbereby did net recolonize the rapid at Ouokoukro rapidly. Therefore data about t.his species are more fragmentary. Probably the periods of increased reproduction are greatly similar with those of Chezrmatopsyche, and the main difference between Clzezzmafopsyche and A. dispar was, that the lat.ter had to recolonize a stream aft,er the period without flow by means of eggs laid by llying females migrating from rapids with continous flow.
high discharge
medium.
low
zero
2 high abundance/m
medium
low
zero
FIG. 12. - Patt.ern of the discharge, the abundance changes, and the dominant pat.tern of the development on an untreated rapid of the WZi during a tlow rcgime as found in t.his study. The pattern shown here is valid for Cheumnfopsyche spp. rl. dispar recolonized t.he rapid aft.er the resumpt.ion of flow by means of eggs. Ovi. indicates the main periods of oviposition or hatching, T indicat.es pupation and emergence (see text
for further detailsj
7. DISCUSSION AND CONCLUSION
The data allow the reconstruct.ion of general patterns of the population dynamic.s of the species studied here in untreated sections as they occurred in t.he N’Zi River. These patterns (fig. 12) are based on the combination of t.he data from five rapids, excluding the periods of insecticide application.
The overall influence of the discharge pattern is well document,ed by the corresponding densities of

larvae pw ms. The cross-section of a rapid and the result.ing surface area with current. during periods of decreasing discharge intluenct~s the density per unit. of area.
The current. investigat.ions indicat.e that Ch~urnc~- fopqcke spp. survived the period without, flow in their aquai% stqe. 5nc.e the larvae probably rearhed an agr of about, three mont.hs before the slow stcrpped, t-he inclividuals surviving t.he drought. of t,he rapids pmhably reached 5-6 months or morp ancl then emerged. From June/July until the start of the spate, a generat-ion with a shorter life span was produced. Chat happenetl during the high wat-er is u~k~mv~~ M’hrn discharge decreased populat;ions started getting older again. This was observed until the enc1 of f-l,& sf.udy.
It must, be st-ressed here, 1 bat. the pat.t.ern shown in fig. 12 is probably t-ypical of most. specimens in Wre iqulat.ion. The& is no doubt. that some of the ~pecimens behavrd differently, as indicated by brulien lines.
-4 similirr pattern to that given in fig. 12 probably would havr occurred in --I . dispcrr if t.he sout hern part of the lS’Zi had remaincd untrcated. HowevPr, thiu species had to rrcnlonize a dry rapid after the resuniption cif flo\v by means of eggs.
In tlrtlir larval stage, aquat.ic caddis flies survive periods without3 any op~:n wat.er only esceptional (BouvE.r, 1978; HARRISON, 1!366; IMHOF and HAR- HISoN, I!@l). Ofien species in tempornry waters eshibit. a rrlat.ivel;v long arlult. life compared 10 thr length of t.lie larval life (BOL~~ET, 1978, WrcxrNd, 1973). This ca11 also bc assumed for three of the species studied here, since the adult Chrzzmntops~/che were alA» cxapt-ured at. t.he entl of the period withoul. flow, during whicll rn1ergenc.e is assumed t,o be zero. Whet.her these specimens c*ontribut,e t.o t-he repro- duction in ,June-Julg or not remains unc:ert.ain. For the grographical area under st.ut:ly such a long life of adults as compared with that. of t-he larvae is well clociimen ted for Sinzrzlizznz tlanzrzosrzm (LE BERHE, 19M).
Considfbring t.lre long larval and imagina1 lifo of specimens, probably only t,wo or three generat.ions per year ean be ccornpleted. Thus t.hr life of the species under the environment a1 conditions studied here lastcd longer than is qenerally assumed for -4fric,nn insect.5 living in rrlatively warm wat,er streains neaI L t.hl? l?l.[UiitOr.
The term generation is user1 aboce in the sense of thr total 1engt.h of i..he period which was nceded by the avrrage individu& hatched from ergs at idrntical tirnes until oviposition of females respec.t.i- vely t.he death of imagines. It was also used in this sense in a former paper on t.he Trichoptera of thr Kelengo Rivrr (Zaïre) (STaTzNER, 1%%; p. 122), i.e.
several generations cari exist, at, the same t.inie in the population (SCHTVERDTFECEH 1979, p. 77). Bot.11 LEHMANN (1979) and KUPELKE (1981) disçussed t.he t.erm generation from tbe st.udy on the Kalengo Trichopt.era much more in the sense of single gene- rat.ions present. at. one time, as is frequently t,he c,ase in ternperate zonrs. Suc11 distinct, generations (cohorts) are difficult. to define in stream inverte- brates of t.he tropics.
Neverthrless sorte dominant patterns of develop- ment., caused b‘y’ a seasonality in t.he running waters, cari be recogmzed in t.he tropics. The rhythmic flut:tnations in thcb ernergence of the Kalengo Tri- choptera (STATZNER, 1976) as well as t.he changing instar distribution demonst.rat.ed in t-bis st.udy suggest. t.hat. suçh tlominant pat-terns of development exist.
In the N’Zi, it. is the changing flow regime which seenis t,o drterrnine t,he rlominant. pattern of the development. in tdlr EIydrnpsychidae. Due t.o the poor knowledgr of f1uct.uat.q factors, suc11 as the flow rngime in the N’Zi, in the case of the Kalengo, tAe light. parameter was put into this categnry. LEHMANN (1979) criticized thr: latter approach, argueing that day 1engt.h is so const-ant. near the equator that the light parameter must. also be constant. Considering brooks in a dense for&, t,his is probably c,orrect, as suggestlecl by data from DIHMHIRN (1961 ). However, ronsidering brooks in more open areas, in which the Iialengo was sitnated, inanlation is affec.t8ecl periodi- cally by fnd.ors net relat.ed to day length in the African tropics as ca11 be seen from GRIFFITHS (1972), XIALAIssE (1974) and PAGES rlt a/. (1981).
Rep0rt.s on the development of Chironomidae anrl Ephemeroplera (DEJoux, 1971; CORRET et cd., 1974) suggest that. bot.h qroups havr probably short,er life cycles in African ‘wat.ers t.han most, Trichopt,era. Therefore su& dominant ~Jat,kmS of t.he develop- mrnt. are presurnably more dif&ult. t,o recognize in t.he Chirrmomidne (st.udied by LEHMANN, 1979) and t-lie Ephemeroptera (studied by KOPELKE, 1981) t.han in the Triehoptera.
The approach of the dominant pat.t.prn of develop- ment. enables t.he rough estirnat,ion of the product,ivity of African stream invertebrates, which is otherwise very diff~cult. t.o ralculat,e in the t.ropic.s (BEADLE, 1974; BIYHOP, 1%3).
The maximal hiornass (in dry weight.) of the hydropsychid larvae and pupae of t.he N’Zi (tab. IV) exceeded t.he wet weight, values report,ed by PETR (1970) from Ihe Black Vo1t.a River. However, in PETR'S stur1-y sanlplec were collected only onc.e, at the end of the dry season. The annual mean in the N’Zi exceeded the total c.atldis fly biomass of the Tshinganda-Luhoho, a river system in the Zaïre (STATZNER, 1975). Hvwever, the rapids in the N’Zi represent.ed only a reliifirely Small area of the t0tal

HYDROPSYCHIDAE IN THE N'ZI 173
TABLE IV
Monthly biomass, annual mean biomass, and estimation of annual product.ion (cf. t.ext for mode of calculation) per ma on the rapid at Ouokoukro (a11 items in mg dry weight)
1977 1978 Biomass Production J F M/A M J J A S/O N D J F M 1977 1977
Cheumafopsyche digifafa.. . . . . . . . 158 9397 0 14 0 4 364 n 1 0 97 1877 9333 828 9846 e
Cheumafopsyche sp. III. . . . . . . . . 98 1391 0 1 0 6 442 a 3 0 26 542 1854 162 1937 r
Chsumafopsyche sp. VII’. . . . . . , . 34 105 0 0 0 0 161 0 6 0 18 56 21 208
Aefhalopfera dispar... . . . . . . . . . . . 1982 3510 0 0 0 0 0 Z? 0 238 203 406 275 477 8872
SU~........................... 2272 14493 0 15 0 10 822 OP 4 244 326 2843 11518 1488 20863
* Weights and growth rate based on the instar weights of Cheumafopsyche sp. III
river, i.e. the hydropsychid biomass of an average m2 of a whole section of the N’Zi. cari be expected to be very low.
Due to the fact, that larval densities per m2 were highest during the periods when the popula’tions reached maximal mean weight,s, productivity cannot be calculated according to most of the freyuently applied methods (EDMONDSON and WIN.BERG, 1971; WATERS, 1977). Thus annual production (P) was roughly estimated by applying t.he inst.antaneous growth rate met,hod over an entire life cycle of the specimens, a method described and discussed in detail by WATERS (1969), using the formula
P=in weight, of pupa weight of larva 1
x mean biomass.
It was assumed that one generation was completed in the period November until May/June (Cheumato- pjs@e) respect,ively November unt.il February (A. dispar) and anot.her one from June unt.il August (Cheumatopsyche), the period of the spate was completely neglect.ed.
The annual production yielded by this estimation (Tab. IV) seemed to be qu1t.e high on the flrst view, especially if the figures are doubled due to the hypo- rheic abundance of specimens. However, as men- tioned above (section 3), the rapids of the N’Zi represented lake outflows during long periods of the year. In these lake outflow communities the hydro- psychids were the most dominant benthic group (biomass), especially if one considers solely that group of bent,hic invertebrates, which are able t,o feed by filtration. When cornparing the production est,imations of the hydropsychids at Ouokoukro with studies evaluating t.he product,ion of filt.er feeders s.l. in t,emperate lake out,flows (STATZNER, 1978c, 1979; ULFSTR~ND, 1968), the figures of the N’Zi are yuite low.
This underlines data obtained from stagnant Afric.an waters (DEJOUX et al., 1969; HART, 1950; HAHT and ,~LLANSON, in press; LÉVÊQUE et a!.,
I972), a11 suggesting, that the generally assumed high productivity of t.ropical macroinvertebrates in African wat,ers has not yet been proved conclusively.
Considering t.he eradication of a11 four species studied here, which Will almost certainly occur if Chlorphoxim is applied as an operational insecticide in t,he O.C.P.-area, the following is concluded:
(1) the relat.ively low productivity of these species on a rapid as well as the small area of a rapid in comparison to the large area of a pool indicate, that t,heir eradication seems to be of minor importance concerning the relat.ionships between productivity of invertebrates and edibles flshes;
(2) it has been established that Chezzmalopsyche spp. are predators of Simzzlium damnosum (BURTON and Mc. RAE, 1972; SERVICE and EL~UARD, 1980). The possibility of Chezzmatopsyche eradication and the generally occurring effects of target species’ predator eradicat,ion by man should be seriously considered before Chlorphoxim is applied against S. damnosum.
LManuscrif rqu au Service des Édifions de I’O.R.S.T.0.M. le 7 décembre 1981.
Al1 colleagues and friends from the O.R.S.T.O.M. Hydro- biological Dnit and fhe O.C.C.G.E. as well as their families in Bouake received me in a warm and kind manner. Mr. G. NON- KODABA accompanied mc on a11 excursions int.o the bush and, together with Mr. T. MAYADOU and Mr. M. Brnounr, carefuily sorted out t.he material in the laboratory. Al1 August samples were takcn by Mr. J. J. TROUPAT. Hydrobiological colleagues from O.R.S.T.O.M. and O.C.P. freyuently discussed with me limnological problems of the region and offered a lot of hclp to overcome a11 technical problems. Miss B. HOLLER, Miss C. PETERSEN, Mr. L. GROWNOW, and hIr. T. THOMAS kindly prepared most. of the graphs. hlrs. L. MOLE, hir. D. Mann, and two anonymous reviewcrs commented on, and corrected the English manuscript, and Mr. F. GIBON translated the summary into trench.
Al1 this help is very gratefully acknowledged.
Rev. Hydrobiol. irop. 15 (2): 15Y-176 (1982).

B. STATZNER
REFERENCES
Raococ~~ (R. AI.), 1963. - Observation of oviposit.ion 177x11~
water of thc aorial insect Hydropsyche nngusfipennis (Gurtis) (Trichoptera j. Hydrobioloyio 5 : 2?2-225.
BE \I~LE (L. C:.), 1c)ï-t. - Thr inland waters of t.ropical Africa. Longman, London, 365 pp.
Brsnor (J. E.), lYï3. - Lirnnology of a small hialayan rive] Sungai Gon~bak. .Junk, The Hagut~, Monogr. Biol. i-c :
.a35 p.
BÜTTC~BK (K.), lHï5. - F’rodllktionsbioloCrischt? Stxtlien an tir117 zrntralafrikanischPII Brrgbaah Kalengo. .Irch. Hydrflbild ï5 . l-11 . . C.
RouvEi (IM.), 197.Y. - Adaptations physiologiyutrs et cornpor- tementales des Sfern~phylaz (Limnephilidar) RUS eaux tenq~orairrs. Proc. 2nd Inf. Symp. Trichoptcru, 197i, Jmk, The Htryue: Ilï-119.
BURTOIY (G. J.) ami hic. RAE (T. bl.j, 1972. - Observations on tricliopi era predators of aquat.ir stages of Simuliurn ticz»~n~~szzn~ and other Sin~rzliwn species in Ghana. J. dirai. Ed., Lt : 389-294.
C:.~rus (I-I.), 1971. - Hydrologie 1111 Bandama II. Les aflluents (111 Bandama Blanc. Rapport. O.R.S.T.O.M., Adiopo- dnllm~, 190 pp.
(::fiRRET <s. A.), SELLICIi (n. U.), a7ld \~ILLOUUHWY (N. c;.),
197-1. - Ntrt.es 017 th& biology of the maglly PoniU(z tzdzznfa in \I’est. Africa. J. zo>l., Lorzd., 17?. : i91-SO?.
C;RIDL.kND ii:. Ci.), 195% - Ecological fact.ors affecting the numbers of snails in a permanent. stream. J. frop. Med. Hyg., Al : 3-7.
(-:RISP iG.1, 1956. - .4n ephameral faunn of torrents in thc northern trrritorirs of thr Gold C:oasf., wi1.h special reference t.0 the enrrnies of SSimzzlizzm. ;Innuls frop. nid. 50 : c:,j(J-=$7 - .
DE.JWX (Ci.), 1!571. - Rroherches sur le cycle dr dé~el«pp~-
ment dr Chironnmzzs pzzfrhcr (Diptrra : Ghironomidnc). cm. Ed., 103 : 466-170.
DEJ~IJS (ci.), 1975. - Nouvelle twhniqua pour test.er in sifu l’i7npact. dr pest.iüides sur la faune aquatique non cible Cah. 0. R.S.T.O.AI., s&. Enf. rned. ef Pnrasifol., 13 : ïF>-NJ.
t)E.rOtrX (r:.), L.\I.JZANNE CL.), t!t L&&UE (G.), lt%!). Évoluf-ion qualitative of. quantitative de la faune benthique dans ta partie est du lac Tchad. Cah. O.R.S.T.O.AI., sér. Hydrobiol., vol. III, nu 1 : 3-58.
DE hl~r<oiva (B.i, 1981. - Z( mation ichtyologicp~e du bassin ~II Banda~~~:~ tC;tXr d’ivoire). Rez~. Hydrobiof. trop., 1-I : W-SS.
DIRN~EIJC (1.1, 1961. - Entomological st.udies from a high towtr in SIp711ga For&, Uganda III. Light int.ensit.ies at different. lrvels. Trans. R. Enf. Soc. Land. 113 : 270-27-l.
E~JIWT»N (J. M.), 196X. - Habitat preferences in net-spinning caddis larvac with special rrference to fhr influence of mater velocity. .J. .-lnirrz. Ecol. 37 : 675-69’3.
Er~~conrsa~ (W. T.) and WINRERC~ (T. T.), 1971. - A manuat on methods for the assessment of sccondary produc- tivity in fresh waters - IBP Handbook 17, London, 358 pp.
E’EY (J. h1.j und SCHUHMACHER (H.), 197S. - Zum EinfluB wrchsclnder Temperat.ur aut den Netzbau yon Larven der Kiicherflieprn-.4rt. Hydropsyche pelluçidzrla (Tricho- ptera : Hydrops)-chidar). Enf. Gernz., 4 : 1-l 1.
GIBBS (D. G.), 1973. - The Trichoptrra of Ghana. Dfsch. Enf. %. N.F., 20, 363-124.
GLMS (L. kV.1 and BI~VB.IERCT (R. Y.), l!X!l. - Density and dispersion in laboratory populations of caddistly larvao (Chezznztrfopagche, Hydropsychidae). Ecoloyy, 50 : lOS% 1084.
GRIFMTIIS (J. F.j (ed.), 1972. - Climates of Africa. Elseviev, Amsterdam, M’orld Szzrzwy of climafoly 10 : 60-t p.
H~~RRISON (-4. D.), l!.WX. - Rec.«l«nisat.i«Il of a Rhodcsian stream after drnught.. Arch. Hydro6ioI., 6” : 405421.
HART (R. C:.), 19PO.- Emhryonic duration and post-emùryonic growth rates of the tropical freshmater shrimp Cnridina niloficu (Decapotin : Atyitlas) under laboratory and rspori7nentnl fieltt c.ont.tit.ions. Freshnxzf. Biol., 10 : 297-315.
~%ART (R. C:.) ami ALLANSON (B. R.), in press. - Energy requirements of the tropical frcshwater shrimp Caridinu niloficu (Decapotla : Atyidacj. Verh. Infernnf.
T’erein. Limnol., 21.
Hunes (H. B. N.) and Wrr.~rxtrs (T. R.), 19@2. -- The offe&. of DDT on the t’auna of a Central African stream. Annls Trop. bled. Pnrasifol., 56 : 78-91.
HYNEY (J. D.), 19% a. -Annual cycles of rnacr»-invertebrstes of a river in southern Ghana. Freshzonf. Biol., ù : 71-83.
HYNES (J. D.), 1975 b. - Downst.ream drift of imiertehrates in a riverin southern Glraua. Freshavzf. BioLj 5 : 51553%
ILLIES (J.), 1959. - Retardierte Schlupfzeit. von Baefis- Gelegen (Iris., E~~hem.).~~ufzzrcz~issensrhaferz,~6: 119-12lJ.
IMHOF (.J. G. ,A.) and H~\RRIRC~N (.4. D.), 19Yl. - Survival of Diplccfronu modcsfu Banks j’I’richopt.rra : Rytlropsy- chidaej durinp short periods of dessiccat.ion. Hydro- bioZoyiu 77 : 61-63.

HYDROPSYCHIDAE IN THE N’ZI 175
JANSSON (A.) and VUORI~TO (T.), 1979. - Signiflcance of stridulation in larval Hydropsychidae (Trichoptera). Behauioar 71, 167-186.
JOHNSTONE (G. ‘iv.), 1964. - St.ridulation by larval Hydropsy- chidae. Proc. R. enf. Soc. Land. (A) 39 : 146-150.
KI~IMINS (D. E.), 1963. - On the Trichoptera of Ethiopia. Bull. Br. Mus. naf. Hisf. Enf. 12 : 119-170.
KOPELKE (J.-P.), 1981. - Okologische Studien an Eintags- fiiegcn am Beispiel der Emergenz des zcnfralafrika- nischen Bergbachcs Kalengo (Zaïre). Enf. Abhnndl. Sfaafl. Alus. Tierk. Dresdcn 44 : 9-43.
LE BERRE (R.), 1966. - Contribution à l’étude biologique et écologique de Simulium damnoszzm Theobald, 1903 (Diptera, Simuliidae). M&n. O.R.S.T.O.AI., 17 : 204 p.
LEIIYANN (J.), 1979. - Chironomidac (Diptera) aus FlicBge- wassern Zentralafrikas (Systematik, Okologie, Ver- breitung und Produktionsbiologie). Teil 1 : Kivu- Gebiet, Ostznire. Spiricrna, Suppl. 3 : 144 p.
LkvÊ~un (C.), CARMOLIZE (J.-P.), DEJOUX (C.), DURAND (J.- R.), GRAS (R.), ILTIS (A.), LnnroAmx (J.), LO~DENS (G.), LAUZAIL’NE (L.) et SAINT-JEAN (L.), 1972. - Recherches sur les biomasscs et la productivito du Lac Tchad. In : Kajak (Z.) and Hillbricht-11kowslca (A.) (eds.). Productivity problems of freshwaters. Warszama, Krakow : 165-1Sl.
LÉvEgun (C.), ODEI (M.), and PUGH THOMAS (M.), 1977. - The Onchocerciasis Control Programme nnd the moni- toring of its effrct on t.he riverinr biology of the Volta River Basin. In : Perring (F. H.) and Mellanby (K.) (rds.). Ecological offects of pesticides. Linn. Soc. Symp. Ser. 5 : 133-143.
hbLAISSE (F. P.), 1974. - Phenology of the Zambezian woodland area with emphasis on tho Miombo ecosystem. In : Lieth (H.) (ed.) : Phenology ami seasonality modeling. Springer, New York, Heidelberg, Berlin, 269-286.
MANG~LSDORF (J.) und SCHEURMANN (K.), 1980. - P~UB- morphologie. Oldcnbourg, Miinchen, Wien, 262 pp.
hIxLs (hi. L.), 1976. - The Chaoborus (Diptera : Chaoboridae) component of the invertebrat.e drift of t.he Mwenda River, Lake Kariba, Central Africa. Hydrobiologin, 48 : 247-250.
O.R.S.T.O.M.-O.M.S., 1976. - Rapport annuel d’activit,é. Convention O.R.S.T.O.M.-O.M.S. : Surveillance du milieu aquatique, Bouaké, 55 pp.
PAGBS (J.), LEnrassoN (L.), and DUFO.LTR (P.), 1981. - Primary production measurement in a brackish tropical lagoon effect. of light, as st.udied at some stations by the 1% method. Reu. Hydrobiol. frop., 14 : 3-13'
PETR (T.), 1971). - The bottorn fauna of the rapids of the Black Volta River in Ghana. Hydrobiologia, 36 : 399418.
RAMANANKASINA (R. E.), 1973. - Contribution a l’étude dC la biologie de deux especes d’Éphémere : Elassoneuria insulicolu Demoulin, Proboscidoplociu sikorai Vayssiere, de la rivieie Andriandrano (Mandraka), sous-préfec’urc de hfanjakandriana (RIadagascar). C.R. Acad. S:. Paris, 277 : 1513-1515.
REZI. Hydrobiol. trop. 15 (2): l.tïï-176 (1982).
SATTLER (W.), 1958. -Beitrage ZUP Kenntnis von Lebensweise und Korperbau de.r Larve und Puppe von Hydropsyche Pict. (Tiichoptera) mit. brsonderer Berücksichtigung des Netzbaues. Z. Morph. okol. Tiere, 47 : 115-192.
SATTLMR (W.), 1963. - Uber dcn Kbrperbau, die Gkologie und Ethologie der Larve und Puppe von Macronemn Pict. (Hydropsychidae), ein ais Larve sich von @ hlikro- Drift 8 erntihrendes Trichopter aus dem Amazonas- gcbict. Arch. Hydrobiol., 59 : 26-60.
SATTLER (W.), 196% - Weitere Mitteilungen über die Oketho- logie einer neot.ropischen dincronema-Larve (Hydro- psychidae, Trichoptera). Amuzoniana 1 : 211-229.
SCHROUER (P.), 1976. - Zur Nahrung der Larvenstadien der Kocherfliege Hydropsyche insfabilis (Trichoptera : Hydropsychidae). Ent. Germ., 3 : 260-264.
SCHUHMACHER (II.), 1970. - Untersuchungen zur Taxonomie, Biologie und Gkologie einiger Kocherfliegenart.en der Gattung Hydropsyche Pict. (Insecta, Trichoptera). Inf. Revue ges. Hydrobiol., 55 : 511-557.
SCHWERDTFEGER (F.), 1979. - Gkologie der Tiere II. Demüko- logie (2. Aufl.). Parey, Hamburg, Berlin, 449 p.
SCH\~OERBEL (J.), 1967. - Das hyporheische Interstitial ais Grenzbiotop zwischen oberirdischem und subterranem Gkosystem und Seine Bedeutung für die Primar- Evolution von Kleinsthühlenbewohnern. Arch. Hydro- bioZ./Szzppl.j 33 : l-62.
SERVICE (hf. W.) and EL~UARD (J.-M.), 1980. - Serological identification of the predators of the 5‘. damnosum comples in Ivory Coast. Bull. Enf. Res., 70 : 657-663.
SIOLI (H.), 1965. - Bemerkungen zur Typologie amazonischer Flüsse. Amazoninna, 1 : 7-i-83.
SIOLI (H.), 1975. - Tropical rivers as expressions of t.heir tcrrestrial environments. In : Golley (F. B.) and Medind (E.) (eds.). Tropical Ecological systems. Trends in terrestrial and aquat.ic research. Springer, New York, Berlin, 275-288.
STATZNER (B.), 1975. - Zur Longitudinalzonierung eines zentralafrikanischen FlieBpew%scrsystems unter beson- derer Beriicksichtigung drr Kijcherfliegen (Trichopt.era, Insecta). Arch. Hydrobiol., 76 : 153-180.
STATZNER (B.), 1976. - Dit Kocherflirgen-Emergenz (Tricho- pt.era, Insecta) aus dem zent.ralafrikanischen Bergbach Kalengo. Arch. Hydrobiol., 78 : 102-137.
STATZNER (B.), 1978 a. - Organization of large-scale fleld triais on the effects of larvicide application on non- target. organisms (with particular refcrence to benthic invertebrates). WHO/OCP/SWG/78.8 : 8 pp. (mulfigr.).
STATZNER (B.), 1978 11. - The effects of flight behaviour on the larval abundance of Trichoptera in the Schierensee- brooks (North Germany). Proc. 2nd Inf. Symp. Tri- chopfera, 19ii, .Junk, The Hague : 121-134.
STATZNER (B.), 1978 c. - Factors that detcrmine the benthic secondary production in t.wo lake outflows - a cyber- netic model. Verh. Internat. Serein. Limnol., 20 : 1517- 1522.

1X R. STATZNER
STATZNEH (B.), 1979. - Dei- obere und Untere Schierensceharh iSîhltislvig-Holstrin). Strukturen und Funktioncn in zwci norddeutschrn Se(~-AnsflIlSs-S~~al.enlrrl, untrr I~~»II-
derrr Bc~rücksiclit.i~ilr~F dw BIakroinvertebratrn. Dis~. Kiel, 551 lbp.
ST.~TZR’ER (B.), 1981 a. - The relation betwren <! hydraulic stress 6, and microdistribution of ùenthic macro- invertehrates in a lowland running water system, the Schiererlseebrc,c,ks jNort.11 Gcrmany). &ch. Hydrobiot. 91 : 192-L 1%
STATLNER (B.), 1981 1~. - h progress report on Hydropsychidar from the Ivory Cioast : characters for the specitlc identification of larvae and populations dynamics of four abundant. species. Serics Entomol. 213 : 329-335.
‘I’o%E.R (IV. E.1, DESH (V. H.) and SOLEM (J. CJ.j, 1981. - Bionomics and atlult. behavior of a lentic cadtlis fly, Nccfop,sfyche afbidu (Walkert. Amer. Mid/. NU~., 106 : 171-1-14 *< .
Tmrnn.4~ f.I.-.J.), l!ldl. - Dispositif à gouttieres multiples destine à tester in situ la toxicité des insecticides vis-à-vis des invertt~bres brnthirlues. Reu. Hydrohfol. frop., 14 : 119-152.
I~LF’~TR.~ND (S.1, l!it.#. - Rent.hic animal communities in Lapland strrams. Oi!i«s/.SuppZ., 10 : I-126.
VISSER (s. zi.) and \~ILLENEIKE (J.-P.), 1975. - Similarities and differences in t.he rhemical composition of waters from West, C:entral and East Africa. Verh. Infernczf. Tierein. Limnof., I!i : 1416-14%.
WALLACE (J. R.) and ~UBHRITT (B. W.), 1980. - Filter-feeding ecology of aquatic insrcts. .47m. Reo. Enfomol., 25 : 103-130 .?.
WATERS (T. F.), I%!l. - The t.urnover ratio in product.ion ecology of freshwater invertebrates. .4m. Nnf., 103 : 173-185.
WATERS (T. F.), 19ïï. - Secondary production in inlnnd waters. dtfu. Ecoi. RRR., 10 : 91-104.
WIGGINS (C;. B.), 1973. - .A e.ontribution to t.he hiology of caddistlies (Trichopt,eraj in temporary pools. Life Sci. Contr., H. Onf. Mlzs., 88 : 28 pp.
WILLIAMS (III. D.) and HYNEY (H. B. N.), 1977. - The ecology of t.emporxy StrWDls II. General remarks on t.emporary streams. Inf. Reuzze yes. Hydrobiol., 62 : 53-61.
ZWICIC (,P. i, 1976. - Neoperfa (Plccoptera, Perlidae) emerging from a mountain stream im central Africa. Int. Revue yes. Hydrobiol., 61 : 6X3-697.
While the manuscript was in print. 1 was ahle to identify- the two unnamed Chewncztc~pysche spp. with the help of material kindly send to me hy Dr. P. Ci. B.&aN.&an (Brit.. MUS.~. Chezznzafopsyche sp. III is C. fafcifern (ULRIER) and Chezzmafopsyche sp. VII is 1.:. ropioso Kr.warws.
Rev. Hydrobiol. frop. lb (2): 1.5;-176 (lSS2).





























