Embed Size (px)
Citation preview

Experimental Parasitology 128 (2011) 127–132
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
Plasmodium vivax: Collaborative roles for plasmepsin 4 and vivapainsin hemoglobin hydrolysis
Sung-Ung Moon a,1, Jung-Mi Kang a, Tong-Soo Kim b, Yoon Kong c, Woon-Mok Sohn a, Byoung-Kuk Na a,⇑a Department of Parasitology, Brain Korea 21 Biomedical Center, and Institute of Health Sciences, Gyeongsang National University School of Medicine, Jinju 660-751, Republic of Koreab Department of Parasitology and Inha Research Institute for Medical Sciences, Inha University College of Medicine, Incheon 400-751, Republic of Koreac Department of Molecular Parasitology, Center for Molecular Medicine, Samsung Biomedical Research Institute, Sungkyunkwan University School of Medicine,Suwon 440-746, Republic of Korea
a r t i c l e i n f o a b s t r a c t
Article history:Received 25 August 2010Received in revised form 8 December 2010Accepted 14 February 2011Available online 18 February 2011
Keywords:Plasmodium vivaxAspartic proteasePlasmepsin 4VivapainHemoglobin hydrolysis
0014-4894/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.exppara.2011.02.015
⇑ Corresponding author. Fax: +82 55 759 4022.E-mail address: [email protected] (B.-K. Na).
1 Department of Anatomy, Yonsei University CollegeRepublic of Korea.
Plasmepsins, a family of aspartic proteases of Plasmodium species, are known to participate in a wide vari-ety of cellular processes essential for parasite survival. Therefore, the plasmepsins of malaria parasiteshave been recognized as attractive antimalarial drug targets. Although the plasmepsins of P. falciparumhave been extensively characterized, the plasmepsins of P. vivax are currently not well known. To expandour understanding of the plasmepsins of P. vivax, we characterized plasmepsin 4 of P. vivax (PvPM4). Thebacterially expressed recombinant PvPM4 was insoluble, but it was easily refolded into a soluble protein.The processing of PvPM4 into a mature enzyme occurred through autocatalytic activity under acidic con-ditions in a pepstatin A-sensitive manner, in which process a portion of prodomain was essential for cor-rect folding. PvPM4 could hydrolyze native human hemoglobin at acidic pHs, but preferred denaturedhemoglobin as a substrate. PvPM4 acted synergistically with vivapain-2 and vivapain-3, cysteine prote-ases of P. vivax, in the hydrolysis of hemoglobin. The vivapains also mediated processing of PvPM4 into amature enzyme. These results collectively suggest that PvPM4 is an active hemoglobinase of P. vivax thatworks collaboratively with vivapains to enhance the parasite’s ability to hydrolyze hemoglobin.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
Malaria, which is caused by the apicomplexa parasitePlasmodium spp., is one of the most important parasitic diseasesworldwide, with approximately 200–300 million clinical casesand 1–3 million deaths each year worldwide. Hemoglobin hydroly-sis by malaria parasites is an essential biological process that notonly for providing amino acids for parasite growth but also makingspace for parasites in the erythrocyte or maintaining osmotic bal-ance (Krugliak et al., 2002; Lew et al., 2003,2004). A series of differ-ent classes of proteases, cysteine, aspartic, and metalloproteases,play critical roles in hemoglobin hydrolysis in the food vacuole ofPlasmodium falciparum (Francis et al., 1994; Eggleson et al., 1999;Shenai et al., 2000; Sijwali et al., 2001; Banerjee et al., 2002;Rosenthal, 2002; Klemba et al., 2004a; Singh et al., 2006). Proteaseinhibitors inhibit hemoglobin hydrolysis in the parasite and blockthe parasite development (Rosenthal et al., 1991; Francis et al.,1994; Olson et al., 1999; Jiang et al., 2001; Moura et al., 2009).Recent gene knockout studies also have demonstrated pivotal roles
ll rights reserved.
of Medicine, Seoul 120-752,
for these enzymes in hemoglobin hydrolysis, parasite developmentand survival (Omara-Opyene et al., 2004; Sijwali and Rosenthal,2004; Sijwali et al., 2006; Bonilla et al., 2007a,b). These studies col-lectively suggest that plasmodial proteases are attractive targetsfor new antimalarial chemotherapeutics.
Plasmepsins, a family of aspartic proteases of the Plasmodiumspecies, have been well characterized in P. falciparum. A total of10 genes that encode plasmepsins or related enzymes have beenidentified in the parasite (Coombs et al., 2001). Four of them(PfPM1, PfPM2, PfHAP and PfPM4) are localized in the food vacuoleof P. falciparum and play important roles in hemoglobin hydrolysis(Francis et al., 1994; Banerjee et al., 2002). Meanwhile, the others(PfPM5–PfPM10) seem to be expressed in other regions of theasexual erythrocytic stage parasite or in different stages of the par-asite’s life cycle (Banerjee et al., 2002). BLAST searches of P. vivaxgenome sequence database using amino acid sequences of eachof the P. falciparum plasmepsins as the queries suggest that P. vivaxhas 7 orthologues of plasmepsins, PfPM4–PfPM10, but the ortho-logues of PfPM1, PfPM2 and PfHAP were not identified (Dameet al., 2003). An in silico study also suggests that PfPM4 orthologueis the only putative food vacuole plasmepsin in non-falciparummalaria species (Dame et al., 2003). These suggest that plasmepsin4 of P. vivax (PvPM4) may be a plausible food vacuole plasmepsinof P. vivax. PvPM4 has been partially characterized by several

128 S.-U. Moon et al. / Experimental Parasitology 128 (2011) 127–132
research groups, but these studies have mainly focused on thestructural properties and substrate specificity of the enzyme (Wes-tling et al., 1997; Bernstein et al., 2003; Li et al., 2004; Beyer et al.,2005). As such, its biological and biochemical properties still re-main to be characterized.
In this study, we characterized the processing event of PvPM4as well as its hemoglobin hydrolysis property in order to expandour understanding of the enzyme. Processing of PvPM4 occurredvia autocatalytic activity under acidic conditions in which a por-tion of the prodomain was required for correct folding. PvPM4showed collaborative hemoglobin hydrolysis activity with cysteineproteases of P. vivax, vivapain-2 (VX-2) and vivapain-3 (VX-3), atacidic pHs and hemoglobin hydrolysis activity by the enzymeswas increased under reduced conditions. Moreover, both VX-2and VX-3 were shown to mediate the processing of PvPM4. Takentogether, our results demonstrate that PvPM4 is an active hemo-globinase of P. vivax that works collaboratively with vivapains toimprove hemoglobin hydrolysis by the parasite.
2. Materials and methods
2.1. Expression and purification of recombinant PvPM4
The gene for PvPM4 (Gene ID: PVX_086040) was amplified bypolymerase chain reaction (PCR) using 200 ng of P. vivax genomicDNA, Ex Taq DNA polymerase (Takara, Otsu, Japan) and oligonu-cleotide primers (50-ATGGATATAGCAGTGAAAGAA-30 and 50-TTAATTCTTTGCGATGGCAAA-30). The amplified product of 1353 bp in sizewas gel-purified, cloned into pGEM-T Easy vector (Promega, Mad-ison, WI, USA), and transformed into competent Escherichia coliDH5a cells. Nucleotide sequence of the cloned gene was analyzedby ABI 377 automatic DNA sequencer (Applied Biosystems, FosterCity, CA, USA). To make expression construct of PvPM4, a fragmentharboring a portion of the prodomain and the entire mature do-main of PvPM4 was amplified using the primers 50-GGATCCCAAAACACGGAACATTTAACTTTA-30, which contained a 50 BamHI site,and 50-CTGCAGTTAATTCTTTGCGATGGCAAAACC-30, which har-bored a 50 Pst I site. The PCR product was gel-purified, ligated intothe pGEM-T Easy vector (Promega), and transformed into E. coliDH5a. The resulting plasmid DNA was digested with the appropri-ate restriction enzymes and ligated into the pQE-30 expressionvector (Qiagen, Valencia, CA, USA), which was then transformedinto E. coli M15 [pREP4] cells (Qiagen). Selected clones were grownand induced with 1 mM isopropyl-1-thio-b-D-galactopyranoside(IPTG) for 3 h at 37 �C. The cells were collected by centrifugationat 10,000g for 15 min at 4 �C, and suspended in 8 M urea lysis buf-fer. The recombinant protein was purified by nickel–nitrilotriaceticacid (Ni–NTA) affinity chromatography (Qiagen) followed bymanufacturer’s instructions. The purification and purity of therecombinant protein was analyzed by sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE).
2.2. Refolding and activation of PvPM4
To determine the optimal refolding condition, the purifiedPvPM4 was refolded in different refolding conditions. In brief,10 ll of the purified protein (1 mg/ml) was slowly added to290 ll of different pH buffers, 100 mM sodium acetate (pH 5.0),100 mM sodium phosphate (pH 6.0), 100 mM Tris–HCl (pH 7.0),100 mM Tris–HCl (pH 8.0), or 100 mM Glycine–NaOH (pH 9.0)and the samples were incubated at 4 �C for overnight with gentleshaking. The enzyme activity was assayed by using the fluorogenicpeptide substrate, Ac–Glu–Asp(EDANS)–Lys–Pro–Ile–Leu–Phe–Phe–Arg–Leu–Gly–Lys(DABCYL)–Glu–NH2 (Bachem, Bubendorf,Switzerland). Briefly, 10 ll of refolded enzyme solution was added
to 140 ll of sodium acetate buffer (pH 4.5) containing 5 lM fluoro-genic substrate and the release of fluorescence (excitation, 405 nm;emission, 510 nm) over 20 min at room temperature was assessedwith a Fluoroskan Ascent FL (Thermo, Vantaa, Finland). Activationof refolded PvPM4 was carried out in different pH buffers; 100 mMsodium acetate (pH 4.0–5.0), 100 mM sodium phosphate (pH 6.0),and 100 mM Tris–HCl (pH 7.0–8.0). The refolded PvPM4 was addedto each activation buffer and incubated at 37 �C. Aliquot was takenat indicated time and analyzed by SDS–PAGE or measured the en-zyme activity as the same method described above. For large-scalepreparation of PvPM4, 10 mg of Ni–NTA affinity purified PvPM4was added to refolding buffer, 100 mM Tris–HCl (pH 8.0), in a dilu-tion factor of 200 and incubated at 4 �C for overnight with gentlestirring. The refolded sample was dialyzed against 10 mM Tris–HCl (pH 7.5) and concentrated with Centricon Plus (cut-off10 kDa; Millipore, Billerica, MA, USA). To allow processing to activeenzyme, the pH of the refolded sample was adjusted to 4.5 with3.5 M sodium acetate (pH 2.6) and the sample was incubated at37 �C for 1 h. The precipitated material was removed by centrifuga-tion at 12,000g for 15 min at 4 �C, and the sample was dialyzedagainst 10 mM Tris–HCl (pH 7.0) and concentrated with Centriprep(cut-off 10 kDa, Millipore).
2.3. N-terminal amino acid sequencing
Fully processed mature PvPM4 was separated by SDS–PAGE,transferred to a polyvinylidene difluoride (PVDF) membrane,stained with Coomassie blue, and destained. The protein bandwas excised and sequenced by Edman sequencing at the Korea Ba-sic Science Institute (Daejeon, Korea).
2.4. Optimal pH and stability
The optimal pH for the maximum activity of fully maturedPvPM4 was assessed in sodium acetate (pH 3.0–5.5), sodium phos-phate (pH 6.0–6.5), and Tris–HCl (pH 7.0). The PvPM4 (50 nM) wasadded to each pH buffer supplemented with 5 lM Ac–Glu–Asp(EDANS)–Lys–Pro–Ile–Leu–Phe–Phe–Arg–Leu–Gly–Lys(DABCYL)–Glu–NH2 (Bachem) and the enzyme activity was measured asdescribed above. For each pH step, the appropriate blank wasseparately measured as a control. The stability of PvPM4 was alsoexamined at pH 5.0 or 7.0 by incubating the enzyme at 37 or 50 �Cin the appropriate buffers. Aliquot was taken at indicated timeintervals and the residual enzyme activity was measured.
2.5. Requirement of prodomain for correct folding of PvPM4
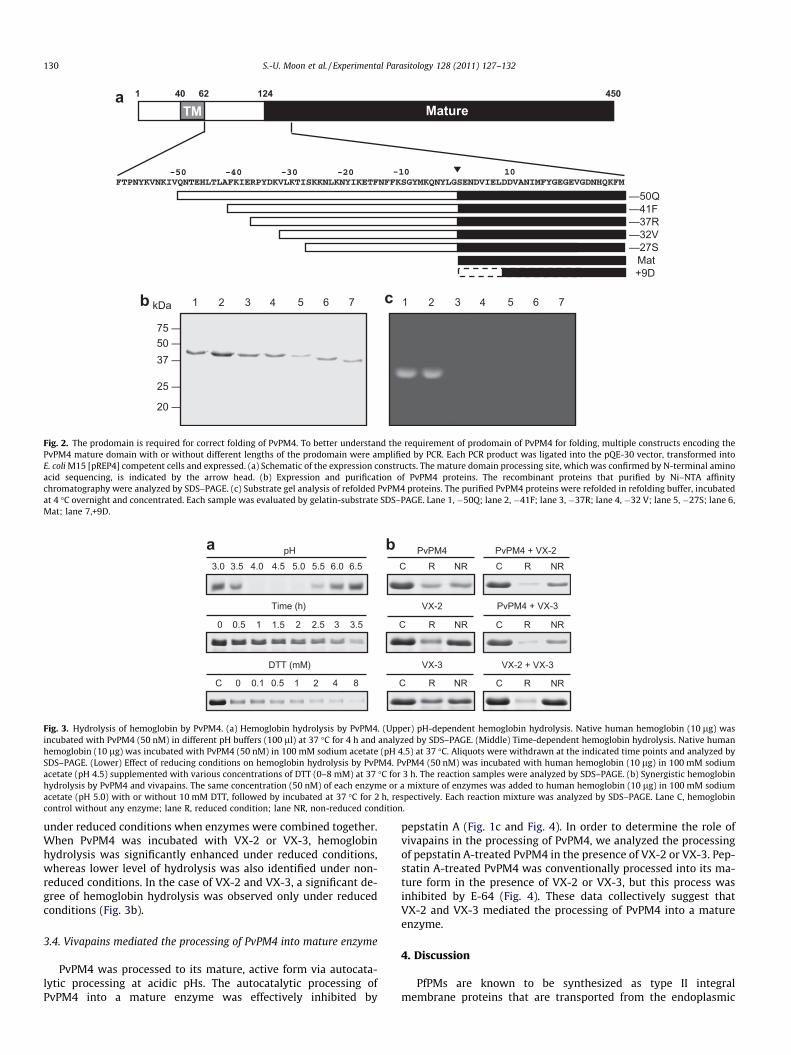
To characterize the requirement of the prodomain for the cor-rect folding of PvPM4, multiple constructs encoding for the PvPM4mature domain and different length of the prodomain were ampli-fied by PCR using the following primers: �50Q EF (50-GGATCC-CAAAACACGGAACATTTAACTTTA-30), �41F EF (50-GGATCCTTTAAAATCGAGAGACCCTATGAC-30), �37 R EF (50-GGATCCAGACCCTATGACAAGGTACTAAAA-30), �32V EF (50-GGATCCGTACTAAAAACGATATCCAAGAAA-30) �27S EF (50-GGATCCTCCAAGAAAAACCTGAAGAATTAC-30), Mat EF (50-GGATCCAGTGAAAATGATGTGATCGAATTA-30), and +9D EF (50-GGATCCGATGATGCAAACATTATGTTCTAT-30); in all cases the reverse primer was 50-CTGCAGTTAATTCTTTGCGATGGCAAAACC-30. The DNA fragments were digested with BamHIand Pst I, ligated into pQE-30 expression vector (Qiagen), andtransformed into E. coli M15 [pREP4] cells. Each protein was ex-pressed, purified, and refolded as described above. The enzymeactivity of each refolded protein was analyzed as described above.Substrate gel analysis was carried out as described previously (Naet al., 2004).
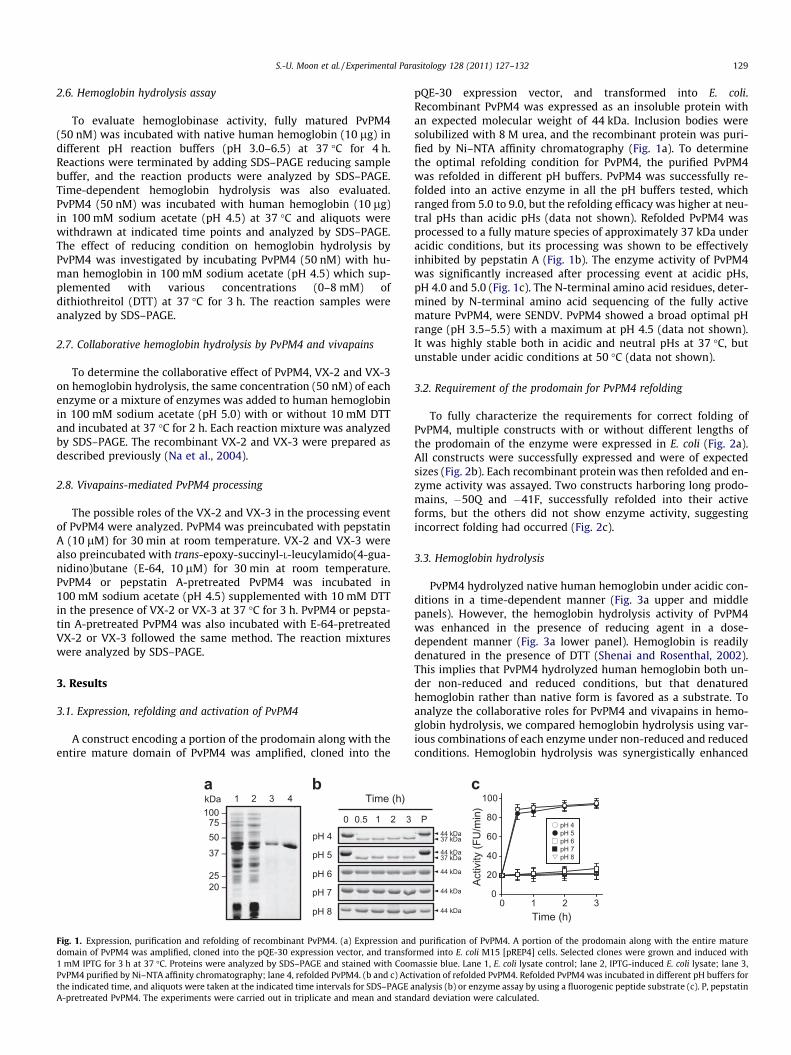
S.-U. Moon et al. / Experimental Parasitology 128 (2011) 127–132 129
2.6. Hemoglobin hydrolysis assay
To evaluate hemoglobinase activity, fully matured PvPM4(50 nM) was incubated with native human hemoglobin (10 lg) indifferent pH reaction buffers (pH 3.0–6.5) at 37 �C for 4 h.Reactions were terminated by adding SDS–PAGE reducing samplebuffer, and the reaction products were analyzed by SDS–PAGE.Time-dependent hemoglobin hydrolysis was also evaluated.PvPM4 (50 nM) was incubated with human hemoglobin (10 lg)in 100 mM sodium acetate (pH 4.5) at 37 �C and aliquots werewithdrawn at indicated time points and analyzed by SDS–PAGE.The effect of reducing condition on hemoglobin hydrolysis byPvPM4 was investigated by incubating PvPM4 (50 nM) with hu-man hemoglobin in 100 mM sodium acetate (pH 4.5) which sup-plemented with various concentrations (0–8 mM) ofdithiothreitol (DTT) at 37 �C for 3 h. The reaction samples wereanalyzed by SDS–PAGE.
2.7. Collaborative hemoglobin hydrolysis by PvPM4 and vivapains
To determine the collaborative effect of PvPM4, VX-2 and VX-3on hemoglobin hydrolysis, the same concentration (50 nM) of eachenzyme or a mixture of enzymes was added to human hemoglobinin 100 mM sodium acetate (pH 5.0) with or without 10 mM DTTand incubated at 37 �C for 2 h. Each reaction mixture was analyzedby SDS–PAGE. The recombinant VX-2 and VX-3 were prepared asdescribed previously (Na et al., 2004).
2.8. Vivapains-mediated PvPM4 processing
The possible roles of the VX-2 and VX-3 in the processing eventof PvPM4 were analyzed. PvPM4 was preincubated with pepstatinA (10 lM) for 30 min at room temperature. VX-2 and VX-3 werealso preincubated with trans-epoxy-succinyl-L-leucylamido(4-gua-nidino)butane (E-64, 10 lM) for 30 min at room temperature.PvPM4 or pepstatin A-pretreated PvPM4 was incubated in100 mM sodium acetate (pH 4.5) supplemented with 10 mM DTTin the presence of VX-2 or VX-3 at 37 �C for 3 h. PvPM4 or pepsta-tin A-pretreated PvPM4 was also incubated with E-64-pretreatedVX-2 or VX-3 followed the same method. The reaction mixtureswere analyzed by SDS–PAGE.
3. Results
3.1. Expression, refolding and activation of PvPM4
A construct encoding a portion of the prodomain along with theentire mature domain of PvPM4 was amplified, cloned into the
a b1 2 3 4
50 –
37 –
25 –
75 –100 –
20 –
kDa
pH 4
pH 5
pH 6
pH 7
pH 8
0 0.5 1 2 3
Time (h)
Fig. 1. Expression, purification and refolding of recombinant PvPM4. (a) Expression anddomain of PvPM4 was amplified, cloned into the pQE-30 expression vector, and transfo1 mM IPTG for 3 h at 37 �C. Proteins were analyzed by SDS–PAGE and stained with CoomPvPM4 purified by Ni–NTA affinity chromatography; lane 4, refolded PvPM4. (b and c) Actthe indicated time, and aliquots were taken at the indicated time intervals for SDS–PAGE aA-pretreated PvPM4. The experiments were carried out in triplicate and mean and stan
pQE-30 expression vector, and transformed into E. coli.Recombinant PvPM4 was expressed as an insoluble protein withan expected molecular weight of 44 kDa. Inclusion bodies weresolubilized with 8 M urea, and the recombinant protein was puri-fied by Ni–NTA affinity chromatography (Fig. 1a). To determinethe optimal refolding condition for PvPM4, the purified PvPM4was refolded in different pH buffers. PvPM4 was successfully re-folded into an active enzyme in all the pH buffers tested, whichranged from 5.0 to 9.0, but the refolding efficacy was higher at neu-tral pHs than acidic pHs (data not shown). Refolded PvPM4 wasprocessed to a fully mature species of approximately 37 kDa underacidic conditions, but its processing was shown to be effectivelyinhibited by pepstatin A (Fig. 1b). The enzyme activity of PvPM4was significantly increased after processing event at acidic pHs,pH 4.0 and 5.0 (Fig. 1c). The N-terminal amino acid residues, deter-mined by N-terminal amino acid sequencing of the fully activemature PvPM4, were SENDV. PvPM4 showed a broad optimal pHrange (pH 3.5–5.5) with a maximum at pH 4.5 (data not shown).It was highly stable both in acidic and neutral pHs at 37 �C, butunstable under acidic conditions at 50 �C (data not shown).
3.2. Requirement of the prodomain for PvPM4 refolding
To fully characterize the requirements for correct folding ofPvPM4, multiple constructs with or without different lengths ofthe prodomain of the enzyme were expressed in E. coli (Fig. 2a).All constructs were successfully expressed and were of expectedsizes (Fig. 2b). Each recombinant protein was then refolded and en-zyme activity was assayed. Two constructs harboring long prodo-mains, �50Q and �41F, successfully refolded into their activeforms, but the others did not show enzyme activity, suggestingincorrect folding had occurred (Fig. 2c).
3.3. Hemoglobin hydrolysis
PvPM4 hydrolyzed native human hemoglobin under acidic con-ditions in a time-dependent manner (Fig. 3a upper and middlepanels). However, the hemoglobin hydrolysis activity of PvPM4was enhanced in the presence of reducing agent in a dose-dependent manner (Fig. 3a lower panel). Hemoglobin is readilydenatured in the presence of DTT (Shenai and Rosenthal, 2002).This implies that PvPM4 hydrolyzed human hemoglobin both un-der non-reduced and reduced conditions, but that denaturedhemoglobin rather than native form is favored as a substrate. Toanalyze the collaborative roles for PvPM4 and vivapains in hemo-globin hydrolysis, we compared hemoglobin hydrolysis using var-ious combinations of each enzyme under non-reduced and reducedconditions. Hemoglobin hydrolysis was synergistically enhanced
c
0 1 2 30
20
40
60
80
100
Time (h)
Activ
ity (F
U/m
in)
pH 4pH 5pH 6pH 7pH 8
P44 kDa37 kDa
44 kDa
44 kDa
44 kDa
44 kDa37 kDa
purification of PvPM4. A portion of the prodomain along with the entire maturermed into E. coli M15 [pREP4] cells. Selected clones were grown and induced with
assie blue. Lane 1, E. coli lysate control; lane 2, IPTG-induced E. coli lysate; lane 3,ivation of refolded PvPM4. Refolded PvPM4 was incubated in different pH buffers fornalysis (b) or enzyme assay by using a fluorogenic peptide substrate (c). P, pepstatin
dard deviation were calculated.
50 –37 –
25 –
75 –
20 –
kDa 1 2 3 4 5 6 7 cb
a 1 45040 62 124
–50Q–41F–37R–32V–27S
Mat+9D
MatureTM
1 2 3 4 5 6 7
Fig. 2. The prodomain is required for correct folding of PvPM4. To better understand the requirement of prodomain of PvPM4 for folding, multiple constructs encoding thePvPM4 mature domain with or without different lengths of the prodomain were amplified by PCR. Each PCR product was ligated into the pQE-30 vector, transformed intoE. coli M15 [pREP4] competent cells and expressed. (a) Schematic of the expression constructs. The mature domain processing site, which was confirmed by N-terminal aminoacid sequencing, is indicated by the arrow head. (b) Expression and purification of PvPM4 proteins. The recombinant proteins that purified by Ni–NTA affinitychromatography were analyzed by SDS–PAGE. (c) Substrate gel analysis of refolded PvPM4 proteins. The purified PvPM4 proteins were refolded in refolding buffer, incubatedat 4 �C overnight and concentrated. Each sample was evaluated by gelatin-substrate SDS–PAGE. Lane 1, �50Q; lane 2, �41F; lane 3, �37R; lane 4, �32 V; lane 5, �27S; lane 6,Mat; lane 7,+9D.
ba pH3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5
Time (h)
0 0.5 1 1.5 2 2.5 3 3.5
0 0.1 0.5 1 2 4 8C
DTT (mM)
C R NRPvPM4
VX-2
VX-3
PvPM4 + VX-2
PvPM4 + VX-3
VX-2 + VX-3
C R NR
C R NR C R NR
C R NR C R NR
Fig. 3. Hydrolysis of hemoglobin by PvPM4. (a) Hemoglobin hydrolysis by PvPM4. (Upper) pH-dependent hemoglobin hydrolysis. Native human hemoglobin (10 lg) wasincubated with PvPM4 (50 nM) in different pH buffers (100 ll) at 37 �C for 4 h and analyzed by SDS–PAGE. (Middle) Time-dependent hemoglobin hydrolysis. Native humanhemoglobin (10 lg) was incubated with PvPM4 (50 nM) in 100 mM sodium acetate (pH 4.5) at 37 �C. Aliquots were withdrawn at the indicated time points and analyzed bySDS–PAGE. (Lower) Effect of reducing conditions on hemoglobin hydrolysis by PvPM4. PvPM4 (50 nM) was incubated with human hemoglobin (10 lg) in 100 mM sodiumacetate (pH 4.5) supplemented with various concentrations of DTT (0–8 mM) at 37 �C for 3 h. The reaction samples were analyzed by SDS–PAGE. (b) Synergistic hemoglobinhydrolysis by PvPM4 and vivapains. The same concentration (50 nM) of each enzyme or a mixture of enzymes was added to human hemoglobin (10 lg) in 100 mM sodiumacetate (pH 5.0) with or without 10 mM DTT, followed by incubated at 37 �C for 2 h, respectively. Each reaction mixture was analyzed by SDS–PAGE. Lane C, hemoglobincontrol without any enzyme; lane R, reduced condition; lane NR, non-reduced condition.
130 S.-U. Moon et al. / Experimental Parasitology 128 (2011) 127–132
under reduced conditions when enzymes were combined together.When PvPM4 was incubated with VX-2 or VX-3, hemoglobinhydrolysis was significantly enhanced under reduced conditions,whereas lower level of hydrolysis was also identified under non-reduced conditions. In the case of VX-2 and VX-3, a significant de-gree of hemoglobin hydrolysis was observed only under reducedconditions (Fig. 3b).
3.4. Vivapains mediated the processing of PvPM4 into mature enzyme
PvPM4 was processed to its mature, active form via autocata-lytic processing at acidic pHs. The autocatalytic processing ofPvPM4 into a mature enzyme was effectively inhibited by
pepstatin A (Fig. 1c and Fig. 4). In order to determine the role ofvivapains in the processing of PvPM4, we analyzed the processingof pepstatin A-treated PvPM4 in the presence of VX-2 or VX-3. Pep-statin A-treated PvPM4 was conventionally processed into its ma-ture form in the presence of VX-2 or VX-3, but this process wasinhibited by E-64 (Fig. 4). These data collectively suggest thatVX-2 and VX-3 mediated the processing of PvPM4 into a matureenzyme.
4. Discussion
PfPMs are known to be synthesized as type II integralmembrane proteins that are transported from the endoplasmic

50 –37 –
25 –
75 –
150 –
15 –
kDa
PvPM4PepstatinAVX-2VX-3E-64
++–––
+–+––
+––+–
+++––
++–+–
+++–+
++–++
+––––C
Fig. 4. Vivapains mediated the processing of PvPM4. PvPM4 or pepstatin A-pretreated PvPM4 was incubated in 100 mM sodium acetate (pH 4.5) supplementedwith 10 mM DTT with or without VX-2 or VX-3. E-64 pretreated VX-2 or VX-3 wasalso analyzed following the same method. The reaction mixtures were analyzed bySDS–PAGE. C, unprocessed PvPM4 control.
S.-U. Moon et al. / Experimental Parasitology 128 (2011) 127–132 131
reticulum to the cytostomes via vesicles, after which they aretransported along with their substrate hemoglobin via cyto-stome-derived vesicles to the food vacuole (Francis et al., 1997;Klemba et al., 2004a,b; Klonis et al., 2007). Once in the acidic foodvacuole, PfPMs are activated and released from the membrane byproteolytic cleavage at a conserved sequence site (Y/H)LG;(S/N)XXD (Francis et al., 1997; Banerjee et al., 2003; Klemba et al.,2004b). Recombinant PfPMs readily undergoes autocatalytic pro-cessing under acidic conditions in both cis- and trans-catalyticfashions (Kim et al., 2006). PvPM4 is also a typical aspartic proteasewith a large prodomain and type II integral transmembrane do-main at its N-terminus. In this study, the bacterially expressed re-combinant protein was obtained as an insoluble protein, but waseasily refolded into a soluble form under a broad pH ranges from5.0 to 9.0. The refolded PvPM4 was rapidly processed into a matureenzyme via autocatalytic cleavage at NYLG;SEND under acidic con-ditions in a pepstatin A-sensitive manner, during which a portionof the prodomain was essential for correct folding. Due to the tech-nical shortcomings of P. vivax stemming from the lack of in vitrocultivation method, it is difficult to determine the trafficking andprocessing progresses of PvPM4 in the parasite. However, consid-ering that PvPM4 shares highly comparable structural and process-ing properties with PfPMs in vitro, it also likely undergoes similartrafficking and processing events as PfPMs in vivo. However, thisshould be elucidated further.
PvPM4 hydrolyzed human hemoglobin under acidic conditions,but favored denatured hemoglobin rather than native form as asubstrate. These results suggest that the principle function of theenzyme is hemoglobin hydrolysis in the acidic food vacuole ofthe parasite, which supports the notion that PvPM4 is a uniquefood vacuole plasmepsin of P. vivax (Dame et al., 2003). Hemoglo-bin hydrolysis in P. falciparum is well characterized. Two families ofproteases, three cysteine protease falcipains (FP-2, FP-2’ and FP-3)and four aspartic protease plasmepsins (PfPM1, PfPM2, HAP andPfPM4), are critically involved in the initial steps of hemoglobincatabolism in the food vacuole of the parasite (Francis et al.,1994; Shenai et al., 2000; Sijwali et al., 2001; Banerjee et al.,2002; Singh et al., 2006). Although an ordered pathway of hemo-globin hydrolysis has not yet been fully understood, several recentgene knockout studies strongly suggest the presence of substantialfunctional redundancy within and between the enzyme familiesregarding their essential and compensable roles in hemoglobinhydrolysis (Omara-Opyene et al., 2004; Liu et al., 2005; Sijwaliet al., 2001,2006; Bonilla et al., 2007a,b). P. vivax is the mostwidespread Plasmodium species around the world and the second
medically important human malaria parasite (Mendis et al.,2001). However, the biology and biochemistry of hemoglobinhydrolysis by this parasite are poorly understood. Two cysteineproteases, VX-2 and VX-3, which are the apparent orthologues ofFP-2 and FP-3 of P. falciparum, respectively, were identified as ac-tive hemoglobinases of P. vivax (Na et al., 2004). Both VX-2 andVX-3 hydrolyzed hemoglobin at acidic pH, suggesting that theirprincipal function is hemoglobin hydrolysis in the food vacuoleof P. vivax. An aspartic protease with hemoglobin hydrolysis activ-ity at acidic pH has been purified from P. vivax and its biochemicalproperties have been partially characterized (Sharma et al., 2005).However, it was not clear if the purified enzyme is PvPM4. In thisstudy, we analyzed the collaborative roles of PvPM4 and vivapainsin hemoglobin hydrolysis. Hemoglobin hydrolysis was enhancedby PvPM4 and vivapains, suggesting both classes of enzymes workcollaboratively to improve hemoglobin hydrolysis in P. vivax. Wealso found that the processing of PvPM4 was mediated by VX-2and VX-3. In the presence of pepstatin A, the autocalytic processingof PvPM4 into a mature enzyme was effectively inhibited. How-ever, this process was recovered by VX-2 and VX-3, suggesting thatVX-2 and VX-3 could mediate PvPM4 maturation. Similar resultswere recently reported in P. falciparum. PfPM2 processing in thefood vacuole is primarily mediated by falcipains. However, auto-catalytic processing of PfPM2 occurs as an alternate system of acti-vation if falcipain activity is blocked, which strongly suggests therole of falcipains as PfPM maturase (Drew et al., 2008). VX-4, thefourth cysteine protease of P. vivax is also known to mediate mat-uration of PvPM4 (Na et al., 2010). Our result also showed that pro-cessing of PvPM4 by vivapains to the size of the mature enzymeproduced due to autoprocessing. We attempted N-terminal aminoacid sequencing analysis of PvPM4 processed by vivapains, butunfortunately we could not obtain clear result. However, as evi-dence by similar molecular size to autoprocessed PvPM4, the pro-cessing site of PvPM4 by vivapains could be similar or identical tothat of autoprocessed PvPM4. It raised a perplexing question whyvivapain-mediated PvPM4 processing is occurred at a specific siteand PvPM4 is protected from hydrolysis by vivapains, pretty pro-miscuous enzymes having multiple cleavage sites? The study toanswer this question is now underway.
In conclusion, here we characterized the processing event ofPvPM4 as well as principal role of the enzyme in hemoglobin hydro-lysis. Collaborative roles for PvPM4 and vivapains in hemoglobinhydrolysis confirm that cysteine and aspartic proteases of P. vivaxcontribute to the improvement of hemoglobin hydrolysis in theparasite. Our finding that VX-2 and VX-3 mediate the processingof PvPM4 also suggests a level of functional redundancy betweenPvPM4 and vivapains in P. vivax hemoglobin hydrolysis. Our studyallows further advances to be made in the understanding of hemo-globin hydrolysis in P. vivax, but further comprehensive studies arestill necessary for in-depth understanding of the event.
Acknowledgment
This study was supported by grants of the Korea Centers for Dis-ease Control and Prevention (2008-E00151-00) and the KoreaHealth 21 R&D Project, Ministry of Health and Welfare, the Repub-lic of Korea (03-PJ1-PG1-CH01-0001).
References
Banerjee, R., Liu, J., Beatty, W., Pelosof, L., Klemba, M., Goldberg, D.E., 2002. Fourplasmepsins are active in the Plasmodium falciparum food vacuole, including aprotease with an active-site histidine. Proceedings of the National Academy ofSciences of the United States of America 99, 990–995.
Banerjee, R., Francis, S.E., Goldberg, D.E., 2003. Food vacuole plasmepsins areprocessed at a conserved site by an acidic convertase activity in Plasmodiumfalciparum. Molecular and Biochemical Parasitology 129, 157–165.

132 S.-U. Moon et al. / Experimental Parasitology 128 (2011) 127–132
Bernstein, N.K., Cherney, M.M., Yowell, C.A., Dame, J.B., James, M.N., 2003. Structuralinsights into the activation of Plasmodium vivax plasmepsin. Journal ofMolecular Biology 329, 505–524.
Beyer, B.B., Johnson, J.V., Chung, A.Y., Li, T., Madabushi, A., Agbandje-McKenna, M.,McKenna, R., Dame, J.B., Dunn, B.M., 2005. Active-site specificity of digestiveaspartic peptidases from the four species of Plasmodium that infect humansusing chromogenic combinatorial peptide libraries. Biochemistry 44, 1768–1779.
Bonilla, J.A., Bonilla, T.D., Yowell, C.A., Fujioka, H., Dame, J.B., 2007a. Critical roles forthe digestive vacuole plasmepsins of Plasmodium falciparum in vacuolarfunction. Molecular Microbiology 65, 64–75.
Bonilla, J.A., Moura, P.A., Bonilla, T.D., Yowell, C.A., Fidock, D.A., Dame, J.B., 2007b.Effects on growth, hemoglobin metabolism and paralogous gene expressionresulting from disruption of genes encoding the digestive vacuole plasmepsinsof Plasmodium falciparum. International Journal for Parasitology 37, 317–327.
Coombs, G.H., Goldberg, D.E., Klemba, M., Berry, C., Kay, J., Mottram, J.C., 2001.Aspartic proteases of Plasmodium falciparum and other parasitic protozoa asdrug targets. Trends in Parasitology 17, 532–537.
Dame, J.B., Yowell, C.A., Omara-Opyene, A.L., Carlton, J.M., Cooper, R.A., Li, T., 2003.Plasmepsin 4 the food vacuole aspartic proteinase found in all Plasmodium sppinfecting man. Molecular and Biochemical Parasitology 130, 1–12.
Drew, M.E., Banerjee, R., Uffman, E.W., Gilbertson, S., Rosenthal, P.J., Goldberg, D.E.,2008. Plasmodium food vacuole plasmepsins are activated by falcipains. TheJournal of Biological Chemistry 283, 12870–12876.
Eggleson, K.K., Duffin, K.L., Goldberg, D.E., 1999. Identification and characterizationof falcilysin, a metallopeptidase involved in hemoglobin catabolism within themalaria parasite Plasmodium falciparum. The Journal of Biological Chemistry274, 32411–32417.
Francis, S.E., Gluzman, I.Y., Oksman, A., Knickerbocker, A., Mueller, R., Bryant, M.,Sherman, D.R., Russell, D.G., Goldberg, D.E., 1994. Molecular characterizationand inhibition of a Plasmodium falciparum aspartic hemoglobinase. EMBOJournal 13, 306–317.
Francis, S.E., Banerjee, R., Goldberg, D.E., 1997. Biosynthesis and maturation of themalaria aspartic hemoglobinases plasmepsins I and II. The Journal of BiologicalChemistry 272, 14961–14968.
Jiang, S.P., Prigge, S.T., Wei, L., Gao, Y.E., Hudson, T.H., Gerena, L., Dame, J.B., Kyle,D.E., 2001. New class of small nonpeptidyl compounds blocks Plasmodiumfalciparum development in vitro by inhibiting plasmepsins. AntimicrobialAgents and Chemotherapy 45, 2577–2584.
Kim, Y.M., Lee, M.H., Piao, T.G., Lee, J.W., Kim, J.H., Lee, S., Choi, K.M., Jiang, J.H., Kim,T.U., Park, H., 2006. Prodomain processing of recombinant plasmepsin II and IV,the aspartic proteases of Plasmodium falciparum, is auto- and trans-catalytic.The Journal of Biochemistry 139, 189–195.
Klemba, M., Gluzman, I., Goldberg, D.E., 2004a. A Plasmodium falciparum dipeptidylaminopeptidase I participates in vacuolar hemoglobin degradation. The Journalof Biological Chemistry 279, 43000–43007.
Klemba, M., Beatty, W., Gluzman, I., Goldberg, D.E., 2004b. Trafficking of plasmepsinII to the food vacuole of the malaria parasite Plasmodium falciparum. Journal ofCell Biology 164, 47–56.
Klonis, N., Tan, O., Jackson, K., Goldberg, D., Klemba, M., Tilley, L., 2007. Evaluation ofpH during cytostomal endocytosis and vacuolar catabolism of haemoglobin inPlasmodium falciparum. Biochemical Journal 407, 343–354.
Krugliak, M., Zhang, J., Ginsburg, H., 2002. Intraerythrocytic Plasmodium falciparumutilizes only a fraction of the amino acids derived from the digestion of host cellcytosol for the biosynthesis of its proteins. Molecular and BiochemicalParasitology 119, 249–256.
Lew, V.L., Tiffert, T., Ginsburg, H., 2003. Excess hemoglobin digestion and theosmotic stability of Plasmodium falciparum-infected red blood cells. Blood 101,4189–4194.
Lew, V.L., Macdonald, L., Ginsburg, H., Krugliak, M., Tiffert, T., 2004. Excesshaemoglobin digestion by malaria parasites: a strategy to prevent prematurehost cell lysis. Blood Cells, Molecules and Diseases 32, 353–359.
Li, T., Yowell, C.A., Beyer, B.B., Hung, S.H., Westling, J., Lam, M.T., Dunn, B.M., Dame,J.B., 2004. Recombinant expression and enzymatic subsite characterization ofplasmepsin 4 from the four Plasmodium species infecting man. Molecular andBiochemical Parasitology 135, 101–109.
Liu, J., Gluzman, I.Y., Drew, M.E., Goldberg, D.E., 2005. The role of Plasmodiumfalciparum food vacuole plasmepsins. The Journal of Biological Chemistry 280,1432–1437.
Mendis, K., Sina, B.J., Marchesini, P., Carter, R., 2001. The neglected burden ofPlasmodium vivax malaria. American Journal of Tropical Medicine and Hygiene64, 97–106.
Moura, P.A., Dame, J.B., Fidock, D.A., 2009. Role of Plasmodium falciparum digestivevacuole plasmepsins in the specificity and antimalarial mode of action ofcysteine and aspartic protease inhibitors. Antimicrobial Agents andChemotherapy 53, 4968–4978.
Na, B.K., Shenai, B.R., Sijwali, P.S., Choe, Y., Pandey, K.C., Singh, A., Craik, C.S.,Rosenthal, P.J., 2004. Identification and biochemical characterization ofvivapains, cysteine proteases of the malaria parasite Plasmodium vivax.Biochemical Journal 378, 529–538.
Na, B.K., Bae, Y.A., Zo, Y.G., Choe, Y., Kim, S.H., Desai, P.V., Avery, M.A., Craik, C.S.,Kim, T.S., Rosenthal, P.J., Kong, Y., 2010. Biochemical properties of a novelcysteine protease of Plasmodium vivax, vivapain-4. PLoS Neglected TropicalDiseases 4, e849.
Olson, J.E., Lee, G.K., Semenov, A., Rosenthal, P.J., 1999. Antimalarial effects in miceof orally administered peptidyl cysteine protease inhibitors. Bioorganic andMedicinal Chemistry 7, 633–638.
Omara-Opyene, A.L., Moura, P.A., Sulsona, C.R., Bonilla, J.A., Yowell, C.A., Fujioka, H.,Fidock, D.A., Dame, J.B., 2004. Genetic disruption of the Plasmodium falciparumdigestive vacuole plasmepsins demonstrates their functional redundancy. TheJournal of Biological Chemistry 279, 54088–54096.
Rosenthal, P.J., Wollish, W.S., Palmer, J.T., Rasnick, D., 1991. Antimalarial effects ofpeptide inhibitors of a Plasmodium falciparum cysteine proteinase. Journal ofClinical Investigation 88, 1467–1472.
Rosenthal, P.J., 2002. Hydrolysis of erythoctye proteins by proteases of malariaparasites. Current Opinion in Hematology 9, 140–145.
Sharma, A., Eapen, A., Subbarao, S.K., 2005. Purification and characterization of ahemoglobin degrading aspartic protease from the malarial parasite Plasmodiumvivax. Journal of Biochemistry 138, 71–78.
Shenai, B.R., Sijwali, P.S., Singh, A., Rosenthal, P.J., 2000. Characterization of nativeand recombinant falcipain-2, a principal trophozoite cysteine protease andessential hemoglobinase of Plasmodium falciparum. The Journal of BiologicalChemistry 275, 29000–29010.
Shenai, B.R., Rosenthal, P.J., 2002. Reducing requirements for hemoglobin hydrolysisby Plasmodium falciparum cysteine proteases. Molecular and BiochemicalParasitology 122, 99–104.
Sijwali, P.S., Shenai, B.R., Gut, J., Singh, A., Rosenthal, P.J., 2001. Expression andcharacterization of the Plasmodium falciparum haemoglobinase falcipain-3.Biochemical Journal 360, 481–489.
Sijwali, P.S., Rosenthal, P.J., 2004. Gene disruption confirms a critical role for thecysteine protease falcipain-2 in hemoglobin hydrolysis by Plasmodiumfalciparum. Proceedings of the National Academy of Sciences of the UnitedStates of America 101, 4384–4389.
Sijwali, P.S., Koo, J., Singh, N., Rosenthal, P.J., 2006. Gene disruptionsdemonstrate independent roles for the four falcipain cysteine proteases ofPlasmodium falciparum. Molecular and Biochemical Parasitology 150, 96–106.
Singh, N., Sijwali, P.S., Pandey, K.C., Rosenthal, P.J., 2006. Plasmodium falciparum:biochemical characterization of the cysteine protease falcipain-2’. ExperimentalParasitology 112, 187–192.
Westling, J., Yowell, C.A., Majer, P., Erickson, J.W., Dame, J.B., Dunn, B.M., 1997.Plasmodium falciparum, P. Vivax, and P. malariae: a comparison of the active siteproperties of plasmepsins cloned and expressed from three different species ofthe malaria parasite. Experimental Parasitology 87, 185–193.





























