Embed Size (px)
Citation preview

Phytopath. Z., 89, 359—365 (1977)© 1977 Verlag Paul Parey, Berlin und HamburgISSN 0031-9481 / ASTM-Coden: PHY2A3
Division of Fruit and Vegetable Storage, Agricultural Research OrganizationThe Volcani Center, Bet Dagan, Israel
Physiological Response of Avocado Fruit to Infectionby different Fusarium Species
By I
GiORA ZAUBERMAN and MINA SCHIFFMANN-NADEL
With J figuresI
Received January 6, 1977I
Introduction
The avocado (Persea americana Mill.) differs from other fruit in that Itdoes not ripen while still on the tree (BIALE and YOUNG 1962), but ripens andsoftens several days after harvest. At softening, various fungi whidi cause rotmay develop on the fruit (HORNE 1934), among them some species of Fusa-rium (JoFFE and SCHIFFMANN-NADEL 1967). Previous observations showed thatavocado fruit naturally infected by Fusarium solani (Mart.) App. and Wr.softened after harvest earlier than uninfected fruit {ZAUBERMAN and SCHIFF-MANN-NADEL 1974). Softening of the infected fruit occurred during the in-cubation period of the fungus, before the appearance of the first sign of rot;softening did not progress from the infection point, but occurred in the wholefruit at once. With the softening of the fruit the rot started at the infectionpoint as a brown spot, whidi grew larger and became covered with whitemycelium. Naturally- and artificially-inoculated fruit behaved similarly(ZAUBERMAN and SCHIFFMANN-NADEL 1974).
In this previous study a question arose whether the nature of the earlysoftening of the avocado fruit infected by F. solani was similar to that insound fruit, and whether the earlier softening was an acceleration of theripening processes that occur during fruit softening.
Contrihution from the Agricultural Researdi Organization, The Volcani Center, BetDagan, Israel. 1976 Series, No. 240-E.

360 ZAUBERMAN and SCHIFFMANN-NADEL
It was found that basic physiological processes related to softening andripening, such as an increasing rate of respiration, ethylene evolution, pecto-lytic enzyme activity and dianges in pectolytic substances, were found to besimilar in infected and uninfected fruit. However, these processes occurredearlier in the infected fruit, resulting in earlier softening and ripening (ZAU-BERMAN and SCHIFFMANN-NADEL 1974).
Further study of avocado fruit infected by species of the genus Fusariumpathogenic to avocado showed that while some of the species of this genusaccelerated softening of the fruit, others did not. On this basis, we dividedFusarmm species isolated from avocado into two groups; A-fungi causingearlier softening of the fruit {F. solani and F. sambucinum), and B-fungi thatdid not accelerate fruu softening {F. avenaceum, F. eqttiseti, F. moniliformeand F. semitectum).
The question then was raised whether, in the same genus of Fusarium,there are some species that cause an earlier softening of the fruits and otherspecies that did not. For this purpose, in the present study, we compared somemetabolic functions during softening in fruits infected by fungi of both groupsand unmfected fruits. The respiration pattern, and the polygalacturonase (PG)and pectin methylesterase (PME) enzyme activities, were chosen as goodmdicators of physiological changes occurring during avocado fruit softeningand ripening (ZAUBERMANN and SCHIFFMANN-NADEL 1974).
Materials and Methods
Avocado {cultivar 'Fuerte') fruit was inoculated by placing 0.1 ml of a suspension ofFusarium spores (lO** sporcs/ml) into a 0.5-cm-deep wound made in the fruit. The inoculationsite was covered with cellotapc and the fruit was kept at 14' and 20 °C and 90 9r relativehumidity until examination. Sound and wounded but not inoculated fruit served as controls.The experiments were conducted three or four times per year over a period of 3 years, withthree to five replicates in each group.
Respiration determinations were made on individual fruits placed in 2-liter j;lasscontainers, and the amount of COg evolved was determined by gas chromatography. Activitiesof PME and PG were determined in rotten and sound fruits according to the methoddescribed previously {ZAUBERMAN and SCHIFFMANN-NADEL 1971).
Results
The physiological behavior of the fruits infected by fungi of groups Aand B, and of uninfected fruit, was somewhat similar; the climacteric patternand respiration values at the climacteric peak were similar in all fruits. How-ever, in fruit infected by the fungi of group A, the climacteric peak appearedearlier than in the other two categories; for example, 4 days after inoculationas compared with 7 days (Fig. 1).
Enzyme activities in general roughly followed the respiration pattern.No PG activity was detected in firm fruit immediately after harvest; PG
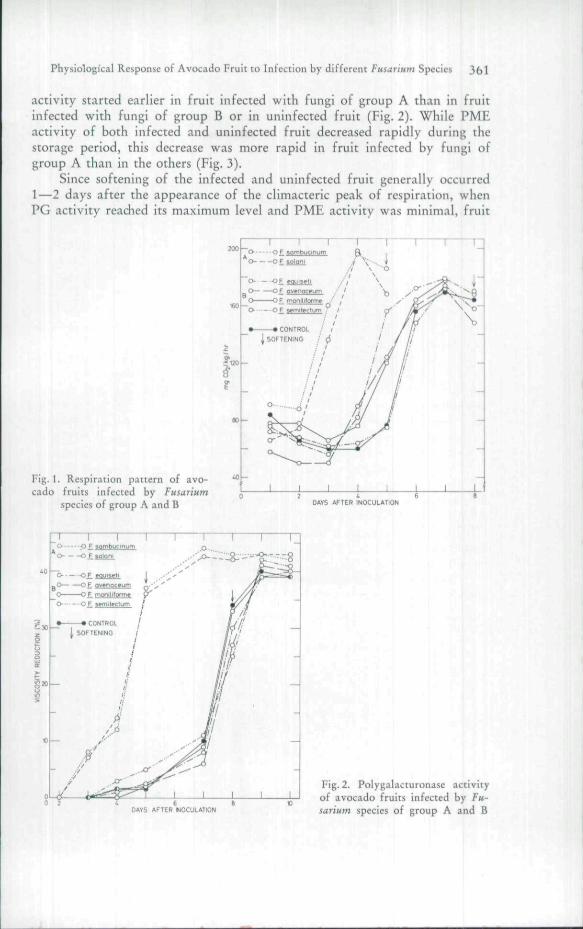
Physiological Response of Avocado Fruit to Infection by different Fusarmm Species 36l
activity started earlier in fruit infected with fungi of group A than in fruitinfected with fungi of group B or in uninfected fruit (Fig. 2). While PMEactivity of both infected and uninfected fruit decreased rapidly during thestorage period, this decrease was more rapid in fruit infected by fungi ofgroup A than in the others (Fig. 3).
Since softening of the infected and uninfected fruit generally occurred1—2 days after the appearance of the climacteric peak of respiration, whenPG activity reached its maximum level and PME activity was minimal, fruit
Fig. 1. Respiration pattern of avo-cado fruits infected by Fusar'tHm
species of group A and B DAYS AFTER INOCULATION
DAYS AFTER HOCULATION
Fig. 2. Polygalacturonase activityof avocado fruits infected by Fu-sarmm species of group A and B
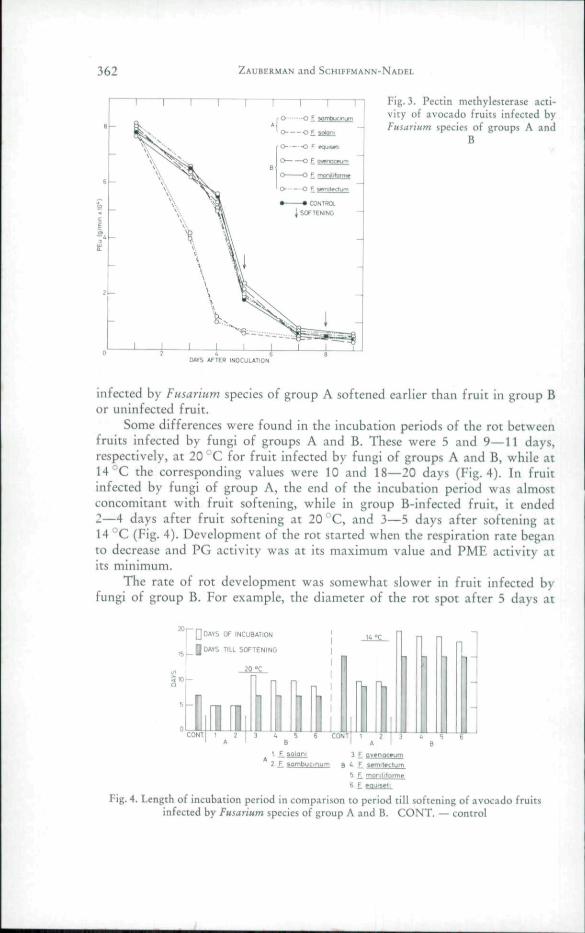
362 ZAUBERMAN and SCHIFFMANN-NADEL
CONTWXSOF TENINC —
Fig, 3. Pectin methylesterase acti-vity of avocado fruits infected byFusarium species of groups A and
B
DAV5 AFTER INOCJLAIION
infected by Fusarium species of group A softened earlier than fruit in group Bor uninfected fruit.
Some differences were found in the incubation periods of the rot betweenfruits infected by fungi of groups A and B. These were 5 and 9—11 days,respectively, at 20 °C for fruit infected by fungi of groups A and B, while at14'^C the corresponding values were 10 and 18—20 days (Fig. 4). In fruitinfected by fungi of group A, the end of the incubation period was almostconcomitant with fruit softening, while in group B-infected fruit, it ended2—4 days after fruit softening at 20 C, and 3—5 days after softening at14 °C (Fig. 4). Development of the rot started when the respiration rate beganto decrease and PG activity was at its maximum value and PME activity .itits minimum.
The rate of rot development was somewhat slower in fruit infected byiungi of group B. For example, the diameter of the rot spot after 5 days at
[ ] DAYS OF INCUBATION
_ JDAVS TILL SOFTENING
iO'C
Fig. 4. Length of incubation period in comparison to period rill softening of avocado fruitsinfected by Fusarium species of group A and B, CONT. — control

Physiological Response of Avocado Fruit to Infection by different Fusarium Species 363
20*^0 was 24—31mm in group-B fungi-infected fruit as compared with 33—34 mmm group A-infected (Fig. 5). Similar resultswere obtained at 14 °C.
Fig. 5. Diameter of rot, after 5 days at 20 '̂ 'C, frominoculation of avocado fruits infected by Fusarium
species of groups A and B
5•
O 20
1 F sambucinum2 F solon
B3. F. eouiseti4. F_ monilitorme5. F_ avenaceum6. F. semilectum
Discussion
Previously (ZAUBERMAN and SCHIFFMANN-NADEL 1974), we found that.Tccelerated softening of fruit infected by F. solani was related to acceleratedphysiological processes connected with fruit ripening, sudi as respiration,ethylene evolution, changes in pectic substances and pectolytic enzymes activ-ity. In this extension of the study, we found that only certain Fusaria speciescause this acceleration of fruit softening and ripening. Thus, Fusaritim samhu-cinum had an effect on fruit softening similar to that of F. solani, while otherspecies sudi as F. avenaceum, F. equiseti, F. moniliforme, and F. semitectnmdid not.
The results of this and the previous study (ZAUBERMAN and SCHIFFMANN-NADEL 1974) may show that whereas physiological processes related to fruitripening seem to be basically similar in fruit infected by fungi of both groupsA and B and uninfected fruit, in fruit infected by fungi of group A, theripening processes are accelerated whereas in fruit infected by fungi of groupB the fungus infection has no effect on the ripening processes, which aresimilar to those in non-infected fruit.
Information on the effect of fungi on changes in metabolic processes isscare. An increased rate of respiration was found in some fruits infected bydifferent fungi; in tissue slices of peadies inoculated with Monilinia fructicola(HALL 1967); in lemons inoculated with Penicillium digitatum (EARS 1955)or infected by Phytophthora citrophthora (COHEN and SCHIFFMANN-NADEL1972); in oranges, due to Diplodia natalensis (ZAUBERMAN and BARKAI-GOLAN1975); and in citrus fruits due to different fungi pathogenic to them (SCHIFF-MANN-NADEL 1974, VINES, EDWARDS and GRIERSON 1965). In avocado, how-ever, we did not find any sudi increase in the rate of respiration, increasedmetabolism being expressed by an earlier peak in the climacteric respirationpattern. Some changes in activity of pectolytic enzymes caused by fungi infruit have been cited in literature: a rise in PG activity was found in lemonpeel infected by P. digitatum (BARASH and ANGEL 1970) and in avocado in-fected by Colletotrichum gloeosportoides (BARASH and KHAZZAN 1970).

364 ZAUBERMAN and SCHJFFMANN-NADEL
Our observations showed that fungi pathogenic to avocado fruit did notdevelop in firm fruits, but started concomitantly with or after fruit softening;consequently, there may be a relationship between the length of the incubationperiod and the length of time until softening. The faster softening of the fruitdue to group A fungi may reduce the time for the appearance of rots asobserved; thus, there may be a relationship between the earlier softening ofthe fruit and the shorter incubation period of fruit infected by fungi ofgroup A. On the other hand, infection by fungi of group B did not cause anacceleration of fruit softening and consequently the incubation period in suchfruit was longer.
The effect of group A fungi on fruit softening and ripening was similarto that caused by ethylene treatment of uninfected avocado fruits. It may bethat infection by the fungi of group A that triggered an earlier ethylene evolu-tion as shown in our previous study (ZAUBERMAN and SCHIFFMANN-NADEL1974), triggers the acceleration of ripening processes.
This study showed different host-parasite relations in fruits infected bythe genus Fusarium. It is thus evident that various fungi, even of the samegenus — in our case Fusarium — differ in their physiological behavior andmay or may not accelerate ripening processes. Those fungi that cause accelera-tion of ripening also have a shorter incubation period.
Summary
Infection of avocado fruit by F. solani and F. sambucinum (group A)caused an accelerated softening of fruit which was accompanied by an earlierrise in climacteric respiration and in enzymatic activity of polygalacturonase(PG) and pectin methylesterase (PME). On the other hand, infection by F.avenaceum, F. equiseti, F. moniliforme and F. semitectum (group B) did notcause this phenomenon and the physiological processes occurred as in un-infected fruits. The incubation period of fungi in group A was shorter thanin group B; development of the rot started when the respiration rate beganto decrease and PG activity was at its maximum and PME activity at itsminimum.
Zusammenfassung
Physiologisdie Reaktion von Avocado-Fruditenauf Infektion durdi versdiiedene Fusarium-hrtcn
Die Infektion von Avocado-Friichten durdi Fusarium solani und F. sam-bucinum (Gruppe A) verursachte ein beschleunigtes Weichwerden der Frijchte,das von einem fruheren Eintreten in den kritisdien Bereidi der Atmung undvon einer enzymatisdien Aktivitat von Polygalacturonase (PG) und Pektin-methylesterase (PME) begleitet war. F. avenaceum, F. equiseti, F. moniliformeund F. semitectum (Gruppe B) dagegen verursachten dieses Phanomen nidit,sondern die physiologischen Prozesse liefen hier wie bei nicht infizierten

Physiological Response of Avocado Fruit to Infection by different Fusarium Species 365
Priiditen ab. Die Inkubationszeit der Pilze in Gruppe A war kurzer als derin Gruppe B. Die Faule setzte ein, wenn die Atmung abzunehmen bcgann unddie PG-Aktivitat ihr Maximum, die PME-Aktivitat ihr Minimum erreiclithatten.
Literature
BARASH, I., and E. ANGEL, 1970: Isolation and properties of an exopolygalacturonase producedby Penicillium digitatum during infection of lemon fruits. Israel J. Bot. 19, 599—608.
, and S. KHAZZAN, 1970: The relationship and properties of pectic glycosidases producedby host and pathogen during Anthracnose disease of avocado. Phytodiemistry 9,1187-1197.
BiALE, J. B., and R. E. YOUNG, 1962: The biodiemistry of fruit maturation. Endeavour 21,164—174.
COHEN, E., and M. SCHIFFMANN-NADEL, 1972: Respiration pattern of lemon fruit infectedwith Phytophthora citrophthora. Phytopathology 62, 932—933.
KAKS, I. L., 1955: Effect of biphenyl on respiration of oranges and lemons. Proc. Amer. Soc.hort. Sci. 66, 135—140.
HALL, R., 1967: Effect of Monilinia frHCticola on oxygen uptake of peadi fruits. Phytopath.2.58, 131 — 136.
HORNE, W. T., 1934: Avocado diseases in California. Calif. Agric. Exp. Sta. Bull. 585,58—72.
JoFFE, A. Z., et M. SCHIFFMANN-NADEL, 1967: Les Fusarium isoles a partir d'avocats etd'avocatiers d'Israel. Fruits 22, 97—100,
SCHIFFMANN-NAUEL, M., 1974: Relation between fungal attack and postharvest fruit matura-tion. Coll. Intern. C.N.R.S. Facteurs et regulation de la maturation des fruits, 238,139—145.
VINES, H . M., G. J. ED-ffARDS, and W. GRIERSON, 1965: Citrus fruit respiration. Proc. Fla.Sta. hort. Soc. 78, 198-202.
/-AuittRMAN, G., and R. BARKAI-GOLAN, 1975: Changes in respiration and ethylene evolutioninduced by Diplodia nataiemis in orange fruit. Phytopathology 65, 216—217.
, and M. ScHiFFMANN-NADEt, 1971: Pectin methylesterase and polygalacturonase inavocado fruit at various stages of development. Plant Physiol. 49, 864—865.
— —, and , 1974: Changes in the ripening process of avocado fruit infected by Fusariitmsolani. Phytopathology 64, 188-190.
Authors' address: Division of Fruit and Vegetable Storage, Agricultural ReseardiOrganization, The Volcani Center, P.O.B. 6, Bet Dagan (Israel).






























