Embed Size (px)
Citation preview

THE JOURNAL OF RIOLOG~~L CHENISTRY Tel. 242, No. 2, Issue of January 25, pp. 256-264, 196i
Printed in U.S.A.
Physical and Chemical Characterization of the Isomerase of Histidine Biosynthesis in Salmonella typhimurium
(Received for publication, July 28, 1966)
~\IICIIAEL IV. ~IARGOLIES* AND ROBERT F. GOLDBERGER
From the Laboratory of Chemical Biology, National Institute of Arthritis and Metabolic Diseases, National Institutes of Health, Bethesda, Maryland 20014
SUMMARY
The amino acid composition and chemical and immuno- logical properties of the isomerase catalyzing the fourth step in histidine biosynthesis in Salmonella typhimwium are re- ported. The molecular weight of the native enzyme is 29,000, as determined by high speed equilibrium centrifugation; it is unchanged after complete reduction and alkylation, indicat- ing the presence of a single polypeptide chain. This conclu- sion is supported by the finding of a single carboxyl-terminal amino acid, valine, both by digestion with carboxypeptidase A and by hydrazinolysis. Furthermore, the number of spots seen on peptide maps of tryptic digests of carboxymethylated isomerase is approximately the same as the number of arginine plus lysine residues found by amino acid analyses. These results agree with results of genetic studies (1) which show that the structural gene for this enzyme consists of a single complementation group. The minor active isomers of the enzyme, previously described, appear to result from modi- fications in vitro of a single protein.
Antibody specific to the isomerase has been prepared. Carboxymethylated isomerase and isomerase derived from extracts of repressed and derepressed histidine auxotrophs with mutations in all regions of the histidine operon, of con- stitutive mutants, and of the wild type strains LT-2 and LT-7 are antigenically identical.
The structural genes for the 10 enzymes for histidine bio- synthesis in Salmonella typhimurium are located on the chromo- some in a cluster known as the histidine operon. Because of the large body of information now available on the biochemical and genetic aspects of this system (l-3), it was thought especially desirable to obtain data on the physical and chemical character- istics of the enzymes themselves. The isolation and character- ization of the enzymes for histidine biosynthesis have therefore been undertaken. The results for several of these enzymes have been reported (4-6).
We previously described a method (6) for the isolation of the enzyme catalyzing the fourth step in the pathway for histidine biosynthesis in S. typhimurium, an isomerase, the structure of
* Present address, General Surgical Services, Massachusetts General Hospital, Boston, Massachusetts 02114.
which is encoded in the A gene of the histidine operon. Hart- man has shown that mutants of this gene show no intragenic complementation in over 150 tests by abortive transduction (2). The amino acid composition and chemical and immunological properties of the enzyme are reported here, together with evi- dence that it consists of a single polypeptide chain corresponding to the single complementation group.
MATERIALS AND METHODS
Preparation of Enzyme
Growth of bacteria, preparation of cell-free extracts, and purification of the isomerase were performed as described previously (6). Polyacrylamide disc gel electrophoresis was performed according to Davis (7), and gels were stained for protein or isomerase activity as previously reported (6).
Assays
Protein was estimated in crude preparations by the method of Lowry et al. (8), with the use of bovine insulin (Lilly) as standard. Amounts of pure isomerase were calculated on the basis of the optical density at 280 rnp of enzyme in 0.05 M Tris-HCl, pH 8.0 (see “Results”). Assays for isomerase activity were done as previously described (6).
Carboxymethylation
Reduction and alkylation of enzyme were performed according to the method of Anfinsen and Haber (9) as modified by Craven, Steers, and Anfinsen (10). To 10 mg of purified isomerase in 0.05 M Tris-HCl, pH 8.0, was added sufficient recrystallized urea to bring the solution to 8 M in urea (1 ml total volume). After addition of P-mercaptoethanol (Eastman) to a concentration of 0.1 M, the tube was stoppered and incubated for 4 hours at 40”. Following reduction, the solution was diluted 4-fold with 0.1 M
Tris-HCl, pH 8.5. A 4-fold molar excess of twice recrystallized iodoacetic acid (Eastman) was dissolved in sufficient Tris base to bring the pH to 8.5. The iodoacetic acid was then added to the reduced enzyme and incubated for 15 min at room tempera- ture without change in pH. An amount of fl-mercaptoethanol equimolar to the iodoacetic acid used was then added to stop the reaction, and the mixture was dialyzed against buffer appropriate for further use (see below).
256
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1967 M. N. Margolies and R. F. Goldberger 257
Molecular Weight Determinations
Molecular weight and homogeneity were estimated in the Spinco model E analytic ultracentrifuge by the meniscus deple- tion method of Yphantis (ll), with the use of a standard double- sector interference cell. The ultracentrifuge was equipped with Rayleigh interference optics and the temperature for all runs was controlled at 20”. Photographs were taken at zero time and after equilibrium was reached. Attainment of equilibrium re- quired 20 hours of centrifugation except for samples run in 8 M urea; in the latter case photographs were taken after 48 hours of centrifugation. To prepare these samples, enzyme solutions (0.2 to 0.4 mg per ml) were dialyzed for 7 to 10 days against 8 M urea in 0.05 M Tris-HCl, pH 8.0. We assumed v to be 0.725 for the calculation of molecular weights.
Acid Hydrolyses and Amino Acid Analyses
Amino acid analyses were performed according to the method of Spackman, Stein, and Moore (12), with the use of a Beckman model 120B automatic amino acid analyzer coupled with an Infotronics model CRS-1OA digital readout system. Purified native enzyme and the carboxymethylated enzyme were each dialyzed against several changes of 0.2 M NH4HC0a, divided into 1-mg aliquots, and lyophilized to dryness. Duplicate samples of both native and carboxymethylated enzyme were hydrolyzed with constant boiling HCl in sealed evacuated tubes for 19, 45, and 69 hours at 110”. The acid was removed under reduced pressure and the timed hydrolysates were analyzed consecutively.
Tryptophan was estimated by the method of Beaven and Holi- day (13) with the use of solutions of carboxymethylated enzyme in 0.1 M NaOH. Spectra were recorded with the model 15 Cary spectrophotometer. In addition, peptide maps prepared from tryptic digests of the carboxymethylated enzyme were stained with Ehrlich’s reagent (14) to determine the minimum number of tryptophan residues. The reagent, p-dimethylaminobenzal- dehyde, was obtained from Eastman.
Trypsin Digestion and Peptide Jdapping
Samples of carboxymethylated isomerase were dialyzed against 0.2 M NH,HCOs and divided into 2-mg (O.O7+mole) aliquots. Two per cent by weight of trypsin (Worthington), previously treated with diisopropyl fluorophosphate (14), was added, and the solution was incubated for 4 hours at 37”. Following lyoph- ilization, peptide maps were prepared by the method of Katz, Dreyer, and Anfinsen (15) on Whatman No. 3MM chromatog- raphy paper and stained with 0.25a/, ethanolic ninhydrin (East- man).
Carboxypeptidase Digestion
The carboxypeptidases A and B were obtained from Dr. J. T. Potts. These proteolytic enzymes had been treated with DFPl to remove all endopeptidase activity (16). Following such treatment, they exhibited only exopeptidase activity when tested with the following substrates: bovine pancreatic ribo- nuclease A, ribonuclease S, ribonuclease S-protein, ribonuclease S-peptide, parathyroid hormone, and an extracellular nuclease from Staphylococcus uureus. In early experiments, 3-mg (O.l- pmole) samples of reduced, alkylated isomerase were incubated (5 mg per ml) with 2% by weight each of carboxypeptidases A
1 The abbreviations used are: DFP, diisopropyl fluorophos- phate; DNP-, 2,4-dinitrophenyl-.
I I I I x2- 500 505 470 x2- 480 490
FIG. 1. Molecular weight determinations by the meniscus de- pletion method of Yphantis (11). Average molecular weight values are indicated for each set of runs. The constant slopes of the plots for log C with respect to X2 indicate homogeneity. A, native isomerase in 0.05 M Tris-HCl, pH 8.0; B, carboxymethylated enzyme in the same buffer plus 8 M urea; C, carboxymethylated isomerase in buffer; D, native enzyme in buffer plus 8 M urea. The agreement in both calculated molecular weight and degree of homogeneity among Samples A, B, and D suggests that the isomer- ase consists of a single polypeptide chain. Aggregation and pre- cipitation of carboxymethylated enzyme, as seen in C, may be prevented by dialysis against 8 M urea (see B).
and B for 16 to 20 hours in 0.1 M NH4HC03. In later experi- ments, 0.2 pmole of carboxymethylated enzyme was incubated with 2 To by weight of carboxypeptidase A alone. Equal aliquots (0.04 pmole) were withdrawn at 0, 2, 4, 6, and 16 hours. Diges- tions were stopped by freezing and lyophilization. All samples were subjected to amino acid analyses (see above). In addition, control samples, containing amounts of the carboxypeptidase preparation identical with the experimental samples, were incu- bated in parallel and subjected to amino acid analyses. The amount of each contaminating amino acid in these latter samples was always less than 0.002 pmole.
Hydrazinolysis
Hydrazinolysis was performed according to the method of Korenman (17). Anhydrous hydrasine (Matheson, Coleman, and Bell) was used without further treatment, and oxidized bovine pancreatic ribonuclease A (Worthington) was employed as a control. Aliquots of 0.05 pmole were withdrawn at zero time and after 10 hours of hydrazinolysis at 110”. Amino acid anal- yses were performed as described above.
Amino-terminal End Group Analyses
Amino-terminal end group analyses of 0.1 pmole of enzyme were done by the fluoroclmitrobenzene method of Sanger (18), with the use of paper chromatographic separation of the ether- soluble 2,4-dinitrophenyl-amino acids, as described by Redfield and Anfinsen (19). The water phase was examined by high voltage electrophoresis (20). The yields of DNP-amino acids were estimated photometrically at 360 rnp after elution of spots from paper into distilled water at 60” for 15 min. Solutions of authentic DNP-amino acids (Calbiochem) were used as stand- ards. Determination of the amino-terminal end group was also attempted by digestion of carboxymethylated isomerase with leucine aminopeptidase (21). Before use, the leucine amino-
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

258 Isomerase of Histidine Biosynthesis in S. typhimurium Vol. 242, No. 2
FIG. 2. Patterns obtained by electrophoresis in polyacrylamide gels of (1) purified native isomerase and (3) carboxymethylated isomerase. The pattern noted in R appears to be due to aggrega- tion of the carboxymethylated enzyme (cf. Fig. 1).
peptidase (Worthington) was dialyzed against 0.05 M NH.J?ICOa with 0.01 M MgC& and incubated at 37” for 3 hours. Three milligrams of carboxymethylated isomerase dissolved in the same buffer were incubated with 0.3 mg of leucine aminopeptidase at 37” for 16 hours. Aliquots 10.05 pmole) were removed initially and at 16 hours, and lyophilized for amino acid analyses. In addition, parallel incubations of identical amounts of leucine aminopeptidase in the same buffer were analyzed.
Immunizations
New Zealand albino male rabbits, weighing 2500 g, were immunized against various samples of isomerase, with the use of 1 ml of enzyme solution emulsified in 1 ml of Freund’s complete adjuvant (Difco) injected subcutaneously, on the 1st and again on the 7th day of immunization. A booster dose was given at 30 days. For crude enzyme preparations, 20 mg were given at each injection; for purified isomer&se, 1 mg was used for each injection. Prior to use, enzyme solutions were rendered sterile by passage through a Millipore filter with a pore size of 0.22 p. Blood was collected prior to immunization, at the time of the booster injection, and at 4-day intervals thereafter. Sera ob- tained by centrifugation were stored at -20”.
Examination of Antibody
Preer Technique--The minimum number of precipitating antigen-antibody systems present was examined quantitatively by double diffusion in agar by the method of Preer (22). Noble agar (Difco), 0.8%, was employed as the diffusing medium. All tubes were run in duplicate, with the use of serial dilutions of
antigen against a constant amount of undiluted antiserum. Tubes were examined for precipitin bands at 4-hour intervals in a dissecting microscope equipped with a calibrated ocular. The equivalence point for each antigen-antibody system was deter- mined from plots of p (position of band relative to antigen-agar interface) with respect to concentration of antigen for each time of observation, as well as from plots of p with respect to time for each concentration of antigen.
Ouchterlony Technique--Antibody was also examined by double diffusion in agar according to the method of Ouchterlony (23), in prepared plates (Hyland Laboratories) with 4-, 5-, and 7-n-m well distances. Plates were incubated at room tempera- ture for 24 hours.
ImmunoeZectrophore~Immunoelectrophoresis was performed on microscope slides (24) in 0.1 M Verona1 buffer, pH 8.6, for 50 min at 250 volts. The slides were incubated with antiserum at room temperature for 16 hours.
Inactivation-Precipitation-Antibody was also tested for its ability to neutraliie the enzymic activity of the isomerase. To 0.1 ml of immune serum in each of several centrifuge tubes was added 0.1 ml of serial dilutions of purified isomerase. Control tubes contained preimmunization serum with identical serial dilutions of enzyme. All tubes were incubated at 25” for 90 min and stored overnight at 4”. Following centrifugation, each supernatant was quantitatively assayed in duplicate for isomerase activity. Equivalence points for antisera were determined from plots of concentration of activity in the super- natant with respect to concentration of antigen added (see Fig. 8).
Adsorption of Antibody
Removal of contaminating antibodies from the immune serum (see “Results”) was accomplished by incubation of antiserum at 37” for 1 hour with one-tenth volume of a derepressed extract (25 mg of protein per ml) of an isomerase deletion mutant, his- 61% The reaction mixture was stored overnight at 4” and the supernatant was separated by centrifugation. Two consecutive adsorptions were necessary to remove all detectable contaminants (see “Results”).
RESULTS
Molecular Weight DeterwGnations-We have previously de- termined the average molecular weight of the native isomerase by the meniscus depletion method of Yphantis (11) in 0.05 M
Tris, pH 8.0, to be 29,000 (6). Plots of log C against X2 for all five experiments conformed to a straight line, indicating homo- geneity with respect to size (see Fig. 1A). The data were also subjected to analysis on a Honeywell 800 computer with the program of Dr. Parker Small and Dr. Robert Resnick. For this run, at 37,020 rpm, the whole cell weight average molecular weight (J&J was 29,700, the whole cell number average molecular weight (M,J was 27,700, and the whole cell z average molecular weight (ME) was 32,500.
The carboxymethylated isomerase was subjected to equilib- rium centrifugation in order to determine whether the enzyme is composed of subunits. Solutions of reduced and alkylated isomerase in 0.05 M Tris-HCl, pH 8.0, at 0.4, 0.2, and 0.15 mg per ml were examined at 35,600, 31,140, and 35,600 rpm, re- spectively, after 24 hours of centrifugation. The results of a typical experiment are shown in Fig. 1C. The range of molecular weights calculated from the slopes of lines tangent to the curve
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1967 M. N. Margolies and R. F. Goldberger
TABLE I
Amino acid composition of purified isomerase from hisEll
Duplicate samples of 1 mg each were hydrolyzed in acid for 19, from the averages of the 45 and 69.hour values, after complete 45, and 69 hours. Numbers of residues of each amino acid are release of these residues had been effected. Cysteine was deter- based upon a molecular weight of 29,000 for the native isomerase as mined as S-carboxymethylcysteine from a separate set of analyses determined by equilibrium centrifugation. The amounts of on carboxymethylated protein. Tryptophan was estimated by serine, threonine, and tyrosine decreased linearly with time and the method of Beaven and Holiday (13). The amounts of all other were therefore calculated by extrapolation to zero time. amino acids were calculated by averaging the results obtained Amounts of methionine, valine, and isoleucine were calculated from the paired samples at all hydrolysis times.
Amino acid
Lysine ...................... Histidine. .................. Arginine .................... S-Carboxymethylcysteine ... Aspartic acid. ............. Threonine .................. Serine ...................... Glutamic acid. ............. Proline ..................... Glycine ..................... Alanine .................... Valine ...................... Methionine ................. Isoleucine .................. Leucine .................... Tyrosine ................... Phenylalanine .............. Tryptophan ................
Duration of hydrolysis
19 hrs 45 hrs
0.186 0.188 0.077 0.081 0.234 0.233 0.065 0.063 0.406 0.400 0.192 0.190 0.178 0.177 0.497 0.488 0.172 0.179 0.432 0.428 0.414 0.414 0.413 0.425 0.037 0.033 0.232 0.221 0.419 0.421 0.103 0.105 0.090 0.093
wnole
0.180 0.185 0.179 0.191 0.074 0.074 0.074 0.079 0.232 0.235 0.229 0.239 0.061 0.059 0.062 0.063 0.406 0.412 0.401 0.407 0.181 0.184 0.170 0.175 0.156 0.157 0.139 0.141 0.518 0.525 0.510 0.520 0.169 0.167 0.173 0.162 0.429 0.436 0.422 0.430 0.420 0.423 0.418 0.421 0.445 0.449 0.445 0.450 0.040 0.041 0.041 0.043 0.233 0.238 0.236 0.243 0.422 0.424 0.420 0.425 0.097 0.096 0.093 0.094 0.095 0.094 0.094 0.093
shown varied up to values above 50,000. Comparison of the pattern obtained upon electrophoresis of the carboxymethylated protein in polyacrylamide gels to that obtained with the use of native enzyme (see Fig. 2) also indicated size heterogeneity due to aggregation of the reduced and alkylated protein. Therefore, carboxymethylated protein was first dialyzed against 8 M urea in the same buffer for 7 to 10 days and then examined in the ultracentrifuge. Under these conditions, the protein no longer aggregated or precipitated, and a pattern identical with that for native enzyme was seen (see Fig. 1B). For these experiments, solutions of carboxymethylated isomerase, 0.3 mg per ml, were centrifuged at 33,450 rpm for 48 and 60 hours, and at 37,020 rpm for 48 hours; the molecular weights were calculated to be 25,900, 25,000, and 26,100, respectively. These calculations were made with the use of the same v for runs in both buffer and in buffered urea solutions. Control samples of native enzyme (0.2 to 0.4 mg per ml) in 8 M urea, 0.05 M Tris-HCl, pH 8.0, produced patterns identical with those for the carboxymethylated enzyme in urea when examined by the same technique (see Fig. 1D). Data obtained by centrifugation at 40,000 and 36,000 rpm gave calculated molecular weights of 28,500 and 24,200, respectively. The average molecular weight for the reduced and alkylated enzyme in urea (25,700) compares favorably with that for the native enzyme in urea (26,400).
Amino Acid AnalysesThe amino acid compositions of paired samples of native enzyme hydrolyzed in acid for 19, 45, and 69 hours are shown in Table I. The amounts of threonine, serine, and tyrosine decreased linearly with the duration of hydrolysis
- I 69 hrs
-
-I- /.mole
0.185 0.076 0.234 0.062 0.405 0.198 0.192 0.510 0.170 0.429 0.418 0.447 0.041 0.237 0.422 0.107 0.093
11.9 4.9
15.1 4.2
26.1 12.8 12.4 32.9 10.9 27.7 27.0 28.9 2.7
15.3 27.2 6.9 6.0 2.9
and were therefore calculated by extrapolation to zero time; the amounts of methionine, valine, and isoleucine were calculated from the average of the 45- and 69-hour samples, after these amino acids had been completely released by hydrolysis; the amounts of all other amino acids were calculated by averaging the results from all samples. Cysteine was estimated from a separate set of timed hydrolyses of paired samples of carboxymethylated protein. The quantitative conversion of cysteine to S-carboxy- methylcysteine was shown by the absence of cysteine or cysteic acid in the latter analyses. The reduced and alkylated enzyme had no detectable isomerase activity.
There were 2.9 residues of tryptophan per mole of protein as determined from the spectrum of the isomerase in 0.1 M NaOH (Fig. 3). Peptide maps prepared from tryptic digests of car- boxymethylated isomerase, stained with Ehrlich’s reagent, revealed two spots.
On the basis of the absorbance at 280 rnp of a solution of native isomerase in 0.05 M Tris-HCl, pH 8.0, together with determina- tions of protein concentration by amino acid analyses, the molar extinction coefficient was calculated to be 3.31 x 104.
Peptide MapsPeptide maps prepared from tryptic digests of carboxymethylated isomerase revealed a reproducible pattern consisting of 34 ninhydrin-positive spots; the origin was free of ninhydrin-positive material (see Fig. 4). This agrees with the total number of lysine plus arginine residues as determined by amino acid analyses.
Carboxyl-terminal End Group AnalysisInitial incubations of 0.1 pmole of carboxymethylated isomerase with carboxypepti-
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

260 Isomerase of Histidine Biosynthesis in S. typhimurium Vol. 242, n-o. 2
munization of rabbits with a crude extract of derepresseds S. typhimurium hisEl1 was examined against purified isomerase by double diffusion in agar, with the Ouchterlony and Preer techniques, only one band was seen. However, the antibody prepared in this way was not sufficiently strong for further examination of homogeneity. As expected, when this antiserum was reacted against the original crude extract, multiple strong bands were seen. However, antisera produced by immunization with purified isomerase were satisfactory for further studies. The results obtained when these antisera were examined in Preer tubes containing serial dilutions of the purified isomerase are shown in Fig. 6. According to plots of p against time, and p against concentration of antigen (and by inspection), the equiva- lence point for the major system seen was found to be at an antigen concentration of 65 pg per ml. In addition, several minor components were revealed at much higher concentrations of antigen. Corresponding patterns were seen in Ouchterlony plates.
The conclusion that the major precipitin system was due to the reaction of the isomerase with its specific antibody is based on three lines of evidence. (a) After the antiserum had been ad- sorbed with a crude extract of S. typhimurium his-612 (a mutant in which the entire A gene is deleted) and then was reacted against purified isomerase, no contaminating system was ob- served (see Fig. 6B). Conversely, when the reaction between unadsorbed antibody and serial dilutions of the same his-612
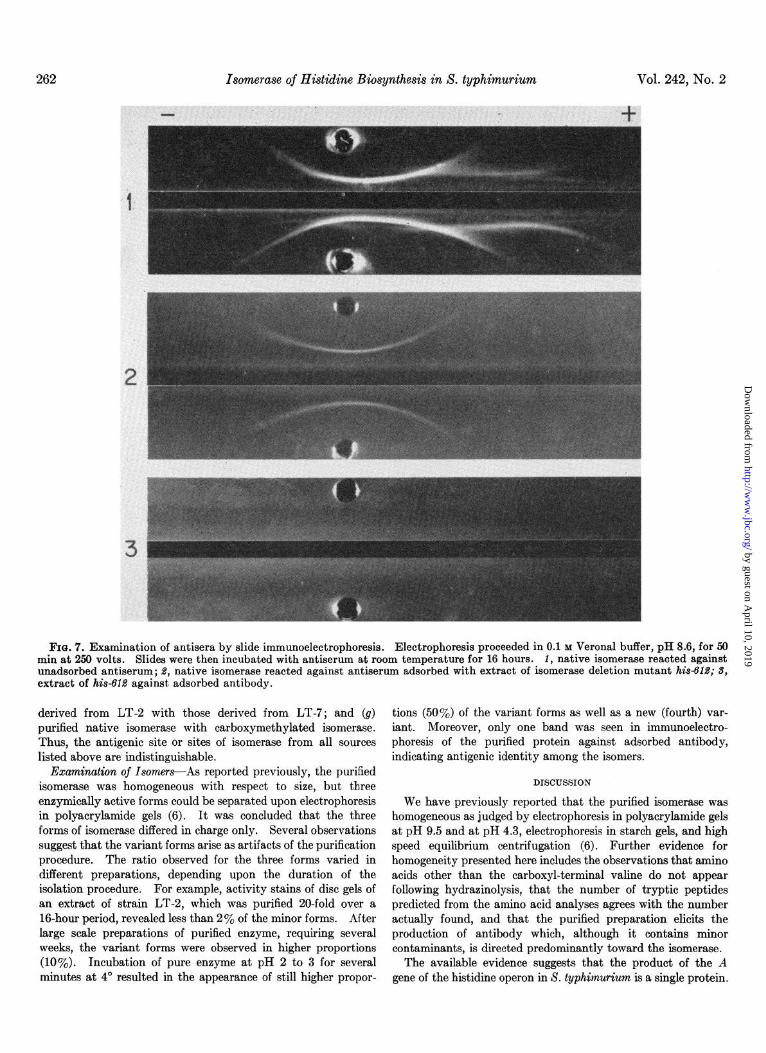
extract was examined in Preer tubes, the major band due to the isomerase was absent. (b) When the unadsorbed antiserum was examined with purified isomerase by immunoelectrophoresis, the major component was due to the isomerase (Fig. 7,1). The contaminating systems were absent when the isomerase was reacted with the adsorbed antiserum (Fig. 7, 2). The minor contaminating bands were also identified by their presence in runs with the use of extracts of the deletion mutant h&61,?
against the unadsorbed antibody and their absence in reactions involving the adsorbed antibody (Fig. 7, S). (c) The reaction of isomerase with its antibody could also be studied by the ability of the antiserum to inactivate and precipitate the isomerase.
The antigen-antibody precipitate in agar did not stain in the color reaction for isomerase activity (6), nor could resuspended precipitates be shown to contain isomerase activity in the standard assay. A plot of isomerase activity remaining in the supernatants after reaction of antiserum with serial dilutions of purified enzyme, with respect to concentration, is shown in Fig. 8. By comparison with the plot obtained from the same experiment done with control (preimmunization) serum, it is clear that the antiserum removed all enzymic activity from solution up to a concentration of 65 pg per ml. The equivalence point obtained in this way is identical with that obtained from studies of double diffusion in agar.
WAVELENGTH (mp)
FIG. 3. Ultraviolet absorption spectra of carboxymethylated isomerase, at a concentration of 0.60 mg per ml, in 0.05 M Tris-HCl. pH 8.0 (-), and, at a concentration-of 0.39 mg per ml, in 0.1 & NaOH (-- -). The tryptophan content of the isomerase was calculated from the latter curve, with the use of the method of Beaven and Holiday (13).
dases A and B for 20 hours resulted in 72% yield of valine as well as yields of 26 y0 for aspartic acid, 27% for leucine, and 19% for alanine. Results of timed digestions of carboxymethylated
isomerase with carboxypeptidase A, with the use of 0.05 pmole of isomerase for each analysis, are shown in Fig. 5. The calcu- lated final yield of valine in this experiment was 150%. This result suggests that the penultimate residue is also valine. Alanine, aspartic acid, and leucine were again obtained in low yield; it has not been possible to distinguish the order in which they appear.
The identity of the carboxyl-terminal residue was confirmed by
hydrazinolysis of 0.07 pmole of the isomerase for 10 hours which resulted in 75 y0 yield of valine No other amino acids were released by hydrazinolysis. A 0.1~pmole sample of oxidized ribonuclease, run in parallel, produced a 52y0 yield of valine.2
Amino-terminal End Group AnaZysisAmino-terminal end group analysis of 0.1 pmole of the isomerase, with the use of 2,4-dinitrofluorobenzene, revealed only a trace of threonine (less than 30/,), although the yield of e-DNP-lysine was greater than 70%. In addition, the carboxymethylated enzyme was refrac- tory to attempts at prolonged digestion by leucine aminopep-
tidase.
Immunological Studies-When antiserum prepared by im-
2 The carboxyl-terminal amino acid residue of bovine pancreatic ribonuclease is known to be valine (25).
The reaction between crude extracts of derepressed histidine
auxotrophs and antibody was also examined by the above techniques. The results obtained with both adsorbed and
3 When S. typhimurium is grown in the presence of excess histi- dine, the histidine operon is repressed: the enzymes for histidine biosynthesis are synthesized at a relatively low rate. In histidine auxotrophs grown under conditions in which the growth rate is limited by the amount of histidine available, the histidine operon becomes derepressed, and the enzymes for histidine biosynthesis are synthesized at a relatively high rate, producing levels of en- zymes up to 30 times those of the wild type (LT-2) organism.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1967 M. N. Margolies and R. F. Goldberger 261
CHROMATOGRAPHY -
FIG. 4. Peptide maps of tryptic digests of 0.1 pmole of car- boxymethylated isomerase prepared by the method of Katz, Dreyer, and hnfinsen (15), stained with ninhydrin. The patterns were reproducible and contained 34 ninhydrin-positive spots.
unadsorbed antibody were similiar to those obtained when putified enzyme was used as antigen. After adsorption, the antiserum formed only one band with pure and with crude preparations of isomerase, and formed no detectable precipitate with extracts of the deletion mutant.
When examined by double diffusion in agar by the Ouchterlony technique, with the use of antiserum prepared against purified isomerase, reactions of identity were seen between the following antigen preparations: (a) purified isomerase with crude extract of derepressed h&El 1; (b) partially purified isomerase from the wild type strain LT-2 (repressed) with crude extract of dere- pressed hisEll; (c) crude extract of derepressed LT-2* with crude extracts of repressed hi&II (grown on excess histidine) ; (d) crude extracts of both repressed and derepressed histidine auxotrophs (other than A gene mutants) with extracts of various constitutive mutants5; (e) crude extracts of LT-2 with crude extracts of another wild type strain, LT-7; fj) crude ex- tracts of both repressed and derepressed histidine auxotrophs
4 Derepression of the wild type organism was effected by addi- tion to the growth medium of nn-2-thiazole alanine (Cycle Chemi- cal Company) at a concentration of 0.5 mM. This compound is known to inhibit the first enzyme in the pathway for histidine biosynthesis (26).
5 The enzymes for histidine biosynthesis are present in rela- tively high concentrations in organisms with certain mutations outside the histidine operon; these are the constitutive mutants (27).
/
0 I I I
0 2 4 6 TIME (hrs.)
FIG. 5. Release of amino acids during timed digestion of car- boxymethylated isomerase with carboxypeptidase A. A total of 0.2 pmole of the reduced and alkylated isomerase was incubated with 2oJc, by weight, carboxypeptidase A at 37”. Valine was ob- tained early and in high yield, but it has not been possible to dis- tinguish the order in which alanine, aspartic acid, and leucine were released.
A Ag I:1 I:2 I:4 1~8 ItI6 I:32 I:64 I:128
Ab I:1 I:1 I:1 I:1 Id I:I I:I I:1
B Ag I:1 I:2 I:4 1:8 I:16 I:32 I:64 I:128
Ab PI I:1 I:1 Id I:1 I:I I:I I:I
FIG. 6. Double diffusion in agar, by the method of Preer (22), of serial dilutions of purified isomerase against constant amounts of undiluted antiserum. A, reaction of isomerase with unadsorbed antiserum reveals a major antigen-antibody system with equiva- lence at 65 pg of isomerase per ml. Minor components are seen only at high antigen concentrations. B, the contaminating sys- tems were no longer detected when pure isomerase was reacted against antiserum previously adsorbed with an extract of the dele- tion mutant !&+6+fd.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
262 Isomerase of Histidine Biosynthesis in S. typhimurium
FIU. 7. Examination of antisera by slide immunoelectrophoresis. Electrophoresis proceeded in 0.1 M Verona1 buffer, pH 8.6, for 50 min at 250 volts. Slides were then incubated with antiserum at room temperature for 16 hours. 1, native isomerase reacted against unadaorbed antiserum; 8, native isomerase reacted against antiserum adsorbed with extract of isomerase deletion mutant his4’ld; 3, extract of hisSld against, adsorbed antibody.
derived from LT-2 with those derived from LT-7; and (g) tions (50%) of the variant forms as well as a new (fourth) var- purified native isomerase with carboxymethylated isomerase. iant. Moreover, only one band was seen in immunoelectro- Thus, the antigenic site or sites of isomerase from all sources phoresis of the purified protein against adsorbed antibody, listed above are indistinguishable. indicating antigenic identity among the isomers.
Examination of Isomers---As reported previously, the purified isomerase was homogeneous with respect to size, but three enzymically active forms could be separated upon electrophoresis in polyacrylamide gels (6). It was concluded that the three forms of isomerase differed in charge only. Several observations suggest that the variant forms arise as artifacts of the purification procedure. The ratio observed for the three forms varied in different preparations, depending upon the duration of the isolation procedure. For example, activity stains of disc gels of an extract of strain LT-2, which was purified 20-fold over a 16-hour period, revealed less than 2% of the minor forms. After large scale preparations of purified enzyme, requiring several weeks, the variant forms were observed in higher proportions (10%). Incubation of pure enzyme at pH 2 to 3 for several minutes at 4” resulted in the appearance of still higher propor-
DISCUSSION
We have previously reported that the purified isomerase was homogeneous as judged by electrophoresis in polyacrylamide gels at pH 9.5 and at pH 4.3, electrophoresis in starch gels, and high speed equilibrium centrifugation (6). Further evidence for homogeneity presented here includes the observations that amino acids other than the carboxyl-terminal valine do not appear following hydrazinolysis, that the number of tryptic peptides predicted from the ammo acid analyses agrees with the number actually found, and that the purified preparation elicits the production of antibody which, although it contains minor contaminants, is directed predominantly toward the isomerase.
The available evidence suggests that the product of the A gene of the hiitidine operon in S. typhimurium is a single protein.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1967 M. N. Margolies and R. F. Goldberger 263
G; ISOMERASE CONCENTRATION (mg/ml)
FIG. 8. Neutralization and precipitation of isomerase by anti- serum. A plot of isomerase activity remaining in the supernatant with respect to antigen concentrations after reaction of serial dilu- tions of purified isomerase with control serum (- - -) and anti- serum (-) reveals that the equivalence concentration is 65 ag of isomerase per ml. This is identical with the equivalence point found in Preer tubes (upper part of the figure), calculated from plots of p against time and p against concentration.
FIO. 9. Reactions of identity among isomerase preparations .- from various sources, as seen in Ouchterlony plates. Center well contains adsorbed antibody; incubation with antigens proceeded at room temperature for 16 hours. Antigen wells contain: 1, puri- fied native isomerase; %, crude extract of derepressed hisElf; 3, partially purified (repressed) LT-2; 4, crude extract of constitutive mutant hisTl601; 6, carboxymethylated isomerase.
The variant forms distinguished by their different electrophoretic mobilities appear to arise as artifacts during large scale prepara- tions of the enzyme. Such variants might be produced, for example, by deamidat,ion of glutamine or asparagine residues during the isolation procedure.
The conclusion that the isomerase is composed of a single polypeptide chain is based upon the following evidence. (a) The molecular weight and homogeneity, as determined by ultra- centrifugation, were not affected by exposure to 8 M urea or by carboxymethylation. (6) There appears to be only one car- boxyl-terminal residue, valine, as determined by both hydra- zinolysis and digestion with carboxypeptidase A. (c) The possibility that the isomerase is composed of two or more identical subunits is ruled out by the finding that the number of peptides produced by tryptic digestion corresponds closely to the total number of lysine plus arginine residues determined by amino acid analyses.
The studies presented here show that the isomerase isolated from a derepressed histidine auxotroph (hisEll) is antigenically identical with that of repressed and derepressed histidine auxo- trophs with mutations in other genes of the histidine operon, constitutive mutants, the wild type strains LT-2 and LT-7, and carboxymethylated isomerase.
Those A gene mutations which result in the production of isomerase with amino acid substitutions that alter antigenicity are widely distributed throughout the map of the A gene.6 Thus, the tertiary conformation of the isomerase which is re- quired for antigenic specificity may be destroyed by a change in any of a large number of amino acids widely distributed along the polypeptide chain. The conformation required for enzymic activity is probably even more specific since it is destroyed by carboxymethylation of the native enzyme while antigenicity is preserved and since extracts of many A gene mutants are enzymically inactive but antigenically competent.
Acknowbdgmenk-The authors are indebted to Dr. Christian B. Anfinsen for critical suggestions during the course of this work. We wish to thank Dr. Edward Steers, Jr., for perform- ance of the ultracentrifugation and for advice in the immunolog- ical studies. Dr. Bruce N. Ames kindly supplied the mutants used. We also thank Mrs. Marilyn Meyers for technical as- sistance and Mr. Clifford Lee for performing the amino acid
REFERENCES
1. LOPER, J. C., GRABNAR, M., STAHL, R. C., HARTMAN, Z., AND HARTMAN, P. E., Brookhaven Symp. Biol., 17, 15 (1964).
2. AYES, B. N., AND HARTMAN, P. E., Cold Spring Harbor Symp. Quant. Biol., 28, 349 (1963).
3. GOLDBERGER, R. F., AND BERBERICH, M. A., in H. J. VOGEL, J. 0. LAMPEN, AND V. BRYSON (Editors), Organizational bio- sunnthesis, Academic Press, New York, in press.
4. LOPER, J. C., AND ADAMS, E., J. Biol. Chem., 249,788 (1965). 5. MARTIN, R. G., AND GOLDBERGER, R. F., J. Biol. Chem., in press. 6. MARGOLIES, M. N., AND GOLDBERGER, R. F., J. Biol. Chem.,
241, 3262 (1966). 7. DAVIS, B. J., Ann. N. Y. Acad. Sci., lai, 494 (1964). 8. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL,
R. J., J. Biol. Chem., 193, 265 (1951). 9. ANFINSEN, C. B., AND HABER, E., J. Biol. Chem., 236, 1361
(1961).
6 M. N. Margolies and R. F. Goldberger, manuscript in prepara- tion.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

264 Isomerase of Hi&dine Biosynthesis in S. typhimurium Vol. 242, No. 2
10.
11.
12.
13.
14.
15.
16.
17.
18.
CRAVEN, G. R., STEERS, E., JR., AND ANFINSEN, C. B., J. Biol. 19. REDFIELD, R. R., AND ANFINSEN, C. B., J. Biol. Chem., 221, Chem., 240, 2468 (1965). 385 (1956).
YPHANTIS, D. A., Biochemistry, 3, 297 (1964). 20. BISERTE, G., HOLLEMAN, J. W., HOLLEMAN-DEHOVE, J., AND
SPACKMAN, D. H., STEIN, W. H., AND MOORE, S., Anal. Chem., SAUTIERE, P., in M. LEDERER (Editor), Chhromatographic
30, 1190 (1958). reviews, VoZ. 2, American Elsevier Publishing Company,
BEAVEN, G. H., AND HOLIDAY, E. R., Advan. Protein Chem., 7, New York, 1960, p. 59.
319 (1952). 21. HILL, R. L., AND SMITH, E. L., J. BioZ. Chem., 231,117 (1953).
CANFIELD, R. E., AND ANFINSEN, C. B., in H. NEURATH (Edi- 22. PREER, J. R., J. Immunol., 77, 52 (1956).
tor), The proteins, Vol. 1, Academic Press, New York, 1963, 23. OUCHTERLONY, O., Acta Path. Microbial. Stand., 26,507 (1949).
p. 319. 24. KABAT, E. A., AND MAYER, M. M., Experimental immuno-
KATZ, A. M., DREYER, W. J., AND ANFINSEN, C. B., J. Biol. chemistry, Charles C Thomas Publisher, Springfield, Ill.,
Chem., 234, 2897 (1959). 1964, p. 642.
POTTS, J. T., BERGER, A., COOKE, J., AND ANFINSEN, C. B., 25. ANFINSEN, C. B., REDFIELD, R. R., CHOATE, W. L., PAGE, J.,
J. Biol. Chem., 237, 1851 (1962). AND CARROLL, W. R., J. Biol. Chem., 207,201 (1954).
KORENMAN, S., Biochem. Biophys. Acta, 124, 160 (1966). 26. MARTIN, R. G., J. Biol. Chem., 238, 257 (1963).
SANGER, F., Biochem. J., 45, 563 (1949). 27. ROTH, J. R., ANTON, D. N., AND HARTMAN, P. E., J. Mol. Biol.,
in press.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Michael N. Margolies and Robert F. GoldbergerSalmonella typhimuriumBiosynthesis in
Physical and Chemical Characterization of the Isomerase of Histidine
1967, 242:256-264.J. Biol. Chem.
http://www.jbc.org/content/242/2/256Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/242/2/256.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from








![Research Article CrystalStructureofL-Histidinium2 ...chloride monohydrate [2], L-histidine tetrafluoroborate [3], L-histidine hydrochloride monohydrate [4], L-histidine hydrofluoride](https://img.pdfslide.us/doc/110x75/60b51c180636315681384205/research-article-crystalstructureofl-histidinium2-chloride-monohydrate-2.jpg)




















