Embed Size (px)
Citation preview

Physiology and Pathophysiology of the Venous System
Fedor Lurie*Jobst Vascular Institute, ProMedica Toledo Hospital, Toledo, OH, USA
Abstract
Venous physiology and pathophysiology are described in relationship to two main functions of theperipheral veins – venous volume and venous return. These two interconnected functions aredetermined by basic physiological characteristics of venous vessels such as compliance, capaci-tance, and resistance. Special attention is paid to the role of the valves as modulators of venousfunction. Pathological changes in the vein wall in primary and postthrombotic disease and theresultant changes in venous function are described, but the details of the molecular mechanisms areleft for further reading.
Glossary of Terms
Compliance Ability of a vein to increase its volume in response to a change in transmuralpressure.
Transmuralpressure
The difference in pressure between two sides of the vein wall.
Valve cycle The phasic changes in flow and in geometrical configuration of venous valvesduring the time period between two consecutive closures of the valve.
Venousresistance
The ratio of the total energy gradient across the venous segment over the flowrate through this segment.
Introduction
The veins are components of the circulatory system, and their function should always be viewed asan integral part of the blood circulation. In addition to serving as a conductor between themicrocirculation and the heart, veins are capacitance vessels, providing the ability to sequestersignificant volume of blood from the circulation and recruit necessary volume back into thecirculation as needed.
Anatomically, the venous system differs significantly from the arterial side of the circulation. Thevenous wall is structurally different, which explains the uniqueness of its mechanical properties suchas collapsibility and compliance. The systematic organization of veins in the extremity into deep andsuperficial networks connected by perforating veins has no arterial analogy. Intramuscular sinusoidsand, most importantly, venous valves are unique to venous system.
*Email: [email protected]
*Email: [email protected]
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 1 of 19

Such profound differences reflect the distinct functions of the venous system. In describing keyaspects of venous physiology and pathophysiology, this chapter will concentrate on the uniquefunctions of venous capacitance and volume and the mechanisms involved in venous return.
Venous Volume
At any given time up to 80 % of intravascular blood volume resides in the veins, underscoring thephysiological importance of venous capacitance. Proportionally small changes in venous volumemay result in significant changes in venous return to supply the heart with the necessary volume ofblood when circulatory demands change. Changing body position, exercising, being exposed tomotor vehicle or aircraft acceleration, or simply digesting causes a dramatic redistribution of bloodvolume in the vascular system without affecting cardiac function. Reduction in blood volume due todehydration or bleeding can be acutely compensated by recruitment of blood from the veins.
In abnormal conditions, however, sudden changes in venous volume may result in insufficientreturn, or cardiac overload, affecting the entire circulatory system. Changes in venous volume are inlarge part determined by mechanical properties of the venous wall and the activity of smooth musclecells (venous tone). The measure of the responsiveness of the venous wall to pressure changes iscompliance, defined as the ratio of the change in volume to the change in transmural pressure. Thevolume-pressure curve of an isolated venous segment is S-shaped and has three distinct areas(Fig. 1). At low levels of transmural pressure, the compliance of the venous wall is high, andsmall changes in pressure cause large volume changes. At high levels of transmural pressure, venouscompliance is very low, and large changes in pressure cause very small volume changes. Whenpressure outside the vein exceeds intramural pressure (negative transmural pressure), the vein cancollapse, and the compliance is very low.
High venous compliance in the physiological range of venous pressure leads to significant shiftsin blood volume when the position of the body changes. For example, the transition from supine toa standing position changes the hydrostatic pressure, leading to a rapid increase in leg vein volumeby approximately 400 ml, with an additional increase of 100 ml after 1 min (Rushmer 1976). Suchvolume shifts trigger a range of vasoactive responses to maintain constant cardiac output. Bloodflow through the liver, kidney, and mesenteric vessels significantly decreases (Culbertsonet al. 1951). Failure or delay in the action of these mechanisms may result in syncope.
Fig. 1 Volume-pressure curve of an isolated venous segment
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 2 of 19

These mechanical properties are only basic components of the complex behavior of the venouswall that determines venous capacitance and volume. Changes in venous tone, anatomical connec-tions between the venous wall and surrounding tissues, and pathological changes in the wall, thelumen, and around the vein significantly affect the mechanical behavior of the venous wall. In situ,veins are intimately connected to surrounding connective tissue structures, which prevent most veinsfrom collapsing, even when transmural pressure is negative. The presence of synechia inpostthrombotic veins limits the ability of the vein to extend its lumen. Fibrosis in the venous walland surrounding tissue as a result of inflammation significantly decreases wall compliance.
Venous tone plays an important physiological role in regulating blood flow. In addition tomechanical autoregulation, changes in smooth muscle activity in the tunica media are mediatedthrough bioactive substances released by nerves, endothelium, or from circulating blood, and arecontrolled largely by sympathetic alpha adrenergic activity. When veins contract to increase cardiacoutput, their resistance increases, but this increase is usually offset by arteriolar dilation and activityof the muscle pump. Intramuscular sinusoids lack adrenergic innervation and do not constrict,maintaining the ability of the muscle pump to accumulate the necessary volume of blood (Stepherd1966). Changes in superficial vein volume constitute one of the thermoregulatory mechanisms.These changes can be mediated by sympathetic activity, but also directly by temperature changes inthe environment. Dilated veins serve as heat exchangers, and constricted veins conserve energy ina cold environment.
Venous Return
Veins are an integral part of the circulation which, from a mechanical point of view, is a closedsystem of blood vessels connected to the heart. The only exception is the microcirculation wherefluid exchange between intravascular and extravascular spaces occurs. This fluid exchange innormal and in chronic pathological conditions is volume neutral, so the changes in intravascularblood volume are negligible. Acute volume changes, such as during shock, are a special concern thatis beyond the subject of this chapter. Blood flow in individual veins and regions may changedramatically over time without affecting the total venous return to the heart or venous return fromthe region or organ. Bending the knee or elbow, for example, can temporarily stop venous flow insome of the extremity’s veins, but the resulting venous outflow from this extremity over timeremains equal to arterial inflow. During acute venous thrombosis (except rare cases of phlegmasia),collateral flow develops over a very short time, balancing the venous outflow with arterial inflow.Thus the term “venous stasis” describes a phenomenon that can only exist in an individual venoussegment and only during a relatively short time.
In general, blood moves because of and in the direction of an energy gradient. Two components oftotal blood energy relevant to a discussion of blood dynamics are kinetic and potential energy.Kinetic energy is related to the energy of a moving mass and is directly proportional to velocity andspecific gravity of the blood. Potential energy is associated with a set of forces that act on a body ina way that depends only on the body’s position in space. It is frequently associated with gravitationalenergy, which is possessed by virtue of height above the surface of the earth. It is important tomention that other components of potential energy are actively involved in the dynamic of thevenous flow. An example is potential energy of the blood in the intramuscular sinusoid duringmuscle relaxation. The kinetic energy of muscle contraction converts this energy of the blood intoa kinetic energy of the flow. This is similar to a bow and arrow, where the potential energy in the bent
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 3 of 19

limbs of the bow when the string is drawn back can become the kinetic energy of an arrow when thestring is released.
While physical principles and laws are helpful in understanding some of the blood flow phenom-ena, they are limited to idealized models, such as ideal nonviscous fluid, rigid tubes, andone-dimensional elementary stream of flow. The real flow in the veins is far from such idealizedmodels, and the data derived from physical and computational models should be interpreted withcaution.
An example where data interpretation may not be straightforward is related to the use of pressuregradient instead of energy gradient. Since some components of pressure are measurable, many of thephysiological studies related to venous flow were focused on pressure changes in veins. Wheninterpreting these data, it is important to remember that measured pressure is only a component ofblood energy, and bloodmay and does flow against pressure gradient when inertial forces and kineticenergy are sufficient. Examples are the flow from left ventricle to aorta at the end of systole and theflow in the venous segment after closure of distal valve. These phenomena take place during veryshort time intervals and are important primarily to hemodynamic events related to the work ofvalves.
Blood flow in the veins of the lower extremities is grossly influenced by conditions in the moreproximal veins. As the venous track passes the abdominal and thoracic compartments, the changesin their volumes and pressures modify the venous return from more distal veins. The abdominalcontents have a density similar to blood-specific gravity; therefore, the changes in hydrostaticpressure in the abdominal veins are balanced by a similar change in intra-abdominal pressure.Subatmospheric pressure in the thorax maintains positive transmural pressure in the intrathoracicveins despite intravascular pressure drops caused by the heart, preventing venous collapse. Theinfluence of respiration on the pattern of blood flow in the veins of the lower extremities depends onthe movements of the diaphragm. During inspiration, the diaphragm descends, increasing intra-abdominal pressure and decreasing the flow in the femoral veins. In expiration, the diaphragm rises,decreasing abdominal pressure and increasing blood flow in the femoral veins. As the relative role ofthe diaphragm in the respiratory cycle changes with the position of the body, so does the pattern ofvenous flow. In a slightly head down position, venous flow in the femoral veins depends solely onthe cardiac cycle, whereas in a standing position, it depends upon the respiratory cycle (Monetaet al. 1988).
Venous ResistanceThe term venous resistance was introduced to quantify the blood flow through a venous segment.Resistance (R) is the ratio of the total energy gradient across the segment (DE) over the flow ratethrough this segment (Q):
R ¼ DE=Q
For practical reasons, the dynamic pressure gradient (DPd) is substituted for the total energygradient, which introduces substantial limitations and sometimes leads to inaccurate interpretationsof physiological phenomena. Resistance of a venous segment is not constant, but changes with thevelocity of blood flow, variations of transmural pressure, and blood viscosity. Around venousvalves, the constantly changing position of the cusps and the dimensions of the sinus causecontinuous changes in resistance. The collapsibility of veins also contributes to the changingresistance, leading to very complex flow dynamic phenomena. Hargens et al. demonstrated thatjugular vein pressure in an upright giraffe does not fall with height, as it would in a vertical column of
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 4 of 19
fluid at rest, but actually increases (Hargens 1987), suggesting considerable resistance to flow in thejugular vein. A likely explanation is that the vein must be significantly collapsed, and a reducedcross-sectional area leads to increased resistance to the flow (Pedley 1987). They also noted thehigher number of venous valves in the distal neck compared to a few valves in the proximal neck.The orifice of the venous valves at their maximal opening have a smaller cross-sectional area thanthe adjoining venous segments, creating additional resistance, which may also explain these findings(Franklin and Richardson 1937; Lurie et al. 2002). Studying similar phenomena in humans isdifficult due to the smaller anatomical scale, but it is likely that similar influences on pressure andflow exist in human veins.
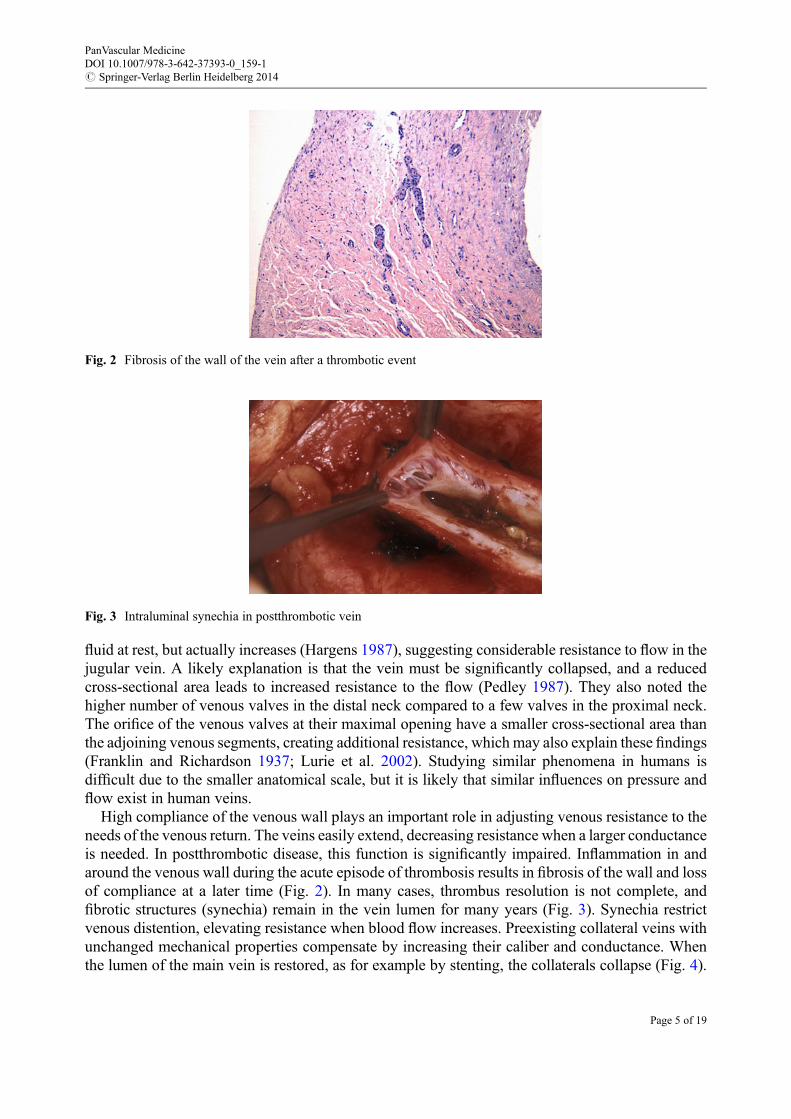
High compliance of the venous wall plays an important role in adjusting venous resistance to theneeds of the venous return. The veins easily extend, decreasing resistance when a larger conductanceis needed. In postthrombotic disease, this function is significantly impaired. Inflammation in andaround the venous wall during the acute episode of thrombosis results in fibrosis of the wall and lossof compliance at a later time (Fig. 2). In many cases, thrombus resolution is not complete, andfibrotic structures (synechia) remain in the vein lumen for many years (Fig. 3). Synechia restrictvenous distention, elevating resistance when blood flow increases. Preexisting collateral veins withunchanged mechanical properties compensate by increasing their caliber and conductance. Whenthe lumen of the main vein is restored, as for example by stenting, the collaterals collapse (Fig. 4).
Fig. 2 Fibrosis of the wall of the vein after a thrombotic event
Fig. 3 Intraluminal synechia in postthrombotic vein
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 5 of 19
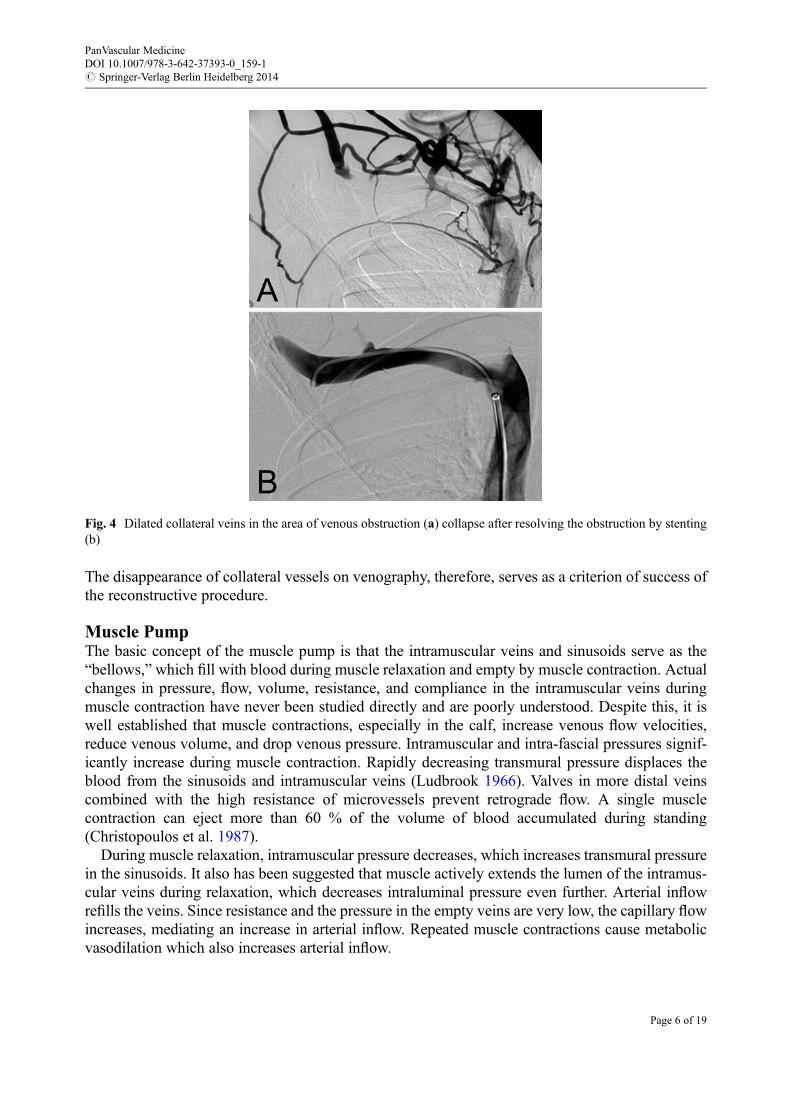
The disappearance of collateral vessels on venography, therefore, serves as a criterion of success ofthe reconstructive procedure.
Muscle PumpThe basic concept of the muscle pump is that the intramuscular veins and sinusoids serve as the“bellows,” which fill with blood during muscle relaxation and empty by muscle contraction. Actualchanges in pressure, flow, volume, resistance, and compliance in the intramuscular veins duringmuscle contraction have never been studied directly and are poorly understood. Despite this, it iswell established that muscle contractions, especially in the calf, increase venous flow velocities,reduce venous volume, and drop venous pressure. Intramuscular and intra-fascial pressures signif-icantly increase during muscle contraction. Rapidly decreasing transmural pressure displaces theblood from the sinusoids and intramuscular veins (Ludbrook 1966). Valves in more distal veinscombined with the high resistance of microvessels prevent retrograde flow. A single musclecontraction can eject more than 60 % of the volume of blood accumulated during standing(Christopoulos et al. 1987).
During muscle relaxation, intramuscular pressure decreases, which increases transmural pressurein the sinusoids. It also has been suggested that muscle actively extends the lumen of the intramus-cular veins during relaxation, which decreases intraluminal pressure even further. Arterial inflowrefills the veins. Since resistance and the pressure in the empty veins are very low, the capillary flowincreases, mediating an increase in arterial inflow. Repeated muscle contractions cause metabolicvasodilation which also increases arterial inflow.
Fig. 4 Dilated collateral veins in the area of venous obstruction (a) collapse after resolving the obstruction by stenting(b)
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 6 of 19

The functional competency of venous valves determines the pathways of blood displaced fromintramuscular veins and the effectiveness of the pump. Competent valves in perforating veins secureunidirectional flow from deep into the superficial veins. Competent valves in the superficial veins arenecessary for further propagation of the flow into more proximal deep veins. Incompetent valvesallow the blood to return into the intramuscular veins and sinusoids, competing with arterial inflowand decreasing effectiveness of the muscle pump.
While it is clear that the muscle pump increases venous flow, other aspects of its function remaincontroversial. Metabolic activity during contractions requires an increase in blood supply to themuscle. One of suggested mechanisms of exercise-induced muscle hyperemia is a combination ofvasodilation and increased cardiac output caused by mobilization of peripheral blood volume(Laughlin 1987). This theory is largely based on the observation that the highest possible perfusionby a muscle pump happens during locomotion in upright position. It suggests that during relaxation,the pressure in the intramuscular veins is negative (subatmospheric), which increases the arteriove-nous gradient causing increased perfusion. Since it is not known whether negative pressure can infact be produced, this mechanism remains largely hypothetical.
If negative pressure in muscular veins drives the arterial perfusion, reflux can affect this bycompeting with arterial inflow and filling of intramuscular veins. More severe reflux, especially indeep veins, should have a larger impact on arterial inflow. The existing evidence, however, makesthis hypothesis questionable. Direct measurements demonstrated that the increase in arterial bloodflow during muscle exercise does not correlate with changes in venous pressure or venous outflow(Valic et al. 2005). Compartment pressures and venous pressures during foot movements andwalking in healthy volunteers demonstrated that dramatic increases in intra-compartment pressurestranslate into a negligible change in venous pressure (Alimi et al 1994). Although foot movementsand walking affected pressures in the superficial veins, they had minimal influence on deep veinpressures. A study of patients with venous disease not only confirmed these findings, but alsoshowed prolonged increase in deep vein pressures caused by exercise in 20 % of the limbs. None ofthe limbs with this exercise-induced pressure increase had great saphenous vein (GSV) reflux, andall limbs with GSV reflux showed a decrease in deep venous pressure with exercise (Neglen andRaju 2000). In other words, superficial vein pressure does not reflect pressure changes in the deepsystem during exercise, and the GSVreflux does not necessarily affect venous pressures. In addition,the arterial flow to the contracting muscle is not entirely dependent on the arterial-venous pressuregradient, or on vasodilation. With muscle vasculature isolated from the systemic circulation, thecontracting muscle can perfuse itself (Sheriff and Van Bibber 1998), and an increase in muscle bloodflow can occur without vasodilation (Sheriff 2003). It also seems quite clear that a vascular waterfalleffect exists during muscle contraction (Naamani et al. 1995). This means that after a certain level,the decrease in venous pressure no longer affects arterial flow.
The concept of the muscle pump is one of the fundamental components of clinical phlebology, butmany of its elements remain theoretical, having never been confirmed experimentally.
Venous ValvesThe apparent simplicity of venous valve anatomy initially left little room for speculation regardingits function. Since its discovery, the venous valve was viewed as a passive membrane that opens andcloses the vessel’s lumen (Scultetus et al. 2001). Experimental demonstration of unidirectionalvenous flow was first described in Exercitatio Anatomica de Motu Cordis et Sanguinis inAnimalibus and was essential for modern understanding of the cardiovascular system (Harvey1978). The challenge to this concept came centuries later when E.B. Carrier described the intricateblood flow around the venous valve leaflets observed in the bat’s wing (Franklin and Richardson
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 7 of 19

1937). He also reported that venous valve leaflets do not open widely enough to touch the sinus wall,a finding confirmed by in vitro experimentation with human saphenous valves (McCaughanet al. 1984). These findings demonstrated that the complex hemodynamics around the valve farexceed the simple sequence of forward and backward flow and challenged the existing concept ofsimple physics behind closing of the venous valve.
Contemporary ultrasound techniques for testing venous valve function rely on Doppler-basedregistration of reversed blood flow in response to Valsalva or rapid compression-decompressionmaneuvers. This approach advanced venous diagnosis by providing a reliable tool for refluxdetection. Unfortunately, the indirect approach also created confusion between the presence ofreversed flow in the vein and function of the valve itself. The terms “reflux time” and “valve closuretime” were falsely used interchangeably. As a consequence, the view that reversed flow through thevalve is necessary for valve closure was promulgated (van Bemmelen et al. 1990). In addition, thedefinition of valve function was reduced to the ability of the venous valve to close in response toextreme forces (van Bemmelen et al. 1989). A new generation of ultrasound equipment, particularlythe introduction of B-flow modality, has made it possible to observe venous valve and blood flow inthe area of the valve in undisturbed physiological conditions. Artificial maneuvers to force the bloodbackward to check valve competency are no longer needed for normal valve observations. One cansimultaneously observe the motions of valve leaflets, changes in venous sinus shape and size, andblood flow through the valve during a normal respiratory cycle in different positions of the body andduring exercises such as dorsal and plantar flexion of the foot.
The engineering of artificial valves revived interest in studying the mechanism of valve closure.As a result, advanced physical and computational models were developed and used to test some ofthe assumptions of the current theory. These models showed that the geometry of valves plays animportant role in its function. The depth of valve pockets, for example, determines its competency:deeper sinuses provide better reaction to reversed flow. At the same time, deep sinuses increasechances of blood stagnation (Buescher et al. 2005). Interestingly, the depth of valve pockets has littleeffect on prograde flow. One of the challenges in designing artificial valves is finding a geometricalcompromise between maximizing the ability of the valve to prevent retrograde flow and minimizingthe valve’s resistance to prograde flow, but the variety of configurations in natural valves demon-strate that optimization to reaction to a reversed flow is not always nature’s intent.
The Valve CycleThe flow andmovements of the valve appear to be two parts of the same physiological process of the“valve cycle” – the time period between two consecutive closures of the valve, which we arbitrarilydivided into four phases (Lurie et al. 2003; Fig. 5).
During the opening phase, the cusps move from the closed position toward the sinus wall. Thisphase lasts on average 0.3 s when the patient is in the horizontal position. After reaching a certainpoint, the valves cease opening and enter the equilibrium phase. During this phase the leading edgesremain suspended in the flowing stream and undergo oscillations that resemble the flutter of flags inthe wind. The valve is maximally open during this phase yet the cusps maintain their position atsome distance from the wall, creating a narrowing of lumen. The cross-sectional area between theleaflets is about two-thirds of the cross-sectional area of the vein distal to the valve. The flowaccelerates in this narrowed area, forming a proximally directed jet. The impact of the jet againsta layer of much slower-moving blood proximal to the valve results in reflection of flow in the muralparts of the stream. While the larger stream located in the center of the vessel is directed proximallyalong the axis of the vein, the smaller part of the flow turns into the sinus pocket behind the valve
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 8 of 19

cusp, forming a vortex along the sinus wall and mural side of the valve cusp before reemerging in themain stream in the vein.
As vortical flow persists, it applies pressure upon the mural surface of the valve cusps. When thepressures on the mural and luminal sides of the cusp are in equilibrium, the valve remains open andthe cusps float in the stream. This dynamic equilibrium is sustained by equilibrium in velocities ofthe two streams – vortex on the mural side and axial flow on the luminal side of the valve cusps.Changes in either stream can lead to valve closure, and the oscillations of the leading edges of theleaflets that occur during this equilibrium phase make the valve evenmore sensitive to small changesin flow.
When the venous flow rate increases distal to the valve, as occurs during foot movements, thevelocity of the flow between the valve cusps rapidly increases. This causes a fall in pressure on theluminal side of the cusp, and the cusps start moving toward the axis of the vessel, further constrictingthe lumen (Fig. 6). With rising pressures on the mural side and falling pressures on the luminal sideof the cusps, valve closure is favored. The closing phase ensues. The leaflets move synchronouslytoward the center. The cusps of the valve assume a symmetrical position at an equal distance fromthe walls on both sides of the sinus. This phase lasts 0.4 s when the patient is at rest and is shorterwhen foot movements are performed. The last phase is the closed phase, during which the cuspsremain closed.
Fig. 5 The “valve cycle”
Fig. 6 The mechanism of valve closure by prograde flow
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 9 of 19

The duration of the valve cycle and of each of its four phases depends upon body position. In thestanding position, the duration of the valve cycle closely relates to respiratory movements. Ina supine position, the duration of the cycle is shorter and is influenced by both respiratory andcardiac cycles. Muscle activity (dorsal and plantar flexions of the foot) causes shortening of theclosing phase. Every single foot movement causes significant increase of venous flow velocities andclosure of the valve. In the absence of forced reversed flow, the valve cusps consistently undergo allphases of the valve cycle. The local hemodynamic events, such as vortical flow in the sinus pocket,play important roles in valve operation. These hemodynamic events are predetermined by the shapeandmechanical properties of the sinus and valve cusps and constitute a self-sustained mechanism forcompetent valve operation.
The existence of two distinct mechanisms of venous valve closure, one as a reaction to reversal inpressure gradient and/or flow and the other by the changes in prograde flow, can explain a number ofphysiological events in the venous system. An example is the flow from the tributary into a segmentof vein between the valves. Henderson and Johnson in 1912 described the phenomenon of jetseparation after abrupt closure of a tube. Briefly, when a tube is clamped upstream, the downstreamjet separates and creates an area of negative pressure at the site of separation and a pressure wavetraveling downstream. This means that when the valve in a vein closes by the prograde flow, thepressure in the segment immediately downstream to this valve drops.When the second valve locatedmore proximally closes (Fig. 7), the low pressure is captured in the venous segment. Theseconditions favor the flow from the tributaries. The importance of this phenomenon is that thelocal flow in the venous net can be driven by the muscle pump, independent of hydrostatic pressureand the position of the body. The pressure trapping in the venous segments between valves has beendemonstrated in venous bypass grafts (Thubrikar et al. 1994; Robicsek et al. 1999). It is challengingto confirm pressure trapping in veins due to low physiological pressure ranges and velocities, but it islikely that the flow conditions demonstrated in these mechanically and geometrically identicalvessels occur in normal veins.
Valve Functions Other Than Prevention of RefluxThe blood flow in peripheral veins is more complex than in arteries and capillaries. At a given time itcan be continuous, pulsatile, or absent. The transitions from one state to another produce even more
Fig. 7 Sequence of flow events in a vein segment between two valves
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 10 of 19

complex fluid dynamic phenomena. The presence of valves complicates this even further as valvesnot only are moved by the flow, but participate in flow changes. In this sense, we can postulate thatthe venous valve has physiological functions other than securing unidirectional flow, broadlycharacterized as flowmodulation. The central role in providing this function belongs to the geometryof the valve and the distinct mechanical properties of leaflets, sinus walls, and the walls of adjoiningsegments.
During the opening and equilibrium phases of the valve cycle, a proximally directed jet forms inthe valve orifice. The shape of this orifice, with fused leaflets on both sides and gradual wideningtoward the center, significantly differs from the circular cross section of adjoining venous segments.As blood passes through the valve orifice, its velocity profile flattens. This makes possible theformation of organized flow patterns such as spiral flow. In the proximal segment of the GSVand inthe proximal femoral vein, for example, paired valves are positioned at small angles to each other(Lurie and Kistner 2012), thus rotating the flow as it passes proximally (Lurie and Kistner 2013).This is more likely to happen in segments with several closely located valves, such as crural veins.The existence of such organized flow patterns may conserve significant energy facilitating venousreturn.
The flow inside the valve pocket deserves special attention. It is essential in keeping valve leafletsaway from the venous wall and in closing of the valve. It also provides a “flush” of the valve pocketduring each valve cycle. Experiments have shown that when this flush does not occur regularly, theoxygen content of the blood inside the valve pockets drops. This occurs during time of continuousflow. With pulsatile flow, this flush occurs regularly, and the oxygen content in valve pockets is notdifferent from the rest of venous blood (Hamer et al. 1981). Avascularity of the valve leafletspredisposes them to ischemic damage, thus, prolonged time intervals of continuous flow followedby reperfusion of valve pockets may play an important role in the formation of venous thrombi(Malone and Agutter 2006). This is one likely explanation of the known fact that valve pockets aresites of initial thrombus development (Sevitt 1974). By closing in reaction to increasing flowvelocity, venous valves convert continuous flow into pulsatile, which secures adequate oxygenationof valve pockets and prevents formation of thrombi.
Recognition that venous valves have important functions beyond reflux protection poses newquestions about the role of valvular dysfunction in venous pathophysiology. Much attention has beendevoted to the hemodynamic consequences of reflux, but no data are available regarding the role ofdysfunctional valves in the development of thrombosis, or its impact on the blood flow. Studies ofblood flow interaction with the vascular wall showed that some flow patterns induce inflammatoryreactions, while other flow conditions can initiate anti-inflammatory protective changes in theendothelium (Began et al. 2006). The relationship between these endothelium reactions and specificchanges in the flow produced by a dysfunctional valve remains to be investigated.
Hemodynamic Role of Perforating VeinsThe venous system is rich in interconnections between the individual vessels. Establishing numer-ous collateral circulatory routes, these connections prevent disruption of the venous flow duringchanges in body and extremity position and compression of the veins by contracting muscles duringlocomotion or by external pressure. Crossing the proper fascia of the extremity, perforating veinsconnect the superficial and the deep venous systems of extremities. There are about 60 perforatingveins in a lower extremity (Thomson 1979). Although some anatomical studies have demonstratedthat all perforating veins have valves, others have shown that some are avalvular (Thomson 1979;Cotton 1961). The latter, however, can be found in a minority of extremities and not more frequentlythan two veins per extremity (Thomson 1979). These anatomical data are consistent with findings
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 11 of 19

from duplex scans of asymptomatic legs, of which only 3.6 % have bidirectional flow in theperforating veins (PVs) (Labropoulos et al. 1995). The hemodynamic role of PVs in normalcirculation has not been addressed by experimental or direct observational studies; it is unlikely,however, that the current theory could be meaningfully challenged. Whether the PVs providecollateral ways for blood flow to bypass areas of obstruction of the superficial veins in situationslike compression from outside during sitting or their presence facilitates the outflow from superficialveins when the pressure drops in the deep veins during walking or both, it is clear that the flow inhealthy PVs is directed from the superficial into the deep system.
The role and significance of incompetent PVs are less obvious. In the absence of functioningvalves, the blood flow in the PV is governed mainly by the energy gradient in adjoined segments ofthe superficial and deep systems.
Direct measurements of blood flow in incompetent PVs and venous pressure in superficial veinsdemonstrated that even when incompetent, the PVs can function similarly to the competent ones.During standing there is no blood flow through incompetent PVs, and during walking the net flow isfrom superficial to deep veins, despite the bidirectional nature of the flow during muscle contractions(Bjordal 1972).
Occlusion of incompetent PVs does not influence venous pressure in superficial veins, whileocclusion of an incompetent GSV normalizes hypertension (Bjordal 1972). In light of these findings,the concept of transmission of the higher blood pressure from deep into superficial veins viaincompetent PVs during ambulation becomes questionable. Indirectly this was confirmed by thedemonstration of independent superficial and deep venous pressure changes during ambulation(Neglen and Raju 2000).
The observations of an increasing number of incompetent PVs in legs affected by venous diseaseover time (Labropoulos et al. 2006; Gohel et al. 2005), and with increasing severity of the disease(Cooper et al. 2003; Sakurai et al. 1997), likely indicate the reaction of PVs to changing hemody-namic conditions rather than their active role in hemodynamic changes. This view is supported bythe resolution of PV incompetence in most limbs after correction of the superficial reflux.
Capillary Exchange
One of the frequent signs of venous disorders is swelling. For this reason much attention has beendevoted to studying the role of venous function in fluid exchange in microcirculation. Ernest Starlingpostulated the main principle governing exchange of fluid through the capillary wall in 1896 bynoting that capillary filtration is proportional to the difference between the hydraulic pressure andosmotic pressure gradients across the wall. The Starling equation reads as follows (Levick 2010):
Jv ¼ LpS Pc � Pi½ � � s pc � pi½ �ð Þ
where:
Jv is the net fluid movement between compartments.[Pc � Pi] �s [pc � pi] is the net driving force.Pc is the capillary hydrostatic pressure.Pi is the interstitial hydrostatic pressure.pc is the capillary oncotic pressure.pi is the interstitial oncotic pressure.
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 12 of 19

Lp is the hydraulic conductance of the wall.S is capillary surface.s is the reflection coefficient.
The interstitial pressure in the skin, subcutaneous adipose, and muscles is usually in the range of�0.3*/�3.6 mmHg in the supine position and does not change significantly with a change inposition from supine to sitting or standing (Hussmann et al. 2006). Interstitium is a complexsubstance containing collagen and elastin fibers and glycosaminoglycan chains (GAGs). GAGsnot only are hydrophilic, but also serve as the major factor determining interstitial permeability towater. Water and solutes occupy the spaces within the GAG matrix. Because of the extremely smallsize of these spaces, their resistance to the flow is high, and the mobility of interstitial water is verylow, preventing its displacement by gravity or local compression.
Measurements in human skin and muscle demonstrated that the net driving pressure is belowvenous capillary pressure (Levick 1991). Therefore, in normal conditions, filtration occurs along theentire capillary length, with no reabsorption. When arterial pressure falls, as during hypovolemicshock, absorption temporarily occurs in the venous capillary, but because of increasingsubglycocalyx protein concentration and decreasing interstitial pressure, it gradually decreasesand returns to a normal state (Michel and Phillips 1987). More detailed explanation of the absenceof reabsorption was provided by a two-layer membrane model that incorporates the glycocalyx andthe endothelial intercellular cleft (Weinbaum et al. 2003, 2007).
The rate of capillary filtration is normally very low. For example, the entire filtration in the foot ofa supine subject is about 0.22 mL/h per 100 g of tissue. Such low filtration is explained by the workof the small-pore system of glycocalyx and by vasomotion. The small-pore system transmits protein-free plasma ultrafiltrate into the subglycocalyx space. From this space, the ultrafiltrate has to passthrough narrow breaks in the junctional strands. At the same time, interstitial plasma proteins arediffused through the filtration system. The result is that the subglycocalyx protein concentration islower than the interstitial concentration, and the true transmembranic osmotic gradient is greaterthan predicted by the classical Starling equation (Adamson et al. 2004; Levick 2004).
Vasomotion, the rhythmic dilations and constriction of arterioles, leads to changes in capillarypressure. Capillary pressure primarily depends on precapillary resistance, which explains why therise of capillary pressure in the foot as a result of the transition from the supine to vertical position issignificantly smaller than the rise in arterial pressure (de Graaff et al. 2003). Each contractionreduces capillary pressure, decreasing filtration and giving time for lymphatic vessels to absorb fluid.As the result, the interstitial pressure also changes rhythmically (Hussmann et al. 2006).
Since no reabsorption occurs in the capillaries, the role of the lymphatic system is critical formaintaining fluid balance in the tissue. The endothelium of the lymphatic vessels is connected to thesurrounding interstitial matrix, especially tightly to adjacent collagen, by the anchoring filaments(Gerli et al. 1991; Leak and Burke 1966). These fibers pull the lymphatic endothelial cells apartwhen interstitial pressure increases, opening the junctions and increasing the passage of fluid intolymphatic vessels. The same mechanism closes junctions when interstitial pressure normalizes(Aukland and Reed 1993). Further transport of the lymph in the lymphatic vessels occurs by severalpumping mechanisms, including active peristaltic contractions of the vessels, function of lymphaticvalves, and movements of surrounding tissues.
Swelling in venous disease may be explained by increased filtration due to elevated venouspressure as a result of acute or chronic occlusion of major veins or by failure of lymphaticreabsorption or both. Inflammation may be another cause or contributing factor to the developmentof edema. The therapeutic goal in treating venous swelling is to decrease capillary filtration. This
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 13 of 19

may be achieved by mechanically increasing interstitial pressure (compression stockings) or bydecreasing intracapillary pressure (leg elevation).
Pathophysiology of the Venous Wall
The traditional focus of studies related to venous diseases has been related to the physiological andpathophysiological aspects of hemodynamics. The advantages of such an approach include theability to directly measure blood velocity, pressure, and volume in the leg of healthy individuals andthose affected by venous disease. Some hemodynamic abnormalities are correctable by surgery, thusproviding measurable physiological outcomes. As a result, a large body of knowledge has beenaccumulated regarding both the normal and pathological physiology of veins. Obviously, sucha reductionistic approach has many limitations. Clinicians may use this knowledge to guide theirpractice but it remains a difficult task to synthesize data from different scientific disciplines andinvestigate what is applicable to a specific patient. Thus, it is important to remember that not allaspects of venous disease can be explained by hemodynamic phenomena and that changinghemodynamics does not always result in the desirable clinical outcome.
Predisposition for development of venous disorders is known to be inherited. Althoughresearchers have proposed that some genetic variants play a role in the pathogenesis of venousdisorders (Zamboni et al. 2005; Tan et al. 2007; Mellor et al. 2007), no specific genetic candidate hasbeen sufficiently established, and the search continues. Regardless of the mechanism, exposure toknown risk factors, such as prolonged standing andmultiple pregnancies, is not sufficient to increasethe chances of venous disease and only does so in predisposed individuals. In normal individuals,exposure to hydrostatic pressure is necessary for maintaining venous tone and is responsible fordecreased distensibility of distal veins (Kolegard et al. 2009). Contrary to hydrostatic pressure-basedtheory, the major difference between healthy individuals and those affected by venous disease is notin their venous response to standing, but in impaired arteriolar vasodilatation (Klonizakis et al. 2006,2009). Age, on the other hand, has a strong association with incidence of primary chronic diseaseand the pattern of affected veins (Caggiati et al. 2006). The age-related changes in the venous wall,especially in the area of valves, are remarkably similar to the age-related changes in arteries (Saphirand Lev 1952; Merryman 2010).
Pathological processes in the venous wall affected by chronic venous disease are similar oridentical to changes in other tissues, suggesting that its cause is more systemic thanhemodynamic. Inflammatory cells in patients with venous disease are activated not only in venules,but in capillaries and arterioles of the skin in areas usually not involved in the disease and unaffectedby hemodynamic conditions (Pappas et al. 1997). Immunohistochemical study of skin biopsyspecimens from patients with primary chronic venous disease showed that predominant infiltratingcell types were T lymphocytes and macrophages. Surprisingly, expression of endothelial leukocyteadhesion molecule-1 and vascular cell adhesion molecule was not elevated, but intercellularadhesion molecule-1 expression did increase in more severely diseased skin. Dermal staining forboth interleukin (IL)-1 alpha and IL-1 beta was increased in severely liposclerotic skin, but this wasnot seen at an early stage. All changes were confined to the subpapillary region of the skin. Thesefindings contradict the hypothesis that inflammation in primary chronic venous disease is initiatedby abnormal blood flow and adhesion of leukocytes to the endothelium (Wilkinson et al. 1993).
Well-described different patterns of pathological changes may occur simultaneously in adjoiningsegments of the same vein exposed to the same flow conditions (Badier-Commander et al. 2001).While some of these changes may be interpreted as adaptive to disturbed flow, the majority of them
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 14 of 19

are the results of remodeling due to factors not related to the flow. The collagen imbalance in the wallof varicose veins, for example, is likely a result of vascular endothelial growth factor (VEGF)activated matrix metalloproteinases (MMPs) (Kockx et al. 1998; Sansilvestri-Morel et al. 2001,2005). However, the altered transcription of genes for VEGF and its receptors in veins is indepen-dent of the presence or absence of valvular incompetence (Hollingsworth et al. 2004).
Investigation of the molecular mechanisms of venous disorders is a very new area of research. Ithas already contributed significantly to our understanding of venous pathophysiology and over timeshould lead to development of new, potentially more effective therapeutic options.
References
Adamson RH, Lenz JF, Zhang X, Adamson GN, Weinbaum S, Curry FE (2004) Oncotic pressuresopposing filtration across non-fenestrated rat microvessels. J Physiol 557:889–907
Alimi YS, Barthelemy P, Juhan C (1994) Venous pump of the calf: a study of venous and muscularpressures. J Vasc Surg 20:728–735
Aukland K, Reed RK (1993) Interstitial-lymphatic mechanisms in the control of extracellular fluidvolume. Physiol Rev 73:1–78
Badier-Commander C, Couvelard A, Henin D, Verbeuren T, Michel JB, Jacob MP (2001) Smoothmuscle cell modulation and cytokine overproduction in varicose veins. An in situ study. J Pathol193:398–407
Bergan JJ, Schmid-Schonbein GW, Smith PD, Nicolaides AN, Boisseau MR, Eklof B (2006)Chronic venous disease. N Engl J Med 355:488–498
Bjordal RI (1972) Circulation patterns in incompetent perforating veins in the calf and in thesaphenous system in primary varicose veins. Acta Chir Scand 138:251–261
Buescher CD, Nachiappan B, Brumbaugh JM, Hoo KA, Janssen HF (2005) Experimental studies ofthe effects of abnormal venous valves on fluid flow. Biotechnol Prog 21:938–945
Caggiati A, Rosi C, Heyn R, Franceschini M, Acconcia MC (2006) Age-related variations ofvaricose veins anatomy. J Vasc Surg 44:1291–1295
Christopoulos DG, Nicolaides AN, Szendro G, Irvine AT, Bull ML, Eastcott HH (1987) -Air-plethysmography and the effect of elastic compression on venous hemodynamics of theleg. J Vasc Surg 5:148–159
Cooper DG, Hillman-Cooper CS, Barker SG, Hollingsworth SJ (2003) Primary varicose veins: thesapheno-femoral junction, distribution of varicosities and patterns of incompetence. Eur J VascEndovasc Surg 25:53–59
Cotton LT (1961) Varicose veins. Gross anatomy and development. Br J Surg 48:589–598Culbertson JW, Wilkins RW, Ingelfinger FJ, Bradley SE (1951) The effect of the upright posture
upon hepatic blood flow in normotensive and hypertensive subjects. J Clin Invest 30:305–311de Graaff JC, Ubbink DT, Lagarde SM, Jacobs MJ (2003) Postural changes in capillary pressure in
the hallux of healthy volunteers. J Appl Physiol (1985) 95:2223–2228Franklin KJ, Richardson KC (1937) A monograph on veins. C C Thomas, SpringfieldGerli R, Ibba L, Fruschelli C (1991) Ultrastructural cytochemistry of anchoring filaments of human
lymphatic capillaries and their relation to elastic fibers. Lymphology 24:105–112Gohel MS, Barwell JR, Wakely C, Minor J, Harvey K, Earnshaw JJ, Heather BP, Whyman MR,
Poskitt KR (2005) The influence of superficial venous surgery and compression on incompetentcalf perforators in chronic venous leg ulceration. Eur J Vasc Endovasc Surg 29:78–82
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 15 of 19

Hamer JD, Malone PC, Silver IA (1981) The PO2 in venous valve pockets: its possible bearing onthrombogenesis. Br J Surg 68:166–170
Hargens AR (1987) Gravitational cardiovascular adaptation in the giraffe. Physiologist 30:S15–S18Harvey W (1978) Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus. The Classics
of Medicine Library, BirminghamHollingsworth SJ, Powell G, Barker SG, Cooper DG (2004) Primary varicose veins: altered
transcription of VEGF and its receptors (KDR, flt-1, soluble flt-1) with sapheno-femoral junctionincompetence. Eur J Vasc Endovasc Surg 27:259–268
Husmann MJ, Barton M, Amann-Vesti BR, Franzeck UK (2006) Postural effects on interstitial fluidpressure in humans. J Vasc Res 43:321–326
Klonizakis M, Yeung JM, Lingam K, Nash JR, Manning G, Donnelly R (2006) Contrasting effectsof varicose vein surgery on endothelial-dependent and -independent cutaneous vasodilation in theperimalleolar region. Eur J Vasc Endovasc Surg 31:434–438
Klonizakis M, Tew G, Michaels J, Saxton J (2009) Exercise training improves cutaneous micro-vascular endothelial function in post-surgical varicose vein patients. Microvasc Res 78:67–70
Kockx MM, Knaapen MW, Bortier HE, Cromheeke KM, Boutherin-Falson O, Finet M (1998)Vascular remodeling in varicose veins. Angiology 49:871–877
Kolegard R, Mekjavic IB, Eiken O (2009) Increased distensibility in dependent veins followingprolonged bedrest. Eur J Appl Physiol 106:547–554
Labropoulos N, Delis KT, Nicolaides AN (1995) Venous reflux in symptom-free vascular surgeons.J Vasc Surg 22:150–154
Labropoulos N, Tassiopoulos AK, Bhatti AF, Leon L (2006) Development of reflux in the perforatorveins in limbs with primary venous disease. J Vasc Surg 43:558–562
Laughlin MH (1987) Skeletal muscle blood flow capacity: role of muscle pump in exercisehyperemia. Am J Physiol 253:H993–H1004
Leak LV, Burke JF (1966) Fine structure of the lymphatic capillary and the adjoining connectivetissue area. Am J Anat 118:785–809
Levick JR (1991) Capillary filtration-absorption balance reconsidered in light of dynamic extravas-cular factors. Exp Physiol 76:825–857
Levick JR (2004) Revision of the starling principle: new views of tissue fluid balance. J Physiol557:704
Levick JR (2010) An introduction to cardiovascular physiology, 5th edn. Hodder Arnold, LondonLudbrook J (1966) The musculovenous pumps of the human lower limb. Am Heart J 71:635–641Lurie F, Kistner RL (2012) The relative position of paired valves at venous junctions suggests their
role in modulating three-dimensional flow pattern in veins. Eur J Vasc Endovasc Surg44:337–340
Lurie F, Kistner RL (2013) On the existence of helical flow in veins of the lower extremities. J VascSurg Venous Lymphat Disord 1:134–138
Lurie F, Kistner RL, Eklof B (2002) The mechanism of venous valve closure in normal physiologicconditions. J Vasc Surg 35:713–717
Lurie F, Kistner RL, Eklof B, Kessler D (2003) Mechanism of venous valve closure and role of thevalve in circulation: a new concept. J Vasc Surg 38:955–961
Malone PC, Agutter PS (2006) The aetiology of deep venous thrombosis. QJM 99:581–593McCaughan JJ Jr, Walsh DB, Edgcomb LP, Garrett HE (1984) In vitro observations of greater
saphenous vein valves during pulsatile and nonpulsatile flow and following lysis. J Vasc Surg1:356–361
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 16 of 19

Mellor RH, Brice G, Stanton AW, French J, Smith A, Jeffery S, Levick JR, Burnand KG, MortimerPS (2007) Mutations in FOXC2 are strongly associated with primary valve failure in veins of thelower limb. Circulation 115:1912–1920
Merryman WD (2010) Mechano-potential etiologies of aortic valve disease. J Biomech 43:87–92Michel CC, Phillips ME (1987) Steady-state fluid filtration at different capillary pressures in
perfused frog mesenteric capillaries. J Physiol 388:421–435Moneta GL, Bedford G, Beach K, Strandness DE (1988) Duplex ultrasound assessment of venous
diameters, peak velocities, and flow patterns. J Vasc Surg 8:286–291Naamani R, Hussain SN, Magder S (1995) The mechanical effects of contractions on blood flow to
the muscle. Eur J Appl Physiol Occup Physiol 71:102–112Neglen P, Raju S (2000) Ambulatory venous pressure revisited. J Vasc Surg 31:1206–1213Pappas PJ, DeFouw DO, Venezio LM, Gorti R, Padberg FT Jr, Silva MB Jr, Goldberg MC, Durán
WN, Hobson RW 2nd (1997) Morphometric assessment of the dermal microcirculation inpatients with chronic venous insufficiency. J Vasc Surg 26:784–795
Pedley TJ (1987) Haemodynamics: how giraffes prevent oedema. Nature 329:13–14Robicsek F, Thubrikar MJ, Fokin A, Tripp HF, Fowler B (1999) Pressure traps in femoro-popliteal
reversed vein grafts. Are valves culprits? J Cardiovasc Surg (Torino) 40:683–689Rushmer RF (1976) Cardiovascular dynamics, 4th edn. Saunders, PhiladelphiaSakurai T, Matsushita M, Nishikimi N, Nimura Y (1997) Hemodynamic assessment of
femoropopliteal venous reflux in patients with primary varicose veins. J Vasc Surg 26:260–264Sansilvestri-Morel P, Rupin A, Badier-Commander C, Kern P, Fabiani JN, Verbeuren TJ, Vanhoutte
PM (2001) Imbalance in the synthesis of collagen type I and collagen type III in smooth musclecells derived from human varicose veins. J Vasc Res 38:560–568
Sansilvestri-Morel P, Rupin A, Jullien ND, Lembrez N, Mestries-Dubois P, Fabiani JN, VerbeurenTJ (2005) Decreased production of collagen type III in cultured smooth muscle cells fromvaricose vein patients is due to a degradation by MMPs: possible implication of MMP-3.J Vasc Res 42:388–398
Saphir O, Lev M (1952) The venous valve in the aged. Am Heart J 44:843–850Scultetus AH, Villavicencio JL, Rich NM (2001) Facts and fiction surrounding the discovery of the
venous valves. J Vasc Surg 33:435–441Sevitt S (1974) The structure and growth of valve-pocket thrombi in femoral veins. J Clin Pathol
27:517–528Shepherd JT (1966) Role of the veins in the circulation. Circulation 33:484–491Sheriff DD (2003) Muscle pump function during locomotion: mechanical coupling of stride
frequency and muscle blood flow. Am J Physiol Heart Circ Physiol 284:H2185–H2191Sheriff DD, Van Bibber R (1998) Flow-generating capability of the isolated skeletal muscle pump.
Am J Physiol 274:H1502–H1508Tan J, Smith A, Abisi S, EasthamD, Burnand KG (2007) Tissue and urinary haemosiderin in chronic
leg ulcers. Eur J Vasc Endovasc Surg 34:355–360Thomson H (1979) The surgical anatomy of the superficial and perforating veins of the lower limb.
Ann R Coll Surg Engl 61:198–205Thubrikar MJ, Robicsek F, Fowler BL (1994) Pressure trap created by vein valve closure and its role
in graft stenosis. J Thorac Cardiovasc Surg 107:707–716Valic Z, Buckwalter JB, Clifford PS (2005) Muscle blood flow response to contraction: influence of
venous pressure. J Appl Physiol (1985) 98:72–76van Bemmelen PS, Bedford G, Beach K, Strandness DE (1989) Quantitative segmental evaluation
of venous valvular reflux with duplex ultrasound scanning. J Vasc Surg 10:425–431
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 17 of 19

van Bemmelen PS, Beach K, Bedford G, Strandness DE Jr (1990) The mechanism of venous valveclosure. Its relationship to the velocity of reverse flow. Arch Surg 125:617–619
Weinbaum S, Zhang X, Han Y, Vink H, Cowin SC (2003) Mechanotransduction and flow across theendothelial glycocalyx. Proc Natl Acad Sci U S A 100:7988–7995
Weinbaum S, Tarbell JM, Damiano ER (2007) The structure and function of the endothelialglycocalyx layer. Annu Rev Biomed Eng 9:121–167
Wilkinson LS, Bunker C, Edwards JC, Scurr JH, Smith PD (1993) Leukocytes: their role in theetiopathogenesis of skin damage in venous disease. J Vasc Surg 17:669–675
Zamboni P, Tognazzo S, Izzo M, Pancaldi F, Scapoli GL, Liboni A, Gemmati D (2005) Hemochro-matosis C282Y genemutation increases the risk of venous leg ulceration. J Vasc Surg 42:309–314
Further ReadingLurie F (2008) The function of venous valves in normal and pathological conditions. Medicographia
30(2):95–99, This article provides a summary of physiology and pathophysiology of venousvalves
Pirner F (1981) On the valves of the perforating veins. In: May R (ed) Perforating veins. Urban &Schwarzenberg, M€unchen, pp 46–48
Stuart WP, Lee AJ, Allan PL, Ruckley CV, Bradbury AW (2001) Most incompetent calf perforatingveins are found in association with superficial venous reflux. J Vasc Surg 34:774–778
Sumner D, van Bemmelen P (1997) Hemodynamics of venous system: calf pump and valvefunction. In: Raju S, Villavincencio L (eds) Surgical management of venous disease. William& Wilkins, Baltimore, pp 16–59, This text provides an excellent overview of venoushemodynamics
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 18 of 19

Index Terms:
Blood dynamics 3Capillary filtration 12Compliance 2Flow modulation 11Interstitial pressure 13Muscle pump 6Perforating veins 11Pressure gradient 4Reflux 7Synechia 5Transmural pressure 2Valve closure 8Valve cycle 8Valve leaflets 8Venous capacitance 2Venous resistance 4Venous return 3Venous tone 3Venous valve 7Venous volume 2Venous wall 14Vortical flow 9
PanVascular MedicineDOI 10.1007/978-3-642-37393-0_159-1# Springer-Verlag Berlin Heidelberg 2014
Page 19 of 19





























