Embed Size (px)
Citation preview

Vol. 265, No. 23, Issue of August 15, pp. 14023-14029,199O Printed in U.S.A.
Physicochemical Properties of the Lipopolysaccharide Unit That Activates B Lymphocytes*
(Received for publication, February 14, 1990)
Kuni Takayama$$, Zafeer Z. Din+, Pasupati Mukerjeen, Peter H. CookelI**, and Theo N. Kirkland** From the $Mycobacteriology Research Laboratory, William S. Middleton Memorial Veterans Hospital, Madison, Wisconsin 53705, Department of Bacteriology, College of Agricultural and Life Sciences, KS’chool of Pharmacy, and (Jlntegrated Microscopy Resources for Biomedical Research, University of Wisconsin, Madison, Wisconsin 53706, and the @Division of Infectious Diseases, Veterans Administration Medical Center and Department of Pathology and Medicine, University of California, San Diego, California 92161
We have examined the physical state of highly puri- fied deep rough chemotype lipopolysaccharide (ReLPS) from Escherichia coli D31m4 as an aqueous suspension and as complexes with bovine serum albu- min (BSA). The ReLPS suspension showed large ellip- soidal particles 12-38 nm wide and 40-100 nm long. The solubility of this form of ReLPS was determined by equilibrium dialysis experiments to be 3.3 X lo-’ M at 22 “C and 2.8 x lo-’ M at 37 “C in 150 mM Tris- KCl, pH 7.5; 3.0 x lOwe M at 37 “C in 0.75 mM Tris- KCl, pH 7.5.
The BSA-ReLPS complexes were fractionated on a Sephacryl S-200 column to yield peaks I and II with apparent masses of about 240 and 70 kDa, respec- tively. Peak II was a BSA monomer with estimated BSA:ReLPS molar ratios of l:l-1:7. The ReLPS sus- pension and the two complexes were compared as an- tigens in enzyme-linked immunosorbent assays using three select monoclonal antibodies to lipopolysaccha- ride. The results were consistent with the high state of disaggregation of the ReLPS in both peaks I and II. Since the ReLPS in these complexes were not visible by electron microscopy, they did not contain vesicles or large particles.
All forms of ReLPS tested were capable of stimulat- ing 70213, a lipopolysaccharide-responsive murine pre-B cell line. However, peak II was consistently more stimulatory at very low concentrations than the other preparations. The maximally stimulatory concentra- tion of ReLPS for 7OZ/3 cells was 40 rig/ml (1.6 x lo-’ M) for peak II and 70 rig/ml (2.8 X lo-’ M) for the ReLPS suspension. As expected, the above concentra- tions were at or below the solubility of the ReLPS. These results suggested that the highly disaggregated form of ReLPS (possibly the monomer) is the active unit that stimulates the cellular response in 7OZ/3 cells.
* This work was supported in part by the Medical Research Service of the Department of Veterans Affairs and by Grant GM-36054 from the National Institutes of Health. The Integrated Microscopy Re- sources for Biomedical Research is supported by National Institutes of Health Grant RR-570. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC. Section 1734 solely to indicate this fact.
§ To whom correspondence should be addressed: Mycobacteriology Research Laboratorv, William S. Middleton Memorial Veterans Hos- pital, 2500 OverlookTerrace, Madison, WI 53705. Tel.: 608-256-1901 (ext. 7812).
** Present address: EM Center, USDA-ARS-ERCC, 600 E. Mer- maid Ln., Philadelphia, PA 19118.
Since lipopolysaccharide (LPS)’ is an amphipathic macro- molecule, it is highly aggregated in an aqueous preparation (I-5). This is due to a compromise between the tendency of the six fatty acyi groups on a single lipid A moiety to avoid contact with water and a strong affinity of the carbohydrate and phosphate groups for water. An important question has remained unanswered regarding the basic biology of LPS action. How does an aggregated LPS interact with responding cells (B cells and macrophages, among others) to elicit their numerous biological activities under physiological conditions? More specifically, what is the active LPS unit? An answer to this question is needed to determine whether there is a phys- iological receptor for LPS in responding cells and what the nature of this receptor is, if it exists. A study of the structure- to-function relationship requires knowledge of the active LPS unit. At present it is difficult to answer this question on the basis of the stoichiometric ligand-to-receptor interaction be- cause the ligand in this case is highly water-insoluble. How- ever, we can consider three possibilities: (i) that LPS yields a small but finite concentration of monomeric LPS molecules that serve as the ligand; (ii) that the LPS aggregate serves as the ligand; and (iii) that the LPS aggregate or monomer is internalized by the responding cells and then transported to specific sites inside the cells for its interaction (6). This is an alternative to cell surface interaction. This problem can only be studied by using a highly purified and well characterized LPS preparation. The ReLPS from Escherichia coli D31m4 is now available and is suitable for such a study (7).
Serum albumin binds long-chain fatty acids (B-12) and phospholipids (13, 14), which suggests that it can also bind LPS and lipid A. Thus Galanos et al. (15,16) tested and found that BSA binds and “solubilizes” lipid A in aqueous medium. Although they did not study the physical nature of the BSA- lipid A interaction, it appeared likely that the BSA actually reduced the state of aggregation of the lipid A suspension, possibly to the monomeric state. This disaggregating property of BSA would be useful in the study of the nature of the active LPS unit.
We have examined some of the important physicochemical properties of the LPS that interact with B cells and activate them. For this study we used the ReLPS obtained from E. coli in the form of BSA.ReLPS complexes. This complex mixture was purified by column chromatography and char- acterized by using selective mAb against different epitopes on the ReLPS molecule. By complexing with BSA, we were able
’ The abbreviations used are: LPS, lipopolysaccharide; ReLPS, deep rough chemotype lipopolysaccharide; BSA, bovine serum albu- min; mAb, monoclonal antibody; KDO, 2-keto-3-deoxyoctonate; CMC, critical micelle concentration.
14023

14024 Properties of the Active LPS Unit
to obtain ReLPS preparations in highly disaggregated forms. We studied the physical nature of the ReLPS particles and the BSA.ReLPS complex fractions, and how well they were able to activate the 7OZ/3 cells. We suggest that these acti- vations occurred by the direct interaction of the highly dis- aggregated form of ReLPS with the 7OZ/3 cells. We have now established the solubility of the simplest of LPS, the hexaacyl ReLPS from E. coli.
EXPERIMENTAL PROCEDURES’
RESULTS
Determination of Solubility of ReLPS-When the donor cell of an equilibrium dialysis apparatus containing 82.5 pM [“‘Cl ReLPS in 50 mM Tris-HCl, pH 7.5,lOO mM KC1 was allowed to operate at 22 “C, equilibrium was reached in the dialysate in 24 h at 3.3 X lOma M (Fig. 2).” We believe this represents the solubility of the ReLPS with the structure shown in Fig. 1.
This experiment was performed in two different ways. In one series, the dialysate side was sampled successively with increasing time at 2, 6, 15, 17, and 24 h. In another series, sampling was done successively at 24 and 48 h, and then at 2, 6, 18, and 72 h. Sampling involved the removal of the entire volume, replacement with fresh buffer, and continuation of the experiment. The donor side and membrane remained the same in each series. We found a time-dependent increase in the [YJReLPS on the dialysate side, regardless of whether sampling was done as early as 3-24 h or as late as 7 days in a discontinuous manner (Fig. 2). It reached equilibrium in about 24 h at the same concentration of 3.3 x 10TR M. At the end of the experiment (7 days), the following distribution of [‘“Cl ReLPS was determined for the donor side: bound to mem- brane, 3% (2.5 nmo1/4.5 cm*); bound to Teflon housing, 60% (49.5 nmol); recoverable, 35% (28.9 nmol); lost, 2% (1.7 nmol). Although “sticky,” the [i4C]ReLPS did not clog the membrane pores to the extent that it interfered with diffusion over the entire period of the experiment. The [Y]ReLPS remained intact during this period.4 At lower concentrations of ReLPS it took longer to reach this equilibrium concentration. For example, equilibrium was expected to be reached in about 4 days at 41.3 pM [“C]ReLPS and 6 days at 27.5 pM. When we used the high concentration of [“‘C]ReLPS with the culture medium for the 7OZ/3 cell activation assay, we obtained identical results at 36, 48, and 60 h: 3.2 X lo-‘, 3.3 X 10e8, and 3.2 X lo-* M, respectively.
A time course of solubilization of an 82.5 pM suspension of [Y]ReLPS in 0.25 mM Tris-HCl, pH 7.5, 0.50 mM KC1 at 37 “C showed that equilibrium is reached in about 4 h (Fig. 2). The average value of nine points past 4 h in three experi- ments was 3.0 + 0.3 X lOma M. The solubility of [“C]ReLPS in 50 mM Tris-HCl, pH 7.5, 100 mM KC1 at 37 “C was 2.8 X
lOma M. The radiolabeled material in the dialysate side was shown to be [Y]ReLPS by thin layer chromatography and radio high performance liquid chromatography. This repre-
? Portions of this paper (“Experimental Procedures,” Figs. 3-6, and Tables I and II) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
a For reasons discussed later, we believe that this equilibrium dialysate concentration represents the saturation solubility of ReLPS rather than its CMC (27, 28).
4 The [‘“C]ReLPS is adequately stable for the equilibrium dialysis experiment. This compares with the [“Clmonophosphoryl and [‘“Cl bisphosphoryl lipid As which release fatty acids, thus masking the solubility determination of lipid A.
FIG. 1. Structure of the hexaacyl ReLPS from E. coli D31m4, i%f, 2238, as the free acid. A, chemical structure. B, space-filling molecular model. The right side is the reducing end of the GlcN disaccharide unit. The KDO disaccharide is situated at the upper left corner; it projects away from and perpendicular to the plane of the picture, Two of the six fatty acyl groups are behind the two shown at the left side. Elemental color code: carbon, black; hydrogen, white; oxygen, red, nitrogen, blue; phosphorus, yellow.
sents the first successful attempt to determine the solubility of an LPS.
Preparation and Fractionation of BSA . ReLPS Complex Mixture-BSA. ReLPS complex mixture was prepared by the method of Appelmelk et al. (24) with one modification: after the BSA solution was mixed with the ReLPS suspension, the

Properties of the Active LPS Unit 14025
4- I I I
l l
I I 12 24
Time (hours)
I
48
;,
72
FIG. 2. Time course of dialysis of an 82.5 pM [“C]ReLPS suspension. Left panel, in 0.25 mM Tris-HCl, pH 7.5, 0.50 mM KC1 at 37 “C (A); in 50 mM Tris-HCl, pH 7.5, 100 &M KC1 at 37 “C (A). R&t nanel. in 50 mM Tris-HCl. DH 7.5. 100 mM KC1 at 22 “C. Samplkg was performed in sequenGa1 order (0) or in discontinuous manner as indicated by the numbers in parentheses (0). The equilib- rium dialysis apparatus was used.
combination was sonicated for 15 min at 22 “C to achieve maximal formation of the complexes. Such a mixture had a BSA:ReLPS dry weight ratio of 1O:l (molar ratio, 1:2.8). This preparation was dissolved in 1.0 ml of 50 mM Tris-HCl, pH 7.5, 100 mM KC1 at a concentration of 10 mg of BSA/ml and applied to a short (3.2 X 15 cm) Sephacryl S-200 column for the initial fractionation. The column was eluted with the same Tris-HCl buffer, and l.O-ml fractions were collected. The protein profile of such a fractionation is shown in Fig. 3A. Effluent volume fractions 40-56 ml (crude peak I) and 57-80 ml (crude peak II) were pooled and lyophilized. Then each pooled fraction was dissolved in 2.0 ml of water and fractionated for the second time on a long (1.6 x 90 cm) Sephacryl S-200 column. The results of this fractionation are shown in Fig. 3, B and C. Effluent volume fractions 68-84 ml (purified peak I) and 85-97 ml (purified peak II) were pooled and analyzed for BSA and ReLPS content by measuring protein and total phosphorus (Table I). Such preparations were characterized for their ability to bind mAb and then used to activate 7OZ/3 cells.
The Sephacryl S-200 columns used in these fractionations required preconditioning. The column was charged with at least 3 aliquots of BSA (1 ml each of a 10 mg/ml solution) and washed with 10 bed volumes of the Tris-KC1 buffer. Then 2.0 ml of the BSA. ReLPS preparation containing 10 mg of BSA and 1 mg of ReLPS/ml was placed on the column and washed with the same buffer.
When the effluent volume of BSA (10 mg) was determined with a Sephacryl S-200 column, both absorbance at 280 nm and total phosphorus content (for LPS) were determined. The BSA control was essentially devoid of the BSA. LPS complex.
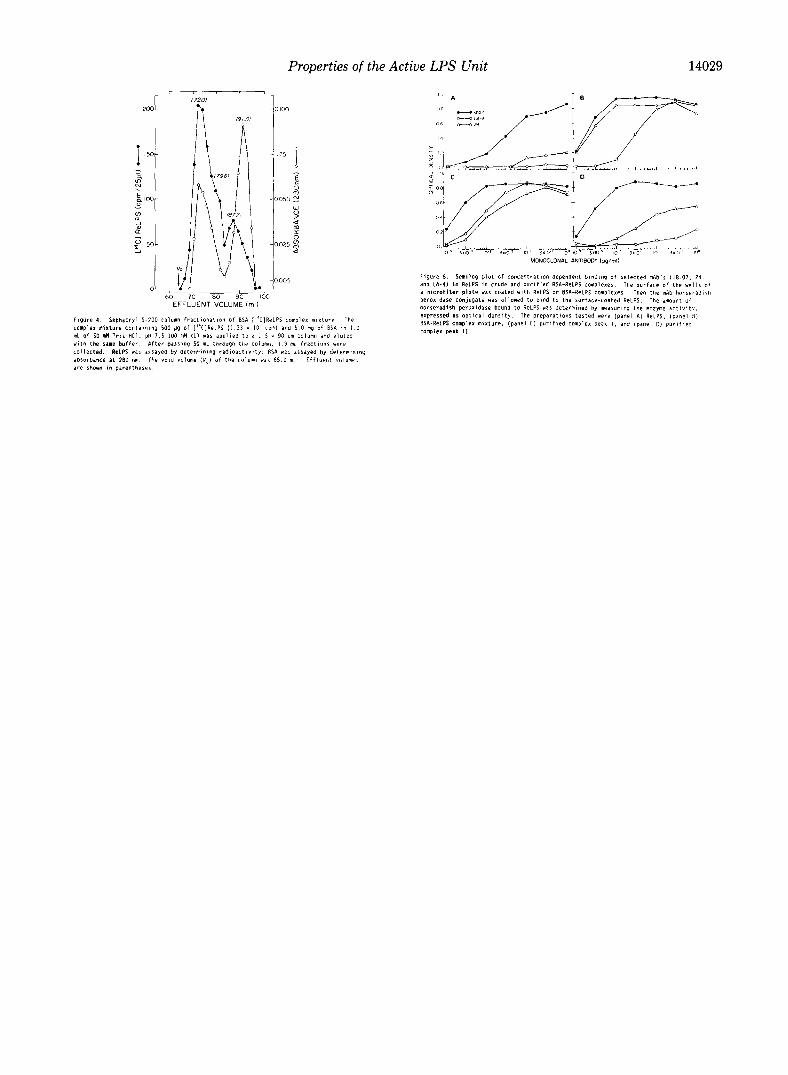
Fractionation and Characterization of BSA . [‘T]ReLPS Complexes-The BSA . [ 14C]ReLPS complex mixture was pre- pared and fractionated on a Sephacryl S-200 column as shown in Fig. 4. Select column fractions were analyzed for size of the complexes and BSA and [i4C]ReLPS contents were deter- mined (Table II). About 25% of the [14C]ReLPS was not complexed, and thus adsorbed to the Sephacryl column. All of the BSA and the remaining 75% of the [Y]ReLPS applied to the column eluted within the void volume (V, = 65.0 ml) and the effluent volume of the BSA (VBS* = 91.0 ml). Two BSA-positive peaks (absorbance at 280 nm) were observed at
72.0 and 91.0 ml effluent volumes. The [i4C]ReLPS gave a large peak at 72.0 ml, a shoulder at 79.6 ml, and a small peak at 87.2 ml effluent volumes. The size of the BSA. [“C]ReLPS complexes ranged from just above 66 kDa to >240 kDa. The large BSA. [‘4C]ReLPS complexes in fractions at effluent volumes 72.0 and 73.9 ml (Table II) gave sizes of 240 and 220 kDa, with BSA:[14C]ReLPS molar ratios of 1.0:7.4 and 1.0:7.3, respectively. These two peaks could represent the BSA trimer containing about 22 [“‘C]ReLPS molecules. The shoulder fraction at 79.6 ml could represent the [i4C]ReLPS-containing BSA dimer and trimer mixture. According to the previous designation (Fig. 3), peak I would be pooled effluent volumes 70-83 ml, and peak II would be pooled effluent volumes 84- 94 ml.
The BSA and [i4C]ReLPS plots in peak I were similar, whereas the [Y]ReLPS plot in peak II showed that the peak was offset to the larger sizes over the BSA peak (Fig. 4). This indicated that there is an excess of free BSA in peak II. Thus, a determination of the BSA:[‘4C]ReLPS molar ratio would be misleading. We conclude that peak II consists of free BSA and a mixture of monomeric BSA. [‘“CJReLPS complex with a range of BSA:[14C]ReLPS molar ratios of 1:l to about 1:7.
The BSA. [14C]ReLPS complexes were also prepared with fatty acid-depleted BSA, fractionated by column chromatog- raphy, and analyzed (data not presented). The monomeric peak II was the major fraction. The minor oligomeric BSA fraction (peak I) had a BSA:[14C]ReLPS ratio on the order of l:l-1:2, which was different from the previously described preparation of peak I. Native BSA was used in this study because it would resemble the BSA found naturally under physiological conditions.
Electron Microscopy of BSA . ReLPS Complexes-Nega- tively stained samples of the ReLPS suspension and the BSA. ReLPS complexes were examined electron microscopically (Fig. 5). We needed to analyze thick samples at protein concentrations up to 10 mg/ml to visualize more clearly the smaller particles of ReLPS in BSA. Therefore, we used the high-voltage electron microscope operating at an accelerating voltage of 1 MeV. Fig. 5A shows that, as expected, the ReLPS suspension is composed of large ellipsoidal particles 12-38 nm
FIG. 5. Electron micrographs of negatively stained ReLPS from E. coli D31m4 and its complexes with BSA. A, ReLPS in 50 mM Tris-HCl, pH 7.5, 100 mM KCl; B, BSA mixed with ReLPS (lO:l, w/w); C, peak I from Sephacryl S-200 column; D, peak II from Sephacryl S-200 column. The occasional particles seen in D represent trace amounts of those seen in B, which presumably eluted from the column. Magnification X 30,000; bar equals 100 nm.

14026 Properties of the Active LPS Unit
OoL 20 40 60 80 ReLPS (rig/ml)
FIG. 7. Activation of 70213 pre-B cells by ReLPS suspen- sion and purified BSA.ReLPS complexes in the absence of serum. 7OZ/3 cells were incubated for 40 h with various concentra- tions of ReLPS. and then stained with fluoresceinated anti-IeM antibody and analyzed by cytofluorometry. The percent of exposed cells induced to become IgM-positive was calculated as percent of fluorescent cells minus background fluorescence in unexposed cells. This was plotted against ReLPS concentration.
wide and 40-100 nm long. When the ReLPS was combined with BSA, the large particles were absent, and in their place were many small particles with a fairly uniform size of about 8 x 20 nm (Fig. 5B). These particles were smaller than the trilamellar discs described by Shands (l), but could represent a shortened form of the lo-nm diameter tubular or rod-shaped micelles described by Hayter et al. (5). Fig. 5, C and D, shows that the Sephacryl S-200 column fractions (peaks I and II) were essentially devoid of the 8 X 20-nm particles. They could represent the 25% of the ReLPS lost during the column fractionation. Peaks I and II showed little more than the BSA background observed in control preparations. These and other results suggest that the BSA . ReLPS mixture contains at least three physical forms of ReLPS: the 8 X 20-nm particles, peak I and peak II.
Inability of BSA f ReLPS Complexes to Entrap Small Mole- cr.&s-Nixdorff et al. (29) showed that like phospholipids (30), vesicles made with LPS can entrap water-soluble mole- cules. We tested the BSA. ReLPS complex mixture for this ability. For this experiment, 5.7 nmol of [U-“‘Clglucose (0.5 &i) was added to 10.0 mg of BSA and 1.0 mg of ReLPS in the preparation of the complex mixture. One milliliter of this was sonicated and immediately chromatographed on the 2.3 x E-cm Sephacryl S-200 column (data not presented). Prac- tically all of the [‘4C]glucose (5.6 nmol, 98%) eluted from the column at 104 ml effluent volume. Within the BSA.ReLPS complex peak fractions (40-73 ml effluent volume), only a trace amount (less than 0.1 nmol) of the [‘4C]glucose was detected and the radioactivity profile was slightly above base line. These results show that the BSA.ReLPS complexes do not have structures that can entrap small molecules. They support the hypothesis that the ReLPS molecules in the BSA. ReLPS complexes are highly disaggregated.
Accessibility of ReLPS to mAb in BSA . ReLPS Complexes- We wanted to determine the degree of disaggregation of the ReLPS in the BSA.ReLPS complexes based on the accessi- bility of the bound ReLPS to several key mAbs. We chose three mAbs: 18.07 (31), which has high avidity for the KDO region of ReLPS; 24 (31), with low avidity for the lipid A region of ReLPS; and LA-4 (32), with very high avidity for the lipid A region of ReLPS. The enzyme-linked immunosor- bent assay of Appelmelk et al. (24) was used. This procedure involves binding the ReLPS to a plastic surface. We found that the [i4C]ReLPS in all forms (suspension and BSA com- plexes) bound equally well to plastic. The ReLPS suspension, BSA . ReLPS complex mixture, and purified BSA. ReLPS complex fractions I and II were tested for binding by the
mAb. The results of one such study are shown in Fig. 6. The mAb 18.07 bound to uncomplexed ReLPS and all forms
of the BSA complexes, suggesting that the KDO region was exposed in all preparations. The minimum concentration of mAb 18.07 needed for detectable binding to the BSA com- plexes was slightly lower than that for binding to the uncom- plexed ReLPS, indicating that the BSA complex did allow more exposure of the KDO region than uncomplexed ReLPS. However, this was minor compared to the results of anti-lipid A antibody binding. Neither mAb 24 nor LA-4 bound to the uncomplexed ReLPS, suggesting that the lipid A regions was not exposed (Fig. 6A). Both anti-lipid A mAbs bound well to peak I (Fig. 6C) and significantly less to peak II (Fig. 6D). This suggests that the lipid A region of ReLPS is less acces- sible in peak II than in peak I and (or) that the multivalent antibody binding occurs to peak I but not to peak II. We favor the former interpretation because we know that LA-4 has a higher affinity than mAb 24 for ReLPS (31), but LA-4 binds peak II less well than mAb 24. In addition, LA-4 binds to the lipid region of lipid A, whereas mAb 24 appears to bind to the 1,4’-bisphosphate disaccharide unit (31), so that LA-4 might be less able to bind to lipid that is tightly bound to BSA.
Activation of 70213 Cells by ReLPS-Results of the concen- tration-dependent activation of 7OZ/3 cells by several ReLPS preparations in the absence of serum are shown in Fig. 7. Peak II was much more active than either ReLPS suspension or peak I. This activation was characterized by a sharp rise at the low concentration range of 2-8 rig/ml ReLPS (0.8-3.2 X lo-’ M). Maximal activation with peak II occurred at about 40 rig/ml ReLPS (1.6 X lo-* M). Peak II was also the most active at low concentrations in the B lymphocyte proliferation assay in the absence of serum (data not presented). The three forms of ReLPS gave identical dose-response curves in the presence of 5% fetal calf serum, with maximal activation at about 70 rig/ml (2.8 x 1O-8 M) (data not presented).
DISCUSSION
In this study, we wanted to answer a basic question about the nature of the active LPS unit. How can an aggregated LPS initiate the numerous biological responses in the re- sponding cells? To study this problem we first had to develop a highly disaggregated form of the LPS without resorting to either derivatization or covalent attachment to a water-solu- ble carrier. We prepared and studied the physical properties of the noncovalent BSA + ReLPS complexes designated I and II. Peak II contained the highly disaggregated form of ReLPS, which was precisely what we needed for our study.
We expected the state of aggregation of ReLPS to greatly influence antibody binding. In particular, we predicted that antibody directed to the lipid A moiety would bind disaggre- gated LPS more readily than aggregated LPS. We found this to be true. Two anti-lipid A mAb (24 and LA-4) bound to BSA.ReLPS complexes much better than to particles of ReLPS. We also found the lipid A region of the ReLPS in peak II to be less available for interaction with mAbs LA-4 and 24 than that in peak I.
We then tested the ReLPS suspension and BSA complexes for activation of 7OZ/3 cells. If the biologically active ReLPS is the aggregate, we predicted that peak II would be inactive. This was not the case. At the lower concentrations (4-20 ng/ ml) and in the absence of serum, peak II activated 7OZ/3 cells to a greater extent than the ReLPS suspension or peak I. Thus, BSA appeared to allow the ReLPS in peak II to be delivered to 7OZ/3 cells more effectively than the ReLPS that was added to the cell system by serial dilution of a suspension.
To understand the states of ReLPS molecules in solution

Properties of the Active LPS Unit 14027
or dispersion, the equilibrium dialysate concentration of ReLPS should be established as a measure of the solubility of ReLPS, rather than the CMC. A study of the dialysis behavior of sodium dodecyl sulfate, a well known micellar system (27), is relevant here. With membranes permeable to monomers but not to micelles, the dialysis rate of monomers from a high donor-side concentration (well above the CMC) was slower when the dialysate concentration was >CMC than <CMC. Nevertheless, micelles (which are in dynamic equilibrium with monomers (33, 34)), formed in the dialysate when the CMC was exceeded. At equilibrium the donor and dialysate sides achieved equal concentrations of monomers and micelles. In our ReLPS system, in contrast, the equilibrium dialysate concentration of 3.3 X 10e8 M remains only 0.04% of the donor-side concentration, indicating that a solubility limit has been reached and the donor compartment contains solid solute. That this low solubility is primarily due to monomers and not micelles is indicated by the very slight effect of temperature (22 uersus 37 “C) on equilibrium dialysate con- centrations. The solubility of micelle-forming electrolytes above the Krafft temperature increases rapidly with temper- ature when the solubility exceeds the CMC (35). For example, the solubility of sodium decyl sulfonate increases 60-fold as the temperature increases from 23 to 37 “C (35). With ReLPS, the solubility in 50 mM Tris-HCl, pH 7.5,lOO mM KC1 changes from 3.3 X lo-’ M at 22 “C to the slightly lower value of 2.8 X 10m8 M at 37 “C. It is also noteworthy that at 37 “C the solubility change is minimal when the medium is changed from 0.25 mM Tris-HCl, pH 7.5, 0.5 mM KC1 (solubility 3.0 X 10e8 M) to 50 mM Tris-HCl, pH 7.5, 100 mM KC1 (solubility 2.8 X lo-’ M). In ionic micellar systems, the effect of counter- ions on monomer-micelle equilibria would be expected to produce major changes in solubility above the CMC (36). The solubility we observe is most likely due to dissociation of the free acid form of ReLPS from the solid and ionization to anions. These processes are not expected to be strongly de- pendent on counterion concentration.
The minor effects of changes in temperature and counterion concentrations provide evidence that free ReLPS molecules in solution are mostly monomeric. When ReLPS concentra- tion exceeds the low solubility, ReLPS appears to exist as a microsuspension at 22-37 “C! with no micelles present. The available monomer concentration (which corresponds to the solubility) is much less than the CMC of lipid X, a monosac- charide precursor of lipid A, at pH 8.2. The CMC of lipid X was 4 x 10m5 M (37). The lower solubility of ReLPS can be ascribed to ReLPS having six fatty acyl groups (Fig. l), whereas lipid X has only two. The space-filling model of ReLPS (Fig. 1B) emphasizes the predominance of hydropho- bic groups over hydrophilic. We believe that the distinction between a microsuspension and a micellar system (28), which is often difficult, is important for many lipid systems with low solubility.
The slow rate of equilibration in dialysis at 22 “C probably results in part from the microsuspension’s slow rate of disso- lution. The more rapid rate at 37 “C can be ascribed to a lower viscosity and, possibly, to higher rates of dissolution and membrane permeation caused by a higher rate of agitation at the higher temperature.
This study, suggesting that monomeric LPS is the basic unit that activates B lymphocytes, establishes a basis for studying the receptor-to-LPS or -lipid A interaction and the structure-to-biological function relationship.
Acknowledgments-We thank Todd Sievert for growing the cells and extracting the radiolabeled ReLPS and Nilofer Qureshi for as- sistance in purifying the ReLPS in the initial phase of the study.
5.
6. 7.
8. 9.
10.
11.
12.
13. 14.
15.
16.
17. 18. 19.
20.
21.
22.
23.
24.
25.
26. 27.
28.
29.
30. 31.
32.
33. 34.
REFERENCES
Shands, J. W., Jr., Graham, J. A., and Nath, K. (1967) J. Mol. Biol. 25, 15-21
Emmerling, G., Henning, U., and Gulik-Krzywicki, T. (1977) Eur. J. Biochem. 78,503-509
Van Aluhen. L.. Verkleii. A.. Burnell, E., and Luatenbera. B. (198Oj Biochik Bioph& A& 597, 502-517 - -’
Labischinski H.. Barnickel. G.. Bradaczek. H.. Naumann. D.. Rietschel, E. T., and Giesbrecht, P. (1985) J.‘Bacteriol. i62; 9-20
Hayter, J. B., Rivera, M., and McGroarty, E. J. (1987) J. Biol. Chem. 262,5100-5105
Bradley, S. G. (1979) Annu. Reu. Microbial. 3,67-94 Qureshi, N., Takayama, K., Mascagni, P., Honovich, J., Wong,
R., and Cotter, R. J. (1988) J. Biol. Chem. 263, 11971-11976 Saifer, A., and Goldman, L. (1961) J. Lipid Res. 2, 268-270 Reynolds, J., Herbert, S., and Steinhardt, J. (1968) Biochemistry
7,1357-1361 Spector, A. A., John, K., and Fletcher, J. E. (1969) J. Lipid Res.
10,56-67 Reed, R. G., Feldhoff, R. C., Clute, 0. L., and Peters, T., Jr.
(1975)Biochemistr-y 14,4578-4583 Peters. T.. Jr. (1975) in The Plasma Proteins (Putnam, F. W..
ed) pp. i33-181, Academic Press, New York Klonfenstein. W. E. (1969) Biochim. Bioohvs. Acta 181.323-325 Nakagawa, M., and Nishida, T. (1973) 3. &o&m. (Tokyo) 74,
1263-1266 Galanos, C., Rietschel, E. T., Liideritz, O., and Westphal, 0.
(1971) Eur. J. Biochem. 19, 143-152 Galanos, C., Rietschel, E. T., Luderitz, O., and Westphal, 0.
(1972) Eur. J. Biochem. 31, 230-233 Bartlett, G. R. (1959) J. Biol. Chem. 234, 466-471 Osborn, M. J. (1963) Proc. Nat/. Acad. Sci. U. S. A. 50,499-506 Takayama, K., Qureshi, N., Mascagni, P., Nashed, M. A., Ander-
son, L., and Raetz, C. R. H. (1983) J. Biol. Chem. 258, 7379- 7385
Galanos, C., Liideritz, O., and Westphal, 0. (1969) Eur. J. Biochem. 9, 245-249
Qureshi, N., Takayama, K., Heller, D., and Fenselau, C. (1983) J. Biol. Chem. 258, 12947-12851
Qureshi, N., Cotter, R. J., and Takayama, K. (1986) J. Microbial. Methods 5. 65-77
Bligh, E. G., and Dyer, W. J. (1959) Can. J. Biochem. Physiol. 37,911-917
Apnelmelk. B. J., Verweii-Van Vuaht. A. M. J. J.. MacLaren. D. -M., and Thijs, L. G. (1585) J. Zim&ol. Metho& 82, 199-iO7
Paine. C. J.. Kincade. P. W.. and Ralnh. P. (1978) J. Zmmunol. l>i,6411647 * ' . '
Mosier, D. E. (1981) J. Immunol. 127, 1490-1493 Mysels, K. J., Mukerjee, P., and Abu-Hamdiyyah, M. (1963) J.
Phys. Chem. 67, 1943-1944 Mysels, K. J., and Mukerjee, P. (1979) Pure Appl. Chem. 51,
1083-1089 Nixdorff, K., Gmeiner J., and Martin, H. H. (1978) Biochim.
Biophys. Actu 510,87-98 Papahadjopoulos, D. (1978) Ann. N. Y. Acod. Sci. 308, l-462 Kirkland, T. N., Ziegler, E. J., Tobias, P., Ward, D. C., Michalek,
S. M., McGhee, J. R., Macher, I., Urayama, K., and Appelmelk, B. J. (1988) J. Zmmunol. 141.3208-3213
Kirkland, T. N., Colwell, D. E., Michalek, S. M., McGhee, J. R., and Ziegler, E. J. (1986) J. Zmmurwl. 137, 3614-3619
Mukerjee, P. (1967) Adu. Colloid Interface Sci. 1, 241-275 Lindman B., and Wennerstrom, H. (1980) Top. Curr. Chem. 87,
1-83 35. Tartar, H. V., and Wright, K. A. (1939) J. Am. Chem. Sot. 61,
539-544 36. Mukerjee, P., and Mysels, K. J. (1971) Critical Micelk Concentru-
tions of Aqueous Surfuctunt Systems, NSRDS-NBS publication No. 36. Sunerintendent of Documents. U. S. Government Print- ing Office, Washington, D. C. ’
37. Lipka, G., Demel, R. A., and Hauser, H. (1988) Chem. Phys. Lipids 48, 267-280

14028 Properties of the Active LPS Unit
lx t
Properties of the Active LPS Unit 14029





























