Embed Size (px)
Citation preview

REVIEW
Physiological and molecular changes in plants grownat low temperatures
Andreas Theocharis • Christophe Clement •
Essaıd Ait Barka
Received: 30 November 2011 / Accepted: 13 March 2012 / Published online: 20 April 2012
� Springer-Verlag 2012
Abstract Apart from water availability, low temperature
is the most important environmental factor limiting the
productivity and geographical distribution of plants across
the world. To cope with cold stress, plant species have
evolved several physiological and molecular adaptations to
maximize cold tolerance by adjusting their metabolism.
The regulation of some gene products represents an addi-
tional mechanism of cold tolerance. A consequence of
these mechanisms is that plants are able to survive expo-
sure to low temperature via a process known as cold
acclimation. In this review, we briefly summarize recent
progress in research and hypotheses on how sensitive
plants perceive cold. We also explore how this perception
is translated into changes within plants following exposure
to low temperatures. Particular emphasis is placed on
physiological parameters as well as transcriptional, post-
transcriptional and post-translational regulation of cold-
induced gene products that occur after exposure to low
temperatures, leading to cold acclimation.
Keywords C-repeat binding factor � Low temperatures �Plant acclimation � Signal perception
Abbreviations
ABA Abscisic acid
CBF C-repeat binding factor
COR Cold-responsive genes
CRT C-repeat elements
DRE Dehydration-responsive elements
DREB Dehydration-responsive element binding
ICE Inducer of CBF expression
LT Low temperature
Introduction
Among various environmental stresses, low temperature
(LT) is one of the most important factors limiting the
productivity and distribution of plants. Low temperatures,
defined as low but not freezing temperatures (0–15 �C), are
common in nature and can damage many plant species. In
order to cope with such conditions, several plant species
have the ability to increase their degree of freezing toler-
ance in response to low, non-freezing temperatures, a
phenomenon known as cold acclimation. It is well estab-
lished that some of the molecular and physiological chan-
ges that occur during cold acclimation are important for
plant cold tolerance (Hsieh et al. 2004; Zhu et al. 2007).
Accordingly, it has been concluded that cold tolerance that
develops in initially insensitive plants is not entirely con-
stitutive and at least some of it is developed during expo-
sure to low temperatures.
This review addresses plant adaptive responses to cold
stress, with a special emphasis on understanding (i) the
key elements involved in cold signal perception and
transduction, (ii) the major physiological and biochemi-
cal changes that occur following cold exposure, (iii)
cold-inducible gene products that help the plant to
accomplish a synergistic response to cold, and finally
(iv) products that may play major roles in cold accli-
mation and tolerance.
A. Theocharis � C. Clement � E. A. Barka (&)
Laboratoire de Stress, Defense et Reproduction des Plantes,
URVVC, UPRES EA 2069, Universite de Reims
Champagne-Ardenne, BP 1039, 51687 Reims Cedex 2, France
e-mail: [email protected]
123
Planta (2012) 235:1091–1105
DOI 10.1007/s00425-012-1641-y

How do plants perceive cold?
As a result of exposure to low temperatures, many
physiological and biochemical cell functions have been
correlated with visible symptoms (wilting, chlorosis, or
necrosis) (Ruelland and Zachowski 2010). Often, these
adverse effects are accompanied by changes in cell mem-
brane structure and lipid composition (Uemura and
Steponkus 1999; Matteucci et al. 2011), cellular leakage of
electrolytes and amino acids, a diversion of electron flow to
alternate pathways (Seo et al. 2010), alterations in proto-
plasmic streaming and redistribution of intracellular cal-
cium ions (Knight et al. 1998) (Fig. 1). They also involve
changes in protein content and enzyme activities (Ruelland
and Zachowski 2010) as well as ultrastructural changes in a
wide range of cell components, including plastids, thyla-
koid membranes and the phosphorylation of thylakoid
proteins, and mitochondria (Zhang et al. 2011). Brief
exposures to low temperatures may only cause transitory
changes, and plants generally survive. However, prolonged
exposure to stress causes plant necrosis or death.
To overcome stresses generated by exposure to low
non-freezing temperatures, plants can trigger a cascade of
events that cause changes in gene expression and thus
induce biochemical and physiological modifications that
enhance their tolerance (Smallwood and Bowles 2002; Zhu
et al. 2007). This phenomenon is known as chilling or cold
acclimation.
Mechanisms of acclimation to low non-freezing
temperatures
The primary mechanisms involved in cold acclimation are
related to a number of processes discussed below. These
include molecular and physiological modifications occur-
ring in plant membranes, the accumulation of cytosolic
Ca2?, increased levels of ROS and the activation of ROS
scavenger systems, changes in the expression of cold-
related genes and transcription factors, alterations in
protein and sugar synthesis, proline accumulation, and
biochemical changes that affect photosynthesis (Fig. 2).
Modifications to plant cell membranes
Membranes are a primary site of cold-induced injury
(Fig. 1). Several studies have demonstrated that membrane
rigidification, coupled with cytoskeletal rearrangements,
calcium influxes, and the activation of MAPK cascades,
triggers LT responses (Uemura and Steponkus 1999; Orvar
et al. 2000; Xin and Browse 2000; Sangwan et al. 2002).
The lipid composition of the plasma membrane and chlo-
roplast envelopes in acclimated plants changes such that
the threshold temperature for membrane damage is lowered
relative to that for non-acclimated plants (Uemura and
Steponkus 1999). This is achieved by increasing the cold-
adapted membranes’ unsaturated fatty acid content, which
makes them more fluid (Vogg et al. 1998). The process of
Transition phase
Liquid-Crystalline Solid gel
‘’FLUID’’ ‘’SOLID’’
Solute leakage and disrupted ion balance
Injury and death of cells and tissues
Enhanced permeability
Increased activation of energy-bound enzymes
Cessation of protoplasmic streaming
Imbalance in metabolism
Accumulation of toxic metabolites
Reduced ATP supply
Return to normal metabolism
Brief exposure and return to 20 C
Prolonged exposure
Survival
Chilling stress
Cell membranes
°
Fig. 1 A model to explain
symptoms of chilling injury in
chilling-sensitive plants.
Membranes are the primary site
of cold-induced injury, leading
to a cascade of cellular
processes with adverse effects
on the plant. When exposure to
low temperature is brief, the
effects may be transitory and
plants survive. However, the
plant will exhibit necrosis or die
if exposure is maintained
(Lyons 1973; Raison and Lyons
1986)
1092 Planta (2012) 235:1091–1105
123

cold acclimation promotes the stabilization of membranes,
which prevents damage leading to cell death. The accli-
mation process also activates mechanisms that protect
membrane fluidity by ensuring the optimal activity of
associated enzymes (Matteucci et al. 2011).
Photosynthesis and photosynthesis-related pigments
At the physiological level, photosynthesis is strongly affec-
ted by exposure to cold. The cessation of growth resulting
from cold stress reduces the capacity for energy utilization,
causing feedback inhibition of photosynthesis (Ruelland and
Zachowski 2010). In cold-acclimated winter annuals, pho-
tosynthetic activity is maintained by increases in the abun-
dance and activity of several Calvin cycle enzymes (Goulas
et al. 2006). This recovery is associated with elevated levels
of thylakoid plastoquinone A and a concomitant rise in the
apparent size of the intersystem electron donor pool to PSI
(Baena-Gonzalez et al. 2001). Consequently, non-photo-
chemical quenching increases in cold-stressed leaves in
parallel with increased zeaxanthin levels to compensate for
the reduced electron consumption by photosynthesis. Zea-
xanthin protects the PSII reaction centres from over-excita-
tion (Krol et al. 1999). Nevertheless, Ruelland and
Zachowski (2010) reported that energy dissipation via non-
photochemical quenching (NPQ) and electron transport was
not only enhanced following cold acclimation but also con-
tributed to protection from oxidative damage.
Xanthophylls
Although they are not considered photosynthetic pigments
per se, the xanthophylls (notably, violaxanthin, anthera-
xanthin, and zeaxanthin) help in protecting the photosystems
and their abundance increases at low temperatures (Ivanov
et al. 2006). Xanthophylls have structural roles and act as
natural antioxidants, quenching triplet Chl and singlet oxy-
gen, which are potentially harmful to the chloroplast
(Passarini et al. 2009; Han et al. 2010). It has also been
postulated that unbound zeaxanthin and other carotenoids
may also stabilize thylakoid membranes against putative
peroxidative damage and heat stress (Laugier et al. 2010).
Flavonoids
These accumulate in leaves and stems in response to low
temperatures. They are synthesized via the phenylpropanoid
pathway, which is controlled by key enzymes, including
phenylalanine ammonia-lyase and chalcone synthase
(Sharma et al. 2007). Recently, it has been reported that cold
stress induces transcriptomic modifications that increase
flavonoid biosynthesis, including reactions involved in
anthocyanin biosynthesis and the metabolic pathways that
supply it (Crifo et al. 2011).
Calcium and cold temperatures
Calcium acts as a mediator of stimulus–response coupling
in the regulation of plant growth, development, and
responses to environmental stimuli (Sanders et al. 2002; Du
and Poovaiah 2005). Cold stress-induced rigidification of
plasma membrane microdomains can cause actin cyto-
skeletal rearrangement. This may be followed by the acti-
vation of Ca2? channels and increased cytosolic Ca2?
levels (Fig. 3), which may be involved in the cold accli-
mation process (Sangwan et al. 2001; Catala et al. 2003).
The Ca2? released from internal cellular reserves, mediated
by inositol triphosphate, is upstream of the expression of
Accumulation of [Ca2+cyt]
Accumulation of ROS and activation of scavenge system
Modification in plant membranes
Photosynthetic acclimation
Accumulation of cryoprotectantssugars, proline, ...
Changes in gene expression and protein synthesis
Change in lipid composition
Increase in desaturated fatty acids
Increased fluidity of membranes
Reduction of lower threshold temperature in acclimated plants
ColdAcclimation
Fig. 2 Different cellular
processes induced as a
consequence of plant
acclimation to cold
Planta (2012) 235:1091–1105 1093
123

CBFs (C-repeat binding factors) and COR (cold respon-
sive) genes in the cold-signalling pathway(s) (Chinnusamy
et al. 2007, 2010). Recently, Doherty et al. (2009) provided
more evidence for a link between calcium signalling and
cold induction of the CBF pathway, showing that cal-
modulin binding transcription activator (CAMTA) factors
bind to a regulatory element in the CBF2 gene promoter.
As the CAMTA proteins are calmodulin binding tran-
scription factors, they may act directly in the transduction
of LT-induced cytosolic calcium signals into downstream
regulation of gene expression (Doherty et al. 2009). Sim-
ilarly, CRLK1, a novel calcium/calmodulin-regulated
receptor-like kinase, was reported to be crucial for cold
tolerance in plants (Yang et al. 2010).
Role of reactive oxygen species in acclimation
to low temperatures
The role of ROS in abiotic stress management has become
a subject of considerable research interest, particularly
since ROS have been reported to be involved in processes
leading to plant stress acclimation (Suzuki et al. 2011).
This finding indicates that ROS are not simply toxic
by-products of metabolism, but act as signalling molecules
that modulate the expression of various genes, including
those encoding antioxidant enzymes and modulators of
H2O2 production (Neill et al. 2002; Gechev et al. 2003;
Suzuki et al. 2011) (Fig. 3). In addition, LT stress has been
reported to cause significant increases in the levels of the
soluble non-enzymatic antioxidants ascorbate and gluta-
thione, as well as the activity of the main NADPH-gener-
ating dehydrogenases (Airaki et al. 2011).
Cold-mediated transcription regulation
Plant acclimation to LT depends on changes in the
expression of specific genes encoding products that confer
increased cold tolerance (Gilmour et al. 1998; Doherty
et al. 2009) (Fig. 4). The process involves modifying pre-
existing proteins and up- or down-regulating gene expres-
sion and protein synthesis. Several studies have suggested
that activity of cold/chilling-induced genes may facilitate
the metabolic changes that confer LT tolerance (Gilmour
et al. 1998; Doherty et al. 2009). They may also be
involved in the signal transduction of the stress-response
(Ingram and Bartels 1996; Thomashow 2010). HOS15, one
of 237 predicted WD40-repeat proteins in Arabidopsis,
functions as a repressor that modifies chromatin and
thereby controls the expression of genes involved in cold
tolerance (Zhu et al. 2008). In addition, HOS1 is a RING-
type ubiquitin E3 ligase that mediates the ubiquitination
and proteosomal degradation of ICE1 and thus negatively
regulates CBF regulons. The constitutive HOS9 and
HOS10 regulons have a role in the negative regulation of
CBF-target genes (Fig. 4).
The CRT/DREB1 regulatory pathway
A wide variety of COR genes have been isolated from cold-
acclimated plants (Svensson et al. 2006). The cloned
genes’ products can be classified as (i) proteins that protect
cells against environmental cold/chilling stress, and (ii)
proteins that regulate gene expression during the adaptation
response (Fowler and Thomashow 2002). Further classifi-
cation divides the gene products into (i) mediators of
Inactivation of enzymes
Lipid peroxidation
Protein degradation
Damage to DNA
PS I, II
Plasma membranes
Cessation of cytoplasm streaming
Inhibition of ethylene biosynthesis
Activation or repression of phosphatase activity
Changes in cytoplasm viscosity
Inhibition of photosynthesis
Induction of cold related genes (COR )
Elevation of proline andtotal soluble sugars contents
Chilling stress
ROS
[Ca2+cyt]
Fig. 3 Ca2? and reactive
oxygen species (ROS) responses
to chilling in sensitive plants.
ROS are not simply toxic
by-products of metabolism; they
also act as signalling molecules
by modulating the expression of
various genes, including those
encoding antioxidant enzymes
and modulators of H2O2
production. Changes to the
plasma membrane can cause
actin cytoskeletal
rearrangements that may be
followed by the activation of
Ca2? channels and increased
cytosolic Ca2? concentrations,
triggering a series of reactions
such as the expression of
cold-regulated genes
1094 Planta (2012) 235:1091–1105
123

biochemical and physiological changes required for growth
and development at low temperatures and (ii) gene prod-
ucts with a direct role in chilling and freezing tolerance
(Thomashow 2010). A transcriptome analysis of resistant
Arabidopsis treated at 13 �C indicated that 20 % of about
8,000 genes were affected by treatment, particularly those
involved in protein biosynthesis (Provart et al. 2003;
Chinnusamy et al. 2010). The mRNA profiles for the
chilling-lethal mutants were very similar and included
extensive chilling-induced and mutant-specific alterations
in gene expression. The expression pattern of the mutants
upon chilling suggests that the normal function of the
mutated loci is to prevent a wide-ranging damaging effect
of LT on transcriptional regulation (Provart et al. 2003).
Role of CBF transcription factors in cold acclimation
The signal transduction pathways that control COR
expression incorporate a regulatory network (Fig. 4) in
which a few regulatory genes control those involved in the
cold response (Fowler and Thomashow 2002; Yamaguchi-
Shinozaki and Shinozaki 2006; Chinnusamy et al. 2010;
Thomashow 2010). Attempts to isolate the regulatory ele-
ments responsible for the initiation of the COR gene tran-
scription under LT have primarily focused on Arabidopsis.
The ability to express all of the COR genes in concert at
warm temperatures was described following the discovery
of the CBF family of transcriptional activators (Gilmour
et al. 2004; Skinner et al. 2005; Chinnusamy et al. 2010),
which are also known as DREs (drought responsive ele-
ments) or LTREs (low temperature responsive elements)
(Shinwari et al. 1998; Shinozaki and Yamaguchi-Shinozaki
2000; Yamaguchi-Shinozaki and Shinozaki 2006). The
CBF1, CBF2, and CBF3 genes follow one-another in
sequence on Arabidopsis chromosome 4 (Shinwari et al.
1998; Gilmour et al. 2004). DRE/LTREs stimulate gene
expression in response to cold, high salinity and drought,
but not in response to exogenously applied abscisic acid
(Shinozaki and Yamaguchi-Shinozaki 2000). Using the
yeast one-hybrid system, the DRE/CRT elements have
been used as bait to isolate DRE/CRT-binding proteins.
The five different DRE binding proteins isolated to date
have been classified into two groups, DREB1 and DREB2
(Liu et al. 1998), which bind specifically to the DRE/CRT
elements and transcriptionally activate the expression of
COR genes. The promoters of CBF/DREB-regulated COR
genes contain a cold- and dehydration-responsive DNA
regulatory element known as CBF/DRE (Shinozaki and
Yamaguchi-Shinozaki 2000).
Overexpression of CBF1 in Arabidopsis has been shown
to activate the expression of the entire battery of known
CBF/DREB-regulated COR genes and to enhance whole
plant freezing survival without a low temperature stimulus
(Jaglo-Ottosen et al. 1998). In Arabidopsis, overexpression
of CBF1 and CBF3 activates COR gene expression and
enhances freezing tolerance (Liu et al. 1998; Maruyama
et al. 2004). Additionally, when overexpressed in trans-
genic Arabidopsis plants, the homolog of the CBF/DREB1
protein CBF4 activates a C-repeat/dehydration-responsive
element containing downstream genes that are involved in
cold acclimation (Haake et al. 2002).
The three CBF genes are cold-induced. Indeed, CBF
transcript levels start to increase within 15 min of exposing
plants to low temperatures, and transcripts from the tar-
geted CBF/DRE-regulated COR genes start to accumulate
within approximately 2 h (Mantyla et al. 1995). The pre-
cise mechanism whereby the CBF genes are activated by
LT does not involve autoregulation (Gilmour et al. 1998,
2004) but is controlled by a set of redundant interacting
transcription factors (Vogel et al. 2005; Chinnusamy et al.
2007, 2010).
Many of these genes are also induced by abscisic acid
(Knight et al. 2004) or by dehydration (Shinozaki and
Yamaguchi-Shinozaki 2000), which is consistent with the
fact that both of these factors can increase freezing toler-
ance in transgenic plants (Jaglo-Ottosen et al. 1998; Liu
et al. 1998). However, the existence of CBF-parallel
pathways involved in cold acclimation has been supported
by transcription profiling of plants overexpressing the three
members of the CBF family (Fowler and Thomashow
2002). On the other hand, the Arabidopsis mutant eskimo1
displays freezing tolerance in the absence of cold treat-
ment, without changes in expression of the components of
the CBF pathway, but with a high level of proline accu-
mulation. This suggests that distinct signalling pathways
activate different aspects of cold acclimation. The activa-
tion of one pathway can result in considerable freezing
tolerance without activation of other pathways (Xin and
Browse 2000).
Subsequent to their discovery in Arabidopsis, many
CBF homologues have been found in both monocotyle-
donous and dicotyledonous species capable of cold accli-
mation, but they are also found in species that are not
(Ruelland et al. 2009). For instance, grapevines have five
CBF/DREB1-like genes (CBFs), CBF1 to CBF4 (Xiao
et al. 2006, 2008) and one undefined CBF-like transcription
factor, CBFL (XM_002270601). CBF expression was
reported to be induced within a few hours of exposure to
low temperatures, particularly CBF4 (Xiao et al. 2008).
More recently, Takuhara et al. (2011) have demonstrated
that LT enhanced the expression of VvCBF2, VvCBF4, and
VvCBFL within 3 h, but not VvCBF1 or VvCBF3 expres-
sion. CBF1, 2 and 3 transcripts also accumulated in
response to drought and treatment with exogenous abscisic
acid (ABA), indicating that grapevines contain unique CBF
genes. The expression of the endogenous Vitis CBF4 genes
Planta (2012) 235:1091–1105 1095
123
was low at ambient temperature but increased on exposure
to LT (4 �C). This expression was maintained for several
days, which is uncommon for CBF genes. Further, in
contrast to the previously described Vitis CBF1-3, vCBF4
was expressed in both young and mature tissue. Altogether,
these results suggest that CBF4 represents a second type of
CBF in grape that might be more important for the over-
wintering of grapevine plants.
This raises the question as to whether the CBF tran-
scription factors are limited to activating the expression
of COR genes encoding cryoprotective polypeptides, or
alternatively have a role in activating multiple components
of the cold acclimation response. When both CBF1 and
CBF3 are shut down, the cold induction of all CBF targets
is lower, suggesting that a basal level of CBF proteins
would be needed for the induction of all CBF targets and
for the activation of cold acclimation in Arabidopsis. This
level would be reached only when the CBF1, CBF2, and
CBF3 genes are properly induced in a co-ordinated fashion
(Novillo et al. 2007). On the other hand, overexpression of
CBF3 in Arabidopsis results in multiple biochemical
changes that ultimately increase the concentration of
LOW TEMPERATURES
UNKNOWN SENSORS
Ca2+ influx
ICE1
MYB15
MYBRS
Kinase/Phosphatase
COLD ACCLIMATION/TOLERANCE
ICE1box
ICE1 like
ABRE
METABOLISM
ROS
PROTECTIVE PROTEINS and METABOLITES
Proteolysis
Unknown cis-elements
MYB/MADS/NAC/others
HOS1
HOS1
STZZAT12
LOS2
RAV1
Target
HOS9HOS10
Rd29aCor15a
ZAT10
EP2
RAP 2.6
SFR6
Rd29aRd29b
Rd22AtADH1
ICE1
CBF2CBF3 CBF1
ICE1
COR/KIN/RD/LT? MYCR/MYBR
CBF
U
P
S
CRT/DRE ?
1096 Planta (2012) 235:1091–1105
123

proline and total soluble sugars, including sucrose, raffi-
nose, glucose and fructose (Gilmour et al. 2000). Never-
theless, transcriptome analysis of transgenic Arabidopsis
overexpressing CBF revealed that only about 12 % of the
cold-responsive genes are components of the CBF regulon
(Fowler and Thomashow 2002), suggesting that other
transcriptional activators/repressors also play a significant
role in cold acclimation.
Inducer of CBF expression (ICE), a regulator
of cold acclimation
Since CBF genes are cold-induced, it may be that an
upstream transcription factor present in the cell at normal
growth temperatures is activated by cold stress and, in turn,
induces the expression of CBFs. A constitutive transcrip-
tion factor, inducer of CBF expression 1 (ICE1), which
acts upstream of the CBFs in the cold-response pathway,
has been identified (Chinnusamy et al. 2003). ICE1 binds
to the CBF3 promoter and may activate CBF3 expression
upon cold treatment (Figs. 3, 4). The dominant ice1
mutation blocks the cold induction of CBF3, but not CBF1
or CBF2, and decreases the expression of many CBF-target
genes (Chinnusamy et al. 2003, 2004). The serine 403
residue of ICE1 is involved in regulating the transactiva-
tion and stability of the ICE1 protein. Interestingly, the
substitution of serine 403 by alanine was reported to
enhance the transactivational activity of ICE1 in Arabid-
opsis protoplasts (Miura et al. 2011). These authors also
reported that overexpression of ICE1(S403A) conferred
more freezing tolerance than ICE1(WT) in Arabidopsis,
and the expression of cold-regulated genes such as CBF3/
DREB1A, COR47 and KIN1 (cold-induced gene) was
enhanced in plants overexpressing ICE1(S403A). Whereas
ICE1 primarily affects CBF3/DREB1A expression
(Chinnusamy et al. 2003), the protein encoded by ICE2 (a
homolog of ICE1; At1g12860) primarily influences the
expression of CBF1/DREB1B but has little effect on CBF3/
DREB1A (Fursova et al. 2009). Therefore, ICE1 and ICE2
play pivotal roles in the transcriptional regulation of the
CBF/DREB1 genes (Miura et al. 2011).
The MYB transcription factor and the ABA-independent
cold acclimation pathway
ABA-dependent gene expression is regulated by tran-
scription factors that belong to the bZIP (ABRE binding
factors—or AREBs), MYC and MYB families. In A. thali-
ana, the MYB family transcription factor PAP2 regulates
the flavonoid biosynthesis pathway, which is reported to be
involved in cold tolerance (see above). OsMYB4, a member
of the MYB family of transcription factors, has also been
shown to be inducible by cold, but not by ABA in rice
(Park et al. 2010). Surprisingly, Su et al. (2010) reported
that MYBS3 repressed the DREB1/CBF-dependent cold-
signalling pathway in rice at the transcriptional level.
Further, DREB1 responded quickly and transiently,
whereas MYBS3 responded slowly to cold stress, suggest-
ing distinct pathways acting sequentially to acclimate rice
to cold stress. In A. thaliana the abundance of MYBC1
transcripts was not affected by overexpression of CBF1,
CBF2, and CBF3, suggesting that MYBC1 is not down-
regulated by these CBF family members (Zhai et al. 2010).
Abscisic acid (ABA)-dependent cold signal pathway
and other phytohormones
(a) Abscisic acid: ABA serves as a secondary signal that
plays at least some role in the transduction of cold signals
via second messengers, such as H2O2 and Ca2?. This is
demonstrated by the los5 (low expression of osmotically
responsive genes) mutant, which exhibits significantly
decreased cold- and salt/drought-induced expression of
COR. ABA enhanced antioxidant defence and slowed
down the accumulation of ROS caused by low tempera-
tures (Liu et al. 2011). ABA can also induce the expression
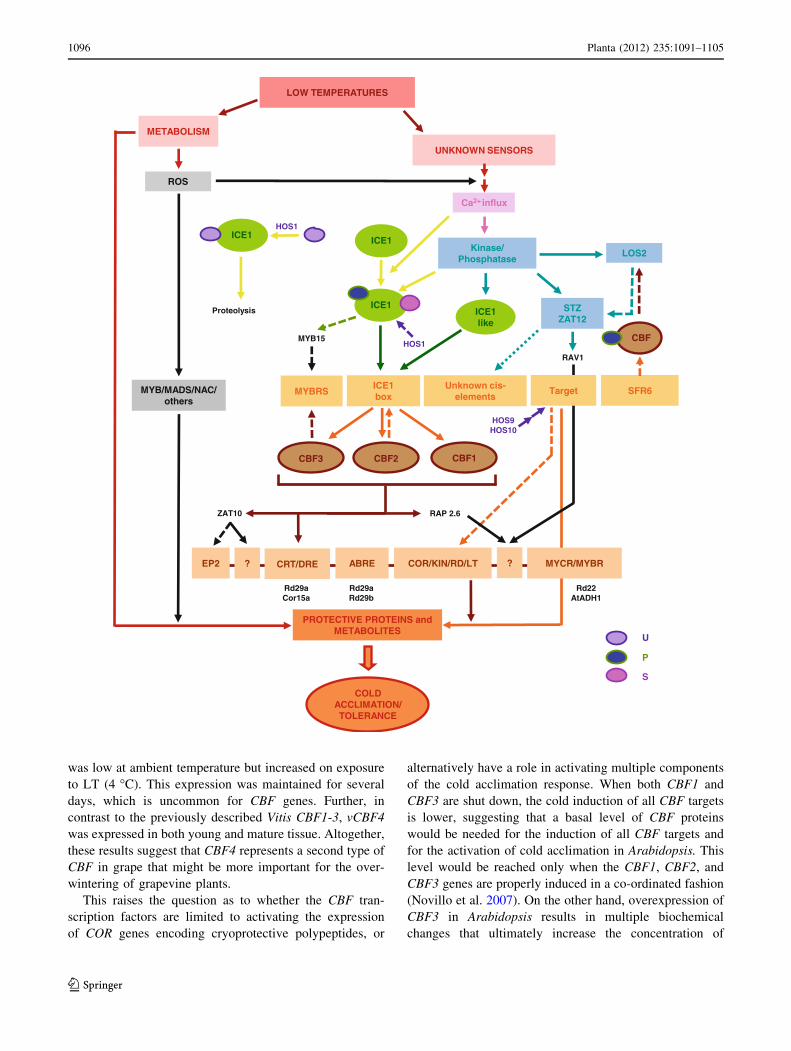
Fig. 4 Schematic diagram of the regulatory network involved in low
temperature responses. Plants sense low temperatures and activate a
calcium signal mediated via protein kinases, and other species that
activate multiple transcriptional cascades, one of which involves
ICE1 and CBFs. The CBF genes play important roles in cold
acclimation and are regulated by multiple pathways. They activate the
transcription of CBFs and repress MYB15. ICE1, a constitutively
expressed gene, is activated by cold stress via sumoylation and
phosphorylation. HOS1 is a RING-type ubiquitin E3 ligase that
negatively regulates cold-induced DREB1/CBF expression. The
constitutive HOS9 and HOS10 regulons have a role in the negative
regulation of CBF-target genes. CBFs regulate the expression of CORgenes that confer freezing tolerance. The expression of CBFs is
negatively regulated by MYB15 and ZAT12. HOS1 mediates the
ubiquitination and proteosomal degradation of ICE1 and, thus,
negatively regulates CBF regulons. Transcription of CBFs might be
cross-regulated. Transcription factor binding sites are represented at
the bottom of the diagram, with the representative promoters listed
below. Yellow arrows indicate post-translational regulation; solidarrows indicate activation, whereas broken lines show negative
regulation; small circles indicate post-transcriptional modification,
such as phosphorylation; question marks indicates unknown cis-
elements. ABRE ABA responsive element, CBF C-repeat binding
factor (an AP2-type transcription factor); COR cold-responsive genes,
CRT C-repeat elements, DRE dehydration-responsive elements,
HOS1 high expression of osmotically responsive genes1, HOS9 and
HOS10, ICE1 inducer of CBF expression 1, KIN cold-induced genes,
LOS2 low expression of osmotically responsive genes 2 (a bifunc-
tional enolase with transcriptional repression activity), LTI low
temperature-induced genes, MYB myeloblastosis, MYBRS MYB
recognition sequence, MYCRS MYC recognition sequence, RDresponsive to dehydration) genes, ROS reactive oxygen species,
SIZ1 SAP and MiZ1 (a SUMO E3 ligase), P phosphorylation,
S SUMO (small ubiquitin-related modifier), U ubiquitin
b
Planta (2012) 235:1091–1105 1097
123

of the CBF1, CBF2, and CBF3 genes, but to a significantly
lower level than that caused by cold (Knight et al. 2004).
(b) Polyamines: There are relatively few reports
regarding the involvement of polyamines in LT stress
(Alcazar et al. 2010; Gill and Tuteja 2010). Putrescine
accumulation under cold stress is essential for proper cold
acclimation and survival at freezing temperatures (Cuevas
et al. 2008). Indeed, Arabidopsis mutants defective in
putrescine biosynthesis (adc1, adc2) exhibit reduced
freezing tolerance compared to wild-type plants, suggest-
ing that the detrimental consequences of putrescine
depletion during cold stress are at least partially due to
changes in the concentration of ABA. On the other hand,
the accumulation of putrescine in the early stages of mango
fruit ripening promoted by LT stress did not prevent
chilling injury (Nair and Singh 2004). During treatment,
the spermidine content of leaves increased substantially in
cold-tolerant cucumber cultivars but not in sensitive ones,
while the concentrations of putrescine and spermine did not
change (Shen et al. 2000). The depletion of endogenous
spermidine and spermine in response to chilling and the
reduced severity of chilling injuries in mango fruits that
had been treated with these polyamines prior to storage
both suggest that polyamine biosynthesis influences cold
sensitivity. Polyamines have also been reported to have a
role in alleviating oxidative stress: inhibiting polyamine
synthesis causes increased oxidative damage in cold-trea-
ted plants (Groppa and Benavides 2008).
(c) Other regulators affecting cold acclimation: A
number of genes involved in the biosynthesis or signalling
of plant hormones, such as gibberellic acid and auxin, may
be also regulated by cold stress. The regulation of these
genes might be important in co-ordinating cold tolerance
with growth and development. A role for cytokinin in
mediating plant growth rates at LT was recently reported
(Xia et al. 2009). Mutants with elevated cytokinin levels
(amp1) displayed enhanced cell division at 4 �C. More-
over, no changes in CBF expression were recorded in amp1
or NahG plants at LT, suggesting that the effects of cyto-
kinin and salicylic acid (SA) on temperature-regulated
growth are independent of the CBF regulon. In addition to
its role in plant pathogen defences, SA is also involved in
suppressing plant growth during chilling and accumulates
at LT (Scott et al. 2004). However, SA is not required for
thermotolerance (Clarke et al. 2004).
Post-transcriptional regulation
Transcriptomic analyses of gene expression at the mRNA
level have contributed greatly to our understanding of cold
responses in Arabidopsis (Kreps et al. 2002; Zhou et al.
2008, 2011). However, the abundance of individual
mRNAs does not always correlate well with that of the
corresponding proteins, which are the crucial agents in the
cell (MacKay et al. 2004; Tian et al. 2004). Consequently,
it is not sufficient to simply predict protein expression
levels from quantitative mRNA data, mainly due to the
effects of post-transcriptional regulation mechanisms
(Pradet-Balade et al. 2001).
The involvement of post-transcriptional cold-induced
regulation on the abundance of specific mRNAs has been
reported in several species, including Arabidopsis and
alfalfa. Maize plants exposed to cooler temperatures
respond by phosphorylating a minor chlorophyll a/b pro-
tein rather than synthesizing a new protein from a cold-
regulated gene (Bergantino et al. 1995). Although plant cell
membranes exhibit significantly higher levels of unsatura-
tion at lower temperatures, there is no apparent increase in
rate of transcription or stability of fatty acid desaturase
mRNA at lower temperature. The exception is the Ara-
bidopsis fatty acid desaturase gene FAD8 (Matsuda et al.
2005), which suggests that plant fatty acid desaturases are
regulated at the post-transcriptional level.
Expression of homology to pathogenesis-related (PR)
genes and synthesis of antifreeze proteins (AFPS)
Studies in different plant species have shown that several
cold-induced genes encode cryoprotective proteins (Hincha
2002). In the last few decades, research has focused on
specific proteins with antifreeze activity that accumulate in
the apoplast during cold acclimation, thereby offering plant
resistance against freezing (Griffith and Yaish 2004;
Griffith et al. 2005; Yaish et al. 2006) (Fig. 5). These
proteins have been found in many overwintering vascular
plants (Griffith and Yaish 2004; Venketesh and Dayananda
2008), but antifreeze activity is present only after their
exposure to LT and only in plants that tolerate the presence
of ice in their tissues (Griffith and Yaish 2004; Yaish et al.
2006). These proteins were identified as b-1,3-glucanase-
like proteins, chitinase-like proteins, thaumatin-like pro-
teins and as polygalacturonase inhibitor proteins (Wang
et al. 2006; Yaish et al. 2006). Although they were present
in non-acclimated plants, they were found in different
locations and did not exhibit antifreeze activity, which
suggests that different isoforms of pathogenesis-related
proteins are produced under LT conditions (Antikainen
et al. 1996; Wang et al. 2006).
Until now, no plant has been reported to have consti-
tutive antifreeze activity. Rather, all studies have shown
that transcripts and translation products of AFP genes
accumulate during cold acclimation (Yeh et al. 2000;
Huang and Duman 2002; Wang et al. 2006). Recent studies
have shown that many PR genes are induced and disease
resistance is enhanced after exposure to LT, linking cold
signals with pathogenesis in plants (Seo et al. 2010).
1098 Planta (2012) 235:1091–1105
123

By accumulating PR proteins during cold acclimation,
overwintering grasses and cereals acquire a systemic, non-
specific, pre-emptive defence against pathogens and thus
exhibit greater disease resistance.
Cold shock proteins and RNA binding proteins
Cold shock domain proteins (CSDPs) play important roles
in development and stress adaptation in a variety of
organisms, ranging from bacteria to mammals (Chaikam
and Karlson 2010). In higher plants, cold shock domain
proteins are involved in the cold response (Nakaminami
et al. 2006; Sasaki et al. 2007). Recently, it was demon-
strated that CSP Arabidopsis 3 (AtCSP3), which shares a
cold shock domain with the CSDPs, is involved in the
acquisition of freezing tolerance in plants (Kim et al.
2009).
COR/LEA and dehydrins
The accumulation of hydrophilic proteins predicted to form
an amphipathic a-helix is one of the best documented
responses of plants to cold treatment (Eriksson et al. 2011).
Most of these proteins have therefore been named COR
(cold responsive), LTI (low temperature induced), RAB
(responsive to abscisic acid), KIN (cold induced) or ERD
(early responsive to dehydration). These proteins include the
dehydrins, which belong to group II of the late embryogen-
esis abundant (LEA) proteins (Bies-Etheve et al. 2008).
The accumulation of one particular dehydrin (WCO
R410) is correlated with the capacity to develop freezing
tolerance in wheat. However, the overexpression of single
dehydrins does not necessarily lead to enhanced freezing
tolerance. For instance, the overexpression of RAB18, a
cold-induced dehydrin, has no effect on freezing tolerance in
Arabidopsis (Lang and Palva 1992). This suggests that to
fully play their role, dehydrins may need to be activated by a
cold-induced mechanism such as protein phosphorylation.
One of the roles that have been also attributed to dehydrins is
the prevention of membrane destabilization during dehy-
dration. In addition to this postulated function, it has been
proposed that dehydrins possess cryoprotective (Bravo et al.
2003) or antifreeze (Puhakainen et al. 2004) activities.
Cold-induced osmolites/osmoprotectants
In response to cold and other osmotic stresses, plants
accumulate a range of compatible solutes including cer-
erosides, free sterols, sterol glucosides and acylatedsterols,
glucosides, raffinose, arbinoxylans, and other soluble sug-
ars. In addition, plants accumulate other solutes such as
glutamic acid, amino acids (alanine, glycine, proline, and
serine), polyamines and betaines (Hekneby et al. 2006;
Patton et al. 2007; Ruelland and Zachowski 2010). These
different molecules, which are often degraded once the
stress has passed, are referred to as osmolytes, osmopro-
tectants or compatible solutes.
Carbohydrate changes
Carbohydrate metabolism has been reported to have greater
instantaneous low temperature sensitivity than other com-
ponents of photosynthesis (Fernandez et al. 2012). Although
the precise function of soluble sugars remains to be deter-
mined, their accumulation in cold-acclimated plants sug-
gests roles as osmoregulators, cryoprotectants or signalling
molecules (Welling and Palva 2006). Sugars play multiple
roles in low temperature tolerance. As typical compatible
osmolytes, they contribute to the preservation of water
within plant cells, thereby reducing water availability for ice
nucleation in the apoplast (Uemura and Steponkus 1999;
Ruelland et al. 2009). Sugars might protect plant cell mem-
branes during cold-induced dehydration, replacing water
molecules in establishing hydrogen bonds with lipid mole-
cules (Uemura et al. 2003; Ruelland et al. 2009). Moreover,
carbohydrates may also act as scavengers of reactive oxygen
Resistance to pathogens
Cold acclimation
Accumulation of antifreeze proteins (AFPs)
Cold acclimation
Chilling stress
Accumulation of transcripts homology to PR genes
Pathogen attack
Accumulation of PR proteins
Expression of genesEncoding for PR proteins
Salicylic acidAbscisic acid
EthyleneJasmonic acid
Fig. 5 Accumulation of antifreeze proteins (AFPs) in cold-accli-
mated plants with antifungal activity (adapted from Yeh et al. 2000;
Huang and Duman 2002; Griffith and Yaish 2004; Yaish et al. 2006).
By accumulating PR proteins during cold acclimation, overwintering
plants may acquire a systemic, non-specific, pre-emptive defence
against pathogens and exhibit greater disease resistance
Planta (2012) 235:1091–1105 1099
123

species and contribute to increased membrane stabilization
(Bohnert and Sheveleva 1998). Sugar signalling is also clo-
sely associated with hormone signalling, the control of
growth and development, and stress responses in plants
(Zeng et al. 2011).
Depending on the plant species, various forms of soluble
sugars are involved in physiological reactions to cold
stress. For example, treatment of rice seedlings with fruc-
tose or glucose prior to LT treatment increases their
resistance to cold. Cotton cotyledon discs floating on a
sucrose solution in the dark were less injured by cold than
those on non-sugar solutions (Couee et al. 2006). On the
other hand, King et al. (1988) reported that cold tolerance
in tomato seedlings decreased following pronounced
reductions in their starch and sugar levels during a dark
period.
The soluble carbohydrate content of grasses can undergo
a tenfold increase within 8 h of transfer from a warm to a
cold environment (Pollock and Lloyd 1987). The oligo-
saccharides raffinose and stachyose are especially associ-
ated with cold hardiness, low temperature and dormancy
(Aıt Barka and Audran 1996; Couee et al. 2006). More-
over, the concentration of sucrose, the most easily detect-
able sugar in cold-tolerant species, increases several fold
during exposure to LT (Bohnert and Sheveleva 1998;
Tabaei-Aghdaei et al. 2003). The accumulation of sucrose
in cane sugar exposed to salt stress or to LT stress supports
the role of this sugar as an osmoprotectant that stabilizes
cellular membranes and maintains turgor (Jouve et al.
2004). In addition, high sucrose levels correlate with the
priming of defence responses in rice that overexpresses the
PRms gene from maize, which encodes a PR-1 type protein
(Casacuberta et al. 1991).
Trehalose is a non-reducing disaccharide of glucose that
is found in a variety of organisms including bacteria, yeast,
fungi, insects and invertebrates, where it serves as a stress
protectant and/or a reserve carbohydrate (Penna 2003;
Fernandez et al. 2010). Although increased levels of tre-
halose are associated with abiotic stress tolerance in
transgenic plants expressing heterologous microbial genes,
the function of endogenous trehalose in higher plants
remains unclear. This sugar possesses the unique capacity
for reversible water absorption and appears to be superior
to other sugars in protecting biological molecules from
desiccation-induced damage (Fernandez et al. 2010). Fur-
ther, transgenic A. thaliana plants that accumulated treha-
lose displayed significantly enhanced freezing tolerance
(Miranda et al. 2007). Increases in trehalose concentration
may also be involved in starch accumulation (Fernandez
et al. 2010).
During exposure to LT, starch content typically declines
following hydrolysis, and there is a corresponding increase
in the concentration of free saccharides (Pollock and Lloyd
1987; Bohnert and Sheveleva 1998). However, in several
cases, increases in the levels of both soluble sugars and
starch have been reported during cold acclimation. For
instance, in cabbage seedlings grown at 5 �C, the concen-
trations of starch and all soluble sugars (myo-inositol aside)
in the leaves increased gradually during cold acclimation
(Sasaki et al. 1996). However, the induced freezing toler-
ance was lost after only 1 day of acclimation at control
temperatures and this change was associated with a large
reduction in sugar content.
Carbohydrate accumulation at LT may be explained
through the activation of specific enzymes (Bohnert and
Sheveleva 1998; Couee et al. 2006). This suggests that
although LT inhibits sucrose synthesis and photosynthesis,
various biochemical and physiological adaptations to LT
counteract these effects. These adaptations include the
post-translational activation and enhanced expression of
enzymes involved in the sucrose synthesis pathways and
those of Calvin cycle—in particular, the cytosolic enzymes
fructose-1,6-bisphosphatase, sucrose phosphate synthase
and sucrose synthase (Stitt and Hurry 2002).
Compatible osmotica other than sugars
(a) Proline: The positive correlation between the accu-
mulation of endogenous proline (Pro) and improved cold
tolerance has been found mostly in LT-insensitive plants
such as barley, rye, winter wheat, grapevine, potato,
chickpea and A. thaliana (Verbruggen and Hermans 2008;
Szabados and Savoure 2010; Kaur et al. 2011). Proline
plays multiple roles in plant stress tolerance, as a mediator
of osmotic adjustment, a stabilizer of proteins and mem-
branes, an inducer of osmotic stress-related genes, and as a
scavenger of ROS (Verbruggen and Hermans 2008;
Szabados and Savoure 2010; Theocharis et al. 2011). The
most probable roles of proline are to (a) regulate cytosol
acidity, (b) stabilize the NAD?/NADH ratio, (c) increase
the photochemical activity of the photosystem II in thyla-
koid membranes and (d) decrease lipid peroxidation
(Kishor et al. 2005). Most chilling-sensitive plants that
accumulate Pro under LT conditions do not acquire cold
tolerance (Kushad and Yelenosky 1987), unless a high
concentration of Pro was applied prior to stress (Xin and Li
1993). It appears therefore that proline possesses the
potential to alleviate LT injury in chilling-sensitive plants,
but for some reason this system fails under natural
conditions.
(b) Glycine betaine: The accumulation of glycine beta-
ine (GB) usually correlates with the plant’s level of stress
tolerance. Both the genetically engineered biosynthesis of
GB in plants that do not naturally accumulate GB and the
exogenous application of GB enhance the tolerance of such
plants to various abiotic stresses (Chen and Murata 2008).
1100 Planta (2012) 235:1091–1105
123

Possible roles for GB include stabilization of the tran-
scriptional and translational machinery. GB stabilizes
protein complexes and membranes in vitro and may indi-
rectly induce H2O2-mediated signalling pathways.
Effect of microorganisms on cold tolerance
Deleterious effects
Both the aerial parts of the plant and the rhizospheric zone
harbour hundreds of species of bacteria, yeast and fungi.
Several bacterial and fungal species have the ability to
nucleate ice at high sub-freezing temperatures. Bacterial
species with ice nucleation activity (Ice? bacteria) such as
Pseudomonas syringae contribute to frost injury in many
frost-sensitive plant species by reducing their ability to
supercool and avoid the formation of membrane damaging
ice crystals (Lindow and Leveau 2002). Other ice nucle-
ating bacterial species include P. fluorescens, Erwinia
herbicola, and some strains of Xanthomonas campestris, as
well as related strains. Some species of Fusarium and
related genera of fungi are also active in ice nucleation.
Beneficial effects
One way to reduce the incidence of LT damage is to use
beneficial microorganisms that enhance plant growth and
improve their resistance to stress. Alternatively, beneficial
bacteria may also be used to eliminate the Ice? bacteria
from plant surfaces. Since the ice nucleation temperature
increases with the population size of Ice? bacteria, pre-
emptive competitive exclusion of Ice? bacteria by natu-
rally occurring non-ice nucleating active bacteria could be
an effective and practical method for managing frost in
cold-sensitive plants (Lindow and Leveau 2002). The
bacteria could also be genetically altered to not carry the
genetic instructions needed to produce the ice nucleating
protein. If such new bacteria (INA-) were sprayed onto
plants at very high concentrations, naturally occurring
bacteria (INA?) would not be able to compete. Recombi-
nant Ice- bacteria, the first engineered microorganisms
released into the open environment in field experiments,
have been used to illustrate the specificity with which
competitive exclusion of Ice? bacteria occurs (Skirvin
et al. 2000).
Several endophytic bacteria have been reported to
induce resistance against biotic stress and tolerance to
abiotic stress in several plants. For instance, a plant
growth-promoting rhizobacterium (PGPR), Burkholderia
phytofirmans strain PsJN, is able to reduce chilling-induced
damage (Ait Barka et al. 2006). Similar conclusions were
reported during the interaction between Chorispora bun-
geana and the endophyte Clavibacter sp. strain Enf12
(Ding et al. 2011). In attempt to explain how some bene-
ficial bacteria may influence cold tolerance, Theocharis
et al. (2011) reported that several stress-related gene tran-
scripts (e.g., StSy, PAL, Chit4c, Chit1b, Gluc and LOX) and
changes in levels of several stress-related metabolites (e.g.,
proline, malondialdehyde and other aldehydes known to be
lipid peroxidation markers, and hydrogen peroxide)
increased earlier, faster, and were more pronounced in
chilled-PsJN-bacterized plantlets. This is consistent with
the ‘priming’ concept (Theocharis et al. 2011), supporting
the establishment of a mutualistic relationship between the
bacterium and the grapevine. The endophyte participates in
the cold acclimation process via a scavenging system (Ding
et al. 2011; Theocharis et al. 2011). The reported increase
in the expression of Chit1b, Chit4c and Gluc was not
surprising because chitinases and glucanases can be clas-
sified as either antifreeze (AFPs, Griffith and Yaish 2004)
or PR (Van Loon and Van Strien 1999) proteins; this
confirms the link between cold signals and pathogenesis in
plants, as shown in Fig. 5. Further, we demonstrated
recently that photosynthesis is modulated by the presence
of beneficial bacteria in grapevine plantlets, suggesting that
the modification of carbohydrate metabolism is one of the
major modes by which PGPR reduces chilling-induced
damage (Fernandez et al. 2012).
Concluding remarks and future perspectives
Plant physiologists and plant molecular biologists have
always been interested in mechanisms involved in plant
tolerance to cold and how plants may react to withstand
damage following stress. The biological and physiological
changes that occur following cold exposure have been
particularly well-studied. Studies conducted in recent years
have analysed the cold signal, the genes that act down-
stream of it to induce cold acclimation, and the overall
cascade of molecular events that occur following cold
perception. This has generated a large amount of data that
requires collation and interpretation. Thus, there is a real
need for a comprehensive model that encapsulates this
multi-step process.
Acknowledgments The first author (A.T.) was supported by a Grant
from the Greek State Scholarship Foundation (I.K.Y.).
References
Airaki M, Leterrier M, Mateos RM, Valderrama R, Chaki M, Barroso
JB, Del Rio LA, Palma JM, Corpas FJ (2011) Metabolism of
reactive oxygen species and reactive nitrogen species in pepper
(Capsicum annuum L.) plants under low temperature stress.
Plant Cell Environ 35:281–295
Planta (2012) 235:1091–1105 1101
123

Aıt Barka E, Audran JC (1996) Reponse des vignes champenoises aux
temperatures negatives: effet d’un refroidissement controle sur
les reserves glucidiques du complexe gemmaire avant et au cours
du debourrement. Can J Bot 74:492–505
Ait Barka E, Nowak J, Clement C (2006) Enhancement of chilling
resistance of inoculated grapevine plantlets with a plant growth-
promoting rhizobacterium, Burkholderia phytofirmans strain
PsJN. Appl Environ Microbiol 72:7246–7252
Alcazar R, Altabella T, Marco F, Bortolotti C, Reymond M, Koncz C,
Carrasco P, Tiburcio AF (2010) Polyamines: molecules with
regulatory functions in plant abiotic stress tolerance. Planta
231:1237–1249
Antikainen M, Griffith M, Zhang J, Hon WC, Yang D, Pihakaski-
Maunsbach K (1996) Immunolocalization of antifreeze proteins
in winter Rye leaves, crowns, and roots by tissue printing. Plant
Physiol 110:845–857
Baena-Gonzalez E, Gray JC, Tyystjarvi E, Aro EM, Maenpaa P
(2001) Abnormal regulation of photosynthetic electron transport
in a chloroplast ycf9 inactivation mutant. J Biol Chem 276:
20795–20802
Bergantino E, Dainese P, Cerovic Z, Sechi S, Bassi R (1995) A post-
translational modification of the photosystem II subunit CP29
protects maize from cold stress. J Biol Chem 270:8474–8481
Bies-Etheve N, Gaubier-Comella P, Debures A, Lasserre E, Jobet E,
Raynal M, Cooke R, Delseny M (2008) Inventory, evolution and
expression profiling diversity of the LEA (late embryogenesis
abundant) protein gene family in Arabidopsis thaliana. Plant
Mol Biol 67:107–124
Bohnert HJ, Sheveleva E (1998) Plant stress adaptations—making
metabolism move. Curr Opin Plant Biol 1:267–274
Bravo LA, Gallardo J, Navarrete A, Olave N, Martınez J, Alberdi M,
Close TJ, Corcuera LJ (2003) Cryoprotective activity of a cold-
induced dehydrin purified from barley. Physiol Plant 118:262–
269
Casacuberta JM, Puigdomenech P, San Segundo B (1991) A gene
coding for a basic pathogenesis-related (PR-like) protein from
Zea mays. Molecular cloning and induction by a fungus
(Fusarium moniliforme) in germinating maize seeds. Plant Mol
Biol 16:527–536
Catala R, Santos E, Alonso JM, Ecker JR, Martinez-Zapater JM,
Salinas J (2003) Mutations in the Ca2?/H? transporter CAX1
increase CBF/DREB1 expression and the cold-acclimation
response in Arabidopsis. Plant Cell 15:2940–2951
Chaikam V, Karlson DT (2010) Comparison of structure, function
and regulation of plant cold shock domain proteins to bacterial
and animal cold shock domain proteins. BMB Rep 43:1–8
Chen TH, Murata N (2008) Glycinebetaine: an effective protectant
against abiotic stress in plants. Trends Plant Sci 13:499–505
Chinnusamy V, Ohta M, Kanrar S, Lee BH, Hong X, Agarwal M, Zhu
JK (2003) ICE1: a regulator of cold-induced transcriptome and
freezing tolerance in Arabidopsis. Genes Develop 17:1043–1054
Chinnusamy V, Schumaker K, Zhu JK (2004) Molecular genetic
perspectives on cross-talk and specificity in abiotic stress
signalling in plants. J Exp Bot 55:225–236
Chinnusamy V, Zhu J, Zhu JK (2007) Cold stress regulation of gene
expression in plants. Trends Plant Sci 12:444–451
Chinnusamy V, Zhu JK, Sunkar R (2010) Gene regulation during cold
stress acclimation in plants. Methods Mol Biol 639:39–55
Clarke SM, Mur LA, Wood JE, Scott IM (2004) Salicylic acid
dependent signaling promotes basal thermotolerance but is not
essential for acquired thermotolerance in Arabidopsis thaliana.
Plant J 38:432–447
Couee I, Sulmon C, Gouesbet G, El Amrani A (2006) Involvement of
soluble sugars in reactive oxygen species balance and responses
to oxidative stress in plants. J Exp Bot 57:449–459
Crifo T, Puglisi I, Petrone G, Recupero GR, Lo Piero AR (2011)
Expression analysis in response to low temperature stress in
blood oranges: implication of the flavonoid biosynthetic path-
way. Gene 478:1–9
Cuevas JC, Lopez-Cobollo R, Alcazar R, Zarza X, Koncz C, Altabella
T, Salinas J, Tiburcio AF, Ferrando A (2008) Putrescine is
involved in Arabidopsis freezing tolerance and cold acclimation
by regulating abscisic acid levels in response to low temperature.
Plant Physiol 148:1094–1105
Ding S, Huang CL, Sheng HM, Song CL, Li YB, An LZ (2011) Effect
of inoculation with the endophyte Clavibacter sp. strain Enf12
on chilling tolerance in Chorispora bungeana. Physiol Plant
141:141–151
Doherty CJ, Van Buskirk HA, Myers SJ, Thomashow MF (2009)
Roles for Arabidopsis CAMTA transcription factors in cold-
regulated gene expression and freezing tolerance. Plant Cell
21:972–984
Du L, Poovaiah BW (2005) Ca2?/calmodulin is critical for brassi-
nosteroid biosynthesis and plant growth. Nature 437:741–745
Eriksson SK, Kutzer M, Procek J, Grobner G, Harryson P (2011)
Tunable membrane binding of the intrinsically disordered
dehydrin Lti30, a cold-induced plant stress protein. Plant Cell
23:2391–2404
Fernandez O, Bethencourt L, Quero A, Sangwan RS, Clement C
(2010) Trehalose and plant stress responses: friend or foe?
Trends Plant Sci 15:409–417
Fernandez O, Theocharis A, Bordiec S, Feil R, Jacquens L, Clement
C, Fontaine F, Ait Barka E (2012) Burkholderia phytofirmansstrain PsJN acclimates grapevine to cold by modulating carbo-
hydrates metabolism. Mol Plant Microbe Interact 25:496–504
Fowler S, Thomashow MF (2002) Arabidopsis transcriptome profil-
ing indicates that multiple regulatory pathways are activated
during cold acclimation in addition to the CBF cold response
pathway. Plant Cell 14:1675–1690
Fursova OV, Pogorelko GV, Tarasov VA (2009) Identification of
ICE2, a gene involved in cold acclimation which determines
freezing tolerance in Arabidopsis thaliana. Gene 429:98–103
Gechev T, Willekens H, Van Montagu M, Inze D, Van Camp W,
Toneva V, Minkov I (2003) Different responses of tobacco
antioxidant enzymes to light and chilling stress. J Plant Physiol
160:509–515
Gill SS, Tuteja N (2010) Polyamines and abiotic stress tolerance in
plants. Plant Signal Behav 5:26–33
Gilmour SJ, Zarka DG, Stockinger EJ, Salazar MP, Houghton JM,
Thomashow MF (1998) Low temperature regulation of the
Arabidopsis CBF family of AP2 transcriptional activators as an
early step in cold-induced COR gene expression. Plant J 16:433–
442
Gilmour SJ, Sebolt AM, Salazar MP, Everard JD, Thomashow MF
(2000) Overexpression of the Arabidopsis CBF3 transcriptional
activator mimics multiple biochemical changes associated with
cold acclimation. Plant Physiol 124:1854–1865
Gilmour SJ, Fowler SG, Thomashow MF (2004) Arabidopsistranscriptional activators CBF1, CBF2, and CBF3 have matching
functional activities. Plant Mol Biol 54:767–781
Goulas E, Schubert M, Kieselbach T, Kleczkowski LA, Gardestrom
P, Schroder W, Hurry V (2006) The chloroplast lumen and
stromal proteomes of Arabidopsis thaliana show differential
sensitivity to short- and long-term exposure to low temperature.
Plant J 47:720–734
Griffith M, Yaish MW (2004) Antifreeze proteins in overwintering
plants: a tale of two activities. Trends Plant Sci 9:399–405
Griffith M, Lumb C, Wiseman SB, Wisniewski M, Johnson RW,
Marangoni AG (2005) Antifreeze proteins modify the freezing
process in planta. Plant Physiol 138:330–340
1102 Planta (2012) 235:1091–1105
123

Groppa MD, Benavides MP (2008) Polyamines and abiotic stress:
recent advances. Amino Acids 34:35–45
Haake V, Cook D, Riechmann JL, Pineda O, Thomashow MF, Zhang
JZ (2002) Transcription factor CBF4 is a regulator of drought
adaptation in Arabidopsis. Plant Physiol 130:639–648
Han H, Gao S, Li B, Dong XC, Feng HL, Meng QW (2010)
Overexpression of violaxanthin de-epoxidase gene alleviates
photoinhibition of PSII and PSI in tomato during high light and
chilling stress. J Plant Physiol 167:176–183
Hekneby M, Antolın MC, Sanchez-Dıaz M (2006) Frost resistance
and biochemical changes during cold acclimation in different
annual legumes. Environ Exp Bot 55:305–314
Hincha DK (2002) Cryoprotectin: a plant lipid-transfer protein
homologue that stabilizes membranes during freezing. Philos
Trans R Soc Lond B Biol Sci 357:909–916
Hsieh TH, Lee JT, Yang PT, Chiu LH, Charng YY, Wang YC, Chan
MT (2004) Heterology expression of the Arabidopsis C-repeat/dehydration response element binding factor 1 gene confers
elevated tolerance to chilling and oxidative stresses in transgenic
tomato. (vol 129, pg 1086, 2002). Plant Physiol 135:1145–1155
Huang T, Duman JG (2002) Cloning and characterization of a thermal
hysteresis (antifreeze) protein with DNA-binding activity from
winter bittersweet nightshade, Solanum dulcamara. Plant Mol
Biol 48:339–350
Ingram J, Bartels D (1996) The molecular basis of dehydration
tolerance in plants. Annu Rev Plant Physiol Plant Mol Biol
47:377–403
Ivanov AG, Sane PV, Krol M, Gray GR, Balseris A, Savitch LV,
Oquist G, Huner NP (2006) Acclimation to temperature and
irradiance modulates PSII charge recombination. FEBS Lett
580:2797–2802
Jaglo-Ottosen KR, Gilmour SJ, Zarka DG, Schabenberger O,
Thomashow MF (1998) Arabidopsis CBF1 overexpression
induces COR genes and enhances freezing tolerance. Science
280:104–106
Jouve L, Hoffmann L, Hausman JF (2004) Polyamine, carbohydrate,
and proline content changes during salt stress exposure of aspen
(Populus tremula L.): involvement of oxidation and osmoreg-
ulation metabolism. Plant Biol 6:74–80
Kaur G, Kumar S, Thakur P, Malik JA, Bhandhari K, Sharma KD,
Nayyar H (2011) Involvement of proline in response of chickpea
(Cicer arietinum L.) to chilling stress at reproductive stage. Sci
Hortic 128:174–181
Kim MH, Sasaki K, Imai R (2009) Cold shock domain protein 3
regulates freezing tolerance in Arabidopsis thaliana. J Biol
Chem 284:23454–23460
King AI, Joyce DC, Reid MS (1988) Role of carbohydrates in diurnal
chilling sensitivity of tomato seedlings. Plant Physiol 86:764–
768
Kishor PBK, Sangam S, Amrutha RN, Laxmi PS, Naidu KR, Rao K,
Rao S, Reddy KJ, Theriappan P, Sreenivasulu N (2005)
Regulation of proline biosynthesis, degradation, uptake and
transport in higher plants: Its implications in plant growth and
abiotic stress tolerance. Curr Sci 88:424–438
Knight H, Brandt S, Knight MR (1998) A history of stress alters
drought calcium signalling pathways in Arabidopsis. Plant J
16:681–687
Knight H, Zarka DG, Okamoto H, Thomashow MF, Knight MR
(2004) Abscisic acid induces CBF gene transcription and
subsequent induction of cold-regulated genes via the CRT
promoter element. Plant Physiol 135:1710–1717
Kreps JA, Wu Y, Chang HS, Zhu T, Wang X, Harper JF (2002)
Transcriptome changes for Arabidopsis in response to salt,
osmotic, and cold stress. Plant Physiol 130:2129–2141
Krol M, Ivanov AG, Jansson S, Kloppstech K, Huner NP (1999)
Greening under high light or cold temperature affects the level of
xanthophyll-cycle pigments, early light-inducible proteins, and
light-harvesting polypeptides in wild-type barley and the Chlo-rina f2 mutant. Plant Physiol 120:193–204
Kushad MM, Yelenosky G (1987) Evaluation of polyamine and
proline levels during low temperature acclimation of citrus. Plant
Physiol 84:692–695
Lang V, Palva ET (1992) The expression of a rab-related gene, rab18,
is induced by abscisic acid during the cold acclimation process
of Arabidopsis thaliana (L.) Heynh. Plant Mol Biol 20:951–962
Laugier E, Tarrago L, Vieira Dos Santos C, Eymery F, Havaux M,
Rey P (2010) Arabidopsis thaliana plastidial methionine
sulfoxide reductases B, MSRBs, account for most leaf peptide
MSR activity and are essential for growth under environmental
constraints through a role in the preservation of photosystem
antennae. Plant J 61:271–282
Lindow SE, Leveau JHJ (2002) Phyllosphere microbiology. Curr
Opin Biotech 13:238–243
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki
K, Shinozaki K (1998) Two transcription factors, DREB1 and
DREB2, with an EREBP/AP2 DNA binding domain separate
two cellular signal transduction pathways in drought- and low-
temperature-responsive gene expression, respectively, in Ara-bidopsis. Plant Cell 10:1391–1406
Liu Y, Jiang H, Zhao Z, An L (2011) Abscisic acid is involved in
brassinosteroids-induced chilling tolerance in the suspension
cultured cells from Chorispora bungeana. J Plant Physiol 168:
853–862
Lyons JM (1973) Chilling injury in plants. Annu Rev Plant Physiol
Plant Mol Biol 24:445–466
MacKay VL, Li X, Flory MR, Turcott E, Law GL, Serikawa KA, Xu
XL, Lee H, Goodlett DR, Aebersold R, Zhao LP, Morris DR
(2004) Gene expression analyzed by high-resolution state array
analysis and quantitative proteomics: response of yeast to mating
pheromone. Mol Cell Proteomics 3:478–489
Mantyla E, Lang V, Palva ET (1995) Role of abscisic acid in drought-
induced freezing tolerance, cold acclimation, and accumulation
of LT178 and RAB18 proteins in Arabidopsis thaliana. Plant
Physiol 107:141–148
Maruyama K, Sakuma Y, Kasuga M, Ito Y, Seki M, Goda H, Shimada
Y, Yoshida S, Shinozaki K, Yamaguchi-Shinozaki K (2004)
Identification of cold-inducible downstream genes of the
Arabidopsis DREB1A/CBF3 transcriptional factor using two
microarray systems. Plant J 38:982–993
Matsuda O, Sakamoto H, Hashimoto T, Iba K (2005) A temperature-
sensitive mechanism that regulates post-translational stability of
a plastidial omega-3 fatty acid desaturase (FAD8) in Arabidopsisleaf tissues. J Biol Chem 280:3597–3604
Matteucci M, D’Angeli S, Errico S, Lamanna R, Perrotta G, Altamura
MM (2011) Cold affects the transcription of fatty acid desatu-
rases and oil quality in the fruit of Olea europaea L. genotypes
with different cold hardiness. J Exp Bot 62:3403–3420
Miranda JA, Avonce N, Suarez R, Thevelein JM, Van Dijck P,
Iturriaga G (2007) A bifunctional TPS-TPP enzyme from yeast
confers tolerance to multiple and extreme abiotic-stress condi-
tions in transgenic Arabidopsis. Planta 226:1411–1421
Miura K, Ohta M, Nakazawa M, Ono M, Hasegawa PM (2011) ICE1
Ser403 is necessary for protein stabilization and regulation of
cold signaling and tolerance. Plant J 67:269–279
Nair S, Singh Z (2004) Chilling injury in mango fruit in relation to
biosynthesis of free polyamines. Headley, Ashford
Nakaminami K, Karlson DT, Imai R (2006) Functional conservation
of cold shock domains in bacteria and higher plants. Proc Nat
Acad Sci USA 103:10122–10127
Neill SJ, Desikan R, Clarke A, Hurst RD, Hancock JT (2002)
Hydrogen peroxide and nitric oxide as signalling molecules in
plants. J Exp Bot 53:1237–1247
Planta (2012) 235:1091–1105 1103
123

Novillo F, Medina J, Salinas J (2007) Arabidopsis CBF1 and CBF3
have a different function than CBF2 in cold acclimation and
define different gene classes in the CBF regulon. Proc Natl Acad
Sci USA 104:21002–21007
Orvar BL, Sangwan V, Omann F, Dhindsa RS (2000) Early steps in
cold sensing by plant cells: the role of actin cytoskeleton and
membrane fluidity. Plant J 23:785–794
Park MR, Yun KY, Mohanty B, Herath V, Xu F, Wijaya E, Bajic VB,
Yun SJ, De Los Reyes BG (2010) Supra-optimal expression of
the cold-regulated OsMyb4 transcription factor in transgenic rice
changes the complexity of transcriptional network with major
effects on stress tolerance and panicle development. Plant Cell
Environ 33:2209–2230
Passarini F, Wientjes E, Hienerwadel R, Croce R (2009) Molecular
basis of light harvesting and photoprotection in CP24: unique
features of the most recent antenna complex. J Biol Chem
284:29536–29546
Patton AJ, Cunningham SM, Volenec JJ, Reicher ZJ (2007)
Differences in freeze tolerance of zoysiagrasses: II. Carbohy-
drates and proline accumulation. Crop Science Society of
America, Madison
Penna S (2003) Building stress tolerance through over-producing
trehalose in transgenic plants. Trends Plant Sci 8:355–357
Pollock CJ, Lloyd EJ (1987) The effect of low temperature upon
starch, sucrose and fructan synthesis in leaves. Ann Bot
60:231–235
Pradet-Balade B, Boulme F, Beug H, Mullner EW, Garcia-Sanz JA
(2001) Translation control: bridging the gap between genomics
and proteomics? Trends Biochem Sci 26:225–229
Provart NJ, Gil P, Chen W, Han B, Chang HS, Wang X, Zhu T (2003)
Gene expression phenotypes of Arabidopsis associated with
sensitivity to low temperatures. Plant Physiol 132:893–
906
Puhakainen T, Hess MW, Makela P, Svensson J, Heino P, Palva ET
(2004) Overexpression of multiple dehydrin genes enhances
tolerance to freezing stress in Arabidopsis. Plant Mol Biol
54:743–753
Raison JK, Lyons JM (1986) Chilling injury—a plea for uniform
terminology. Plant Cell Environ 9:685–686
Ruelland E, Zachowski A (2010) How plants sense temperature.
Environ Exp Bot 69:225–232
Ruelland E, Vaultier M-N, Zachowski A, Hurry V, Kader J-C,
Delseny M (2009) Cold signalling and cold acclimation in
plants. Adv Bot Res 49:35–150
Sanders D, Pelloux J, Brownlee C, Harper JF (2002) Calcium at the
crossroads of signaling. Plant Cell 14(Suppl):S401–S417
Sangwan V, Foulds I, Singh J, Dhindsa RS (2001) Cold-activation of
Brassica napus BN115 promoter is mediated by structural
changes in membranes and cytoskeleton, and requires Ca2?
influx. Plant J 27:1–12
Sangwan V, Orvar BL, Beyerly J, Hirt H, Dhindsa RS (2002)
Opposite changes in membrane fluidity mimic cold and heat
stress activation of distinct plant MAP kinase pathways. Plant J
31:629–638
Sasaki H, Ichimura K, Oda M (1996) Changes in sugar content during
cold acclimation and deacclimation of cabbage seedlings. Ann
Bot 78:365–369
Sasaki K, Kim MH, Imai R (2007) Arabidopsis cold shock domain
protein2 is a RNA chaperone that is regulated by cold and
developmental signals. Biochem Biophys Res Commun 364:
633–638
Scott IM, Clarke SM, Wood JE, Mur LA (2004) Salicylate
accumulation inhibits growth at chilling temperature in Arabid-opsis. Plant Physiol 135:1040–1049
Seo PJ, Kim MJ, Park JY, Kim SY, Jeon J, Lee YH, Kim J, Park CM
(2010) Cold activation of a plasma membrane-tethered NAC
transcription factor induces a pathogen resistance response in
Arabidopsis. Plant J 61:661–671
Sharma N, Cram D, Huebert T, Zhou N, Parkin IA (2007) Exploiting
the wild crucifer Thlaspi arvense to identify conserved and novel
genes expressed during a plant’s response to cold stress. Plant
Mol Biol 63:171–184
Shen W, Nada K, Tachibana S (2000) Involvement of polyamines in
the chilling tolerance of cucumber cultivars. Plant Physiol 124:
431–439
Shinozaki K, Yamaguchi-Shinozaki K (2000) Molecular responses to
dehydration and low temperature: differences and cross-talk
between two stress signaling pathways. Curr Opin Plant Biol
3:217–223
Shinwari ZK, Nakashima K, Miura S, Kasuga M, Seki M, Yamaguchi-
Shinozaki K, Shinozaki K (1998) An Arabidopsis gene family
encoding DRE/CRT binding proteins involved in low-tempera-
ture-responsive gene expression. Biochem Biophys Res Commun
250:161–170
Skinner JS, von Zitzewitz J, Szucs P, Marquez-Cedillo L, Filichkin T,
Amundsen K, Stockinger EJ, Thomashow MF, Chen TH, Hayes
PM (2005) Structural, functional, and phylogenetic character-
ization of a large CBF gene family in barley. Plant Mol Biol
59:533–551
Skirvin RM, Kohler E, Steiner H, Ayers D, Laughnan A, Norton MA,
Warmund M (2000) The use of genetically engineered bacteria
to control frost on strawberries and potatoes. Whatever happened
to all of that research? Sci Hortic 84:179–189
Smallwood M, Bowles DJ (2002) Plants in a cold climate. Philos
Trans R Soc Lond B Biol Sci 357:831–846
Stitt M, Hurry V (2002) A plant for all seasons: alterations in
photosynthetic carbon metabolism during cold acclimation in
Arabidopsis. Curr Opin Plant Biol 5:199–206
Su CF, Wang YC, Hsieh TH, Lu CA, Tseng TH, Yu SM (2010) A
novel MYBS3-dependent pathway confers cold tolerance in rice.
Plant Physiol 153:145–158
Suzuki N, Koussevitzky S, Mittler R, Miller G (2011) ROS and redox
signaling in the response of plants to abiotic stress. Plant Cell
Environ 35:259–270
Svensson JT, Crosatti C, Campoli C, Bassi R, Stanca AM, Close TJ,
Cattivelli L (2006) Transcriptome analysis of cold acclimation in
barley albina and xantha mutants. Plant Physiol 141:257–270
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid.
Trends Plant Sci 15:89–97
Tabaei-Aghdaei SR, Pearce RS, Harrison P (2003) Sugars regulate
cold-induced gene expression and freezing-tolerance in barley
cell cultures. J Exp Bot 54:1565–1575
Takuhara Y, Kobayashi M, Suzuki S (2011) Low-temperature-
induced transcription factors in grapevine enhance cold tolerance
in transgenic Arabidopsis plants. J Plant Physiol 168:967–975
Theocharis A, Bordiec S, Fernandez O, Paquis S, Dhondt-Cordelier S,
Baillieul F, Clement C, Ait Barka E (2011) Burkholderiaphytofirmans strain PsJN primes Vitis vinifera L. and confers a
better tolerance to low non-freezing temperatures. Mol Plant
Microbe Interact 25:241–249
Thomashow MF (2010) Molecular basis of plant cold acclimation:
insights gained from studying the CBF cold response pathway.
Plant Physiol 154:571–577
Tian Q, Stepaniants SB, Mao M, Weng L, Feetham MC, Doyle MJ,
Yi EC, Dai H, Thorsson V, Eng J, Goodlett D, Berger JP, Gunter
B, Linseley PS, Stoughton RB, Aebersold R, Collins SJ, Hanlon
WA, Hood LE (2004) Integrated genomic and proteomic
analyses of gene expression in mammalian cells. Mol Cell
Proteomics 3:960–969
Uemura M, Steponkus PL (1999) Cold acclimation in plants:
relationship between the lipid composition and the cryostability
of the plasma membrane. J Plant Res 112:245–254
1104 Planta (2012) 235:1091–1105
123

Uemura M, Warren G, Steponkus PL (2003) Freezing sensitivity in
the sfr4 mutant of Arabidopsis is due to low sugar content and is
manifested by loss of osmotic responsiveness. Plant Physiol
131:1800–1807
Van Loon LC, Van Strien EA (1999) The families of pathogenesis-
related proteins, their activities, and comparative analysis of
PR-1 type proteins. Physiol Mol Plant Pathol 55:85–97
Venketesh S, Dayananda C (2008) Properties, potentials, and
prospects of antifreeze proteins. Crit Rev Biotechnol 28:57–82
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a
review. Amino Acids 35:753–759
Vogel JT, Zarka DG, Van Buskirk HA, Fowler SG, Thomashow MF
(2005) Roles of the CBF2 and ZAT12 transcription factors in
configuring the low temperature transcriptome of Arabidopsis.
Plant J 41:195–211
Vogg G, Heim R, Gotschy B, Beck E, Hansen J (1998) Frost
hardening and photosynthetic performance of Scots pine (Pinussylvestris L.). II. Seasonal changes in the fluidity of thylakoid
membranes. Planta 204:201–206
Wang R, Li R, Sun Z, Ren Y, Yue W (2006) Anti-freezing proteins
and plant responses to low temperature stress. Ying Yong Sheng
Tai Xue Bao 17:551–556
Welling A, Palva ET (2006) Molecular control of cold acclimation in
trees. Physiol Plant 127:167–181
Xia XJ, Wang YJ, Zhou YH, Tao Y, Mao WH, Shi K, Asami T, Chen
Z, Yu JQ (2009) Reactive oxygen species are involved in
brassinosteroid-induced stress tolerance in cucumber. Plant
Physiol 150:801–814
Xiao H, Siddiqua M, Braybrook S, Nassuth A (2006) Three grape
CBF/DREB1 genes respond to low temperature, drought and
abscisic acid. Plant Cell Environ 29:1410–1421
Xiao H, Tattersall EA, Siddiqua MK, Cramer GR, Nassuth A (2008)
CBF4 is a unique member of the CBF transcription factor family
of Vitis vinifera and Vitis riparia. Plant Cell Environ 31:1–10
Xin Z, Browse J (2000) Cold comfort farm: the acclimation of plants
to freezing temperatures. Plant Cell Environ 23:893–902
Xin Z, Li PH (1993) Relationship between proline and abscisic acid in
the induction of chilling tolerance in maize suspension-cultured
cells. Plant Physiol 103:607–613
Yaish MW, Doxey AC, McConkey BJ, Moffatt BA, Griffith M (2006)
Cold-active winter rye glucanases with ice-binding capacity.
Plant Physiol 141:1459–1472
Yamaguchi-Shinozaki K, Shinozaki K (2006) Transcriptional regu-
latory networks in cellular responses and tolerance to dehydra-
tion and cold stresses. Annu Rev Plant Biol 57:781–803
Yang T, Chaudhuri S, Yang L, Du L, Poovaiah BW (2010) A
calcium/calmodulin-regulated member of the receptor-like
kinase family confers cold tolerance in plants. J Biol Chem
285:7119–7126
Yeh S, Moffatt BA, Griffith M, Xiong F, Yang DS, Wiseman SB,
Sarhan F, Danyluk J, Xue YQ, Hew CL, Doherty-Kirby A,
Lajoie G (2000) Chitinase genes responsive to cold encode
antifreeze proteins in winter cereals. Plant Physiol 124:1251–
1264
Zeng Y, Yu J, Cang J, Liu L, Mu Y, Wang J, Zhang D (2011)
Detection of sugar accumulation and expression levels of
correlative key enzymes in winter wheat (Triticum aestivum) at
low temperatures. Biosci Biotechnol Biochem 75:681–687
Zhai H, Bai X, Zhu Y, Li Y, Cai H, Ji W, Ji Z, Liu X, Li J (2010) A
single-repeat R3-MYB transcription factor MYBC1 negatively
regulates freezing tolerance in Arabidopsis. Biochem Biophys
Res Commun 394:1018–1023
Zhang S, Jiang H, Peng S, Korpelainen H, Li C (2011) Sex-related
differences in morphological, physiological, and ultrastructural
responses of Populus cathayana to chilling. J Exp Bot 62:675–
686
Zhou X, Wang G, Sutoh K, Zhu JK, Zhang W (2008) Identification of
cold-inducible microRNAs in plants by transcriptome analysis.
Biochim Biophys Acta 1779:780–788
Zhou MQ, Shen C, Wu LH, Tang KX, Lin J (2011) CBF-dependent
signaling pathway: a key responder to low temperature stress in
plants. Crit Rev Biotechnol 31:186–192
Zhu JH, Dong CH, Zhu JK (2007) Interplay between cold-responsive
gene regulation, metabolism and RNA processing during plant
cold acclimation. Curr Opin Plant Biol 10:290–295
Zhu J, Jeong JC, Zhu Y, Sokolchik I, Miyazaki S, Zhu JK, Hasegawa
PM, Bohnert HJ, Shi H, Yun DJ, Bressan RA (2008) Involve-
ment of Arabidopsis HOS15 in histone deacetylation and cold
tolerance. Proc Natl Acad Sci USA 105:4945–4950
Planta (2012) 235:1091–1105 1105
123





























