Embed Size (px)
Citation preview

Plant Physiol. (1991) 96, 1053-10590032-0889/91/96/1 053/07/$01 .00/0
Received for publication October 31, 1990Accepted March 9, 1991
Polyamines in Plants Infected by Citrus Exocortis Viroid orTreated with Silver Ions and Ethephon'
Jose M. Belles, Juan Carbonell, and Vicente Conejero*Laboratorio de Bioquimica y Biologia Molecular, Departamento de Biotecnologia, Universidad Polit6cnica de
Valencia, 46022-Valencia, Spain (J.M.B., V.C.); and Instituto de Agroquimica y Tecnologia de Alimentos del CSIC,Jaime Roig 11, 46010 Valencia, Spain (J.C.)
ABSTRACT
The levels of polyamines in leaves of Gynura aurantiaca DCand tomato, Lycoperskon esculentum Mill. cv Rutgers, infectedwith citrus exocortis viroid (CEVd) or treated with silver nitrate orethephon (2-chloroethylphosphonic acid) were measured byHPLC in relation to development of symptoms. Previously it hadbeen demonstrated that treatment of G. aurantiaca plants withsilver nitrate or ethephon closely mimicked the effects of viroidinfection in the plants. In the studies reported here, a markeddecrease in putrescine level was observed in plants infected byCEVd or treated with silver ions or ethephon. There was nosignificant change in either spermidine or spermine levels. Treat-ment of G. aurantiaca plants with specific inhibitors of ethylenebiosynthesis (aminoethoxyvinylglycine, Co2+) or ethylene action(norbomadiene) prevented the decrease of putrescine associ-ated with silver nitrate treatment and had no effect on spermidineor spermine levels. The development of viroid-like symptoms, theproduction of associated pathogenesis-related proteins, and therise in protease activity induced by silver nitrate, were all sup-pressed by exogenous application of putrescine. The decreasedlevel of putrescine as an ethylene-mediated step in the transduc-tion of the viroid and silver or ethephon signaling is discussed.
Viroids are single-stranded, small, circular RNA molecules(about 105 D), which, despite their limited coding capacity,are able to replicate without the aid of a helper virus in hostcells and produce developmental distortions in certain hostplants (15, 27).CEVd2, the causal agent of the exocortis disease of citrus
(28), can be transmitted to Gynura aurantiaca DC and tomato(Lycopersicon esculentum Mill. cv Rutgers) plants. The infec-tion of the plants by CEVd induces changes in plant growthand development, including stunting and leaf epinasty andrugosity. Also, the impairment of the capacity of cuttings toform roots has been described (19). Viroid infection alsocauses an increased production of ethylene in the host plants(3-5, 10) and the enhanced accumulation of the so-called PRproteins (6, 11, 23). Interestingly, it has been shown that
'This work has been supported by grant PB87-0662 from Comi-si6n Interministerial de Ciencia y Tecnologia and Diputaci6n Provin-cial de Valencia.
2 Abbreviations: CEVd, citrus exocortis viroid; PR, pathogenesis-related; AVG, aminoethoxyvinylglycine; FTC-, fluorescein thiocar-bamoyl derivative.
treatment of Gynura plants with AgNO3 produces similareffects to those incited by CEVd infection, including anincreased ethylene production (10). The viroid-like effects ofsilver ion has been attributed to its capacity to elicit ethylenebiosynthesis in Gynura plants (10, 14). Analogous effects areproduced by ethephon (2-chloroethylphosphonic acid, an eth-ylene releasing compound) (10). The viroid-like syndromeinduced by AgNO3 in Gynura plants was prevented (4) byinhibitors of ethylene biosynthesis (aminooxyacetic acid orCo2+) (39, 40) and ethylene action (norbornadiene) (30).Based on those findings, a view of viroid pathogenesis, inwhich viroid molecules are considered as pure replicativesignals, has been proposed (14). In this hypothesis, viroids areelicitors of a general, ethylene-mediated response of the hostplant.Polyamines have been implicated in a range ofphysiological
plant responses (20, 22). The possible involvement of polya-mines in the transduction of the viroid pathogenic signalinghas also been hypothesized (14). The present work is the firstexperimental approach to test this hypothesis. As was donepreviously (4), we have used the Gynura-silver interactions asa model system to study the possible involvement of polya-mines in the transduction of the plant response to differenttypes of stress. This system has a number of advantagescompared to host-viroid interactions: (a) quickness of theresponse, i.e. epinasty of the leaves and the production ofPRproteins (P1 and P2) are observed within 48 h in silver-treatedplants, while it takes 15 d in CEVd-infected plants. Conse-quently, (b) the effects ofputrescine and inhibitors ofethylenebiosynthesis or action on the plant pathogenic reaction canbe studied without the need for repeated applications over aprolonged time, which could mask the effects due to nonspe-cific toxicity. We have focused our attention on the levels ofputrescine, spermidine, and spermine, the most widespreadpolyamines in plant tissues, in CEVd-infected or silver- andethephon-treated plants as compared with uninfected controlplants; the effect of inhibitors of ethylene biosynthesis oraction on polyamine levels; and the effect of exogenous ap-plication of polyamines on the viroid-like syndrome andprotease activity induced by treatments with silver ions.
MATERIALS AND METHODS
Plant Material
Plants of Gynura aurantiaca DC were grown from rootedcuttings. Cuttings (7 cm) were obtained from matched blocks
1053
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 96, 1991
of healthy Gynura plants and put into a rooting chamber for15 d and then were transplanted in pots. Tomato (Lycoper-sicon esculentum Mill. cv Rutgers) plants were grown fromseeds. The plants (one per pot) were grown in a greenhouseat 30C/24°C (day/night) (RH 70%) in 20-cm-diameter potscontaining a mix of two parts peat and one part grit/sand.The pots were watered twice a week to saturation with Hoag-land solution. Gynura plants were inoculated 15 d after trans-planting with either buffer or the 2 M LiCl-soluble fraction ofnucleic acids from CEVd-infected Gynura tissue as describedpreviously (29). Inoculation of tomato was carried out bypuncturing the stems of the seedlings with a needle dipped ineither buffer or the nucleic acid preparation. Symptoms ap-peared 15 to 17 d after inoculation in both Gynura and tomatoplants.
Plant Treatments
Healthy Gynura or tomato plants, equivalent in age toCEVd-infected when commencing the expression of theirsymptoms, were utilized in these experiments. Treatmentswith AgNO3 (3 mM), ethephon (2-chloroethylphosphonicacid) (30 mM) as an ethylene generator, AVG (50 ALM), CoSO4(200 Mm), norbornadiene (5 mM), and putrescine (1 mM) wereperformed as described elsewhere (4). Briefly, individualplants were sprayed until run-off with 10 mL of the appropri-ate solution (pH adjusted to 5.5) containing 0.1% (v/v) Tween80 as a wetting agent. Plants were left in a shaded area for afew hours before returning to the greenhouse. Control plantswere sprayed with water containing 0.1% (v/v) Tween 80.
Assay of Proteolytic Activity
Apical leaves (apex + four youngest leaves) were frozen inliquid nitrogen, thawed, and homogenized with 3 mL/g cold50 mm phosphate-citrate buffer (pH 6.6) supplemented with20 mm KCl, 2 mM MgCl2, 5% sucrose, and 15 mM 2-mercap-toethanol. The homogenate was filtered through cheeseclothand centrifuged at 30,000g for 30 min at 4°C. The resultingclarified supernatant was used to determine protease activity.
Protease activity was measured with FTC-casein as sub-strate as described by Vera and Conejero (34). The reactionmixture (total volume of 150 ,uL) contained 20 ,L of enzyme,120 gL assay buffer, and 10 gL of 0.5% (w/v) FTC-casein.The assay buffer was 150 mm phosphate-citrate buffer (pH6.6) containing 0.5 mm DTT. The solution was thoroughlymixed and incubated at 37°C for 1 h. The reaction wasstopped by addition of 150 ML of 10% (w/v) TCA. Thetubes were allowed to stand on ice for 1 h, and the TCA-insoluble material was sedimented by centrifugation for 10min at maximum speed in a Beckman microfuge. A l00-uLaliquot of the supernatant was diluted to 3 mL with 500mM Tris-HCl (pH 8.5). Fluorescence was determined with anexcitation wavelength of 490 nm and an emission wave-length of 525 nm in a Perkin-Elmer 650-1OS fluorescencespectrophotometer.Enzyme activity was linear with time and with concentra-
tion of crude extracts. One unit of FTC-casein-degradingactivity is defined as the enzyme needed to produce 1 unit offluorescence increase under the standard assay conditions.Enzyme activity is expressed as units/mg protein/h.
Protein Extraction and Electrophoretic Analysis
Apical leaves were harvested 48 h after the treatment withAgNO3. Soluble proteins were extracted as previously de-scribed (13) with some modifications. Leaves were frozen inliquid nitrogen, thawed, and homogenized with 3 mL/g cold20 mm Tris-HCI buffer (pH 7.4) supplemented with 20 mMKC1, 2 mM MgCl2, 5% sucrose, and 15 mM 2-mercaptoetha-nol. The homogenate was filtered through cheesecloth andcentrifuged at l00,OOOg for 2 h. Fractions of the supernatantwere used for electrophoresis according to the procedure ofConejero and Semancik (12).
Extraction of Polyamines
Extraction of soluble polyamines was based on the methoddescribed by Flores and Galston (18). Apical leaves werehomogenized with a pestle in a chilled mortar in 0.2 Mperchloric acid (4 mL/g tissue) containing 1,6-diaminohexaneas internal standard. After extraction for 1 h in an ice bath,homogenates were centrifuged at 48,000g for 30 min.
Benzoylation and HPLC Analysis
Apical leaf extracts were benzoylated according to Floresand Galston (18). Samples were ultrafiltered through Milli-pore (Molsheim, France) HV filters (0.45 ,m) and analyzedimmediately. HPLC analysis was done with a Waters model510 liquid chromatograph. The solvent system consisted ofmethanol:water run isocratically at 64% methanol at a flowrate of 1 mL/min. Benzoylated samples were eluted at roomtemperature through a C18 reverse phase column (4 x 250mm, 5-,gm particle size, Merck RT), and detected at 254 nmwith a UV-detector (Waters model 440). The results werequantified with a Hewlett-Packard 3396 A integrator using arelative calibration procedure described by Smith and Davies(33) for dansyl derivatives. With this procedure, peak arearatios of individual polyamines to the internal standard, 1,6-diaminohexane, were linearly correlated with the weight(nmol) ratios. With benzoyl-1,6-diaminohexane as internalstandard, linear calibration curves were obtained up to 100,M (putrescine: y= 0.682x, spermidine: y= 1.205x, spermine:y = 1 .548x). Identity of polyamines was determined by reten-tion time and coinjection of a known standard. The polya-mine breakdown product, 1,3-diaminopropane, has a reten-tion time very close to that of putrescine. However, changingthe solvent to 60% methanol (18) allowed for precise discrim-ination and estimation of 1,3-diaminopropane and putrescinein the extracts and demonstrated that the observed peak wasputrescine and not 1,3-diaminopropane.The data presented in the "Results" are for single experi-
ments with triplicate samples and are representative ofa groupof three or more experiments.
RESULTS
Polyamine Alteration Induced by CEVd-Infection andSilver or Ethephon Treatment
Polyamine levels were followed during 16 d in healthy(uninfected) and CEVd-infected Gynura and tomato plants
BELLES ET AL.1 054
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
POLYAMINES IN HOST-VIROID INTERACTIONS
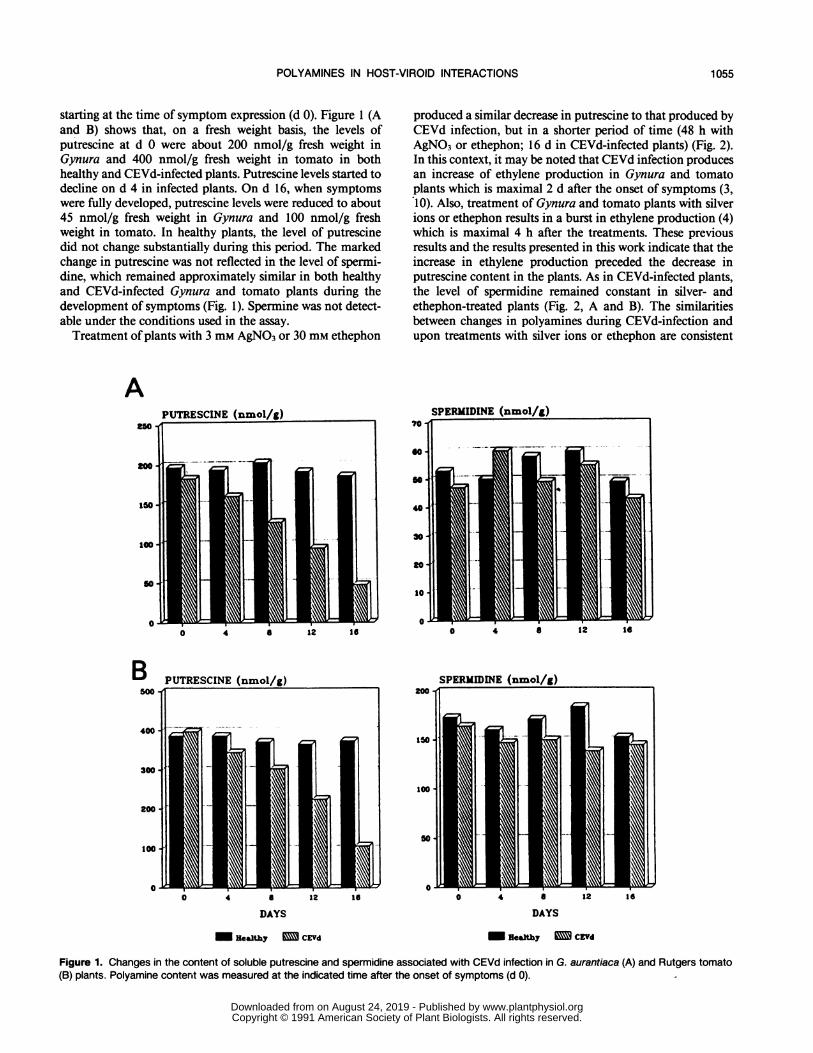
starting at the time of symptom expression (d 0). Figure 1 (Aand B) shows that, on a fresh weight basis, the levels ofputrescine at d 0 were about 200 nmol/g fresh weight inGynura and 400 nmol/g fresh weight in tomato in bothhealthy and CEVd-infected plants. Putrescine levels started todecline on d 4 in infected plants. On d 16, when symptomswere fully developed, putrescine levels were reduced to about45 nmol/g fresh weight in Gynura and 100 nmol/g freshweight in tomato. In healthy plants, the level of putrescinedid not change substantially during this period. The markedchange in putrescine was not reflected in the level of spermi-dine, which remained approximately similar in both healthyand CEVd-infected Gynura and tomato plants during thedevelopment of symptoms (Fig. 1). Spermine was not detect-able under the conditions used in the assay.Treatment of plants with 3 mM AgNO3 or 30 mM ethephon
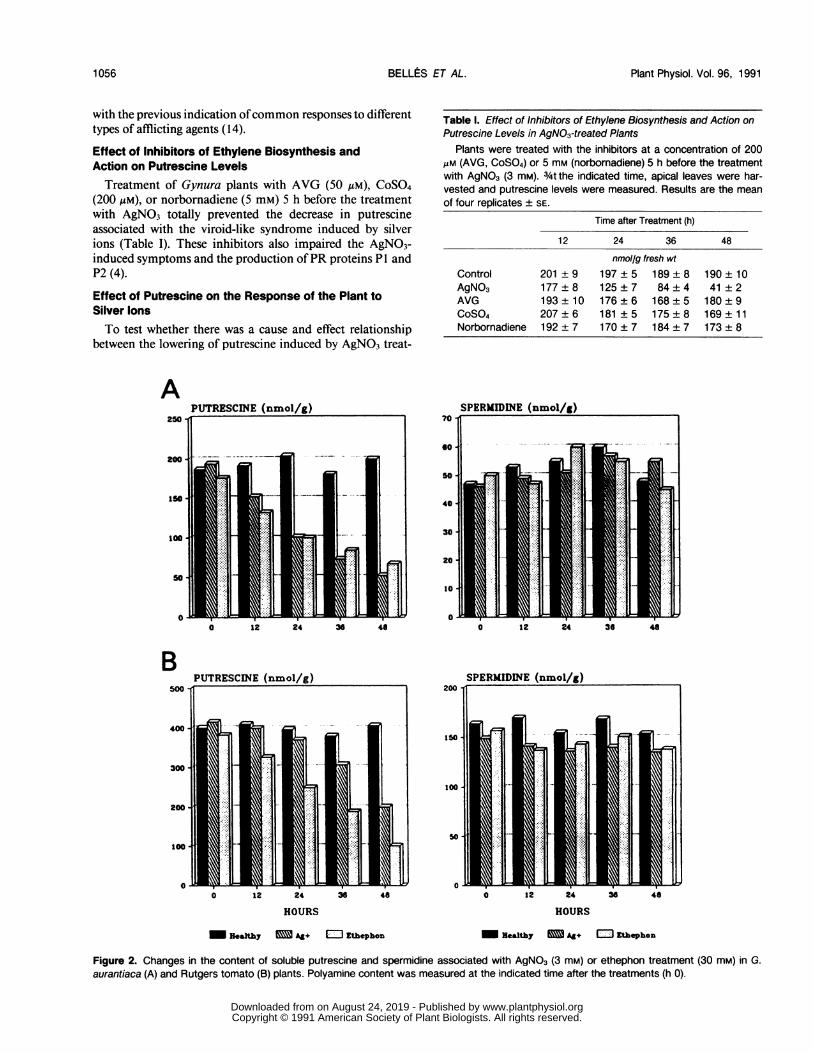
produced a similar decrease in putrescine to that produced byCEVd infection, but in a shorter period of time (48 h withAgNO3 or ethephon; 16 d in CEVd-infected plants) (Fig. 2).In this context, it may be noted that CEVd infection producesan increase of ethylene production in Gynura and tomatoplants which is maximal 2 d after the onset of symptoms (3,10). Also, treatment of Gynura and tomato plants with silverions or ethephon results in a burst in ethylene production (4)which is maximal 4 h after the treatments. These previousresults and the results presented in this work indicate that theincrease in ethylene production preceded the decrease inputrescine content in the plants. As in CEVd-infected plants,the level of spermidine remained constant in silver- andethephon-treated plants (Fig. 2, A and B). The similaritiesbetween changes in polyamines during CEVd-infection andupon treatments with silver ions or ethephon are consistent
A250
200
150
50-
0
BNMO -
400 -
300-
250
100
0'
SPERMIDINE (nmol/g)
60 -
40
30
20O
10
0
200
150
150
50
0o
0 4 a 12 le
4 a 12 is 0 4 a 12
DAYS DAYS
_ Healthy M CEVd _ Beastly = CEYd
Figure 1. Changes in the content of soluble putrescine and spermidine associated with CEVd infection in G. aurantiaca (A) and Rutgers tomato(B) plants. Polyamine content was measured at the indicated time after the onset of symptoms (d 0).
---
1055
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 96, 1991
with the previous indication ofcommon responses to differenttypes of afflicting agents ( 14).
Effect of Inhibitors of Ethylene Biosynthesis andAction on Putrescine LevelsTreatment of Gynura plants with AVG (50 jiM), CoS04
(200 gM), or norbornadiene (5 mM) 5 h before the treatmentwith AgNO3 totally prevented the decrease in putrescineassociated with the viroid-like syndrome induced by silverions (Table I). These inhibitors also impaired the AgNO3-induced symptoms and the production ofPR proteins PI andP2 (4).
Effect of Putrescine on the Response of the Plant toSilver IonsTo test whether there was a cause and effect relationship
between the lowering of putrescine induced by AgNO3 treat-
A250
200
150
t00
50
0 12 24 36 48
BF
500 -l
400
300 -
200 -
100
PUTRESCINE (nmol/g)
0 12 24 36 46
HOURS
_ Healthy M As+ ED Ethephon
Table I. Effect of Inhibitors of Ethylene Biosynthesis and Action onPutrescine Levels in AgNO3-treated Plants
Plants were treated with the inhibitors at a concentration of 200JLM (AVG, CoS04) or 5 mm (norbornadiene) 5 h before the treatmentwith AgNO3 (3 mM). 3/4t the indicated time, apical leaves were har-vested and putrescine levels were measured. Results are the meanof four replicates ± SE.
Time after Treatment (h)
12 24 36 48
nmol/g fresh wtControl 201 ± 9 197 ± 5 189 ± 8 190 ± 10AgNO3 177 ± 8 125 ± 7 84 ± 4 41 ± 2AVG 193 ± 10 176 ± 6 168 ± 5 180 ± 9CoS04 207 ± 6 181 ± 5 175 ± 8 169 ± 11Norbornadiene 192 ± 7 170 ± 7 184 ± 7 173 ± 8
0 12 24 36 48
200
150
100
SO
0 12 24 M 46
HOURS
_ Healthy EM A+ Ml thephon
Figure 2. Changes in the content of soluble putrescine and spermidine associated with AgNO3 (3 mM) or ethephon treatment (30 mM) in G.aurantiaca (A) and Rutgers tomato (B) plants. Polyamine content was measured at the indicated time after the treatments (h 0).
BELLES ET AL.1 056
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
POLYAMINES IN HOST-VIROID INTERACTIONS
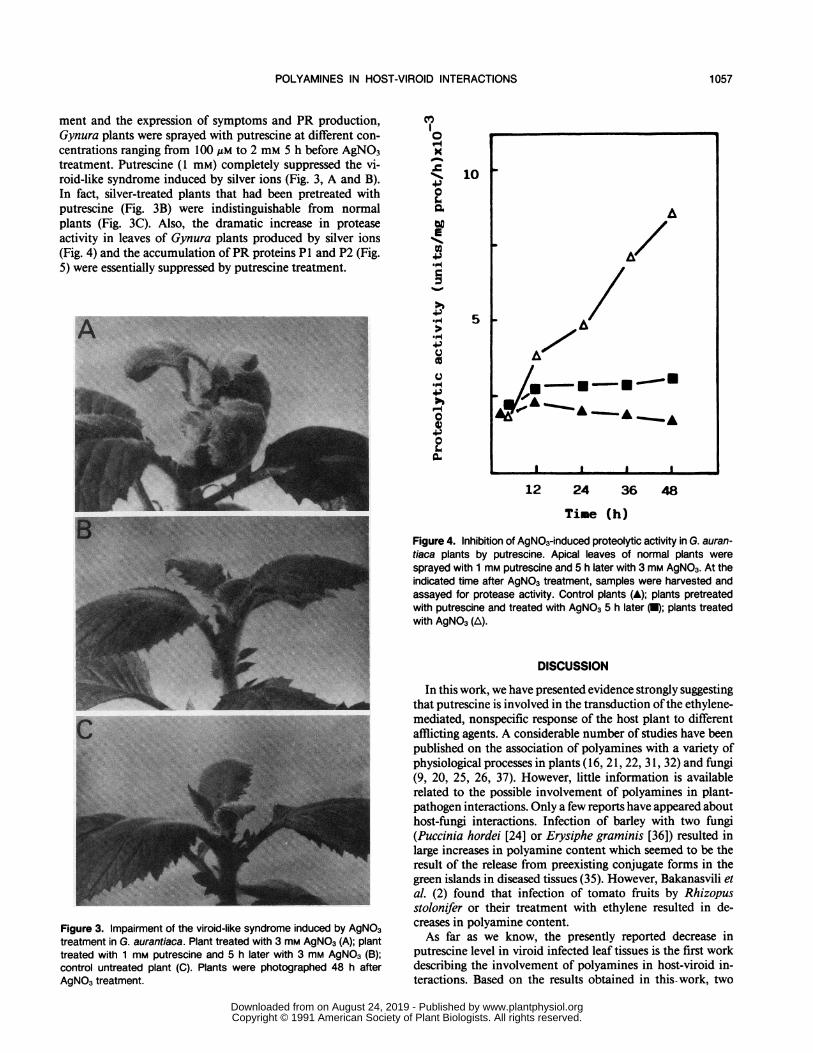
ment and the expression of symptoms and PR production,Gynura plants were sprayed with putrescine at different con-
centrations ranging from 100 JM to 2 mM 5 h before AgNO3treatment. Putrescine (1 mM) completely suppressed the vi-roid-like syndrome induced by silver ions (Fig. 3, A and B).In fact, silver-treated plants that had been pretreated withputrescine (Fig. 3B) were indistinguishable from normalplants (Fig. 3C). Also, the dramatic increase in proteaseactivity in leaves of Gynura plants produced by silver ions(Fig. 4) and the accumulation ofPR proteins P1 and P2 (Fig.5) were essentially suppressed by putrescine treatment.
Figure 3. Impairment of the viroid-like syndrome induced by AgNO3treatment in G. aurantiaca. Plant treated with 3 mm AgNO3 (A); planttreated with 1 mm putrescine and 5 h later with 3 mm AgNO3 (B);control untreated plant (C). Plants were photographed 48 h afterAgNO3 treatment.
12 24 36 48
Time (h)
Figure 4. Inhibition of AgNO3-induced proteolytic activity in G. auran-tiaca plants by putrescine. Apical leaves of normal plants weresprayed with 1 mm putrescine and 5 h later with 3 mm AgNO3. At theindicated time after AgNO3 treatment, samples were harvested andassayed for protease activity. Control plants (A); plants pretreatedwith putrescine and treated with AgNO3 5 h later (U); plants treatedwith AgNO3 (A).
DISCUSSION
In this work, we have presented evidence strongly suggestingthat putrescine is involved in the transduction ofthe ethylene-mediated, nonspecific response of the host plant to differentafflicting agents. A considerable number of studies have beenpublished on the association of polyamines with a variety ofphysiological processes in plants (16, 21, 22, 31, 32) and fungi(9, 20, 25, 26, 37). However, little information is availablerelated to the possible involvement of polyamines in plant-pathogen interactions. Only a few reports have appeared abouthost-fungi interactions. Infection of barley with two fungi(Puccinia hordei [24] or Erysiphe graminis [36]) resulted inlarge increases in polyamine content which seemed to be theresult of the release from preexisting conjugate forms in thegreen islands in diseased tissues (35). However, Bakanasvili etal. (2) found that infection of tomato fruits by Rhizopusstolonifer or their treatment with ethylene resulted in de-creases in polyamine content.As far as we know, the presently reported decrease in
putrescine level in viroid infected leaf tissues is the first workdescribing the involvement of polyamines in host-viroid in-teractions. Based on the results obtained in this-work, two
10 r
0
4-,
.,-
4-,
0
94
a-,
8co4a)
0
A
A
IL-LI
1 057
5 p
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 96, 1991
5
...- . .
conclusions can be supported. First, the enhancement ofethylene production is responsible either directly or throughother intermediary messenger signal(s) for the decrease inputrescine. This conclusion can be deduced from the fact thatspecific inhibitors ofeither ethylene biosynthesis (AVG, Co2")or its action (norbornadiene) prevent putrescine depletion. Ithas been reported (17) that ethylene and polyamines competefor S-adenosylmethionine, a precursor common to both eth-ylene and polyamine pathways (31). Interestingly, we havedemonstrated that norbornadiene prevents the depletion ofputrescine (Table I) without affecting ethylene synthesis (4).This indicates that the link between the increase in ethylenesynthesis (4) and the decrease in putrescine presented in thispaper is not due to metabolic competition for S-adenosyl-methionine. Second, the lowering of putrescine content is a
signaling step in the transduction chain leading to the plantresponse. This can be deduced from the finding that impedingthe decrease in the putrescine level through exogenous appli-cation of this compound prevents the silver-induced expres-sion of viroid-like symptoms (leaf malformations [Fig. 3]),inhibition of root growth (data not shown), the accumulationof PR proteins (Fig. 5), and the enhancement of proteolyticactivity (Fig. 4) associated with the plant response. Our con-
clusion that enhancement of ethylene production is respon-sible for the decrease in putrescine is consistent with the
finding that arginine decarboxylase, the main enzyme respon-sible for putrescine synthesis in plants (7, 8, 38), is inhibitedby ethylene at physiological concentrations (1).
LITERATURE CITED
1. Apelbaum A, Goldlust A, Icekson E (1985) Control by ethyleneof arginine decarboxylase activity in pea seedlings and itsimplication for hormonal regulation of plant growth. PlantPhysiol 79: 635-640
2. Bakanashvili M, Barkai-Golan R, Kopeliovitch E, Apelbaum A(1987) Polyamine biosynthesis in Rhizopus-infected tomatofruits: possible interaction with ethylene. Physiol Mol PlantPathol 31: 41-50
3. Belles JM, Conejero V (1989) Evolution of ethylene production,ACC and conjugated ACC levels accompanying symptomdevelopment in tomato and Gynura aurantiaca DC leavesinfected with citrus exocortis viroid (CEV). J Phytopathol 127:81-85
4. Belles JM, Conejero V (1989) Ethylene mediation of the viroid-like syndrome induced by Ag' ions in Gynura aurantiaca DCplants. J Phytopathol 124: 275-284
5. Belles JM, Duran-Vila N, Granell A, Conejero V (1989) ACCsynthesis as the activated step responsible for the rise of ethyl-ene production accompanying citrus exocortis viroid infectionin tomato plants. J Phytopathol 125: 275-284
6. Belles JM, Vera P, Duran-Vila N, Conejero V (1989) Ethyleneproduction in tomato cultures infected with the citrus exocortisviroid. Can J Plant Pathol 11: 256-262
7. Birecka H, Bitonti AJ, McCann PP (1985) Assaying ornithineand arginine decarboxylases in some plant species. Plant Phys-iol 79: 509-514
8. Birecka H, Bitonti AJ, McCann PP (1985) Activities of arginineand ornithine decarboxylases in various plant species. PlantPhysiol 79: 515-519
9. Birecka H, Garraway MO, Baumann RJ, McCann PP (1986)Inhibition of ornithine decarboxylase and growth ofthe fungusHelminthosporium maydis. Plant Physiol 80: 798-800
10. Conejero V, Granell A (1986) Stimulation of a viroid-like syn-drome and the impairment of viroid infection in Gynuraaurantiaca DC plants by treatment with silver ions. PhysiolMol Plant Pathol 29: 317-323
11. Conejero V, Semancik JS (1977) Exocortis viroid: alteration inthe proteins of Gynura aurantiaca accompanying viroid infec-tion. Virology 77: 221-232
12. Conejero V, Semancik JS (1977) Analysis ofthe proteins in crudeextracts by polyacrylamide slab gel electrophoresis. Phytopath-ology 66: 1424-1426
13. Conejero V, Picazo I, Segado P (1979) Protein alteration indifferent hosts following viroid infection. Virology 97: 454-456
14. Conejero V, Belles JM, Garcia-Breijo F, Garro R, Hernandez-Yago J, Rodrigo I, Vera P (1990) Signalling in Viroid Patho-genesis. In RSS Fraser, ed, Recognition and Response in Plant-Virus Interactions, Vol 41. Springer-Verlag, Berlin, pp 233-261
15. DienerTO (1979) Viroids and viroid diseases. Wiley Interscience,New York
16. Evans PT, Malmberg RL (1989) Do polyamines have roles inplant development? Annu Rev Plant Physiol Plant Mol Biol40: 235-269
17. Even-Chen Z, Mattoo AK, Goren R (1982) Inhibition of ethylenebiosynthesis by aminoethoxyvinylglycine and by polyaminesshunts label from 3,4-(Q4C)methionine into spermidine in agedorange peel discs. Plant Physiol 69: 385-388
18. Flores HE, Galston AW (1982) Analysis of polyamines in higherplants by high performance liquid chromatography. PlantPhysiol 69: 701-706
19. Flores R, Rodriguez JL (1981) Altered pattern of root formationon cuttings of Gynura aurantiaca infected by citrus exocortisviroid. Phytopathology 71: 964-966
20. Foster SA, Walters DR (1990) The effects of polyamine biosyn-
BELLES ET AL.1 058
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

POLYAMINES IN HOST-VIROID INTERACTIONS
thesis inhibitors on mycelial growth, enzyme activities andpolyamine levels in the oat-infecting fungus, Pyrenophora av-enae. J Gen Microbiol 136: 233-239
21. Galston AW (1983) Polyamines as modulators of plant devel-opment. Bioscience 33: 382-388
22. Galston AW (1989) Polyamines and plant response to stress. InU Bachrach, YM Heimer, eds, The Physiology of Polyamines,Vol II, Polyamines in Microbes. Polyamines and plants. Poly-amines and Diseases. CRC Press, Boca Raton, FL, pp 99-106
23. Granell A, Belles JM, Conejero V (1987) Induction ofpathogen-esis-related proteins in tomato by citrus exocortis viroid, silverion and ethephon. Physiol Mol Plant Pathol 31: 83-89
24. Greenland AJ, Lewis DH (1984) Amines in barley leaves infectedby brown rust and their possible relevance to formation of"green islands". New Phytol 96: 283-291
25. Khan AJ, Minocha SC (1989) Polyamine biosynthetic enzymesand the effect of their inhibition on the growth of somephytopathogenic fungi. Plant Cell Physiol 30: 655-663
26. Rajam MV, Weinstein LH, Galston AW (1985) Prevention of aplant disease by specific inhibition of fungal polyamine biosyn-thesis. Proc Natl Acad Sci USA 82: 6874-6878
27. Semancik JS (1979) Small pathogenic RNA in plants. Theviroids. Annu Rev Phytopathol 17: 461-484
28. Semancik JS, Weathers LG (1972) Exocortis disease: evidencefor a new species of "infectious" low molecular weight RNAin plants. Nature New Biol 237: 242-244
29. Semancik JS, Morris TJ, Weathers LG, Roldorf GF, KearnsDD (1975) Physical properties of a minimal infectious RNA(viroid) associated with the exocortis disease. Virology 63: 160-167
30. Sisler EC, Yang SF (1984) Anti-ethylene effect of cis-2-buteneand cyclic olefins. Phytochemistry 23: 2765-2768
31. Slocum RD, Kaur-Sawhney R, Galston AW (1984) The physiol-ogy and biochemistry of polyamines in plants. Arch BiochemBiophys 235: 283-303
32. Smith TA (1985) Polyamines. Annu Rev Plant Physiol 36: 117-143
33. Smith MA, Davies PJ (1985) Separation and quantitation ofpolyamines in plant tissue by high performance liquid chro-matography of their dansyl derivatives. Plant Physiol 78: 89-91
34. Vera P, Conejero V (1988) Pathogenesis-related proteins of to-mato. P-69 as an alkaline endoproteinase. Plant Physiol 87:58-63
35. Walters DR, Wylie MA (1986) Polyamines in discrete regions ofbarley leaves infected with the powdery mildew fungus, Ery-siphe graminis. Physiol Plant 67: 630-633
36. Walters DR, Wilson PWF, Shuttleton MA (1985) Relativechanges in levels of polyamines and activities of their biosyn-thetic enzymes in barley infected with the powdery mildewfungus, Erysiphegraminis D.C. ex. Merat f.sp. hordei Marchal.New Phytol 101: 695-705
37. Weinstein LH, Osmeloski JF, Wettlaufer SH, Galston AW(1987) Protection of wheat against leaf and stem rust andpowdery mildew diseases by inhibition of polyamine metabo-lism. Plant Sci 51: 311-316
38. Winer L, Apelbaum A (1986) Involvement of polyamines in thedevelopment and ripening of avocado fruits. J Plant Physiol126: 223-233
39. Yu YB, Yang SF (1979) Auxin-induced ethylene production andits inhibition by aminoethoxyvinylglycine and cobalt ion. PlantPhysiol 64: 1074-1077
40. Yu YB, Adams DO, Yang SF (1979) I-Aminocyclopropanecar-boxylate Synthase, a key enzyme in ethylene biosynthesis. ArchBiochem Biophys 198: 280-286
1 059
www.plantphysiol.orgon August 24, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.





























