Embed Size (px)
Citation preview

Journal of Comparative and, Physiological Psychology1976, Vol. 90, No. 0, 660-571
Polarized Light Discrimination by Pigeonsand an Electroretinographic Correlate
Juan D. Delius, Robert J. Perchard, and Jacky EmmertonUniversity of Durham, Durham, England
Pigeons placed in a multiple-key Skinner-box could be trained to choose re-liably keys that were aligned in a specific way with the polarization axisof an overhead, randomly rotating light source. On the basis of these re-sults and those of additional control experiments, it is concluded thatpigeons can discriminate the axis orientation of linearly polarized lightand, furthermore, that they can orient themselves spatially by this cue.Electrophysiological recording experiments showed that the shape of theb-wave of the pigeons' electroretinogram is affected by the axis orientationof linearly polarized flash stimuli. This phenomenon seems to be due to thepresence of retinal polarization analyzers that may be tied to color visionmechanisms.
It is now a well-established fact that manyinvertebrates, especially arthropods, arecapable of perceiving the linear polarizationof a light source (von Frisch, 1967). Indeed,many orient themselves spatially by this cuein normal life, as the light from the skydisplays a polarization pattern that is linkedwith the position of the sun (Sekera, 1956).Humans are unware of this pattern sincethey are insensitive to light polarizationexcept under rather special circumstanceswhen they can perceive a faint, foveallycentered colored pattern (Haidinger'sbrushes), the orientation of which dependson the polarization axis (Lerner, 1970).
Until recently it was thought that allvertebrates shared this inability with us, butit has now been shown that at least certainfish (Forward & Waterman, 1973) andsalamanders (Taylor & Adler, 1973) are
This paper is dedicated with gratitude to NikoTinbergen on the occasion of his retirement asProfessor of Animal Behaviour at Oxford Uni-versity.
The work was supported by a Science ResearchCouncil (London) grant to J. D. Delius. The as-sistance of A. Perry, D. Barton, D. Harper, andMarie Cawton is gratefully acknowledged. Weare also indebted to K. Adler, W. T. Keeton, K.Schmidt-Koenig, and H. G. Wallraff for criticismof an earlier draft of the manuscript.
J. Emmerton is now at the Department ofPsychology, University of Liverpool.
Requests for reprints should be sent to JuanD. Delius, Department of Psychology, Ruhr-Universitat, Bochum, West Germany.
capable of orienting themselves guided bylinearly polarized light. Contrary to what anearlier study (Montgomery & Heinemann,1952) using a standard operaut discrimina-tion procedure suggested, pigeons are alsoable to detect polarization-axis changes oflight sources. Kreithen and Keeton (1973),using a classical heart rate conditioning pro-cedure, demonstrated that pigeons could betrained to distinguish between a light with astationary polarization plane and a lightwith rotating polarization plane.
This article reports a behavioral experi-ment that was begun while we were unawareof Kreithen and Keeton's work but that ex-tends their findings by showing that pigeonscan behave polarotactically, i.e., orient them-selves with respect to the polarization axisof an overhead surface light source. We alsopresent physiological evidence that certaincharacteristics of the pigeon's ocular bioelec-tric response to a light stimulus depend onthe polarization-axis orientation of thatlight.
EXPERIMENT 1: BEHAVIOR
MethodThe method was adapted from that commonly
used in operant discrimination learning. Therationale was that pigeons placed in a multiple-key Skinner box would be reinforced for peckingkeys that were lined up at some specific angle tothe polarization axis of an overhead source, whilenot being reinforced when they pecked keys linedup at 90° to that angle. From trial to trial, the
560

POLARIZED LIGHT DISCRIMINATION BY PIGEONS 561
^^
FIGURE 1. Diagrammatic top and obliqueviews of the apparatus used for the behavior ex-periment.
overhead polarization source would turn to lineup randomly with a new set of keys.
Subjects. Eight pigeons (Columba livia) oflocal homing stock and unknown sex were used.These animals had at least 1 yr of free-ranging ex-perience but were otherwise of varied age. Theywere kept in an artificially lit room and at 80%of their normal weight throughout the experiment.Before the experiment proper they were shaped topeck a key for grain reward in a conventionalSkinner box. They were then pretrained for sev-eral sessions in the chamber described below,which was not yet equipped with the polarizationfilter and was programmed so that a peck to anyof the four keys would deliver a grain reward. Atthis stage one pigeon responded unreliably andwas excluded from the experiment.
Apparatus. The apparatus consisted of a tall(50 cm) Skinner box (Figure 1) with an octagonalperimeter (8 X 20 cm). Four translucent keys(2.5 cm diameter) were placed recessed 1 cm intoeach alternate wall 15 cm from the floor. Eachkey was illuminated from behind with a miniaturebulb. A solenoid-operated grain hopper had itsopening (3 cm diameter) in the center of the boxfloor. The lid of the box had a circular cut-out(28 cm diameter). In some experiments (see below)this opening was covered with a diffusing sheet of3-mm-thick glass, frosted on one side. Centeredover this opening was a lamp house of inverted,truncated conical shape (30 cm high, 30 cm maxi-mum diameter, and 20 cm minimum diameter).Centered inside at the top of the lamp house wasa 150-W tungsten-iodine bulb with a semispherical,frosted glass diffuser. The inner walls of the upperhalf of the lamp house were lined with crumpledaluminum foil serving as a diffusing reflector.Two glass diffusing plates, frosted on both sidesand of 4-rnm thickness, were located 3 and 7 cmfrom the bottom. Finally, below these and 1.5cm above the Skinner box lid, a polarization filter(Polaroid HN 38) was mounted in a detachable
circular ring. Measured with a SEI photometerthe polarization source had a luminance of 2.7 logftL. («1,700 cd/m2) at the center fading to 2.5log ftL. («1,100 cd/m2) at the edges. The in-tensity could be reduced by lowering the supplyvoltage to the tungsten-iodine bulb, with a conse-quent change in color temperature.
The top of the lamp house was attached to ahollow axle conducting the leads for the bulb.This axle in turn was held by a ballrace mountedon a support stand. A small, silent electric motorattached to the stand rotated the axle through awormgear. The lamp house with the polarizationfilter could thus be turned to any cardinal direc-tion. A disk with stops, mounted on the axle,activated microswitches fixed to the stand suchthat lamp house positions at 90° intervals couldbe sensed by the programming equipment. Theapparatus was controlled automatically by con-ventional relay equipment. The animal's responseswere recorded on counters. In all critical experi-ments the programming gear was in a room ad-jacent to the one containing the experimental box.This latter room was kept totally dark. A loud-speaker producing white noise was situated im-mediately over the box and gave a sound level of75 dB (re 20 juN/m2) within the box. It masked anyunintentional auditory cues. To minimize reflec-tions the interior of the chamber was carefullycoated with a rough-surfaced, matt-black paint.
During its construction the design of the ap-paratus was empirically adjusted towards re-moving unwanted cues arising from the insidious,axis-dependent, differential reflection of polarizedlight. Details that contributed markedly to thiswere the bright transillumination of the keys thatmasked all reflections on them, the large-surfacesource of polarized light that gave a diffuse illum-ination, and the grained, matt finish of the cham-ber interior that enhanced that diffusion.
Procedure. A trial began with a time-out periodof 20 sec duration when all lights were switchedoff. To avoid the development of key preferencesa correction procedure was used. Thus, if the pre-vious trial had ended with an unrewarded re-sponse, the next trial took place with the stimuluscondition unaltered (correction trial). If, how-ever, the preceding trial ended with a rewardedresponse, the stimulus condition was controlledas follows: Three seconds after the beginning ofthe time-out period, the overhead polarized lightsource turned by 90° (p = .5), by 0° (p = .25), orby 180° (p = .25) according to Gellermann semi-random sequences. In the last two cases, of course,the direction of the polarization axis remainedidentical to that of the previous trial. The clock-wise (p = .5) or anticlockwise (p = .5) directionof rotation was also determined by a semirandomGellermann sequence. In every case the turningmaneuver was terminated before the end of thetime-out period was over and before the polarizedlight source and the key lights came on. When abird pecked one of the pairs of keys lined up withthe polarization plane in the manner specified as

562 J. DELIUS, R. PERCHARD, AND J. EMMERTON
i25
sess/onsFIGURE 2. Polarized light orientation discrimination; learning curves of three pigeons. (The inserts
illustrate the discrimination task of the particular subject. The double arrow symbolizes the polari-zation axis. Sessions in which the diffusing plate was used are marked with asterisks.)
"correct" for the particular subject, the key lightswent off, and the hopper came up for 7 sec offeringgrain as reward. Immediately the reinforcementperiod was over, the next trial began with thetime-out period. When a bird pecked one of thepair of keys lined up in the "incorrect" manner,all lights went off, beginning an additional 10-sec "punishment" time-out before the time-outperiod proper preceding the next trial started.Pecks at the darkened keys during the time-outperiods led to a reinitiation of the time-out.
As is adequate for the type of correction proce-dure used, only those responses that were emittedduring noncorrection trials, i.e., in those trials inwhich the subject could not predict the position ofthe polarization plane by previous events, wererecorded. Each subject was run on one dailysession at about the same time each day. A sessionconsisted of 30 (for the first three subjects) or 40noncorrection trials.
Results
Two subjects were trained with the twopositive keys aligned in parallel with the
polarization axis, three subjects were trainedwith the correct keys lined up 45° anticlock-wise from the polarization plane, and twoof these latter three subjects were later re-trained with the positive keys at 90° fromthe polarization axis. The session-by-sessionlearning curves of three subjects are shownin Figure 2. The course of the four remaininglearning curves fell within the range of theones shown. All of them showed clearly thatthe animals could perform consistentlyabove 50 % trials correct after an acquisitionperiod varying between one and nine ses-sions. The proportion of correct versus in-correct responses over the last four sessionsfor each bird was tested by chi-square testsagainst the 50 % null hypothesis and yieldedin each case a p = .01 or better, suggestingthat the pigeons were capable of orientingthemselves by the plane of polarization of
POLARIZED LIGHT DISCRIMINATION BY PIGEONS
C
563
70 iV.
co0.
30 J
70 -i
oo
\/ \/-A8 15
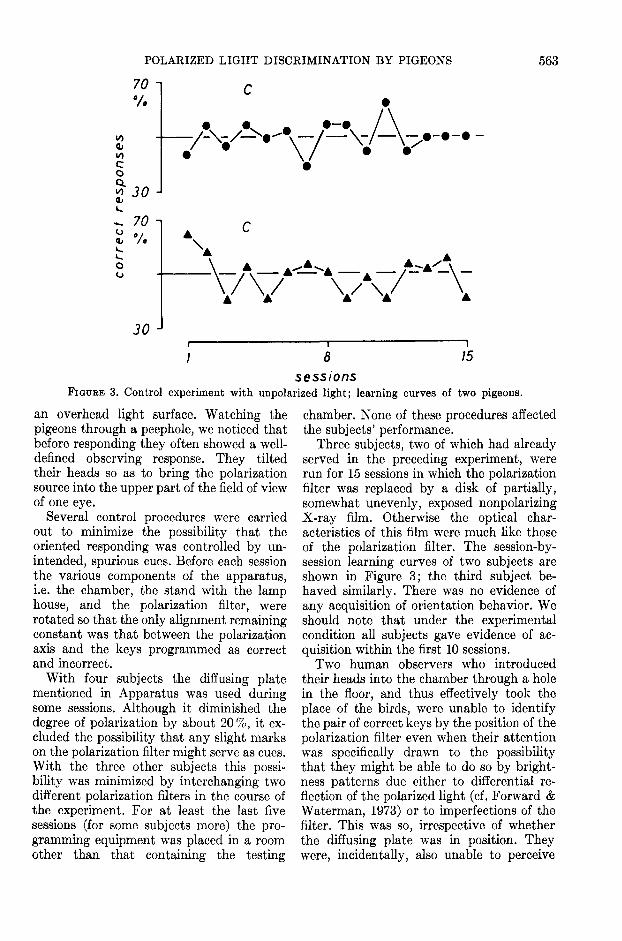
sessionsFIGURE 3. Control experiment with unpolarized light; learning curves of two pigeons.
an overhead light surface. Watching thepigeons through a peephole, we noticed thatbefore responding they often showed a well-defined observing response. They tiltedtheir heads so as to bring the polarizationsource into the upper part of the field of viewof one eye.
Several control procedures were carriedout to minimize the possibility that theoriented responding was controlled by un-intended, spurious cues. Before each sessionthe various components of the apparatus,i.e. the chamber, the stand with the lamphouse, and the polarization filter, wererotated so that the only alignment remainingconstant was that between the polarizationaxis and the keys programmed as correctand incorrect.
With four subjects the diffusing platementioned in Apparatus was used duringsome sessions. Although it diminished thedegree of polarization by about 20%, it ex-cluded the possibility that any slight markson the polarization filter might serve as cues.With the three other subjects this possi-bility was minimized by interchanging twodifferent polarization filters in the course ofthe experiment. For at least the last fivesessions (for some subjects more) the pro-gramming equipment was placed in a roomother than that containing the testing
chamber. None of these procedures affectedthe subjects' performance.
Three subjects, two of which had alreadyserved in the preceding experiment, wererun for 15 sessions in which the polarizationfilter was replaced by a disk of partially,somewhat unevenly, exposed nonpolarizingX-ray film. Otherwise the optical char-acteristics of this film were much like thoseof the polarization filter. The session-by-session learning curves of two subjects areshown in Figure 3; the third subject be-haved similarly. There was no evidence ofany acquisition of orientation behavior. Weshould note that under the experimentalcondition all subjects gave evidence of ac-quisition within the first 10 sessions.
Two human observers who introducedtheir heads into the chamber through a holein the floor, and thus effectively took theplace of the birds, were unable to identifythe pair of correct keys by the position of thepolarization filter even when their attentionwas specifically drawn to the possibilitythat they might be able to do so by bright-ness patterns due either to differential re-flection of the polarized light (cf. Forward &Waterman, 1973) or to imperfections of thefilter. This was so, irrespective of whetherthe diffusing plate was in position. Theywere, incidentally, also unable to perceive

564 J. DELITJS, R. PERCHARD, AND J. EMMERTON
Haidinger's brushes. They were, however,just able to choose the correct keys whenobserving the control X-ray film because ofa faint brightness unevenness. The failureof the pigeons to orient to this cue is prob-ably because of their relatively poor light-intensity discrimination capabilities (Ment-zer, 1966; but see Hodos & Bonbright, 1972).We believe, therefore, that the pigeons werecapable of perceiving the polarization-axisorientation of the overhead light surfacedirectly and were not responding to unin-tentional or secondary cues.
Two subjects were run in an additionalexperiment. After they had mastered thediscrimination, we reduced in four successivesessions the intensity of the overhead polar-ized light by .5 log units, starting from thenormal 2.7 log ftL. (^1,700 cd/m2) lumi-nance. The performance of both birds im-proved slightly with the two initial dimmingsteps but fell drastically from above 80 %correct to close to 50% correct from thethird to the fourth step as the intensity ofthe polarized light fell to .7 log ftL. (;^17cd/m2), i.e., to a scotopic level for humanobservers. Over the luminance range of 2.7to and including 1.2 log ftL. (?»54 cd/m2),however, pigeons appear to be able to main-tain their polarotactic behavior. They differin this respect from the fish that have beentested (Forward & Waterman, 1973), whosepolarotactic behavior is apparently de-pendent on high light intensities (however,see Waterman & Aoki, 1974).
In two interspersed sessions each we alsotested the same two subjects at a daytime6 hr earlier than the usual one. We expectedthat if their polarotactic behavior was timecompensated, as the pigeon's sun-compassorientation is (Schmidt-Koenig, 1958), theywould tend to choose the keys at 90° to thecorrect ones, i.e., the incorrect ones conform-ing with the approximately 90° azimuthrotation that the sky polarization patternundergoes in 6 hr. Their performance, how-ever, was unaffected, but this is not a proofagainst time compensation in conjunctionwith polarotactic behavior. Rather, it mayhave to do with the experimental design.Depending on the details of the experiments,pigeons do or do not time compensate their
orientation with respect to the sun (Schmidt-Koenig, 1958).
EXPERIMENT 2: ELECTROPHYSIOLOGYMethod
The method for this experiment was to recordthe pigeon's ocular bioelectric response to po-larized light flashes, while the orientation of thepolarization axis was varied. If the eye respondeddifferentially to different orientations of thepolarization axis, then it might be possible toobserve electroretinogram shape alterations.
Subjects. Seven pigeons of similar provenanceto those used in the previous experiment, of atleast 2 yr of age and of unknown sex, served assubjects. Before the experiment proper each birdwas anesthetized with Equi-Thesin (.25 ml/100g of body weight). While the head was held in aspecial holder (Karten & Hodos, 1967), the scalpwas incised and retracted, the skull was pittedwith a dental drill, and a small (.5 X .7 X .3 cm)block of brass with two tapped holes was cementedto the skull with dental acrylic. The pigeons werethen left to heal and recover for at least 5 days.
Apparatus. The apparatus consisted of a pro-jection system built on a standard optical bench(Figure 4). Collimated light from a 100-Wtungsten-iodine bulb was passed through an in-frared- and an ultraviolet-blocking filter, whichlimited the spectrum to between 380 and 750 nm.A second lens focused light onto a 2-mm aperture,next to which was a sector-disk shutter driven bya variable speed electric motor. A further colli-mating lens transmitted the light through aneutral-density optical wedge and an adjustablepolarization filter (Polaroid HN 38) to a focusinglens (in some experiments the order of these lasttwo elements was reversed), which yielded aconical beam subtending 14°. All lenses wereachromatic doublets. Calibrations were performedwith a Hilger-Watts FT 17 thermopile placed atthe focal point. The neutral-density filter was setso that the thermopile gave a reading of 20 juW.Small neutral-density wedge adjustments werenecessary to maintain the energy reading as thepolarizing filter was rotated to four standard
IR UV
t>FIGURE 4. Optical stimulator used for the elec-
troretinogram experiment. (Abbreviations: L,lamp; IR and UV, infrared and ultraviolet block-ing filters; S, sector disk shutter; ND, neutral-density wedge; CF, color filter; P, polarizer; E,eye. In some experiments the polarizer was placedbetween the focusing lens and the eye.)

POLARIZED LIGHT DISCRIMINATION BY PIGEONS 565
positions 45° apart. This was necessary becauseof a partial linear polarization of the light emittedby the tungsten-iodine bulb. When in the experi-ment the spectral band was narrowed further withcolor filters, the light intensity was adjusted togive approximately the same electroretinogramamplitude as white light at the baseline intensity.Compensatory intensity adjustments for the var-ious polarization filter positions were madethroughout.
The metal block on the subject's head couldbe screwed to an adjustable arm. An Ag-AgCl,circum-corneal ring electrode was placed againstthe subject's left eye with the help of another ad-justable arm. Electric potentials from the ringelectrode and another Ag-AgCl electrode placedunder the scalp were amplified differentially witha Grass P15 amplifier set with a bandwidth of .1-100 Hz. The bird was earthed by a subcutaneousneedle electrode located on the rump. The elec-troretinograms were displayed on a Tektronix 564storage scope and averaged on a Biomac 500transient averaging computer together with astimulus trace from a photocell. An adjustablephotoelectric gate device on the shutter providedthe necessary triggering pulses. When required,oscilloscope displays of the averages were re-corded photographically.
Procedure. Each subject was anesthetized withEqui-Thesin (im .20 ml/100 g of body weight).Supplementary doses (.07 ml/100 g) were given asnecessary until eye movements were absent. Theleft eye was treated topically with an atropine(2%), succinylcholine (5%), Xylocaine (2%), andbenzalkonium (.1%) solution until the eyelid,nictitating membrane, and pupil were totally re-laxed and the corneal reflex was absent.
The bird was then mounted in the apparatus.Mostly the light beam was focused on the centerof the pupil so that the eye was illuminated in anaxial, Maxwellian fashion, but other illuminationmodes were tried as well. The electrodes were setin place, the shutter was set in motion, and re-cording began.
We made no systematic effort to control theadaptation level of the pigeon's retina except thatabout every 10 min the room lights were switchedon for about 5 min (light level at the pigeon'seye 1.5 log ftL.; wllO cd/m2). This, together withthe pigeon's low rate of adaptation (Blough, 1956),presumably ensured a degree of light adaptation.The electroretinographic records suggested aphotopic state, i.e., one in which cones were ac-tive.
All electroretinogram records were based onaverages of 32 individual responses. Usually theprocedure was to record successively four aver-ages each taken with the polarization filter ro-tated by 45° and then to repeat the sequence withthe reverse order of filter positions, thus checkingthe replicability of the responses. In one of thesefilter positions the polarization axis was approxi-mately parallel with the auditory meatus-palatine
line that is used as a stereotactic baseline (Karten& Hodos, 1967). The electroretinogram amplitudesrecorded at the baseline stimulus intensity werebetween 50 and 70 /xV in amplitude.
Varying flash and interflash durations weretried. Those mostly used ranged between 100 and200 msec for the flashes and 500 and 1,000 msec forthe intervals. In some experiments we limited thespectral bandwidth of the stimulus light with ayellow filter (Kodak 12) that cut off the short-wave light below 450 nm (the wavelengths wherethe polarization filter was relatively inefficient)or with broadband blue, green, or red filters(Kodak 50, 58, and 29).
Results
We first noticed a difference in the shapeof averaged electroretinograms taken withdifferent polarization orientations from twosubjects when using 6-Hz flicker stimuli, butthe effects were rather small and somewhatunstable. Flashes of 100 msec or more dura-tion gave less variable, more obvious dif-ferences. Recordings showing these are pre-sented in Figure 5. As can be seen, the maindifference relates to the shape of the b-wave
50 JJV
/00msFIGURE 5. Eleetroretinograms obtained with
various polarization-axis orientations (Subject D,first session). (The lower set of recordings wasobtained with the standard stimulus intensity;the upper set was obtained 55 min later with abrighter [+.5 log units] stimulus intensity.)

566 J. DELIUS, R. PERCHARD, AND J. EMMERTON
100 ms
FIGURE 6. Electroretinograms obtained during an attempt to replicate the polarization orientationeffect (left) with stimulus intensity changes (right; Subject F). (l.u. = log units.)
of the electroretinogram, i.e., the second,positive component of the response to theonset of the light stimulus. When the polari-zation axis is 45° clockwise with respect tothe meatus-palatine line (horizontal arrowson the figures), the summit of that wave issharp and singular. When, however, thepolarization axis is at 45° anticlockwise ofthe meatus-palatine line (vertical arrows onthe figures), the peak is flat or even doubled.At the two other polarization-axis orienta-tions the shape of the b-wave is intermedi-ate. Since the computer we used had nofacilities for a statistical analysis of thewaveforms, the reliability of the effect hadto be assessed by its replicability. Four sub-jects, one of them in two separate sessions,showed these effects unmistakably in 45 outof 47 sets of recordings when the standardstimulus conditions were used. The reader isreferred to four sets of recordings shown inFigures 5, 6, and 8 which illustrate thebasic effect as obtained on different occa-sions and from different pigeons. In anotherpigeon that had an unusual electroretino-gram (sharply peaked b-wave; in otherelectroretinographic investigations we havealso occasionally noticed aberrant individ-uals), the effect was present but very small.It would have certainly passed unnoticednormally.
This polarization-orientation effect wasnot altered if the polarization filter was
placed behind the focusing lens rather thanbefore it. That is to say, it was not due todifferent brightness patterns that mighthave arisen within the optical system be-cause of the axis-dependent reflection/re-fraction properties of linearly polarized light.We were actually unable to detect such apattern in spite of careful inspection.
Note that the overall amplitudes of theelectroretinograms at the various polariza-tion orientations are not appreciably differ-ent. This suggests that the electroretinogramdifferences are not due to secondary, overallintensity differences of the light reachingthe retina, arising perhaps because of aniso-tropic behavior of the ocular media. How-ever, to exclude this possibility further, weattempted to replicate the electroretinogramdifferences by varying the intensity of thestimulus flashes over the range of +.3 to-.3 log units of the baseline intensitywhile keeping the polarization axis constant.Figure 6 shows the results of one such at-tempt. As one might expect, the electro-retinogram amplitude increases with theintensity, but the shape of the b-wave re-mains very nearly constant; at least it doesnot replicate the summit pattern differencesthat are dependent upon the polarizationplane. Apart from this, the polarizationeffect persisted both at higher (up to +1log units) and lower (down to — 1 log units)stimulus intensities than the standard in-

POLARIZED LIGHT DISCRIMINATION BY PIGEONS 567
gretn
red
flash
100ms
FIGURE 7. Electroretinograms obtained withpolarized light flashes of different orientation andcolor (Subject G).
tensity. But the effect was then not so ob-vious because the lower signal-to-noise ratioat low intensities and because the crowdingof the b-wave components at high intensitiesobscured the rather subtle shape differ-ences.
Exclusion of the relatively unpolarizedblue light of the spectrum by a yellow filterseemed to enhance the polarization-orienta-tion effect slightly. Broadband blue light,i.e., light that was only slightly polarized(the polarization coefficient of the polarizingfilter used is 70 % between 700 and 450 nmbut falls off to 20% at 400 nm), did notyield the effect. Both broadband green andred light produced an effect but apparentlymodified in a complex, color-specific wayillustrated in Figure 7.
Since under normal conditions pigeons arelikely to view the polarized light of the skymainly with the upper sections of the visualfield, we ran some experiments in which thehead of the subject was tilted so that thestimulus light fell at 40° off the eye axison the lower half of the retina. This en-hanced the polarization effect (Figure 8).A similar enhancement seemed to occurwhen the light source was focused at theedge of the pupil while the Maxwellianbeam was still parallel to the axis of theeye, i.e., in a situation in which the single
visual cells are struck by the light at anangle off their anatomical axis.
The results relating to the three laststimulus conditions, i.e., colored light,peripheral field, and off-axis illumination,must, however, be considered as prelimi-nary. They are presently being investigatedin more detail. Finally, we noticed, but didnot investigate further, a correspondingpolarization effect in the "off" wave of theelectroretinogram. The type of stimulationwe used yielded only a small response ofthis type; dark "flashes" may reveal the"off" effect better.
DISCUSSIONWe conclude from the preceding experi-
ments that contrary to the findings ofMontgomery and Heinemann (1952) but inaccordance with those of Kreithen andKeeton (1973), pigeons are capable of de-tecting the orientation of linearly polarizedlight. Furthermore, we believe that we haveshown that they can orient themselvesspatially by that cue. Since our behavioralexperiment imitated to some extent theconditions prevailing under natural condi-tions, it seems reasonable to assume thatpigeons can use the sky polarization patternsto supplement their sun-compass orienta-tion when they perform their remarkablehoming feats.
However, polarized light is a notablydifficult stimulus to control. The possibilitythat the discriminative behavior shown bypigeons in our and Kreithen and Keeton'sexperiments might have been due to thepigeons' being capable of perceiving unin-tended secondary cues that were not ap-parent to the human eye can, of course,not be excluded. The fact that the positiveexperiments involved very different kindsof stimulus presentation renders this al-ternative unlikely in our view. Experimentsincorporating further control procedures willbe necessary before there can be completecertainty on this issue.
We suggest that Montgomery and Heine-mann's (1952) inability to show discrimi-nation was because of the nonbiologicalconditioning situation that they used. Innormal life it is unlikely that pigeons couldever derive selective advantages from dis-

568 J. DELIUS, R. PERCHARD, AND J. EMMERTON
100 ms
FIGUBE 8. Electroretinograms obtained with polarized light flashes striking the eye axially (left)and from above (right; Subject D, second session).
languishing the polarization axis of smalllight sources situated in the lower half oftheir visual field because of the absence ofsuch sources in their usual environment.Thus, they may well not have evolved thenecessary sensory and/or neural apparatusfor such a discrimination. Skinner box keysare viewed by pigeons with the upper pos-terior quadrant of the retina, the so-calledred field known to be morphologically andfunctionally different from the remainder ofthe retina (Galifret, 1968; Nye, 1973;Pedler & Boyle, 1969; Yazulla, 1974). Theresponses of this area were not studied in ourelectroretinographic experiment, but thearea was certainly not employed by the sub-jects to examine the overhead polarizationsource in our behavior experiment.
Although the polarotactic behavior ofsalamanders seems to depend on cranialextraocular receptors (Adler & Taylor,1973), it is unlikely that this is so in pigeons.Even though some extraretinal light sensi-tivity, possibly associated with the pineal,has been demonstrated in birds (e.g., Under-wood & Menaker, 1970), it seems improb-able that it could be involved in controlling
the polarotactic behavior of pigeons. Thepineal organ is located too deeply in thecranium under obscuring and diffusing lay-ers of feathers, skin, and bone to be a likelypolarization-axis detector. In any caseMorita (1966) was unable to record anynonocular bioelectric light responses fromthe avian pineal, whereas such neuroelectriclight responses occur in the pineal organ ofamphibians (Dodt & Morita, 1967). Thefact that the pigeon's eye seems electro-physiologically sensitive to the polarizationaxis of light, that the pigeons made clear-cutocular observing responses in our behavioralexperiment, and that in Kreithen and Kee-ton's experiments the stimulus was arrangedso that it is unlikely that the pigeons"viewed" it with the pineal also militatesagainst that possibility. Indeed, in fishWaterman and collaborators (Waterman &Aoki, 1974; Waterman & Hashimoto, 1974)described polarization-axis-sensitive neu-ronal units in the optic tectum that re-sponded to stimulation of the eye.
For two reasons it seems unlikely to usthat the cornea, lens, or vitreous is actingas an analyzer. First, if this were so, one

POLARIZED LIGHT DISCRIMINATION BY PIGEONS 569
would have expected that the electroreti-nographic polarization effect would havebeen reproduced by suitable flash intensityvariations; but this was not so. Second,examination of two freshly excised pigeoneyes with transilluminating polarized lightof various axis orientations did not revealsuch a property.
The observing responses made by thepigeons in our conditioning chamber werevirtually never of sufficient amplitude toallow them a foveal view of the overheadpolarized light source. In addition, periph-eral illumination that did not fall on thefovea in the electroretinogram experimentstill yielded polarization orientation effects,so it is improbable that the polarization de-tection mechanism of pigeons is based onone analogous to that responsible for theHaidinger's brushes in man.
To account for the polarization sensitivityof fish, which is dependent on very brightlight, Snyder (1973) proposed that at highlight intensities, small but sufficient amountsof light would be reflected and refractedfrom neighboring receptor cells onto thesensitive rod outer segments, which whenilluminated at right angles, are known tobe dichroic. Since in both our behavioraland electrophysiological experiments pigeonscontinued to be sensitive to the orientationof the polarization axis at quite low levelsof illumination and since additionally inthe electrophysiological experiment con-tinued to be responsive when illuminatedwith red light (to which rods are largely in-sensitive), we think that Snyder's hypothesisdoes not apply to the pigeon.
Birds possess a peculiar dark, comblikestructure, the pecten, that protrudes fromthe fundus into the ventral half of thevitreous. While it certainly has trophicfunctions (Welsch, 1972), it is widely sus-pected to have optical functions as well(e.g., Barlow & Osterwald, 1972). Attractiveas the idea is, we cannot think of any physi-cal characteristic that could make it a polari-zation analyzer or of any experimental datathat would support that role for it except-ing perhaps its location, which involves itwith the upper parts of the visual fieldwhich might be particularly polarizationsensitive according to the present evidence.
Rather, we would like to suggest thatthe polarization-axis detection in pigeonsis connected with specialized retinal struc-tures, the double cones, that birds sharewith all other nonplacental vertebrates.One of the elements of the double cones,the accessory cone, appears to have, bythe nature of its shape in the light-adaptedstate, a greater acceptance angle for lightthan do other retinal receptors. It alsopossesses a special, electronmicroscopicallystructured, highly refractive glycogen in-clusion, the paraboloid. The other elementof the pair, the chief cone, is intimatelyapposed to only one side of the accessorycone, and it seems likely that this junctionis relatively transparent (Cohen, 1963).It is conceivable that this system mightfunction as a polarization analyzer on thebasis of the differential reflection/refrac-tion properties of polarized light (seeForges, 1974). Furthermore, the lineup ofthe accessory/chief cone pairs, togetherwith their relative positions with respectto the other neighboring visual cells, formsa regular mosaic pattern over the retina(chicken: Morris, 1970) that would enablethe recognition of surface polarization pat-terns such as that of the sky. In pigeonsthese double cones seem to be rare in thered field but common in the remainder ofthe retina. This would correspond with theapparent lack of polarization sensitivity ofthe red field of the retina. The chief conescarry a yellow-colored oil droplet, whichundoubtedly makes them differentially colorsensitive from the accessory cones that havenone and from the single cones that carrya variety of differently colored oil droplets(King-Smith, 1969; see also Richter &Simon, 1974). This would relate to theapparent color dependence of the electro-retinographic polarization effect that webelieve to have found.
Considerations of whether this conceptcould account for the properties of thepolarization-sensitive units in the optictectum of the goldfish studied by Water-man and Aoki (1974) are obstructed bythe fact that the twin cones of fish aremorphologically different from the doublecones of birds and that the tectal responses

570 J. DELIUS, R. PERCHARD, AND J. EMMERTON
are likely to reflect receptor processes al-ready complicated by neural processing.
Relating to the possible interaction be-tween the spectral composition and theorientation of the polarized light, we wouldlike to draw attention to the possibilitythat the b-wave "wavelets," which seem tobring about the basic polarization orienta-tion effect by their varying relative ampli-tudes, may be the same as those describedby Nye (1968). His failure to find a con-sistent correlation between the stimuluswavelength and the relative amplitudes ofthe wavelets may have been due to his nothaving taken into account the uninten-tional linear polarization of his stimuli(their generation involved reflection offmirrors!) and the precise orientation of thesubject's eyes with respect to them.
REFERENCES
Adler, K., & Taylor, D. H. Extraooular perceptionof polarized light by orienting salamanders.Journal of Comparative Physiology, 1973, 87,203-212.
Barlow, H. B., & Osterwald, T. J. Pecten of thepigeon's eye as an interocular eye shade. Na-ture, 1972, 236, 88-90.
Blough, D. S. Dark adaption in the pigeon. Jour-of Comparative and Physiological Psychology,1956, 49, 425-430.
Cohen, A. I. The fine structure of visual receptorsof the pigeon. Experimental Eye Research, 1963,0, 88-97.
Dodt, E., & Morita, Y. Conduction of nerve im-pulses within the pineal system of frog. PflugersArchiv fur die gesamte Physiologie des Menschenund der Tiere, 1967, 298, 184-192.
Forward, B., & Waterman, T. H. Evidence fore-vector and light intensity pattern discrimi-nation by the teleost Dermogenys. Journal ofComparative Physiology, 1973, 87, 189-202.
Frisch, K. von. The dance language and orientationof bees. Cambridge, Mass.: Belknap Press, 1967.
Galifret, Y. Les diverses aires fonctionelles de lare'tine du pigeon. Zeitschrift fur Zellforschungund Mikroskopische Anatomie, 1968, 86, 535-545.
Hodos, W., & Bonbright, J. C. The detection ofvisual intensity differences by pigeons. Journalof the Experimental Analysis of Behavior, 1972,18, 471-479.
Karten, H. J., & Hodos, W. A stereotaxic atlas ofthe brain of the pigeon (Columba livia). Balti-more: Johns Hopkins University Press, 1967.
King-Smith, P. E. Absorption spectra and func-tion of the coloured oil drops in pigeon retina.Vision Research, 1969, 9, 1391-1399.
Kreithen, M. L., & Keeton, W. T. Detection ofpolarized light by the homing pigeon, Columbalivia. Journal of Comparative Physiology, 1973,89, 83-92.
Lerner, G. Haidinger's brushes and the perceptionof polarization. Acta Psychologica, 1970, 34,106-114.
Mentzer, T. L. Comparison of three methods forobtaining psychophysical thresholds from thepigeon. Journal of Comparative and PhysiologicalPsychology, 1966, 61, 96-101.
Montgomery, K. C., & Heinemann, E. G. Con-cerning the ability of homing pigeons to dis-criminate patterns of polarized light. Science,1952, 116, 454-456.
Morita, Y. Absence of electrical activity of thepigeon pineal organ in response to light. Ex-perientia, 1966, 02, 402.
Morris, V. B. Symmetry in a receptor mosaicdemonstrated in the chick from frequencies,spacing and arrangement of the types of retinalreceptor. Journal of Comparative Neurology,1970, HO, 359-398.
Nye, P. W. An examination of the electroretino-gram of the pigeon in response to stimuli ofdifferent intensity and wavelength and follow-ing intense chromatic adaption. Vision Research,1968, 8, 679-696.
Nye, P. W. On the functional differences betweenfrontal and lateral visual fields of the pigeon.Vision Research, 1973, IS, 559-574.
Pedler, C., & Boyle, M. Multiple oil droplets inthe photoreceptors of the pigeon. Vision Re-search, 1969, 9, 525-528.
Forges, A. Telling time. Science, 1974, 184, 1133.Richter, A., & Simon, E. J. Electrical responses of
double cones in the turtle retina. Journal ofPhysiology, 1974, 040, 673-683.
Sehmidt-Koenig, K. Experimented Einnuss-nahme auf die 24 Stunden Periodik bei Brief-tauben und deren Auswirkungen unterbesonderer Beriicksichtigung des Heimfinde-vermogens. Zeilschrift fur Tierpsychologie, 1958,IB, 301-331.
Sekera, Z. Polarization of skylight. In S. Fliigge(Ed.), Handbuch der Physik. Berlin: Springer-Verlag, 1956.
Snyder, A. W. How fish detect polarized light. In-vestigative Ophthalmology, 1973,10, 78-79.
Taylor, D. H., & Adler, K. Spatial orientation bysalamanders using plane polarized light. Science,1973, 181, 285-287.
Underwood, H., & Menaker, M. Photoperiodicallysignificant photoreception in sparrows: Is theretina involved? Science, 1970, 167, 298-301.
Waterman, T. H., & Aoki, K. E-vector sensitivitypatterns in the goldfish optic tectum. Journal ofComparative Physiology, 1974, 95, 13-27.
Waterman, T. H., & Hashimoto, H. E-vectordiscrimination by the goldfish optic tectum.Journal of Comparative Physiology, 1974, 95,1-12.

POLARIZED LIGHT DISCRIMINATION BY PIGEONS 571
Welsch, TJ. Enzymhistochemische und feinstruk- Yazulla, S. Intraretinal differentiation in theturelle Beobachtungen am Pecten oculi von synaptic organization of the inner plexiformTaube und Lachmowe. Zeitschrift fur Zell- layer of the pigeon retina. Journal of Compara-forschung und mikroskopische Anatomie, 1972, five Neurology, 1974, IBS, 309-324.1S8, 231-244. (Received June 13, 1975)





























