Embed Size (px)
Citation preview

Phytochemistry Letters xxx (2013) xxx–xxx
G Model
PHYTOL-595; No. of Pages 5
Phytotoxic activity and phytochemical characterization of Lotusornithopodioides L., a spontaneous species of Mediterranean area
Fabrizio Araniti, Francesco Sunseri, Maria Rosa Abenavoli *
Dipartimento di Agraria, Universita Mediterranea di Reggio Calabria, Salita Melissari, I-89124 Reggio Calabria, RC, Italy
A R T I C L E I N F O
Article history:
Received 30 December 2012
Received in revised form 25 August 2013
Accepted 28 August 2013
Available online xxx
Keywords:
Bio-guided fractionation
Fatty acids
Germination
Phenolic acids
Root growth
Semiochemicals
A B S T R A C T
The search of new bioactive compounds produced by plants represented an eco-friendly and alternative
strategy in sustainable agriculture. In this context, the aerial parts of Lotus ornithopodioides L. (Fabaceae),
annual herbaceous plant of Mediterranean area, were collected in Calabria (Southern Italy). Leaves and
stems of L. ornithopodioides were extracted with methanol and subsequently bio-fractionated with n-
hexane, chloroform and ethyl acetate. In vitro assays on germination and root growth of lettuce (Lactuca
sativa L.) were performed to evaluate the phytotoxicity of the methanolic extract and its fractions, by a
dose–response curve, comparing the ED50 values. Then, the most bioactive fractions were characterized
through GC–MS. The crude methanolic extract strongly inhibited both processes showing, after 48 h of
treatment, ED50 values equal to 9.01 and 1.16 mg/ml for germination and root growth, respectively. The
bioguided-fractionation showed the following hierarchy of phytotoxicity for both processes: ethyl
acetate > chloroform � n-hexane. A large amount of phenolic compounds were found in the ethyl
acetate fraction (522 mg/g of dry fraction) and some of them have been characterized through HPLC
analysis. On the other hand, the n-hexane and chloroform fractions showed the presence of many
semiochemicals, such as phytone, methyl hexadecanoate, ethyl hexadecanoate, methyl octadecanoate
and 4-methyl-2-heptanone. These results make L. ornithopodioides L. a suitable source of natural
compounds potentially usable as natural herbicides. Further studies will be necessary to understand the
phytotoxic mechanisms of these molecules, alone or in combination and their effects on green-house
and field experiments.
� 2013 Phytochemical Society of Europe. Published by Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Phytochemistry Letters
jo u rn al h om ep ag e: ww w.els evier .c o m/lo c ate /p hyt ol
1. Introduction
The reduction in the use of synthetic chemicals represents, insustainable agricultura, one of the most important challengesbecause of their negative impact on the environment and humanhealth. The search of new bioactive compounds produced by plantswith new mode of action, more specific targets, less toxic andharmful may represent a valid alternative strategy. Recently,allelopathy defined as ‘‘any process, direct or indirect, involvingsecondary metabolites produced by plants, microorganisms, fungi,able to influence, positively or negatively, the growth anddevelopment of biological and agricultural systems’’ (IAS, 1996),might be considered as a tool for biological weed control in asustainable agriculture. Many studies in this field have beenfocused on the isolation and identification of specific bioactive
* Corresponding author at: Dipartimento di Agraria, Universita degli Studi
‘‘Mediterranea’’ di Reggio Calabria, Salita Melissari, I-89124 Reggio Calabria, Italy.
Tel.: +39 965 324077; fax: +39 965 311092.
E-mail address: [email protected] (M.R. Abenavoli).
Please cite this article in press as: Araniti, F., et al., Phytotoxic activity
spontaneous species of Mediterranean area. Phytochem. Lett. (2013
1874-3900/$ – see front matter � 2013 Phytochemical Society of Europe. Published by
http://dx.doi.org/10.1016/j.phytol.2013.08.019
molecules synthesized by both plants and microorganisms (Dukeet al., 2000). Through bio-fractionation techniques, using increas-ingly polar solvents (Anjum and Bajwa, 2010; Rimando et al.,2001), crude plant extracts were fractionated and the fractionsassayed on the germination and root growth processes ofsusceptible species, such as lettuce (Gross, 2001; Duke et al.,2000, 2002). Then, once identified the most phytotoxic fraction, itwas chemically characterized and the bioactive compoundsquantified (An et al., 2000; Duke et al., 2000). This is considereda prerequisite for assessing the biological activity of naturalcompounds (Morgan and Overholt, 2005; Yu et al., 2003).
The genus Lotus consists of a group of annual and perennialspecies widely distributed throughout the world and largelycropped as pasture and forage in Europe and in USA because oftheir rusticity, resistance to drought and palatability to animals(Seaney and Henderson, 1970). This genus, represented by 200different species, is largely distributed in the Mediterranean area(Pignatti, 1982), probably considered the center of origin for theOld World species (Seaney and Henderson, 1970). In particular,Lotus ornithopodioides L. is one of the most common and widelydistributed in Italy and the Mediterranean basin. This species,
and phytochemical characterization of Lotus ornithopodioides L., a), http://dx.doi.org/10.1016/j.phytol.2013.08.019
Elsevier B.V. All rights reserved.

F. Araniti et al. / Phytochemistry Letters xxx (2013) xxx–xxx2
G Model
PHYTOL-595; No. of Pages 5
self-reseeding annual legume, is characterized by a rapid growthand a large biomass production (Seaney and Henderson, 1970).
In the present study, the methanolic extract of the aerial parts ofL. ornithopodioides was chemically fractionated using solvents withincreasing polarity and the phytotoxicity of each fraction wasassayed on germination and root elongation processes of Lactuca
sativa cv. Parris Island COS USA, a sensible species. The fractions,characterized by the greatest toxicity, were further analyzed foridentification of constituents and their quantification by gaschromatography (GC) and gas chromatography–mass spectrome-try (GC–MS) and high-performance liquid chromatography (HPLC).
2. Materials and methods
2.1. Plant material, crude extract preparation and fractionation
The aerial parts of L. ornithopodioides L. were collected on Marchin Calabria (Southern Italy). A plant sample was authenticated byProf. Silvio Fici (University of Palermo, Italy) and deposited in theherbarium of the Department of Botany (University of Palermo).Leaves and stems were cut, dried at 40 8C, grounded (‘‘Wiley Mill’’,0.2 mm), and the powders were stored at room temperature untiltheir use. Five hundred grams of powders were extracted inmethanol (MeOH) for 48 h and filtered through Whatman No. 2.This procedure, repeated 3 times, was conducted at roomtemperature and in dark condition to avoid any phenomena ofphotodegradation. The crude extract, evaporated to dryness undervacuum below 40 8C using a rotary evaporator, was completelydried under nitrogen gas for several hours and then weighed.Successively, three solvents characterized by increasing polarity,n-hexane, chloroform, and ethyl acetate, were used to fractionizethe crude extract and, then the fractions were dried as previouslydescribed. Crude extract and fraction yields were also calculated.
2.2. Bioassay of methanolic extract
The crude extract was diluted with sterile distilled water at thefollowing concentrations: 0, 0.3, 0.6, 1.2, 2.5, 5, 7.5, 10, 15, and20 mg/ml. Distilled water represented the control (0 mg/ml).
2.2.1. Seed germination and root elongation bioassays
Seeds of L. sativa L. (cv Parris Island Cos USA) were surfacesterilized for 15 min in a 15% (v/v) NaClO solution and then rinsedthree times with sterile deionized water. Then, ten seeds wereuniformly distributed into a Petri dish (6 cm 1) between a doublelayer of filter paper moistened with 2 ml of the above concentra-tions of the crude extract. Petri dishes were then placed in a growthchamber at 25 � 1 8C, 70% relative humidity, in dark conditions. After48 h of treatment, seeds showing a root extrusion at least 1 mm longwere considered germinated. The total germination (GT%) wascalculated as previously reported by Chiapusio et al. (1997).
For the root elongation bioassay, five pre-germinated lettuceseeds, selected for uniformity in root length (1 mm), were placed insterile Petri dishes (6 cm 1) and treated as reported for thegermination bioassay. After 48 h of exposure, an image of lettuceroots for each treatment was captured by scanner (EpsonExpression 800, Regent Instruments, Quebec, Canada) and thetotal root length (TRL) was measured using the WinRhizo ProSystem v. 2002a Software (Instruments Regent, Inc., Quebec,Canada).
2.3. Bioassays of three solvent fractions
Each dry solvent fraction was dissolved with MeOH at thefollowing concentrations: 0, 0.625, 1.25, 2.5, 5, and 10 mg/ml.Methanol represented the control. Two milliliters of each
Please cite this article in press as: Araniti, F., et al., Phytotoxic activity
spontaneous species of Mediterranean area. Phytochem. Lett. (2013
concentration were placed in Petri dishes (6 cm 1) containing adouble layer of filter paper and evaporated to dryness at roomtemperature. After evaporation, 2 ml of distilled water were added.Germination and root elongation bioassays were conducted withthe same procedures described above.
2.4. GC–MS analysis
The n-hexane and chloroform fractions obtained from L.
ornithopodioides L. crude extract were analyzed by GC–MS usinga gas chromatograph (Hewlett-Packard, model 5890), equippedwith a methylsilicone SE-30 capillary column (30 m � 0.25 mm),interfaced with a mass spectrometer (model 5972 series 11) usinghelium as carrier gas. The temperature conditions were scheduledas following: 3 min of isothermal at 60 8C, from 60 till 280 8C with arate of 16 8C/min, 10 min of isothermal at 280 8C. Detector andinjector temperature was set at 280 and 250 8C, respectively (flow1 ml/min). Compounds identification was carried out comparingthe relative retention time and mass spectra of the molecules withthose of the Wiley 138 and 275 library data of the GC–MS system(Hewlett-Packard Co.).
2.5. Determination of total phenolic content
The determination of the total phenolic content of the crudeextract and the ethyl acetate fraction was carried out by Singletonand Rossi method (1965). A mixture of acetone, methanol, aceticacid and water (40:40:0.1:20) was used as extraction solvent. Fiftymilligrams of both crude extract and ethyl acetate fraction werepoured in a falcon tube and shacked with 25 ml extraction solvent.After extraction, each sample was heated at 60 8C for 1 h,homogenized for 30 s and allowed to cool down at roomtemperature. Then, 200 ml of solution of each sample wereintroduced into a test tube together with 1 ml of Folin–Ciocalteu’sreagent and 1 ml of 7.5% sodium carbonate solution. The tubeswere then shacked and allowed to stand for 2 h. The absorptionwas measured at 726 nm (Perkin-Elmer Lambda 35 UV/VISspectrophotometer) and the total phenolic content was expressedas milligram of chlorogenic acid equivalents per gram of dry plantmaterial.
2.6. Determination of phenolic acids by HPLC
The phenolic acids detection was carried out between 210 and360 nm using an APC diode array detector (Back UPS CS 650). Thepeak separation was achieved with a 100-5 C18 column; 5 mm;4.6 mm � 250 mm (Knauer). A step-gradient of acetonitrile inwater was used: 15% acetonitrile (5 min, gradient), 30% acetonitrile(20 min, gradient), 40% acetonitrile (25 min, gradient), 60%acetonitrile (30 min, gradient), 60% acetonitrile (35 min, gradient),and 100% acetonitrile (45 min, isocratic). The flow rate was 1 ml/min; the injection volume was 20 ml. A 0.05% ortho-phosphoricacid solution was added to the solvents to avoid tailing of thephenolic acids. Phenolic compounds identification was donecomparing the retention times of the unknown peaks with theretention times of phenolic acid standards. All analyses wereperformed in triplicate.
2.7. Experimental design and statistical analysis
In order to evaluate the effects of the crude extract and itssolvent fractions on germination and root growth a completelyrandom design with 4 replications was adopted. Germination androot growth responses to different doses of crude extract and itssolvent fractions were evaluated by a nonlinear regression modelusing a log-logistic function that allowed to estimate the ED50
and phytochemical characterization of Lotus ornithopodioides L., a), http://dx.doi.org/10.1016/j.phytol.2013.08.019

Table 1Yield of crude extract and its fractions obtained from L. ornithopodioides powders.
Species Dry weight (g) Solvents Total weight (g) Yield %
L. ornithopodioides 500 Methanolic TOT 155.5 31.1
n-Hexane 19 3.8
Chloroform 7 1.4
Ethyl acetate 10.5 2.1
Table 2Effects of crude extract of L. ornithopodioides on germination and root growth of L.
sativa.
Concentration (mg/ml) Methanol
GT% TRL (cm)
0 96.7 (�5.8)a 1.23 (�0.19)a
0.3 96.7 (�5.8)a 0.81 (�0.04)b
0.6 100 (�0)a 0.78 (�0.07)b
1.2 86.7 (�5.8)ab 0.53 (�0.04)bc
2.5 83.3 (�5.8)ab 0.48 (�0.05)bc
5 80 (0)ab 0.4 (�0.03)c
7.5 66.7 (�5.8)b 0.39 (�0.06)c
10 36.7 (�11.5)c 0.38 (�0.02)c
15 0 (�0)d 0.27 (�0.01)c
20 0 (�0)d 0.24 (�0.02)c
Different letters along the columns [for each physiological process GT% and TRL
(cm)] indicated significant differences at P < 0.05 (Tukey’s test). Values within the
brackets indicated the standard error (N = 4). All the dose–response curves pointed
out a significance level of P < 0.001.
F. Araniti et al. / Phytochemistry Letters xxx (2013) xxx–xxx 3
G Model
PHYTOL-595; No. of Pages 5
parameter, the dose required to reduce 50% of the total response(Streibig et al., 1998; Belz et al., 2005). The comparison of thephytotoxicity of the crude extract and solvent fractions wasperformed by one-way ANOVA using the ED50 as a variable and theextraction solvent as main factor. The ED50 data were first checkedfor deviations from normality (Kolmogorov–Smirnov test) andtested for homogeneity (Leven median test). Tukey’s testcomparison was utilized to compare the mean values of ED50
between crude extract and its solvent fractions over time(P < 0.05).
3. Results and discussion
3.1. Extraction yields
Yields of the crude extract and its fractions are expressed inpercentage and referred to the dry weight of the plant materialused (500 g) (Table 1). In particular, the yield of the crude extractwas 31% whereas the yield of the n-hexane, chloroformic and ethylacetate fractions was 3.8%, 1.4% and 2.1%, respectively (Table 1).
3.2. Bioassays with crude extract and its fractions
The crude extract and the fractions obtained from the aerialparts of L. ornithopodioides strongly affected both germination androot growth of lettuce, suggesting the presence of highlyphytotoxic active molecules. In particular, the crude extract didnot significantly affect lettuce seed germination up to 7.5 mg/ml(concentration able to cause 43% of inhibition). At the highestconcentrations (15 and 20 mg/ml) (Table 2), the completeinhibition of lettuce seed germination was reached. Conversely,crude extract was already able to significantly inhibit the lettuceroot elongation at the lowest concentration (0.3 mg/ml), causing a35% of reduction (Table 2). This inhibitory effect increased alongwith increasing concentrations, reaching at the highest ones (15and 20 mg/ml) �80% of root growth inhibition.
Nearly all fractions, obtained through the bio-guided fraction-ation of the crude extract, revealed the presence of germinationand root growth inhibitors. However, ethyl acetate appeared to bethe most active fraction probably due to its ability to extract morephytotoxic compounds than n-hexane and chloroform solvents, asalready demonstrated on Artemisia arborescens (Araniti et al.,2012a) (Table 3). Indeed, whereas all the fractions completelyinhibited lettuce seed germination at highest doses, the ethylacetate fraction was already active at low concentrations (2.5 mg/ml) causing a significative inhibition (15%) of this physiologicalprocess (Table 3).
Concerning root growth, all fractions strongly reduced it in adose dependent-manner. Indeed, the inhibition was alreadyevident at the lowest concentrations, reaching 80–90% of inhibi-tion at the highest one (10 mg/ml) (Table 3). Seed germination androot elongation data of both crude extract and its fractions werefitted through a non-linear regression model using a log-logisticresponse equation largely applied in weed management research(Nielsen et al., 2004; Petersen et al., 2001), which allowed to obtainthe ED50, a specific parameter which defines the effective dosecausing 50% of total response in the population. All the data
Please cite this article in press as: Araniti, F., et al., Phytotoxic activity
spontaneous species of Mediterranean area. Phytochem. Lett. (2013
obtained for both seed germination and root growth werecharacterized by a high statistical significance (P < 0.001) (Tables2 and 3). The ED50 values for both physiological processesconfirmed the inhibitory activity of the crude extract and itsfractions underlying, as already reported by some authors (Chonet al., 2002; Araniti et al., 2012b), the greater sensitivity of rootgrowth than seed germination to chemicals. In particular, the ED50
values of seed germination further confirmed the highestphytotoxicity of ethyl acetate (2.84 mg/ml) followed by thechloroformic (6.32 mg/ml) and the n-hexane (6.9 mg/ml) fractions(Table 4).
Concerning the root growth, the n-hexane fraction, althoughstrongly phytotoxic (1.32 mg/ml), did not cause any significanteffect compared to crude extract, whereas the ethyl acetatefraction (0.56 mg/ml) confirmed its higher inhibitory potentialcompared to the crude extract (1.16 mg/ml) (Table 4). Relativelyless active on the root growth process appeared to be thechloroform fraction showing ED50 value of 2.64 mg/ml (Table 4).The inhibitory activity observed in all fractions on both processescould be due to the presence of non-polar and/or relatively polarmolecules. Several authors (An et al., 2000; Weston et al., 1989)have shown that these solvents might fractionate classes ofcompounds such as terpenoids, fatty acids, alkaloids and poly-phenols, molecules extremely active and generally involved in theallelopathic phenomenon (Reigosa et al., 1999; Macias et al., 2010;Chaimovitsh et al., 2010).
3.3. GC–MS analysis
In the hexane fraction, 10 molecules have been identified: sevenbelonging to fatty acids class a large group of secondarymetabolites well known for their allelopathic activity (Satoshiet al., 2010), two diterpenes (neophytadiene and phytol) and onepeptone (phytone) (Table 5). Among these molecules, phytone,methyl hexadecanoate, ethyl hexadecanoate and methyl octade-canoate, were widely known for their semiochemical activities(Kalinova et al., 2009; Navarro-Silva et al., 2009). The chloroformfraction characterization allowed to identify six molecules(Table 6) including four alcohols (diacetone alcohol, cyclopentanol,
and phytochemical characterization of Lotus ornithopodioides L., a), http://dx.doi.org/10.1016/j.phytol.2013.08.019
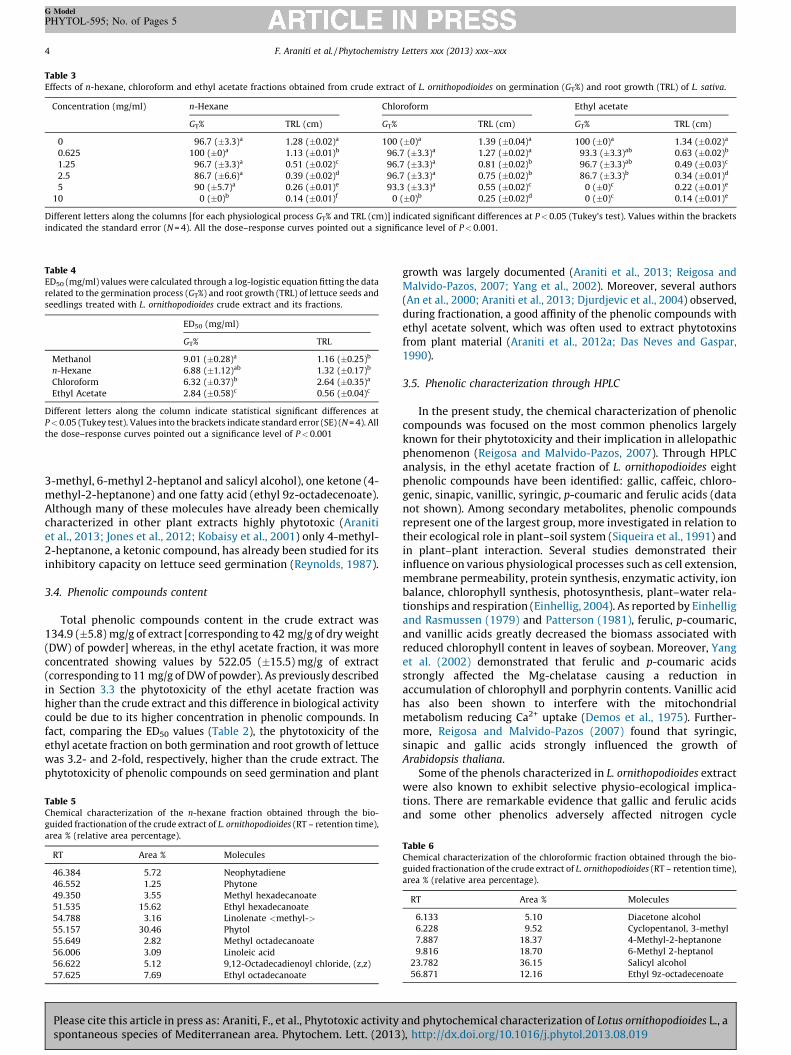
Table 3Effects of n-hexane, chloroform and ethyl acetate fractions obtained from crude extract of L. ornithopodioides on germination (GT%) and root growth (TRL) of L. sativa.
Concentration (mg/ml) n-Hexane Chloroform Ethyl acetate
GT% TRL (cm) GT% TRL (cm) GT% TRL (cm)
0 96.7 (�3.3)a 1.28 (�0.02)a 100 (�0)a 1.39 (�0.04)a 100 (�0)a 1.34 (�0.02)a
0.625 100 (�0)a 1.13 (�0.01)b 96.7 (�3.3)a 1.27 (�0.02)a 93.3 (�3.3)ab 0.63 (�0.02)b
1.25 96.7 (�3.3)a 0.51 (�0.02)c 96.7 (�3.3)a 0.81 (�0.02)b 96.7 (�3.3)ab 0.49 (�0.03)c
2.5 86.7 (�6.6)a 0.39 (�0.02)d 96.7 (�3.3)a 0.75 (�0.02)b 86.7 (�3.3)b 0.34 (�0.01)d
5 90 (�5.7)a 0.26 (�0.01)e 93.3 (�3.3)a 0.55 (�0.02)c 0 (�0)c 0.22 (�0.01)e
10 0 (�0)b 0.14 (�0.01)f 0 (�0)b 0.25 (�0.02)d 0 (�0)c 0.14 (�0.01)e
Different letters along the columns [for each physiological process GT% and TRL (cm)] indicated significant differences at P < 0.05 (Tukey’s test). Values within the brackets
indicated the standard error (N = 4). All the dose–response curves pointed out a significance level of P < 0.001.
Table 4ED50 (mg/ml) values were calculated through a log-logistic equation fitting the data
related to the germination process (GT%) and root growth (TRL) of lettuce seeds and
seedlings treated with L. ornithopodioides crude extract and its fractions.
ED50 (mg/ml)
GT% TRL
Methanol 9.01 (�0.28)a 1.16 (�0.25)b
n-Hexane 6.88 (�1.12)ab 1.32 (�0.17)b
Chloroform 6.32 (�0.37)b 2.64 (�0.35)a
Ethyl Acetate 2.84 (�0.58)c 0.56 (�0.04)c
Different letters along the column indicate statistical significant differences at
P < 0.05 (Tukey test). Values into the brackets indicate standard error (SE) (N = 4). All
the dose–response curves pointed out a significance level of P < 0.001
F. Araniti et al. / Phytochemistry Letters xxx (2013) xxx–xxx4
G Model
PHYTOL-595; No. of Pages 5
3-methyl, 6-methyl 2-heptanol and salicyl alcohol), one ketone (4-methyl-2-heptanone) and one fatty acid (ethyl 9z-octadecenoate).Although many of these molecules have already been chemicallycharacterized in other plant extracts highly phytotoxic (Aranitiet al., 2013; Jones et al., 2012; Kobaisy et al., 2001) only 4-methyl-2-heptanone, a ketonic compound, has already been studied for itsinhibitory capacity on lettuce seed germination (Reynolds, 1987).
3.4. Phenolic compounds content
Total phenolic compounds content in the crude extract was134.9 (�5.8) mg/g of extract [corresponding to 42 mg/g of dry weight(DW) of powder] whereas, in the ethyl acetate fraction, it was moreconcentrated showing values by 522.05 (�15.5) mg/g of extract(corresponding to 11 mg/g of DW of powder). As previously describedin Section 3.3 the phytotoxicity of the ethyl acetate fraction washigher than the crude extract and this difference in biological activitycould be due to its higher concentration in phenolic compounds. Infact, comparing the ED50 values (Table 2), the phytotoxicity of theethyl acetate fraction on both germination and root growth of lettucewas 3.2- and 2-fold, respectively, higher than the crude extract. Thephytotoxicity of phenolic compounds on seed germination and plant
Table 5Chemical characterization of the n-hexane fraction obtained through the bio-
guided fractionation of the crude extract of L. ornithopodioides (RT – retention time),
area % (relative area percentage).
RT Area % Molecules
46.384 5.72 Neophytadiene
46.552 1.25 Phytone
49.350 3.55 Methyl hexadecanoate
51.535 15.62 Ethyl hexadecanoate
54.788 3.16 Linolenate <methyl->
55.157 30.46 Phytol
55.649 2.82 Methyl octadecanoate
56.006 3.09 Linoleic acid
56.622 5.12 9,12-Octadecadienoyl chloride, (z,z)
57.625 7.69 Ethyl octadecanoate
Please cite this article in press as: Araniti, F., et al., Phytotoxic activity
spontaneous species of Mediterranean area. Phytochem. Lett. (2013
growth was largely documented (Araniti et al., 2013; Reigosa andMalvido-Pazos, 2007; Yang et al., 2002). Moreover, several authors(An et al., 2000; Araniti et al., 2013; Djurdjevic et al., 2004) observed,during fractionation, a good affinity of the phenolic compounds withethyl acetate solvent, which was often used to extract phytotoxinsfrom plant material (Araniti et al., 2012a; Das Neves and Gaspar,1990).
3.5. Phenolic characterization through HPLC
In the present study, the chemical characterization of phenoliccompounds was focused on the most common phenolics largelyknown for their phytotoxicity and their implication in allelopathicphenomenon (Reigosa and Malvido-Pazos, 2007). Through HPLCanalysis, in the ethyl acetate fraction of L. ornithopodioides eightphenolic compounds have been identified: gallic, caffeic, chloro-genic, sinapic, vanillic, syringic, p-coumaric and ferulic acids (datanot shown). Among secondary metabolites, phenolic compoundsrepresent one of the largest group, more investigated in relation totheir ecological role in plant–soil system (Siqueira et al., 1991) andin plant–plant interaction. Several studies demonstrated theirinfluence on various physiological processes such as cell extension,membrane permeability, protein synthesis, enzymatic activity, ionbalance, chlorophyll synthesis, photosynthesis, plant–water rela-tionships and respiration (Einhellig, 2004). As reported by Einhelligand Rasmussen (1979) and Patterson (1981), ferulic, p-coumaric,and vanillic acids greatly decreased the biomass associated withreduced chlorophyll content in leaves of soybean. Moreover, Yanget al. (2002) demonstrated that ferulic and p-coumaric acidsstrongly affected the Mg-chelatase causing a reduction inaccumulation of chlorophyll and porphyrin contents. Vanillic acidhas also been shown to interfere with the mitochondrialmetabolism reducing Ca2+ uptake (Demos et al., 1975). Further-more, Reigosa and Malvido-Pazos (2007) found that syringic,sinapic and gallic acids strongly influenced the growth ofArabidopsis thaliana.
Some of the phenols characterized in L. ornithopodioides extractwere also known to exhibit selective physio-ecological implica-tions. There are remarkable evidence that gallic and ferulic acidsand some other phenolics adversely affected nitrogen cycle
Table 6Chemical characterization of the chloroformic fraction obtained through the bio-
guided fractionation of the crude extract of L. ornithopodioides (RT – retention time),
area % (relative area percentage).
RT Area % Molecules
6.133 5.10 Diacetone alcohol
6.228 9.52 Cyclopentanol, 3-methyl
7.887 18.37 4-Methyl-2-heptanone
9.816 18.70 6-Methyl 2-heptanol
23.782 36.15 Salicyl alcohol
56.871 12.16 Ethyl 9z-octadecenoate
and phytochemical characterization of Lotus ornithopodioides L., a), http://dx.doi.org/10.1016/j.phytol.2013.08.019

F. Araniti et al. / Phytochemistry Letters xxx (2013) xxx–xxx 5
G Model
PHYTOL-595; No. of Pages 5
bacteria (Rice and Pancholy, 1974) and the high presence in the soilof phenolics compounds released in the environment by through-fall, litter, and organic layer decomposition was responsible of theregeneration failure of spruce (Gallet, 1994; Gallet and Lebreton,1995).
References
An, M., Haig, T., Pratley, J.E., 2000. Phytotoxicity of vulpia residues: II. Separation,identification, and quantification of allelochemicals from Vulpia myuros. J.Chem. Ecol. 26, 1465–1476.
Anjum, T., Bajwa, R., 2010. Isolation of bioactive allelochemicals from sunflower(variety Suncross-42) through fractionation-guided bioassays. Nat. Prod. Res.24, 1783–1788.
Araniti, F., Lupini, A., Mercati, F., Statti, G.A., Abenavoli, M.R., 2013. Calaminthanepeta L. (Savi) as source of phytotoxic compounds: bio-guided fractionation inidentifying biological active molecules. Acta Physiol. Plant. 1–10.
Araniti, F., Lupini, A., Sorgona, A., Conforti, F., Marrelli, M., Statti, G.A., Menichini, F.,Abenavoli, M.R., 2012a. Allelopathic potential of Artemisia arborescens: isola-tion, identification and quantification of phytotoxic compounds through frac-tionationguided bioassays. Nat. Prod. Res. 1–8.
Araniti, F., Sorgona, A., Lupini, A., Abenavoli, M.R., 2012b. Screening of Mediterra-nean wild plant species for allelopathic activity and their use as bio-herbicides.Allelopath. J. 29, 107–124.
Belz, R.G., Hurle, K., Duke, S.O., 2005. Dose–response – a challenge for allelopathy?Nonlinearity Biol. Toxicol. Med. 3, 173–211.
Chaimovitsh, D., Abu-Abied, M., Belausov, E., Rubin, B., Dudai, N., Sadot, E., 2010.Microtubules are an intracellular target of the plant terpene citral. Plant J. 61,399–408.
Chiapusio, G., Sanchez, A.M., Reigosa, M.J., Gonzalez, L., Pellissier, F., 1997. Dogermination indices adequately reflect allelochemical effects on the germina-tion process? J. Chem. Ecol. 23, 2445–2553.
Chon, S.U., Choi, S.K.C., Jung, S., Jang, H.G., Pyo, B.S., Kim, S.M., 2002. Effects of alfalfaleaf extracts and phenolic allelochemicals on early seedling growth and rootmorphology of alfalfa and barnyard grass. Crop Prot. 21, 1077–1082.
Das Neves, H.J.C., Gaspar, E.M.M., 1990. Identification of active compounds in wheatstraw extracts with allelopathic activity by HRGC–MS and HRGC-FTIR. J. HighResolut. Chromatogr. 13, 550–554.
Demos, K., Woolwine, M., Wilson, R.H., McMillan, C., 1975. The effects of tenphenolic compounds on hypocotyls growth and mitochondrial metabolismof mung bean. Am. J. Bot. 62., 97–102.
Djurdjevic, L., Dinic, A., Pavlovic, P., Mitrovic, M., Karadzic, B., Tesevic, V., 2004.Allelopathic potential of Allium ursinum L. Biochem. Syst. Ecol. 32, 533–544.
Duke, S.O., Dayan, F.E., Rimando, A.M., Shrader, K., Aliotta, G., Oliva, A., Romagni, J.G.,2002. Chemicals from nature for weed management. Weed Sci. 50, 138–151.
Duke, S.O., Dayan, F.E., Romagni, J.G., Rimando, A.M., 2000. Natural products assources of herbicides: current status and future trends. Weed Res. 40, 99–111.
Einhellig, F.A., Rasmussen, J.A., 1979. Effects of three phenolic acids on chlorophyllcontent and growth of soybean and grain sorghum seedlings. J. Chem. Ecol. 5,815–824.
Einhellig, F.A., 2004. Mode of allelochemical action of phenolic compounds. In:Macias, F.A., Galindo, J.C.G., Molinillo, J.M.G., Cutler, H.G. (Eds.), Allelopathy.Chemistry and Mode of Action of Allelochemicals. CRC Press, Boca Raton, pp.217–238.
Gallet, C., 1994. Allelopathic potential in bilberry-spruce forests: influence ofphenolic compounds on spruce seedlings. J. Chem. Ecol. 20, 1009–1024.
Gallet, C., Lebreton, P., 1995. Evolution of phenolic patterns in plants and associatedlitters and humus of a forest ecosystem. Soil Biol. Biochem. 27, 157–165.
Please cite this article in press as: Araniti, F., et al., Phytotoxic activity
spontaneous species of Mediterranean area. Phytochem. Lett. (2013
Gross, E.M., 2001. Seasonal and spatial dynamics of allelochemicals in the sub-mersed macrophyte Myriophyllum spicatum L. Verh. Internat. Verein. Theor.Angew. Limnol. 2, 2116–2119.
International Allelopathy Society, 1996. Constitution. Available at: http://www.a-s.uca.es/bylaws.htm#CONSTI.
Jones, C.D., Woods, K.E., Setzer, W.N., 2012. A chemical ecological investigation ofthe allelopathic potential of Lamium amplexicaule and Lamium purpureum. OpenJ. Ecol. 2 (4) 167–177.
Kalinova, B., Kindl, J., Jiros, P., Zacek, P., Vasickova, S., Budesinsky, M., Valterova, I.,2009. Composition and electrophysiological activity of constituents identifiedin male wing gland secretion of the bumblebee parasite Aphomia sociella. J. Nat.Prod. 72, 8–13.
Kobaisy, M., Tellez, M.R., Webber, C.L., Dayan, F.E., Schrader, K.K., Wedge, D.E., 2001.Phytotoxic and fungitoxic activities of the essential oil of kenaf (Hibiscuscannabinus L.) leaves and its composition. J. Agric. Food Chem. 49 (8)3768–3771.
Macias, F.A., Lacret, R., Varela, R.M., Nogueiras, C., Molinillo, J.M.G., 2010. Isolationand phytotoxicity of terpenes from Tectona grandis. J. Chem. Ecol. 36, 396–404.
Morgan, E.C., Overholt, W.A., 2005. Potential allelopathic effects of Brazilian pepper(Schinus terebinthifolius Raddi, Anacardiaceae) aqueous extract on germinationand growth of selected Florida native plants. J. Torrey Bot. Soc. 132, 11–15.
Navarro-Silva, M.A., Marques, F.A., Duque, J.E., 2009. Review of semiochemicals thatmediate the oviposition of mosquitoes: a possible sustainable tool for thecontrol and monitoring of Culicidae. Rev. Bras. Entomol. 53, 1–6.
Nielsen, O.K., Ritz, C., Streibig, J.C., 2004. Nonlinear mixed-model regression toanalyze herbicide dose–response relationships 1. Weed Technol. 18 (1) 30–37.
Patterson, D.T., 1981. Effects of allelochemicals on growth and physiologicalresponces of soybean (Glycine max). Weed Sci. 29, 53–59.
Petersen, J., Belz, R., Walker, F., Hurle, K., 2001. Weed suppression by release ofisothiocyanates from turnip-rape mulch. Agron. J. 93 (1) 37–43.
Pignatti, S., 1982. Flora d’Italia, 3 vols.. Edagricole, Bologna.Reigosa, M.J., Souto, X.C., Gonzalez, L., 1999. Effect of phenolic compounds on the
germination of six weeds species. Plant Growth Regul. 28, 83–88.Reigosa, M.J., Malvido-Pazos, E., 2007. Phytotoxic effects of 21 plant secondary
metabolites on Arabidopsis thaliana germination and root growth. J. Chem. Ecol.33, 1456–1466.
Reynolds, T., 1987. Comparative effects of alicyclic compounds and quinones oninhibition of lettuce fruit germination. Ann. Bot. Lond. 60, 215–223.
Rice, E.L., Pancholy, S.K., 1974. Inhibition of nitrification by climax ecosystems. III.Inhibitors other than tannins. Am. J. Bot. 1095–1103.
Rimando, A.M., Olofsdotter, M., Dayan, F.E., Duke, S.O., 2001. Searching for riceallelochemicals: an example of bioassay-guided isolation. Agron. J. 93, 16–20.
Satoshi, N., Shingo, Y., Masaaki, H., 2010. Anti-cyanobacterial fatty acids releasedfrom Myriophyllum spicatum. Hydrobiologia 543, 71–78.
Seaney, R.R., Henderson, P.R., 1970. Birdsfoot trefoil. Adv. Agro. 22, 119–157.Singleton, V., Rossi Jr., J.A., 1965. Colorimetry of total phenolics with phosphomo-
lybdic–phosphotungstic acid reagents. Am. J. Enol. Vit. 16, 144–158.Siqueira, J.O., Nair, M.G., Hammerschmidt, R., Safir, G.R., Putnam, A.R., 1991.
Significance of phenolic compounds in plant–soil–microbial systems. Crit.Rev. Plant Sci. 10, 63–121.
Streibig, J.C., Kudsk, P., Jensen, J.E., 1998. A general joint action model for herbicidemixtures. Pestic. Sci. 53, 21–28.
Weston, L.A., Harmon, R., Mueller, S., 1989. Allelopathic potential of sorghum–sudangrass hybrid (sudex). J. Chem. Ecol. 15, 1855–1865.
Yang, C.M., Lee, C.N., Zhou, C.H., 2002. Effects of three allelopathic phenolics onchlorophyll accumulation of rice (Oryza sativa) seedlings. I. Inhibition of supply-orientation. Bot. Bull. Acad. Sin. 43, 299–304.
Yu, J.Q., Ye, S.F., Zhang, M.F., Hu, W.H., 2003. Effects of root exudates and aqueousroot extracts of cucumber (Cucumis sativus) and allelochemicals, on photosyn-thesis and antioxidant enzymes in cucumber. Biochem. Syst. Ecol. 31, 129–139.
and phytochemical characterization of Lotus ornithopodioides L., a), http://dx.doi.org/10.1016/j.phytol.2013.08.019





























