Embed Size (px)
Citation preview

ORIGINAL PAPER
Plant regeneration via somatic embryogenesis in Drimia robusta
Ponnusamy Baskaran • Johannes Van Staden
Received: 27 March 2014 / Accepted: 9 June 2014 / Published online: 20 June 2014
� Springer Science+Business Media Dordrecht 2014
Abstract A simple efficient in vitro plant regeneration
system was developed by direct and indirect somatic
embryogenesis of Drimia robusta, a medicinal plant
extensively used in South African traditional medicine.
Different developmental stages of somatic embryos (SEs:
globular embryos, partial pear-shaped embryos and club-
shaped embryos), club-shaped cotyledon initiation, plu-
mule initiation and plantlets were directly obtained from
leaf explants on Murashige and Skoog (MS) medium
containing 3.5 % (w/v) sucrose and different plant growth
regulators (PGRs). In MS medium containing 3.5 % (w/v)
sucrose and supplemented with 10 lM picloram, 1 lM
thidiazuron (TDZ) and 20 lM glutamine, a higher number
of SEs and plantlets were achieved. These were established
onto half-strength MS medium followed by successful
acclimatization (100 %) in the greenhouse. Liquid somatic
embryo medium (SEML) containing 500 mg of friable
embryogenic callus on MS medium supplemented with
different concentrations and combinations of PGRs and
organic elicitors produced different stages of SEs. Somatic
embryo production was enhanced by 0.5 lM picloram,
1 lM TDZ and mebendazole treatment. The highest
number of plantlets (9.0 ± 0.70) was obtained in SEML
containing 0.5 lM picloram, 1 lM TDZ and 25 mg l-1
haemoglobin. All the cotyledon and plumule embryos
germinated on half-strength MS medium, however 90 % of
SEs germinated on half-strength MS medium containing
0.5 lM naphthaleneacetic acid. All plantlets were suc-
cessfully acclimatized in the greenhouse. This first report
of D. robusta somatic embryogenesis provides an oppor-
tunity to control extinction threats, ensure germplasm
conservation and provides a system for analysis of bioac-
tive compounds and bioactivity.
Keywords Cell suspension � Drimia robusta � Medicinal
plant � Organic elicitors � Picloram � Somatic
embryogenesis � Thidiazuron
Abbreviations
2,4-D 2,4-Dichlorophenoxyacetic acid
BA 6-Benzyladenine
FEC Friable embryogenic callus
HB Haemoglobin
MS Murashige and Skoog medium
MBZ Mebendazole
NAA a-Naphthaleneacetic acid
OEs Organic elicitors
PGRs Plant growth regulators
PPF Photosynthetic photon flux
SEs Somatic embryos
SCV Settled cell volume
SEML Liquid somatic embryo medium
TDZ Thidiazuron
Introduction
Drimia robusta Baker (Hyacinthaceae) is an important
southern African medicinal plant, especially in KwaZulu-
Natal province (Eloff 1998). It contains the medicinally
important bufadienolide, proscillaridin A (PsA). This plant
is used to treat expectorant, emetic, diuretic, heart tonic and
feverish colds (Hutchings 1989; Pujol 1990; Van Wyk and
Gericke 2000). The leaves and bulbs of D. robusta exhibit
P. Baskaran � J. Van Staden (&)
Research Centre for Plant Growth and Development, School of
Life Sciences, University of KwaZulu-Natal Pietermaritzburg,
Scottsville 3209, South Africa
e-mail: [email protected]
123
Plant Cell Tiss Organ Cult (2014) 119:281–288
DOI 10.1007/s11240-014-0532-2

antibacterial, anti-inflammatory, anti-hypertensive and
anticancer activities (Luyt et al. 1999; Fouche et al. 2008).
D. robusta wild populations are over-exploited. Recently,
this species has been included given Data Deficient–
Taxonomically Problematic (DDT) status in the Red List of
South African plants (2013). In vitro plant regeneration of
D. robusta is potentially a valuable method for conserva-
tion, mass propagation and production of bioactive com-
pounds of this species. Although D. robusta
micropropagation has been described (Ngugi et al. 1998;
Baskaran et al. 2013), there is no report of its in vitro plant
regeneration by somatic embryogenesis.
There is growing interest in developing somatic
embryogenesis systems for medicinal plants (Martin
2004a; Baskaran and Van Staden 2012) as a fast and effi-
cient method for clonal mass propagation (Shoyama et al.
1997), production of artificial seeds (Manjkhola et al.
2005), genetic transformation (Parimalan et al. 2010),
cryopreservation (Hua and Rong 2010), and production of
bioactive compounds (Jeong et al. 2005). The technique is
also useful for germplasm conservation, the selection of
genetic variants with desirable characters, the generation of
somaclonal variants and for performing cellular genetic
manipulations (Vasil 1988). Recently, organic elicitors
have been used in improving somatic embryogenesis in
many plants (Martin 2004b; Jayabalan et al. 2004; Bask-
aran and Van Staden 2012) and were useful in enhance-
ment of secondary metabolites for commercial application
(Savitha et al. 2006). Therefore, the development of a
somatic embryogenetic system for plant regeneration
would be helpful for D. robusta in vitro programmes.
The aim of the present study was to develop simple and
efficient protocols for efficient plant regeneration rates via
direct and indirect somatic embryogenesis from leaf
explants of D. robusta, by investigating the effects of plant
growth regulators, and organic elicitors (OEs: haemoglo-
bin, glutamine and mebendazole) on the processes.
Embryogenic suspension cultures can be ideal vehicles for
further research on somatic mutation, protoplast culture,
somatic hybridization and genetic transformation.
Materials and methods
Plant regeneration by direct somatic embryogenesis
Plant material and explant preparation were done as
described previously (Baskaran et al. 2013). Leaf explants
(approximately 10 9 4.0 mm) of D. robusta were excised
from 20-day-old in vitro germinated seedlings and used as
initial explants for both direct and indirect somatic
embryogenesis. Leaf explants were cultured on MS (Mu-
rashige and Skoog 1962) solid [0.8 % (w/v) agar, Sigma,
USA] medium with 3.5 % (w/v) sucrose and different
concentrations and combinations of plant growth regulators
[PGRs: 2,4-dichlorophenoxyacetic acid (2,4-D), picloram,
benzyladenine (BA) and thidiazuron (TDZ)] and 20 lM
glutamine for production of direct somatic embryos (SEs)
and plantlet development. The PGR treatments are indi-
cated in Table 1. The SEs and degree of plantlet induction
were recorded after 10 weeks of culture. All the plantlets
(approximately 10–15 mm) were transferred into half-
strength MS medium for 4 weeks for plant growth and
development. Thereafter, plantlets (approximately
40–60 mm) were acclimatized successfully as described
previously (Baskaran et al. 2013).
In all experiments, the medium lacking plant growth
regulators served as control. The chemicals used were of
analytical grade (Biolab, South Africa; Oxoid, England and
Sigma, USA). All media were adjusted to pH 5.8 with
0.1 N NaOH and/or 0.1 N HCl before gelling with 0.8 %
(w/v) agar and autoclaved at 121 �C for 20 min. The cul-
tures were maintained at a temperature of 25 ± 2 �C and
light intensity of 40 lmol m-2 s-1 provided by cool white
fluorescent light (ORAM L 58 W/740, South Africa) under
a 16 h photoperiod.
Plant regeneration by suspension culture
For production of indirect somatic embryogenesis, the leaf
explants were cultured on MS solid [0.8 % (w/v) agar]
medium and 3.5 % (w/v) sucrose, supplemented with 2,4-
D or picloram in combinations with TDZ and glutamine, as
specified in Table 2. After 4 weeks, friable embryogenic
callus (FEC) from each medium were transferred to liquid
somatic embryo medium (SEML) containing 20 ml liquid
MS medium, 3.5 % (w/v) sucrose and supplemented with
different concentrations and combinations of PGRs (2,4-D,
picloram and TDZ) and organic elicitors [OEs: glutamine
(GM), haemoglobin (HB) and mebendazole (MBZ: methyl
5-benzoyl-1H-benzimidazol-2-yl carbamate] in 100 ml
erlenmeyer flasks. The precise concentrations and combi-
nations are outlined in Table 2. Suspension culture of D.
robusta was performed as previously described (Baskaran
and Van Staden 2012).
Different developmental stages of SEs (globular embryo,
partial pear-shaped, club-shaped, torpedo-shaped, and coty-
ledon and plumule stages) and plantlets per flask of culture
was determined and were recorded after 6 weeks of culture.
All embryo stages were photographed under a Leica M Stereo
Microscope (JVC-Digital Camera: KY-F 1030U type; 0.5X,
Wayne, NJ, USA). All stages of embryos were cultured on
plant induction media [half-strength solid MS medium; half-
strength MS medium plus 0.5 lM a-naphthaleneacetic acid
(NAA)]. The cultures were maintained in darkness for 3 days
and then incubated under 40 lmol m-2 s-1 light intensity
282 Plant Cell Tiss Organ Cult (2014) 119:281–288
123
and a 16 h photoperiod. Embryo germination percentage was
calculated after 8 weeks (number of germinated SE/total
number of SE 9100). The plantlets (approximately
40–60 mm) were successfully acclimatized in a greenhouse
(Baskaran et al. 2013).
Statistical analysis
All experiments were repeated at least three times with 25
replicates for direct somatic embryogenesis and FEC for-
mation, and 5 replicates for SE formation by suspension
culture and germination per treatment. Data were statisti-
cally analyzed using analysis of variance (ANOVA), and
are presented as mean ± standard error of three indepen-
dent experiments. Treatment means were separated using
Duncan’s multiple range test at the 5 % probability level
and analyzed using SPSS Windows version 11 (SPSS Inc.,
Chicago, IL, USA).
Results and discussion
Direct somatic embryo production and conversion
into plantlets
Direct somatic embryogenesis was achieved from leaf
explants on MS medium, 3.5 % (w/v) sucrose and various
plant growth regulators (PGRs) alone or in combination.
Friable embryogenic calli (FEC) formed at wounding sites
of leaf explants after a week in culture and later white
globular embryoids developed directly from leaf explants
in all treatments except the control after 4 weeks (Fig. 1a).
Lincy et al. (2009) have also reported a similar type of
embryo development in Zingiber officinale. SEs develop
either from callus or directly from the explants without an
intermediate callus stage (Jayanthi et al. 2011). In this
study, globular embryos were improved significantly in all
treatments after 8 weeks of culture (Table 1; Fig. 1b).
However, SEs [globular and other different stages of
embryos (partial pear-shaped embryo, club-shaped embryo,
club-shaped cotyledon initiation and the plumule initia-
tion)] and plantlet number were varied after 10 weeks in
treatments (Table 1; Fig. 1c, d). Similar patterns of embryo
development stages are reported for other plant species
(Lincy et al. 2009). Different patterns of developmental
stages of monocotyledon SEs were reported previously
(Shah 1982; Lincy et al. 2009; Raju et al. 2013). Combi-
nation of picloram or 2,4-D, TDZ and glutamine was more
effective in production of SEs and plantlet development
(Table 1). Picloram, TDZ and glutamine induced higher
rates of somatic embryogenesis in other bulbous plants
species (Tribulato et al. 1997; Bakhshaie et al. 2010;
Baskaran and Van Staden 2012). In contrast, cytokinins
alone are considered as the best inducers of SEs in other
Hyacinthaceae species such as Muscari armeniacum
(Wang et al. 2013). In this study, addition of glutamine in
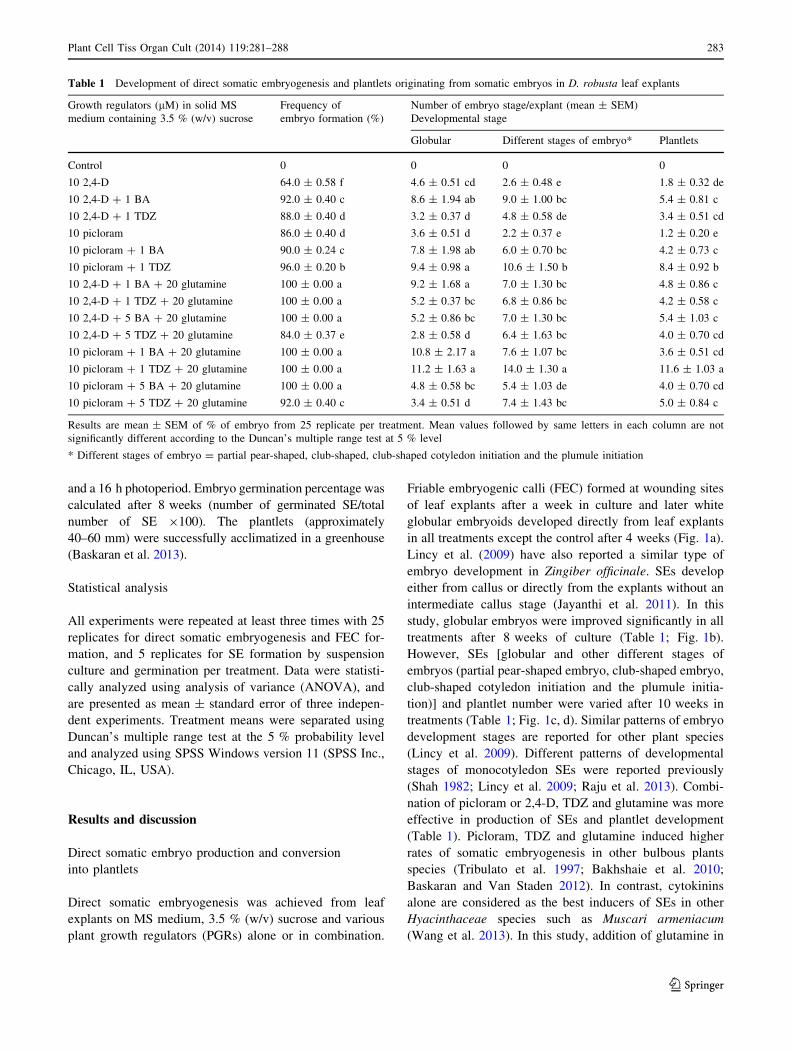
Table 1 Development of direct somatic embryogenesis and plantlets originating from somatic embryos in D. robusta leaf explants
Growth regulators (lM) in solid MS
medium containing 3.5 % (w/v) sucrose
Frequency of
embryo formation (%)
Number of embryo stage/explant (mean ± SEM)
Developmental stage
Globular Different stages of embryo* Plantlets
Control 0 0 0 0
10 2,4-D 64.0 ± 0.58 f 4.6 ± 0.51 cd 2.6 ± 0.48 e 1.8 ± 0.32 de
10 2,4-D ? 1 BA 92.0 ± 0.40 c 8.6 ± 1.94 ab 9.0 ± 1.00 bc 5.4 ± 0.81 c
10 2,4-D ? 1 TDZ 88.0 ± 0.40 d 3.2 ± 0.37 d 4.8 ± 0.58 de 3.4 ± 0.51 cd
10 picloram 86.0 ± 0.40 d 3.6 ± 0.51 d 2.2 ± 0.37 e 1.2 ± 0.20 e
10 picloram ? 1 BA 90.0 ± 0.24 c 7.8 ± 1.98 ab 6.0 ± 0.70 bc 4.2 ± 0.73 c
10 picloram ? 1 TDZ 96.0 ± 0.20 b 9.4 ± 0.98 a 10.6 ± 1.50 b 8.4 ± 0.92 b
10 2,4-D ? 1 BA ? 20 glutamine 100 ± 0.00 a 9.2 ± 1.68 a 7.0 ± 1.30 bc 4.8 ± 0.86 c
10 2,4-D ? 1 TDZ ? 20 glutamine 100 ± 0.00 a 5.2 ± 0.37 bc 6.8 ± 0.86 bc 4.2 ± 0.58 c
10 2,4-D ? 5 BA ? 20 glutamine 100 ± 0.00 a 5.2 ± 0.86 bc 7.0 ± 1.30 bc 5.4 ± 1.03 c
10 2,4-D ? 5 TDZ ? 20 glutamine 84.0 ± 0.37 e 2.8 ± 0.58 d 6.4 ± 1.63 bc 4.0 ± 0.70 cd
10 picloram ? 1 BA ? 20 glutamine 100 ± 0.00 a 10.8 ± 2.17 a 7.6 ± 1.07 bc 3.6 ± 0.51 cd
10 picloram ? 1 TDZ ? 20 glutamine 100 ± 0.00 a 11.2 ± 1.63 a 14.0 ± 1.30 a 11.6 ± 1.03 a
10 picloram ? 5 BA ? 20 glutamine 100 ± 0.00 a 4.8 ± 0.58 bc 5.4 ± 1.03 de 4.0 ± 0.70 cd
10 picloram ? 5 TDZ ? 20 glutamine 92.0 ± 0.40 c 3.4 ± 0.51 d 7.4 ± 1.43 bc 5.0 ± 0.84 c
Results are mean ± SEM of % of embryo from 25 replicate per treatment. Mean values followed by same letters in each column are not
significantly different according to the Duncan’s multiple range test at 5 % level
* Different stages of embryo = partial pear-shaped, club-shaped, club-shaped cotyledon initiation and the plumule initiation
Plant Cell Tiss Organ Cult (2014) 119:281–288 283
123

media influenced effective somatic embryogenesis in D.
robusta. However, media containing 10 lM picloram,
1 lM TDZ and 20 lM glutamine produced significantly
higher frequency (100 %), SEs (11.2–14.0) and number of
plantlets (11.6) compared to other treatments (Table 1;
Fig. 1e). The present study revealed that a combination of
auxin, cytokinin and glutamine is important for significant
SEs production in D. robusta. In this study, all matured
embryos produced plantlets which grew well on half-
strength MS medium. Thereafter, all the plantlets were
successfully established in the greenhouse with a survival
rate of 100 %.
Induction of embryogenic callus
Different PGRs alone or in combinations were used to test
their effect on induction of embryogenic callus (data not
shown). It was observed that combinations of 10 lM
picloram or 2,4-D, 1 lM TDZ and 20 lM glutamine
played an important role in production of D. robusta
embryogenic calli (100 %). These two different combina-
tions promoted fast growing FEC after 2 weeks. In this
study, addition of TDZ and glutamine with picloram was
even more effective in FEC induction (Fig. 2a). Similar
phenomena were also found in other Hyacinthaceae
Table 2 Effect of growth regulators and organic elicitors on somatic embryo (SE) development from embryogenic suspension cells of D.
robusta
FEC induction
medium PGR (lM)
PGR (lM) and organic
supplement in SEML
Frequency of
embryo
formation (%)
Number of embryo stage/explant (mean ± SEM)
Developmental stage
Globular Different
stages
of embryo*
Plantlets
10 picloram ? 1 TDZ ? 20
glutamine
Control (no PGR and supplement) 48.0 ± 0.24 k 2.2 ± 0.37 h 1.2 ± 0.20 f 0.0 ± 0.00
0.5 picloram 72.0 ± 0.40 h 4.2 ± 0.58 fg 3.0 ± 0.54 f 1.0 ± 0.32 ij
0.5 picloram ? 1 TDZ 84.0 ± 0.37 e 5.4 ± 0.86 ef 8.0 ± 1.05 e 2.6 ± 0.24 hi
0.5 picloram ? 2 TDZ 92.0 ± 0.67 c 2.8 ± 0.37 gh 10.6 ± 1.03 cd 8.0 ± 0.92 ab
0.5 picloram ? 1 TDZ ? 20
glutamine
100 ± 0.00 a 3.8 ± 0.32 fg 8.2 ± 1.35 e 4.8 ± 0.37 ef
0.5 picloram ? 1 TDZ ? 40
glutamine
100 ± 0.00 a 4.6 ± 0.51 fg 12.6 ± 1.28 cd 7.2 ± 0.58 ab
0.5 picloram ? 1 TDZ ? 25
haemoglobin
100 ± 0.00 a 10.6 ± 0.92 bc 12.0 ± 1.68 cd 9.0 ± 0.70 a
0.5 picloram ? 1 TDZ ? 50
haemoglobin
76.0 ± 0.37 g 5.6 ± 0.51 ef 9.6 ± 1.36 de 3.8 ± 0.34 fg
0.5 picloram ? 1 TDZ ? 15
mebendazole
88.0 ± 0.40 d 14.0 ± 1.30 a 13.4 ± 1.66 cd 6.0 ± 0.72 bc
0.5 picloram ? 1 TDZ ? 30
mebendazole
80.0 ± 0.44 f 9.0 ± 0.70 cd 19.6 ± 2.01 a 4.6 ± 0.60 ef
10 2,4-D ? 1 TDZ ? 20
glutamine
Control (no PGR and supplement) 56.0 ± 0.58 j 3.0 ± 0.76 gh 1.2 ± 0.37 f 0.0 ± 0.00
0.5 2,4-D 74.0 ± 0.52 g 5.0 ± 0.70 fg 3.0 ± 0.54 f 0.8 ± 0.37 ij
0.5 2,4-D ? 1 TDZ 82.0 ± 0.24 e 6.2 ± 0.58 de 8.4 ± 1.28 e 3.0 ± 0.70 gh
0.5 2,4-D ? 2 TDZ 76.0 ± 0.37 g 4.8 ± 0.37 fg 8.6 ± 0.92 e 7.0 ± 0.64 ab
0.5 2,4-D ? 1 TDZ ? 20 glutamine 96.0 ± 0.20 b 5.8 ± 0.86 ef 9.8 ± 1.24 de 3.4 ± 0.58 gh
0.5 2,4-D ? 1 TDZ ? 40 glutamine 88.0 ± 0.40 d 4.0 ± 0.70 fg 11.8 ± 1.65 cd 4.2 ± 0.51 ef
0.5 2,4-D ? 1 TDZ ? 25
haemoglobin
72.0 ± 0.24 h 8.0 ± 0.92 cd 10.2 ± 1.06 de 7.8 ± 0.58 ab
0.5 2,4-D ? 1 TDZ ? 50
haemoglobin
68.0 ± 0.51 i 8.4 ± 1.20 cd 11.6 ± 1.36 cd 5.0 ± 0.74 de
0.5 2,4-D ? 1 TDZ ? 15
mebendazole
84.0 ± 0.34 e 12.8 ± 1.98 ab 14.8 ± 1.98 bc 5.2 ± 0.92 cd
0.5 2,4-D ? 1 TDZ ? 30
mebendazole
92.0 ± 0.24 c 8.2 ± 1.46 cd 18.6 ± 2.71 ab 5.6 ± 1.36 cd
Values with the mean ± SEM derived from 5 replicate (each 500 mg FEC) with 500 ll settled cell volume of embryogenic suspension cells per
replicate. Mean values followed by same letters in each column are not significantly different according to the Duncan’s multiple range test at
5 % level
* Different stages of embryo = torpedo-shaped, cotyledon-stage and plumule-stage embryos. Haemoglobin and mebendazole = mg l-1
284 Plant Cell Tiss Organ Cult (2014) 119:281–288
123

species (Baskaran and Van Staden 2012). The formation of
embryogenic tissues from somatic cells can be stimulated
by altering either the levels of exogenously applied auxins
or the ratio of auxin to cytokinin (Baskaran and Van Staden
2012). Many reports confirmed that somatic embryogenesis
is influenced by glutamine (Baskaran and Jayabalan 2009;
Deo et al. 2010; Baskaran and Van Staden 2012). The FEC
did not proliferate continuously but differentiated into
somatic embryogenesis after 6 weeks of culture (Fig. 2b).
In the present study, two different combinations of PGRs
that produced calli after 4 weeks were used in SEs for-
mation via cell suspension culture (Table 2).
Somatic embryos formation in cell suspension culture
Plant regeneration via somatic embryonic cell suspension
culture of D. robusta was successfully achieved with SEML
containing various treatments of PGRs alone or in
combination (Table 2). Fast growing FEC from two dif-
ferent combinations were transferred to SEML containing
PGRs (Table 2). FEC units enlarged and produced small
cell aggregates (approximately 0.5–1.0 mm diameter) after
2 weeks in SEML. Thereafter, it produced dense cell
aggregates which were white and soft, or white but watery
callus. However, proliferation rate of cell lines and pro-
portion of cell aggregates varied in settled cell volume
(SCV) of FEC in each treatment. Whitish-soft globular-
shaped, partial pear-shaped, club-shaped and torpedo-
shaped embryos developed from dense cell aggregates after
3 weeks of culture initiation (Fig. 2c). Whitish-green-rigid
globular, torpedo-shaped, cotyledon-stage and plumule-
stage embryos and plantlets were produced and varied in
number from each treatment after 6 weeks (Table 2;
Fig. 2d). Production of globular embryos increased sig-
nificantly on media containing 0.5 lM picloram or 2,4-D
and 1 lM TDZ, while induction of different stages of
Fig. 1 In vitro plant regeneration by direct somatic embryogenesis
form leaf explants of D. robusta: a induction of white globular
embryoids on MS medium plus 10 lM 2,4-D, 5 lM BA and 20 lM
glutamine. b Proliferation of globular embryos in MS medium
containing 10 lM picloram, 1 lM TDZ and 20 lM glutamine.
c Microscopic photograph of developmental stages of somatic
embryos (910). d Proliferation of different stages of embryos and
plantlets on MS medium plus 10 lM picloram, 1 lM TDZ and
20 lM glutamine. e Formation of plantlets on MS medium plus
10 lM picloram, 1 lM TDZ and 20 lM glutamine. Bar (a, b, d and
e) 10 mm
Plant Cell Tiss Organ Cult (2014) 119:281–288 285
123

embryos and plantlets was enhanced in 2 lM TDZ
(Table 2). These embryos were thick and covered with
friable calli. The conversion of the embryos to plantlets
was low (\50 %). Therefore, lower concentration of TDZ
is essential for healthy SEs production.
SEML containing PGRs and organic elicitors (OEs: hae-
moglobin, glutamine and mebendazole) combinations sig-
nificantly increased proliferation of SEs in D. robusta.
Organic additives contains minerals, essential nutrients and
vitamins and have been reported to be effective in somatic
embryogenesis in several plant species (Jayabalan et al. 2004;
Baskaran and Jayabalan 2009; Deo et al. 2010; Al-Khayri
2013; Baskaran and Van Staden 2012). SEs were relatively
large and green in colour in all treatments (Fig. 2e). Meben-
dazole (MBZ) significantly improved somatic embryo
production (globular to cotyledon-stage) after 6 weeks in
culture (Table 2; Fig. 2e). MBZ is a synthetic anthelmintic
drug which acted as a growth regulator on shoot and root
development in M. plumbea and D. robusta micropropagation
(Baskaran et al. 2012, 2013). However, in this study, hae-
moglobin (25 mg l-1) produced a higher number of plantlets
(9.0 ± 0.70 per SCV) from matured embryos (Table 2;
Fig. 2f). A similar effect was also observed with other Hya-
cinthaceae species such as M. plumbea (Baskaran and Van
Staden 2012). Haemoglobin plays a role in enhancement of
mitotic division (Azhakanandam et al. 1997) and cellular O2
supply in vitro (Anthony et al. 1997). Recently, two plant
hemoglobin (Hb) genes (ZmHb1 and ZmHb2) that regulate
the cell survival/death decision that influences somatic
embryogenesis through their cell-specific localization
Fig. 2 In vitro plant regeneration from somatic embryos on cell
suspension culture. a Induction of FEC on MS medium containing
10 lM picloram, 1 lM TDZ and 20 lM glutamine. b Formation of
cotyledon and plumule stages of embryos. c Different developmental
stages from dense cell aggregates. d Production of different stages of
embryos and plantlets on SEML containing 0.5 lM picloram and
2 lM TDZ. e Formation of somatic embryos (globular to cotyledon-
stage) on SEML containing 0.5 lM picloram, 1 lM TDZ and
30 mg l-1 mebendazole. f Formation of plantlets on SEML containing
0.5 lM picloram, 1 lM TDZ and 25 mg l-1 haemoglobin. g Germi-
nation of embryos and plantlet development on half-strength MS
medium supplemented with 0.5 lM NAA. h Well-developed plantlets
from germination medium. i Acclimatized plants of D. robusta in the
greenhouse after 6 months. Bar (a–g) 10 mm
286 Plant Cell Tiss Organ Cult (2014) 119:281–288
123

patterns have been reported (Huang et al. 2014). In this study,
increasing the concentration of haemoglobin significantly
decreased the frequency, number of SEs and plantlets
(Table 2). In contrast, a high concentration of haemoglobin
was effective in formation of SEs in M. plumbea, cotton and
peanut (Jayabalan et al. 2004; Ganesan and Jayabalan 2004;
Baskaran and Van Staden 2012). The medium containing
40 lM glutamine showed a significant improvement in pro-
duction of SEs (Table 2). Improvement of SEs by addition of
glutamine in the medium has also been reported for other
plant species (Baskaran and Jayabalan 2009; Deo et al. 2010;
Al-Khayri 2013; Baskaran and Van Staden 2012). The pres-
ent study indicated that SEs production in D. robusta is
dependent on the type and concentration of OEs. Similar
phenomena have also been reported in other plant species
(Deo et al. 2010; Al-Khayri 2013; Baskaran and Van Staden
2012).
Conversion of plantlets
Successful plant regeneration was achieved from post
somatic embryogenetic developmental stages; globular,
torpedo, cotyledon and plumule on the plant induction
medium. All mature embryos germinated to produce
complete plantlets after 2 weeks in culture. Mostly, glob-
ular embryos failed to germinate on half-strength solid MS
medium, but only rooting occurred with half-strength MS
medium plus 0.5 lM NAA. Half-strength solid MS med-
ium produced 65 % germination, whereas half-strength MS
medium supplemented with 0.5 lM NAA produced a
higher frequency of germination (90 %) and well rooted
plantlets (Fig. 2g). The beneficial effect of auxins and
cytokinins on somatic embryo germination has been
reported in many other monocotyledon plant species
(Kackar et al. 1993; Guo and Zhang 2005; Lincy et al.
2009). Successful germination and plant regeneration via
SEs has been reported with combinations of BA and NAA
in ginger and buffel grass (Lincy et al. 2009; Carloni et al.
2014). After germination, NAA has been required for plant
development in buffel grass (Carloni et al. 2014). Well-
developed plantlets were separated (Fig. 2h) and then
transferred to plastic pots containing a 1:1 (v/v) vermicu-
lite:sand mixture. The plantlets were successfully accli-
matized in a greenhouse (Fig. 2i) with 100 % survival rate.
The present work is a first report for plant regeneration
via direct and indirect somatic embryogenesis from leaf
explants of D. robusta. Promising plant regeneration from
SEs via direct and indirect somatic embryogenesis was
influenced markedly by PGRs combined with OEs (gluta-
mine and haemoglobin). The system will be helpful for
conservation, mass clonal propagation, production of bio-
active compounds and genetic transformation studies.
Acknowledgments The financial support by National Research
Foundation (NRF), Pretoria and the University of KwaZulu-Natal,
Pietermaritzburg is gratefully acknowledged. The authors are grateful
to the Microscopy & Microanalysis Unit (MMU), UKZN, Pieterma-
ritzburg for microscopic assistance.
References
Al-Khayri JM (2013) Complex organic additives-induced changes
during somatic embryogenesis in plant system. In: Aslam J,
Srivastava PS, Sharma MP (eds) Somatic embryogenesis and
gene expression. Narosa Publishing House, New Delhi,
pp 82–123
Anthony P, Davey MR, Power JB, Lowe KC (1997) Enhanced mitotic
division of cultured Passiflora and Petunia protoplasts by
oxygenated perfluorocarbon and haemoglobin. Biotechnol Tech-
nol 11:581–584
Azhakanandam K, Lowe KC, Power JB, Davey MR (1997) Haemo-
globin (Erythrogen)-enhanced mitotic division and plant regen-
eration from cultured rice protoplasts (Oryza sativa L.). Enzyme
Microb Technol 21:572–577
Bakhshaie M, Babalar M, Mirmasoumi M, Khalighi A (2010)
Somatic embryogenesis and plant regeneration of Lilium ledeb-
ourii (Baker) Boiss., an endangered species. Plant Cell Tissue
Organ Cult 102:229–235
Baskaran P, Jayabalan N (2009) In vitro propagation of Psoralea
corylifolia L. by somatic embryogenesis in cell suspension
culture. Acta Physiol Plant 31:1119–1127
Baskaran P, Van Staden J (2012) Somatic embryogenesis of Merwilla
plumbea (Lindl.) Speta. Plant Cell Tissue Organ Cult 109:
517–524
Baskaran P, Ncube B, Van Staden J (2012) In vitro propagation and
secondary product production by Merwilla plumbea (Lindl.)
Speta. Plant Growth Regul 67:235–245
Baskaran P, Singh S, Van Staden J (2013) In vitro propagation,
proscillaridin A production and antibacterial activity in Drimia
robusta. Plant Cell Tissue Organ Cult 114:259–267
Carloni E, Ribotta A, Colomba EL, Griffa S, Quiroga M, Tommasino
E, Grunberg K (2014) Somatic embryogenesis from in vitro
anther culture of apomictic buffel grass genotypes and analysis
of regenerated plants using flow cytometry. Plant Cell Tissue
Organ Cult. doi:10.1007/s11240-014-0441-4
Deo PC, Taylor M, Harding RM, Tyagi AP, Becker DK (2010)
Initiation of embryogenic cell suspensions of taro (Colocasia
esculenta var. esculenta) and plant regeneration. Plant Cell
Tissue Organ Cult 100:283–291
Eloff JN (1998) A sensitive and quick microplate method to
determine the minimal inhibitory concentration of plant extracts
for bacteria. Plant Med 64:711–713
Fouche G, Cragg GM, Pillay P, Kolesnikova N, Maharaj VJ, Senabe J
(2008) In vitro anticancer screening of South African plants.
J Ethnopharmacol 119:455–461
Ganesan M, Jayabalan N (2004) Evaluation of haemoglobin (erythr-
ogen): for improved somatic embryogenesis and plant regener-
ation in cotton (Gossypium hirsutum L. cv. SVPR 2). Plant Cell
Rep 23:181–187
Guo Y, Zhang ZX (2005) Establishment and plant regenerations of
somatic embryogenic cell suspension cultures of the Zingiber
officinale Rosc. Sci Hortic 107:90–96
Hua YM, Rong HN (2010) A simple cryopreservation protocol of
Dioscorea bulbifera L. embryogenic calli by encapsulation-
vitrification. Plant Cell Tissue Organ Cult 101:349–358
Plant Cell Tiss Organ Cult (2014) 119:281–288 287
123

Huang S, Hill RD, Wally OS, Dionisio G, Ayele BT, Jami SK,
Stasolla C (2014) Hemoglobin control of cell survival/death
decision regulates in vitro plant embryogenesis. Plant Physiol
165:810–825
Hutchings A (1989) A survey and analysis of traditional medicinal
plants as used by the Zulu, Xhosa and Sotho. Bothalia
19:111–123
Jayabalan N, Anthony P, Davey MR, Power JB, Lowe KC (2004)
Hemoglobin promotes somatic embryogenesis in peanut cul-
tures. Artif Cells Blood Subst Immobil Biotechnol 32:149–157
Jayanthi M, Mohan N, Mandal PK (2011) Direct somatic embryo-
genesis and plantlet regeneration in oil palm. J Plant Biochem
Biotechnol 20:249–251
Jeong JH, Jung SJ, Murthy HN, Yu KW, Paek KY, Moon HK, Choi
YE (2005) Production of eleutherosides in in vitro regenerated
embryos and plantlets of Eleutherococcus chiisanensis. Biotech-
nol Lett 27:701–704
Kackar A, Bhat SR, Chandel KPS, Malik SK (1993) Plant regener-
ation via somatic embryogenesis in ginger. Plant Cell Tissue
Organ Cult 32:289–292
Lincy AK, Remashree AB, Sasikumar B (2009) Indirect and direct
somatic embryogenesis from aerial stem explants of ginger
(Zingiber officinale Rosc.). Acta Bot Croat 68(1):93–103
Luyt RP, Jager AK, Van Staden J (1999) The rational usage of Drimia
robusta Bak. in traditional medicine. S Afr J Bot 65:1–4
Manjkhola S, Dhar U, Joshi M (2005) Organogenesis, embryogenesis,
and synthetic seed production in Arnebia euchroma—a critically
endangered medicinal plant of the Himalaya. In Vitro Cell Dev
Biol Plant 41:244–248
Martin KP (2004a) Plant regeneration through somatic embryogenesis
in medicinally important Centella asiatica L. In Vitro Cell Dev
Biol Plant 40:586–591
Martin KP (2004b) Plant regeneration protocol of medicinally
important Andrographis paniculata (Burm. F.) Wallich ex Nees
via somatic embryogenesis. In Vitro Cell Dev Biol Plant
40:204–209
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bio assays with tobacco tissue cultures. Physiol Plant 15:473–497
Ngugi G, Jager AK, Van Staden J (1998) In vitro propagation of
Drimia robusta Bak. S Afr J Bot 64:266–268
Parimalan R, Akshatha V, Giridhar P, Ravishankar GA (2010)
Somatic embryogenesis and Agrobacterium-mediated transfor-
mation in (Bixa orellana L.). Plant Cell Tissue Organ Cult
105:317–328
Pujol J (1990) Nature Africa: the Herbalist handbook. Jean Pujol
Natural Healers Foundation, Durban
Raju CS, Kathiravan K, Aslam A, Shajahan A (2013) An efficient
regeneration system via somatic embryogenesis in mango ginger
(Curcuma amada Roxb.). Plant Cell Tissue Organ Cult
112:387–393
SANBI (2013) Statistics: Red List of South African Plants version
2013.1. http://redlist.sanbi.org/stats.php
Savitha BC, Timmaraju R, Bhagyalaksami N, Ravishankar GA
(2006) Different biotic and abiotic elicitors influence betalain
production in hairy root cultures of Beta vulgaris in shake flasks
and bioreactor. Process Biochem 41:50–60
Shah CK (1982) Morpho-histochemical and SEM studies of some
monocotyledonous embryos. Phytomorphol 32:211–221
Shoyama Y, Zhu XX, Nakai R, Shiraishi S, Kohda H (1997)
Micropropagation of Panax notoginseng by somatic embryo-
genesis and RAPD analysis of regenerated plantlets. Plant Cell
Rep 16:450–453
Tribulato A, Remotti PC, Loffler HJM (1997) Somatic embryogenesis
and plant regeneration in Lilium longiflorum Thunb. Plant Cell
Rep 17:113–118
Van Wyk BE, Gericke N (2000) People’s plants, a guide to useful
plants of Southern Africa. Briza Publications, Pretoria
Vasil IK (1988) Progress in the regulation and genetic manipulation
of cereal crops. Biotechnol 6:397–402
Wang S, Yang F, Jiu L, Zhang W, Zhang W, Tian W, Wang F (2013)
Plant regeneration via somatic embryogenesis from leaf explants of
Muscari armeniacum. Biotechnol Biotechnol Equip 27:4243–4247
288 Plant Cell Tiss Organ Cult (2014) 119:281–288
123





























