Embed Size (px)
Citation preview

Development 112, 1115-1121 (1991)Printed in Great Britain © The Company of Biologists Limited 1991
1115
Polyclonal origin of pancreatic islets in aggregation mouse chimaeras
L. DELTOUR1, P. LEDUQUE2, A. PALDI1, M. A. RIPOCHE1, P. DUBOIS2 and J. JAMI1
1 Vnili 257 de ilnstitut National de la Santi et de la Recherche Mtdicale, Institut Jacques Monod du Centre National de la RechercheScientifique et de I'Universttt Paris VII, Tour 43, 2 Place Jussieu, 75251 Paris Cedex 05, France2Uniti de Recherche Assoctee 559 du Centre National de la Recherche Scientifique, Laboratoire d'Histohgie, Faculti de Midecine Lyon Sud,69600 Oullins Cedex, France
Summary
In the present study, we have examined the origin andgrowth pattern of the /J cells in pancreatic islets, todetermine whether a single progenitor cell gave rise to allthe precursors of the islets, or if each of a few progenitorcells is the founder of a different islet, or if each islet is amixture of cells originating from a pool of progenitorcells. Aggregation mouse chimaeras where the pancre-atic fi cells derived from each embryo can be identified inthe islets on histological sections were analyzed. In twochimaeras, all the islets contained cells from both the
aggregated embryo. This clearly demonstrates that eachislet resulted from several independent cells. In addition,the /J cells derived from either embryo component werein very small clusters in the islets, suggesting that in situcell division did not account significantly for isletgrowth.
Key words: endocrine pancreas, /? cell growth, transgenicmice.
Introduction
Pancreas evolves as two evaginations from the duo-denum, beginning in the mouse at day 9.5 and 10.5gestation, respectively, and outgrowing independentlyas ventral and dorsal pancreas that coalesce later at day11 (Rugh, 1968). Early endocrine cells emigrate fromthe cell layer that constitutes the developing exocrinegland and separate from it, most probably by a changein the axis of cell division allowing the escape from theductule lumen of one of the daughter cells (Pictet andRutter, 1972). It is still unclear whether Langerhansislets grow through accumulation of cells originatingfrom the exocrine pancreas, or by proliferation ofprimitive islet cells, or by both processes.
Discrimination between these different possibilities isdifficult to make directly in vivo since individual cellscannot be distinguished from their neighbours. Toovercome this problem, mouse chimaeras could be veryuseful, since these experimental animals are made frommixtures of cells contributed by two different embryoswith different genotypes that are brought together andallowed to develop. If all the fi cells of a givenpancreatic islet are derived from a single progenitorcell, each islet in mouse chimaeras will be entirely ofone genotype or the other. On the contrary, if an islet isderived from multiple progenitor cells, it will be madeof a mixture of cells from both genotypes. For instance,using this method, Thompson etal. (1990) have recentlyshown, by in situ hybridization combined to immuno-cytochemistry on stomach sections from mouse chim-
aeras, that gastrin cells share a common stem cell withthe other cells of a gastric gland. Chimaerism ofpancreas as a whole has been documented (Williams etal. 1988) but, up to date, no evidence for pancreatic isletchimaerism has been presented. A cell marker welladapted to address the question of pancreatic fi cellchimaerism is a human C-peptide/proinsulin productpresent in certain transgenic mouse lines, because thisproduct is readily detectable in single fi cells inhistological sections of pancreas (Bucchini et al. 1989).The origin of every fi cell in any islet of chimaericanimals can thus be determined.
In the transgenic mice carrying the human proinsulingene, the human gene product was easily demonstratedto be expressed specifically in all the fi cells of thepancreatic islets. This was done on tissue sections byimmunocytochemistry using specific antibodies directedagainst the human C-peptide (Bucchini et al. 1989). Inthe present study, aggregation chimaeras were madebetween transgenic and nontransgenic mouse embryos,and islets of these animals were analyzed by immuno-cytochemistry on pancreas sections, using specificantibodies directed against the human C-peptide.Independent of their embryo of origin, the fi cells wereidentified by their staining with an anti-insulin antibodythat did not discriminate between human and mouseinsulins. The ficells derived from the transgenic embryocomponent of the chimaeras stained in addition with aspecific anti-human C-peptide antibody that did notbind the mouse C-peptides. It was thus possible todetermine whether all the $ cells of each pancreatic islet

1116 L. Deltour and others
derived from a single progenitor cell. In this study, weshow that islets result from several independent cells.
Materials and methods
MiceThe two types of embryos used for the construction ofaggregation chimaeras were from crosses of (C57BL/6xDBA/2)F! and of Tg74 transgenic mice, respectively. The(C57BL/6xDBA/2)F! parental mice were brown, but at theF2 generation, coat pigmentation is brown, black, or grey.Tg74 are transgenic mice on SJL/J background and have awhite coat. They carry about 50 copies of an 11 kilobase pair(kb) human DNA fragment including the insulin gene,integrated at a single locus on chromosome 7 (Michalova et al.1988). Human proinsulin is synthesized exclusively in pancre-atic islet cells and can be demonstrated by immunocytochem-istry on pancreas sections, using a specific anti-human C-peptide antibody directed against the other cleavage productof proinsulin. In Tg74 mice, all ft cells, as identified by an anti-insulin antibody that does not discriminate between humanand mouse insulins, stain with the anti-human C-peptideantibody, i.e., synthesize human proinsulin. The human(pro)insulin was separated from mouse (pro)insulins byreverse-phase HPLC and represents half of the total(pro)insulin in the pancreatic islets (Fromont-Racine et al.1990). Tg74 mice have normal plasma levels of glucose andtotal (human-(-mouse) insulin (Bucchini et al. 1986).
Females were superovulated by an intraperitoneal (ip)injection of 5 IU of pregnant mare serum, followed 46h laterby ip injection of 5 IU of human chorionic gonadotropin,placed with males, and checked for vaginal plugs the nextmorning (day 1). 8-cell-stage embryos were collected on day 3of pregnancy. Aggregates were produced between(C57BL/6xDBA/2)F2 and SJL/J-Tg74 embryos. The zonapellucida was removed by treatment with a 0.5 % pronasesolution. Pairs of embryos, one of each type, were aggregatedin microdrops and cultured overnight. Those embryos thatformed chimaeras and developed into single blastocysts weretransferred into uterine horns of day 3 pseudopregnant(C57BL/6xCBA/j)F1 recipient females (Hogan et al. 1986).
Isolation and analysis of DNAThe mice were screened by a dot-blot assay, using 20 \i% DNAprepared from tail fragments as previously described (Buc-chini et al. 1986). The probe was an 11kb //indlll genomicDNA fragment radiolabeled with 32P-dATP and 32P-dCTP byrandom priming. The hybridization was carried out with106ctsmin~1mr1 hybridization buffer. The filters werewashed (final wash) in O.lxSSPE (0.36M NaCl, 0.02MNaH2PO4, pH7.4, 0.002M EDTA, pH7.4) at 65°C andexposed to X-ray films.
Determination of human C-peptideThe mice were analyzed for the presence of human C-peptidein the urine by a radioimmunoassay as described (Bucchini etal. 1986). The assays were carried out with 20[A urine, L25I-labelled (specific activity 2000Cimmon1) human C-peptide(CEA-ORIS) and an antiserum raised against human C-peptide in the goat. The antiserum was kindly provided byM.A. Root and B.H. Frank, the Eli Lilly Company.-
Fixation and tissue preparationMice were killed at 1.5 or 4.5 months. The pancreases were
rapidly removed, and small fragments were fixed in severalmixtures such as glutaraldehyde (2.5% for lh), or parafor-maldehyde (4%) plus glutaraldehyde (0.5%) for 2h, orsolution of Bouin-Hollande sublimate without acetic acid (for6-12 h). The tissues were embedded in Paraplast. Sections3-5 fan thick were cut and mounted on gelatin-coated slides.The sections were deparaffinized and rehydrated beforeprocessing for immunocytochemistry.
AntiseraThe following primary antisera were used: guinea pigpolyclonal anti-bovine insulin N° 8309 (Novo, Copenhagen,Denmark) diluted 1:1000; rat monoclonal anti-human C-peptide N° GN-ID4 (kindly provided by Dr O. Madsen,Hagedorn Research Laboratory, Gentdfte, Denmark) diluted1:2000; rabbit polyclonal anti-C-terminal porcine glucagon N°GAN-8 (kindly provided by Dr A. Kervran, CNRS/INSERM, Unitd 264, Montpellier, France) diluted 1/5000;and rabbit polyclonal anti-somatostatin N° 19608 (kindlyprovided by Dr M.P. Dubois, INRA, Nouzilly, France)diluted 1:2000. Peroxidase-labelled goat anti-guinea pig (oranti-rat) immunoglobulin G (IgG) was purchased fromNordic Immunology (Tilburg, The Netherlands), and peroxi-dase-labelled goat anti-rabbit IgG from Dako-Immunoglobu-lins (Copenhagen, Denmark).
ImmunocytochemistryThe sections were processed at room temperature in a humidchamber by the indirect immunoperoxidase method ofNakane and Pierce (1966), with some modifications (Leduqueet al. 1987a). Briefly, sections were sequentially incubatedwith the following solutions for the times indicated: 1:40 non-immune serum for 15min; primary antiserum for lh; 1:200peroxidase-labelled goat anti-species IgG for 30min; and0.04% diaminobenzidine hydrochloride/0.03 % hydrogenperoxide for 30s-lmin, as suggested by Graham andKarnovsky (1966). Sections were dehydrated, cleared andmounted in DPX.
Immunocytochemical specificity testsThe specificity of the rat monoclonal anti-human C-peptide N°GN-ID4 has been checked by a solution phase equilibriumantibody-binding assay (Madsen et al. 1985), and immunoflu-orescence (Bucchini et al. 1989; Fromont-Racine et al. 1990).The antibody GN-ID4 reacts with both human proinsulin andhuman C-peptide and is species-specific (Madsen et al. 1985).It stains pancreatic /S cells of transgenic mice carrying thehuman insulin gene, but does not react with nontransgenicmouse pancreas. All the cells positive for insulin also stain forhuman C-peptide; no glucagon or somatostatin cells give animmunoreaction for human C-peptide (Bucchini et al. 1986).The RIA and immunocytochemical specificities of the anti-insulin, anti-glucagon and anti-somatostatin have beendescribed elsewhere (Leduque etal. 1985; 1986; 19876; 1989).
In the present study, two types of control procedures wereused to substantiate specific immunocytochemical staining.Staining was absent when non-immune rat serum replaced theanti-human C-peptide, and when either the anti-human C-peptide, or the goat anti-rat IgG conjugated to peroxidase, orhydrogen peroxide was omitted from the immunocytochemi-cal procedure. This demonstrates the absence of electrostaticand hydrophobic binding of immunocytochemical reagents totissues (Leduque et al. 1987a). In addition, no cellsimmunoreactive to anti-human C-peptide antibody weredetected in control nontransgenic mouse pancreas.

Polyclonal origin of pancreatic islets 1117
Results
Obtention of aggregation chimaeras56 aggregation embryos were obtained and transferredinto foster mothers. Five foster females gave rise to 18newborn (Table 1). Six (of which five were males) had atypical chimaeric coat. The (C57BL/6xDBA/2)F2mouse coat component was brown, black, or grey. Tg74component was white, since Tg74 have an albino SJL/Jgenetic background. Red and black mosaic iris was alsofound in one chimaera. The Tg74 mouse contributionwas further documented in these six chimaeras by twocriteria. First, the human proinsulin transgene wasdemonstrated by a tail DNA dot-blot assay, andsecond, human C-peptide was found in urine, using aradioimmunoassay. The latter assay indicated that thetransgenic embryo component participated in thepancreatic fi cells. Six other experimental mice had nowhite coat component. Among them, one (chimaera16) had the transgenic human DNA fragment in theDNA prepared from tails and human C-peptide inurine. Since it had the coat from one embryo and thehuman transgene from the other, it was considered as atrue chimaera. The six experimental albino mice werenot analyzed further by other markers for demonstrat-ing the presence of any (C57BL/6xDBA/2)F2 com-ponent, and were discarded. In total, 7 chimaeric micewere identified, of which one did not have coat colourchimaerism (Table 1). Four of them were used forimmunocytochemistry analysis of the pancreas.
Chimaera 11Staining with anti-human C-peptide antibody allowed
Table 1. Development of aggregation chimaerasNumber ofaggregatestransferred
Number ofrecipients
Number oflive newborns
Number ofchimaeras
56 18
* Six animals had a white/pigmented chimaeric coat and thehuman transgene, and one had no white coat component but hadthe human transgene and human C-peptide. Among the elevenmice that were not considered, five had pigmented coats but thehuman transgene was not demonstrated in tail DNA, and six hada white coat.
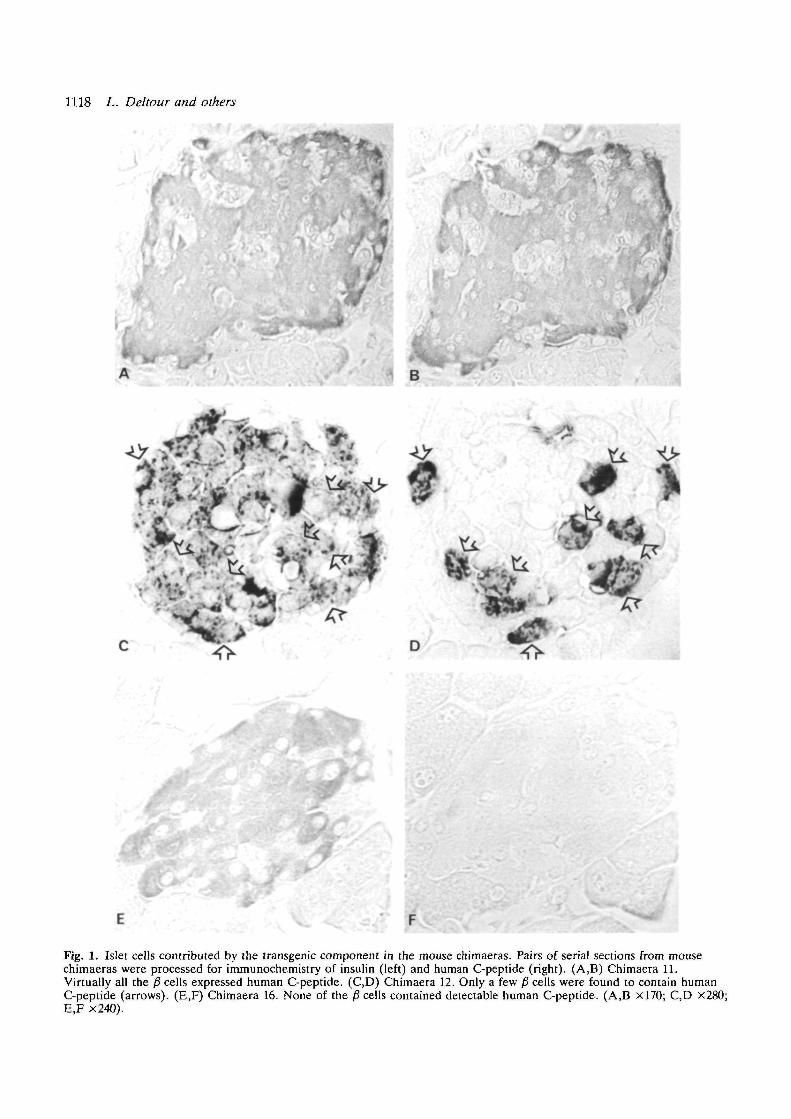
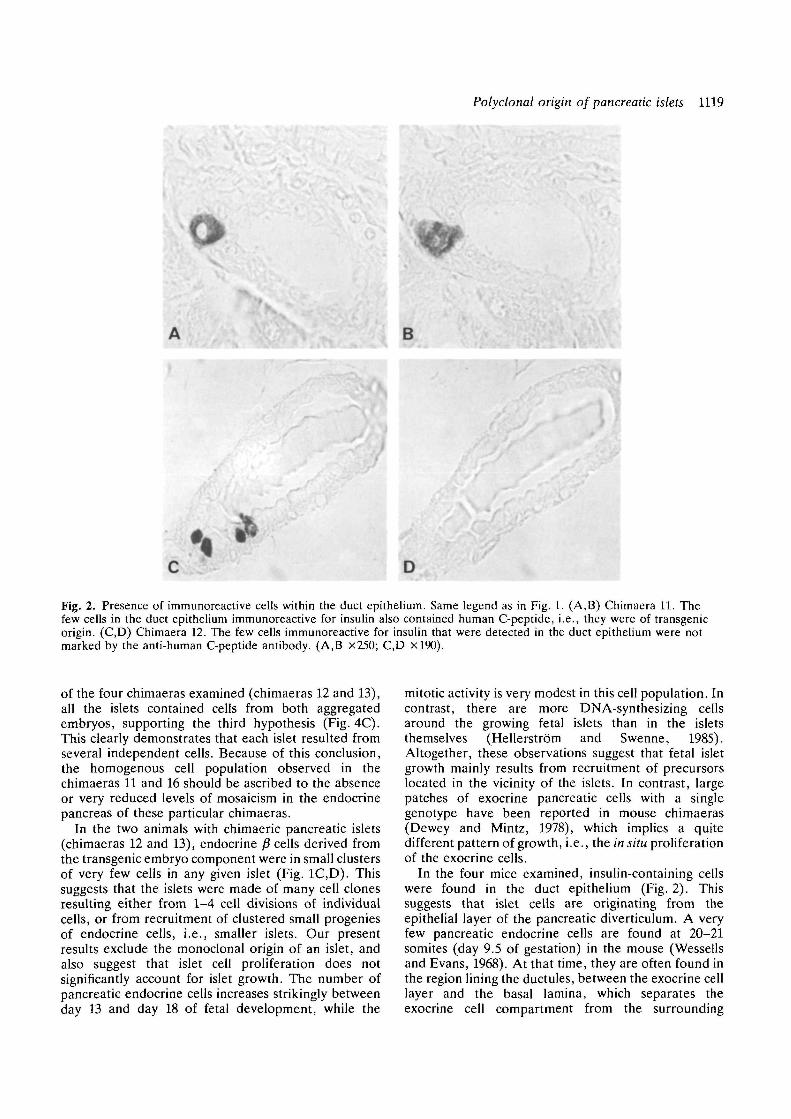
identification of the p1 cells derived from the transgenicembryo. Human C-peptide was found in all the fi cellsof all the islets examined (Fig. 1A,B; Table 2). Pres-ence of endocrine p" cells was occasionally observed inthe duct epithelium by systematic screening of serialsections. These fi cells contained also human C-peptide(Fig. 2A.B).
Chimaeras 12 and 13Human C-peptide immunoreactivity was found in allthe pancreatic islets. Each of the islets examined (238out of 241) was a mosaic of fi cells, which containedhuman C-peptide, and fi cells, which did not(Fig. 1C,D; Table 2). In all the islets, the £ cells derivedfrom the transgenic embryo were only a minority.These cells were associated in small clusters scatteredwithin the islets (Fig. ID). Some fi cells were detectedin the duct epithelium (Fig. 2C), but they were notimmunoreactive for human C-peptide (Fig. 2D).
Chimaera 16We have not been able to detect any human C-peptideimmunoreactivity in any fi cells (Fig. 1E,F; Table 2).Some fi cells were detected in the duct epithelium, andhuman C-peptide was never observed in these cells (notillustrated).
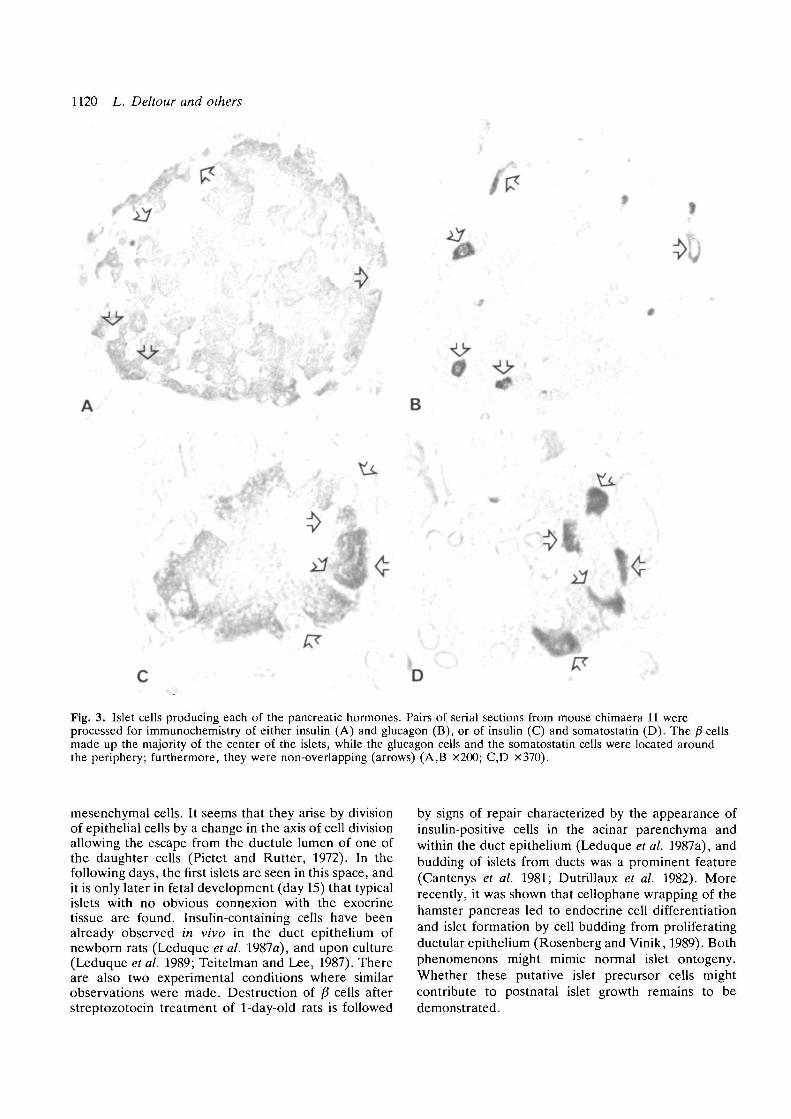
Serial sections of pancreases were stained with anti-insulin, anti-glucagon and anti-somatostatin antibodiesin order to discriminate the p", a, and 6 cells in the islets,respectively (Fig. 3).
Discussion
In the present study, we have examined the origin andthe growth of the pancreatic islets, not to determine thedevelopmental lineage of endocrine islet cells (seereview in Le Douarin, 1988), but to establish whether(1) a single progenitor cell gave rise to all the precursorsof the islets, or (2) if each of a few progenitor cells is thefounder of a different islet, or (3) if each islet is amixture of cells originating from a pool of progenitorcells (Fig. 4). According to the first two hypotheses, oneexpects to find in mouse chimaeras homogenous isletswith only a single cell type, originating from one or theother embryo. According to the third one, individual
Chimaera
11121316
* Presence of humant Presence of human
Coat colour
chimaencchimaericchimaeric
brown
insulin gene (HuC-peptide (HCP)
Table 2. Mosaicism
Hu DNA» HCPt
+ 250+ 100+ 50+ 10, 20
DNA), by tail DNA dot-blot,in urine, by RIA (pgml"1).
of pancreatic
examined
22514299
273
isletsNumber
mosaic
0140980
of islets
homogenous
HCP+
225210
HCP"
000
273
1118 L. Deltour and others
c
E
1
-.' fs
'••-
B - •
1>
Fig. 1. Islet cells contributed by the transgenic component in the mouse chimaeras. Pairs of serial sections from mousechimaeras were processed for immunochemistry of insulin (left) and human C-peptide (right). (A,B) Chimaera 11.Virtually all the /3 cells expressed human C-peptide. (C,D) Chimaera 12. Only a few fi cells were found to contain humanC-peptide (arrows). (E,F) Chimaera 16. None of the /3 cells contained detectable human C-peptide. (A,B xl70; C,D x280;E,F x240).
Polyclonal origin of pancreatic islets 1119
B
C D
Fig. 2. Presence of immunoreactive cells within the duct epithelium. Same legend as in Fig. 1. (A,B) Chimaera 11. Thefew cells in the duct epithelium immunoreactive for insulin also contained human C-peptide, i.e., they were of transgenicorigin. (C,D) Chimaera 12. The few cells immunoreactive for insulin that were detected in the duct epithelium were notmarked by the anti-human C-peptide antibody. (A,B x250; C,D xl90).
of the four chimaeras examined (chimaeras 12 and 13),all the islets contained cells from both aggregatedembryos, supporting the third hypothesis (Fig. 4C).This clearly demonstrates that each islet resulted fromseveral independent cells. Because of this conclusion,the homogenous cell population observed in thechimaeras 11 and 16 should be ascribed to the absenceor very reduced levels of mosaicism in the endocrinepancreas of these particular chimaeras.
In the two animals with chimaeric pancreatic islets(chimaeras 12 and 13), endocrine /3 cells derived fromthe transgenic embryo component were in small clustersof very few cells in any given islet (Fig. 1C,D). Thissuggests that the islets were made of many cell clonesresulting either from 1-4 cell divisions of individualcells, or from recruitment of clustered small progeniesof endocrine cells, i.e., smaller islets. Our presentresults exclude the monoclonal origin of an islet, andalso suggest that islet cell proliferation does notsignificantly account for islet growth. The number ofpancreatic endocrine cells increases strikingly betweenday 13 and day 18 of fetal development, while the
mitotic activity is very modest in this cell population. Incontrast, there are more DNA-synthesizing cellsaround the growing fetal islets than in the isletsthemselves (Hellerstrom and Swenne, 1985).Altogether, these observations suggest that fetal isletgrowth mainly results from recruitment of precursorslocated in the vicinity of the islets. In contrast, largepatches of exocrine pancreatic cells with a singlegenotype have been reported in mouse chimaeras(Dewey and Mintz, 1978), which implies a quitedifferent pattern of growth, i.e., the in situ proliferationof the exocrine cells.
In the four mice examined, insulin-containing cellswere found in the duct epithelium (Fig. 2). Thissuggests that islet cells are originating from theepithelial layer of the pancreatic diverticulum. A veryfew pancreatic endocrine cells are found at 20-21somites (day 9.5 of gestation) in the mouse (Wessellsand Evans, 1968). At that time, they are often found inthe region lining the ductules, between the exocrine celllayer and the basal lamina, which separates theexocrine cell compartment from the surrounding
1120 L. Deltour and others
4
•
'd"
*
B
•
-v
Fig. 3. Islet cells producing each of the pancreatic hormones. Pairs of serial sections from mouse chimaera 11 wereprocessed for immunochemistry of either insulin (A) and glucagon (B), or of insulin (C) and somatostatin (D). The /? cellsmade up the majority of the center of the islets, while the glucagon cells and the somatostatin cells were located aroundthe periphery; furthermore, they were non-overlapping (arrows) (A,B X200; C,D x370).
mesenchymal cells. It seems that they arise by divisionof epithelial cells by a change in the axis of cell divisionallowing the escape from the ductule lumen of one ofthe daughter cells (Pictet and Rutter, 1972). In thefollowing days, the first islets are seen in this space, andit is only later in fetal development (day 15) that typicalislets with no obvious connexion with the exocrinetissue are found. Insulin-containing cells have beenalready observed in vivo in the duct epithelium ofnewborn rats (Leduque et al. 1987a), and upon culture(Leduque et al. 1989; Teitelman and Lee, 1987). Thereare also two experimental conditions where similarobservations were made. Destruction of p cells afterstreptozotocin treatment of 1-day-old rats is followed
by signs of repair characterized by the appearance ofinsulin-positive cells in the acinar parenchyma andwithin the duct epithelium (Leduque et al. 1987a), andbudding of islets from ducts was a prominent feature(Cantenys et al. 1981; Dutrillaux et al. 1982). Morerecently, it was shown that cellophane wrapping of thehamster pancreas led to endocrine cell differentiationand islet formation by cell budding from proliferatingductular epithelium (Rosenberg and Vinik, 1989). Bothphenomenons might mimic normal islet ontogeny.Whether these putative islet precursor cells mightcontribute to postnatal islet growth remains to bedemonstrated.

Polydonal origin of pancreatic islets 1121
Fig. 4. Schematic representation of the possible expansionof the cells from which the islet (3 cells derive. (A) A singleprogenitor cell gives rise to all the precursors of all theislets. (B) There are several independent progenitor cellsfor the /3 cells, but a single progenitor gives rise to all theP cells in an islet. (C) There are several progenitor cells,and mixed progenies make up each islet. The resultsobtained with chimaeras 12 and 13 are relevant to thelatter hypothesis.
We thank O. Madsen for the anti-human C-peptidemonoclonal antibody, M. A. Root and B. H. Frank for theanti-human C-peptide antiserum. A. Paldi was a recipient of afellowship from the Institut National de la Sant6 et de laRecherche M6dicale. This work was supported by grants fromthe Fondation pour la Recherche M6dicale, the LigueNationale contre le Cancer, the Ddipartement de BiologieHumaine, University Claude Bernard Lyon I (985-387), andthe Centre National de la Recherche Scientifique.
References
BUCCHINI, D., MADSEN, O., DESBOIS, P., PICTET, R. AND JAMI, J(1989). B islet cells of pancreas are the site of expression of thehuman insulin gene in transgenic mice. Expl Cell Res. 180,467^74.
BUCCHINI, D., RIPOCHE, M-A., STINNAKRE, M-G., DESBOIS, P.,LORES, P., MONTHIOUX, E., ABSIL, J., LEPESANT, J-A., PICTET,R. AND JAMI, J. (1986). Pancreatic expression of human insulingene in transgenic mice. Proc. nam. Acad. Sci. U.S.A. 83,2511-2515.
CANTENYS, D., PORTHA, B., DUTRILLAUX, M. C , HOLLANDE, E.,ROZE, C. AND PICON, L. (1981). Histogenesis of the endocrinepancreas in newborn rats after destruction by streptozotocin. Animmunocytochemical study. Virchows Arch. B 35, 109-122.
DEWEY, M. J. AND MINTZ, B. (1978). Genetic control of cell-type-specific levels of mouse /J-galactosidase. Devi Biol. 66, 550-559.
DUTRILLAUX, M. C , PORTHA, B., ROZE, C. AND HOLLANDE, E.(1982). Ultrastructural study of pancreatic B cell regeneration innewborn rats after destruction by streptozotocin. VirchowsArch. B 39, 173-185.
FROMONT-RACINE, M., BUCCHINI, D. , MADSEN, O., DESBOIS, P.,LINDE, S., NIELSEN, J. H., RIPOCHE, M-A., JAMI, J. AND PICTET,R (1990). Effect of 5'-flanking sequence deletions on expressionof the human insulin gene in transgenic mice. Molec.Endocrinol. 4, 669-677.
GRAHAM, R. C. AND KARNOVSKY, M. (1966). The early stages ofabsorption of injected horseradish peroxidase in the proximaltubules of mouse kidney: ultrastructural cytochemistry by a newtechnique. J. Hisiochem. Cytochem. 14, 291-302
HELLERSTROM, C. AND SWENNE, I. (1985). Growth pattern ofpancreatic islets in animals. In The Diabetic Pancreas (eds. B.W. Volk and E. R. Arquilla) pp. 53-79. New York: PlenumPublishing Corporation.
HOGAN, B., COSTANTINI, F. AND LACY, E. (1986). Manipulating the
Mouse Embryo. A Laboratory Manual, pp. 332. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory.
LE DOUARJN, N. M. (1988). On the origin of pancreatic endocrinecells. Cell 53, 169-171.
LEDUQUE, P., ARATAN-SPIRE, S., CZERNICHOW, P. AND DUBOIS, P.M. (1986). Ontogenesis of thyrotropin-releasing hormone in thehuman fetal pancreas. / . elm. Invest. 78, 1028-1034.
LEDUQUE, P., ARATAN-SPIRE, S., SCHARFMANN, R.,BASMACIOGULLARJ, A., CZERNICHOW, P. AND DUBOIS, P. M.(1989). Coexistence of thyrotropin-releasing hormone and insulinin culture fetal rat islets: a light and electron microscopicimmunocytochemical study during islet neoformation. Biol. Cell66, 291-296.
LEDUQUE, P., ARATAN-SPIRE, S., WOLF, B., DUBOIS, P. M. ANDCZERNICHOW, P. (1987a). Localization of thyrotropin-releasinghormone and insulin-immunoreactivity in the pancreas ofneonatal rats after injection of streptozotocin at birth. CellTissue Res. 248, 89-94.
LEDUQUE, P., JACKSON, I. M. D., KERVRAN, A., ARATAN-SPIRE, S.,CZERNICHOW, P. AND DUBOIS, P. M. (19876). Histidyl-prolinediketopiperazine (His-Pro-DKP) is present in the glucagon-containing cells of the human fetal pancreas. J. chn. Invest. 79,875-880.
LEDUQUE, P., WOLF, B., ARATAN-SPIRE, S., DUBOIS, P. M. ANDCZERNICHOW, P. (1985). Immunocytochemical location ofthyrotropin-releasing hormone (TRH) in the B-cells of adulthypothyroid rat pancreas. Regul. Pepttdes 10, 281-292.
MADSEN, O. D., COHEN, R. M., FITCH, F. W., RUBENSTEIN, A. H.AND STEINER, D. F. (1985). The production and characterizationof monoclonal antibodies specific for human proinsulin using asensitive microdot assay procedure. Endocrinology 113,2135-2144.
MICHALOVA, K., BUCCHINI, D., RIPOCHE, M-A., PICTET, R. ANDJAMI, J. (1988). Chromosome localization of the human insulingene in transgenic mouse lines. Human Genet. 80, 247-252.
NAKANE, P. K. AND PIERCE, G. B. (1966). Enzyme-labeledantibodies: preparation and application for the localization ofantigens. J. Histochem. Cytochem 14, 929-93.
PICTET, R. AND RUTTER, W. J. (1972). Development of theembryonic endocrine pancreas. In Handbook of Physiology(eds. D. Steiner and N. Freinkel) vol. 1, pp. 25-66. Baltimore,MD: Williams and Wilkins.
ROSENBERG, L. AND VINIK, A. I. (1989). Induction of endocrinecell differentiation: a new approach to management of diabetes.7 Lab. Clin. Med. 114, 75-83.
RUGH, R. (1968). The Mouse. Its Reproduction and Development.pp. 430. Minneapolis, MN: Burgess Publishing Company.
TEITELMAN, G. AND LEE, J. K. (1987). Cell lineage analysis ofpancreatic islet cell development: glucagon and insulin cells arisefrom catecholaminergic precursors present in the pancreaticduct. Devi Biol. 121, 454-466.
THOMPSON, E. M., FLEMING, K. A., EVANS, D. J., FUNDELE, R.,SURANI, M. A. AND WRIGHT, N. A. (1990). Gastric endocrinecells share a clonal origin with other gut cell lineages.Development 110, 477-481.
WESSELLS, N. K. AND EVANS, J. (1968). Ultrastructural studies ofearly morphogenesis and cytodifferentiation in the embryonicmammalian pancreas. Devi Biol. 17, 413-446.
WILLIAMS, R. L., HILTON, D. J., PEASE, S., WILLSON, T. A.,STEWART, C. L., GEARING, D. P., WAGNER, E. F., METCALF, D.,NICOLA, N. A. AND GOUGH, N. M. (1988). Myeloid leukaemiainhibitory factor maintains the developmental potential ofembryonic stem cells. Nature 336, 684-687.
(Accepted 31 May 1991)