Embed Size (px)
Citation preview

Phylogeography and evolutionof freshwater cottid fishes
Tytti Kontula
Department of Ecology and SystematicsDivision of Population Biology
andFinnish Museum of Natural History
University of HelsinkiFinland
Academic dissertation
To be presented, with the permission of the Faculty of Scienceof the University of Helsinki, for public criticism in the Auditorium 1041
of Biocenter 2 (Viikinkaari 5) on October 3rd 2003 at 12 o’clock noon
Helsinki 2003

© Tytti Kontula (chapter 0)© Blackwell Science (chapters I and IV)© authors (chapters II and III)© Academic Press (chapter V)
Cover illustration by Vappu Penttilä
Author’s present address:Finnish Environment InstituteNature DivisionP.O. Box 140FIN-00251 HelsinkiFinland
E-mail: [email protected]
ISBN 952-91-6346-0 (paperback)ISBN 952-10-1384-2 (PDF)http://ethesis.helsinki.fi
Oy Edita AbHelsinki 2003

Original articles and manuscripts
The thesis is based on the following articles and manuscripts, which are referred to inthe text by their Roman numerals:
I Kontula T & Väinölä R (2001) Postglacial colonization of Northern Europe bydistinct phylogeographic lineages of the bullhead, Cottus gobio. MolecularEcology, 10, 1983–2002.
II Kontula T & Väinölä R (2003) Molecular and morphological analysis of sec-ondary contact zones of Cottus gobio in Fennoscandia: geographical discord-ance of character transitions. Biological Journal of the Linnean Society, in press.
III Kontula T & Väinölä R (2003) Comparative phylogeography of freshwater cottidfishes: continent-scale versus European affinities in the Cottus poecilopus andC. gobio complexes. Submitted.
IV Kontula T & Väinölä R (2003) Relationships of Palearctic and Nearctic ‘glacialrelict’ Myoxocephalus sculpins from mitochondrial DNA data. Molecular Ecol-ogy, in press.
V Kontula T, Kirilchik SV & Väinölä R (2003) Endemic diversification of the mono-phyletic cottoid fish species flock in Lake Baikal explored with mtDNAsequencing. Molecular Phylogenetics and Evolution, 27, 143–155.
Supervised by: Doc. Risto VäinöläFinnish Museum of Natural HistoryUniversity of HelsinkiFinland
Reviewed by: Ass. Prof. Maarit JaarolaUniversity of LundSweden
Doc. Jaakko LummeUniversity of OuluFinland
Examined by: Prof. Filip VolckaertKatholieke Universiteit LeuvenBelgium


1Phylogeography and evolution of freshwater cottid fishes
Summary
Introduction
Phylogeographic inferencePhylogeography is a branch of biogeographyconcerned with geographic patterns of evo-lutionary subdivisions within species oramong closely related species (Avise et al.1987; Avise 2000). It is intimately related tothe very core of life sciences, formation ofnew species. Speciation is usually a long-termprocess, which relates to decrease of geneflow between the differentiating taxa. There-fore, phylogeographic studies may offer snap-shots of evolutionary processes, which leadto origin of new species.
Comparative approach to phylogeographyof co-distributed taxa can reveal previouslyunrecognized biogeographic patterns in re-gional biota. In Europe, common subdivi-sions among terrestrial species have been in-terpreted in relation to the Pleistocene gla-cial events (Taberlet et al. 1998; Hewitt1999). Relationships of extant populationshave been examined particularly using mito-chondrial DNA (mtDNA) data, and concord-ance in the intraspecific lineage distributionsamong different species have been explainedby shared glacial refugia and routes ofpostglacial recolonization. Particularly, theIberian Peninsula, Italy, the Balkans andsoutheastern Europe are recognized as themajor postglacial dispersal centres of terres-trial biota in the western Palearctic (Fig. 1).On the scale of whole Eurasia, phylogeo-graphic studies of terrestrial taxa are still few(e.g. Jaarola & Searle 2002; Brunhoff et al.2003).
European freshwater zoogeographyA plain inspection of distributional patternsof European aquatic fauna shows an import-ant difference compared to the terrestrial spe-
cies: The Iberian and Italian peninsulas, andcentral and southern Greece have not con-tributed much to the postglacial recolon-ization of European freshwaters (Banarescu1990, 1992). This is hardly surprising, as noPleistocene river connections are knownacross the Pyrenees or Alps. Instead of south-ern peninsulas, the Danube river basin insoutheastern Europe appears to have beencrucially important for the initial invasion ofEuropean freshwaters; almost all fishes liv-ing in the Atlantic and Baltic Sea basins arefound also in the Danube (Banarescu 1990).
Among fishes, no comprehensive phylo-geographic comparisons have been made inEurope this far, although mtDNA surveyshave been published from several species(e.g. Bernatchez & Dodson 1994; Durand etal. 1999; Nesbø et al. 1999; Englbrecht etal. 2000; Koskinen et al. 2000; Bernatchez2001; Brunner et al. 2001; Nilsson et al.2001). Broad-scale studies have regularlyshown subdivisions into several refugial line-ages within species, but only rarely have ob-vious concordance in the lineage distributions
Fig. 1 The main potential glacial refugia of terres-trial species in Europe, and the maximum ice coverduring the last ice age according to Taberlet et al.(1998) and Hewitt (1999).

2 Kontula
been noted among species. The level of ob-served intermixing between the differentiatedunits has also varied from almost completeallopatry (e.g., chub Leuciscus cephalus;Durand et al. 1999) to thorough intermixingacross large areas (e.g. perch Perca fluviatilis;Nesbø et al. 1999). In any event, apart fromthe Danubian region, several studies havepointed also to more northern refugia, situ-ated relatively close to the ice margin inWestern and Central Europe (e.g. Durand etal. 1999; Nesbø et al. 1999; Hänfling et al.2002).
In Northern Europe, the Baltic Sea hasbeen the single most important dispersal routenorthwards for aquatic fauna during and afterthe deglaciation. The alternating fresh- andbrackish water phases of the Baltic basin(Donner 1995) have variously enabled anddisabled the postglacial immigration of fresh-water species. Apart from Central Europeanrivers in the south, the basin could have beenreached from the east through the ice-lakesystems that were situated along the marginof the Scandinavian ice-sheet (Kvasov 1979;Faustova 1984). The northeasterly ice-lakeshave been proposed as the immediate sourceof several early invaders of the Baltic Sea,e.g., the “glacial relict” crustaceans (Seger-stråle 1982; Väinölä et al. 1994). The sameregion has been regarded also as a glacialrefugium for freshwater fishes, such as theAtlantic salmon Salmo salar (Koljonen et al.1999) and perch (Nesbø et al. 1999).
Fish diversity on Eurasian scaleIn the traditional freshwater zoogeographyof northern Eurasia, the main division occursbetween the Euro-Mediterranean and Sibe-rian subregions (Banarescu 1992). However,the European vs. Siberian distinction does notappear to repeat itself on the level of intra-specific phylogeography, as shown by the fewcontinent-scale surveys accomplished this far(Bernatchez & Dodson 1994; Brunner et al.
2001; Van Houdt et al. 2003). For instance,in the Coregonus lavaretus whitefish com-plex, the deepest split in the HolarcticmtDNA diversity separates the North Ameri-can continental whitefish from the Eurasian+ Beringian populations, and the main divi-sion within the Euro-Siberian region is foundbetween a basically Alpine lineage and theothers (Bernatchez & Dodson 1994).
The ancient Lake Baikal in East Siberiarepresents an environment very differentfrom the rest of Eurasia, and in terms ofaquatic fauna it is referred to a distinct bio-geographical subregion (Banarescu 1992).Baikal is the deepest and oldest lake on earth;one or more lake basins have existed in theBaikal rift valley since the Oligocene (Mats1993). It harbours more than 1000 endemictaxa, including several species flocks of in-vertebrates, such as amphipods, oligochaetesand mollusks (Timoshkin 1997; Kozhova &Izmest’eva 1998; Sherbakov 1999). As re-gards native fishes, the Baikalian fauna com-prises Eurasian and Siberian species, and theendemics belonging to the suborder Cottoidei(Sideleva 2000). The non-endemic and en-demic fishes occupy mainly separate habi-tats in Lake Baikal. The Eurasian and Sibe-rian taxa live primarily in the shallow litto-ral zone, whereas the endemic cottoids in-habit deeper waters.
Cottids, abyssocottids and comephorids - thefish of interest in the thesisThis thesis is a step towards understandingevolution in a group of fishes during the lastfew million years: the history of the north-ern freshwater cottids and their relatives inLake Baikal. The family Cottidae, and theBaikalian endemic families Abyssocottidaeand Comephoridae belong to the suborderCottoidei. In its entirety, Cottoidei compriseseleven families, and its distribution encom-passes all oceans. The diversity of the fam-ily Cottidae (sculpins) is highest in the ma-

3Phylogeography and evolution of freshwater cottid fishes
rine realm of the northern Pacific (c. 130 spe-cies; Froese & Pauly 2003). Freshwatercottids outside Lake Baikal are confined tofour genera Mesocottus, Trachidermus,Myoxocephalus, and Cottus, which has 40species in North America and Eurasia. A com-mon anatomical feature for cottids, and alsofor Baikalian cottoids, is the lack of swim-ming bladder, which basically determinestheir typically benthic life-style.
Only three species of Cottus are wide-spread in Eurasia. The Alpine bullhead C.poecilopus is a cold-adapted fish, which in-habits rivers and lakes from the Russian FarEast to restricted areas of Northern and East-ern Europe (Fig. 2A). The European com-mon bullhead C. gobio is also regarded as acold-adapted species, but in mountain areasit normally lives downstream of the C.poecilopus range (e.g. Witkowski 1979).Both C. poecilopus and C. gobio thrive alsoin the brackish coastal waters of the Baltic
Sea (in salinities up to 6 and 7 p.p.t, respect-ively; Koli 1969; Andreasson 1972). East ofthe Ural Mountains, C. gobio is replaced bythe similarly widespread Siberian C. sibiricus(Fig. 2B), and a close relative, C. ricei, in-habits freshwaters in North America. On thebasis of head sensory canal pores, C. gobio,C. sibiricus, C. ricei, and three taxa onlyknown from the Syr-Darya drainage, havebeen referred to the gobio group (Nybelin1958; McAllister & Lindsey 1961; Koli1969).
Fishes of the Myoxocephalus quadri-cornis complex have a circumpolar distribu-tion primarily in Arctic coastal and estuarinewaters (Fig. 2C). Yet, they also occur in deeplakes of the once-glaciated northern regionsin Europe and North America, where they areregarded as a part of the “glacial relict” zoo-geographical element (Segerstråle 1962;Dadswell 1974). The North American conti-nental Myoxocephalus populations are re-
Fig. 2 Distributions of Cottus poecilopus (A), C. gobio and C. sibiricus (B), and fishes of theMyoxocephalus quadricornis complex (C). The distributions of Cottus species are primarily accordingto Sideleva (2002), with modifications from our own data, and from Andreasson (1972) and Witkowski(1979). For M. quadricornis complex, the Eurasian range is according to Ekman (1953), and NorthAmerican according to Parker (1988) and Houston (1990). The question mark in A denotes an uncer-tain part of C. poecilopus range. The approximate southern limits of the Late Wisconsinian / LateWeichselian continental glaciers in North America and northern Europe are indicated in C.

4 Kontula
ferred to the deepwater sculpin M. q. thomp-sonii or M. thompsonii, distinct from the Arc-tic and European fourhorn sculpin M. q.quadricornis or M. quadricornis (McAllister1961; McAllister & Aniskowicz 1976;Eschmeyer 1998).
In comparison to the widespread Eurasiancottids, their Baikalian relatives show muchmore diverse morphologies, and also life-styles (Sideleva 1982, 2001). Therefore, theyhave been classified into three separate fami-lies of the Cottoidei: Abyssocottidae, Come-phoridae and Cottidae. In Baikal, the evolu-tion of cottoids has proceeded in two direc-tions from the proposed ancestral shallow-water, benthic habitats (Sideleva 1994). Theyhave invaded even the most abyssal environ-ments deeper than 1500 m, and some spe-cies have shifted to a pelagial life-style. Thepelagial adaptations of the Baikalian oilfishesComephorus include a lowered body densitythrough increased fat content (resulting inbuoyancy and a translucent body) and vivipa-rous reproduction in the water column. Asregards the origin of the Baikalian cottoiddiversity, interpretations of the morpholog-ical and molecular evidence have led to ratherdifferent age estimates, which altogetherrange from 1 Myr to Oligo-Miocene ages(e.g. Taliev 1955; Sideleva 1982; Hunt et al.1997; Kiril’chik & Slobodyanyuk 1997).
The original papers of this thesis, I–V, ex-amine the variation of freshwater cottoids onvarious spatial scales, and across the micro-and macroevolutionary time scales. In II, thefocus is on regional variation and on localpopulation structure in the contact zones be-tween intraspecific lineages of Cottus gobio;I+III and IV offer insights to phylogeo-graphic patterns on the Holarctic scale in theC. gobio, C. poecilopus and Myoxocephalusquadricornis species complexes; and in V, amolecular phylogeny is build for the endemiccottoid species flock of Lake Baikal.
Material and methods
SamplesTo examine phylogeographic patterns amongnorthern cottid fishes, samples from over 130locations were studied for mitochondrialDNA, allozyme or morphological variation.The initial sampling of C. gobio was amplein Northern Europe (I). After the localizationof the contact areas between the distinct C.gobio lineages sampling was further focusedon two regions, one in southern Finland andthe other in northern Finland and northernSweden (Fig. 3; II). For the broad-scalephylogeographic study of the C. gobio andC. poecilopus species complexes (III), sam-ples were obtained from ten countries acrossEurasia, and from Canada. The relationshipswithin the Myoxocephalus quadricornis spe-cies group were assessed using material fromthe Canadian Arctic coast, from the BalticSea, and from land-locked populations inNorth America and Northern Europe (IV).The material used in the phylogenetic studyof the Baikalian cottoid species flock (V)included DNA samples of 22 of the 33Baikalian species, and of 8 outgroup taxafrom Japan, USA and Russia.
Molecular and morphological analysesIn recent years, mitochondrial DNA (mt-DNA) has been the most popular marker inphylogeographic studies (Avise 2000). Ap-plicability of mtDNA markers is due to thealmost exclusively maternal inheritance, lackor rarity of recombination and fairly fastmutation rate of the mitochondrial genome.In this thesis, sequencing or restriction frag-ment length polymorphism (RFLP) analyseswere used to recognize distinct mtDNA line-ages and to assess relationships among them.Complete or nearly complete mitochondrialcytochrome b, ATPase 8 and 6 genes, andnon-coding control region were sequenced(in all 2833–2916 bp) to examine phylogeny

5Phylogeography and evolution of freshwater cottid fishes
in the Cottus species complexes (III), and inthe Baikalian cottoid species flock (V); inother studies mtDNA analyses were based onshorter fragments.
Allozyme electrophoresis was used toassess gene frequency variation at nuclearloci in North European C. gobio (I–II). First,variation was scored at 25 loci to character-ize differentiation between the main lineages(I), and the five most informative ones wereused in the analysis of the contact zonepopulations (II).
In addition to molecular markers, fourmorphometric characters, previously foundto differentiate the Fennoscandian lineages
of C. gobio (Koli 1969), were studied formore than 600 individuals, from the contactzone populations or from the “pure” lineages(II).
Data analysesPhylogenetic methods. Phylogenetic analy-ses in I and in III–V applied the maximum-parsimony (MP), maximum-likelihood (ML)or minimum-evolution (ME) criteria. The MPand ML methods aim at inferring the nucle-otide changes separately at each site alongthe sequence for a given topology, whereasME applies genetic distances calculated frompairwise sequence comparisons (e.g. Nei &
Fig. 3 The areas studied in the analysis of the C. gobio contact zones, and distributions of the westernand eastern mtDNA lineages and of GPI-1 allele frequencies (left and right pie charts, respectively)north of the Baltic Sea (A) and in southern Finland (B). The map in the lower left corner shows thelocations of the two study areas, and the corresponding data for the 6 reference populations represent-ing the pure lineages (the hatched circle denotes the third, southeastern haplotype group).

6 Kontula
Kumar 2000). In MP, the mutational changesare assumed to occur equally in all directionsamong the four nucleotides, and the topol-ogy (or topologies), which explains the wholegenealogy of the studied sequences withsmallest number of changes, is chosen to bethe best. In ML, the preferred tree (topologyand branch lengths) is the one, which maxi-mizes the likelihood of obtaining the ob-served set of sequences under a specific sub-stitution model. The MP analysis was appliedas the exclusive or primary method for as-sessing the relationships among mtDNAhaplotypes in I and IV, where the haplotypesof main interest were closely related. Theanalyses of broader species complexes (IIIand V) employed also ML. A hierarchicalseries of likelihood ratio tests were used tofind the most appropriate substitution modelfor each data set (with MODELTEST pro-gram; Posada & Crandall 1998). In themodels selected, the number of different sub-stitution rates varied from two to six (HKY,Hasegawa et al. 1985; TrN, Tamura & Nei1993; GTR, Yang 1994), and in each case,the models were modified to assume a pro-portion of invariable sites (I) and a gamma-distributed substitution rate variation amongthe remaining sites (Γ). Genealogy of themtDNA haplotypes was assessed with MPusing NONA software version 1.5.1 (Golo-boff 1993–1997) or with MP, ML or MEusing PAUP* versions 4.0b8–10 (Swofford1998–2001).
Divergence time estimation. In the estima-tion of divergence times from mtDNA data,the molecular clock assumption has beenextensively used also in phylogeographicalstudies. However, as lineage-specific substi-tution rate variation appears to be a rathercommon phenomenon (e.g. Bromham et al.1996), the divergence times inferred fromDNA should be interpreted with caution (I,III–V). In most divergence time estimations
the rate range used was 0.5–1.3 % / Myr, assuggested for teleostean mtDNA in generalor for particular protein coding genes (forreferences, see V).
In addition to the overall divergence ratevariation among lineages, another confound-ing feature in the divergence time estimationis the varying ratio of non-synonymous tosynonymous substitutions observed, e.g.,among Baikalian cottoids (Kiril’chik &Slobodyanyuk 1997). As the molecular evo-lution at the synonymous sites is assumed tobe more neutral, i.e., not subject to varyingstrengths of purifying selection, the puta-tively more comparable synonymous PBLdistances (Li 1993; Pamilo & Bianchi 1993)were also used for the evaluation of diver-gence times in the study of the Baikaliancottoids (V).
Other analyses. Patterns of allele frequencyvariation in the North European C. gobiowere examined with principal componentanalysis (PCA) with SYSTAT version 8.0 (I).In the study of interlineage contact zone ofC. gobio (see Fig. 3), the differences in fourmorphometric characters were first testedamong and between reference samples rep-resenting the “pure” eastern, western andsoutheastern lineages using ANOVA andTukey´s test (II). Covariance analyses wereused to assess the dependence of these vari-ables on body length, and dependent vari-ables were either transformed or excludedfrom further statistical analyses.
The general patterns of molecular andmorphological variation among referenceand contact zone populations of C. gobiowere summarized with PCA, based on thesample means in three morphological char-acters, and on the frequencies of six molecu-lar characters. The clines within each riverin the northern study area and along thetransect in the southern study area (Fig. 3)were examined by plotting the scaled char-

7Phylogeography and evolution of freshwater cottid fishes
acter means against geographical distance(sample means equal to the value of the Wreference sample were scaled to 0.0, andthose equal to the value of the E referencesample to 1.0). In the northern study area,the coincidence and concordance of the clinesin different characters were also examinedand compared with a transect-independentapproach: Scaled character means were plot-ted against an average over characters (amean hybrid index, PC1) instead of geo-graphical distance (Kruuk 1997; Rohwer etal. 2001).
Several measures were used to examineassociations of characters within populationsin the C. gobio contact zones. The deviationof allozyme genotype frequencies from HWEwas tested within samples or subsamples byheterozygote deficiency tests, using GENE-POP version 3.1a (Raymond & Rousset1995) and GDA version 1.1 (Lewis & Zaykin2001). The genotypic associations betweenpairs of loci were estimated in terms of link-age disequilibrium D , and a standardized dis-equilibrium R was calculated from
pquvDR /= ,
where p, q, u and v are the allele frequenciesat the two loci (Szymura & Barton 1986). Inaddition, variances of morphological charac-ters, and Spearman correlation coefficientsr
S among all characters were estimated within
populations. To see whether the measures ofnon-random association between characterswere highest in the central hybrid zonepopulations, the estimates of R, heterozygotedeficit F
IS, variance and r
S were plotted
against PC1 value in the northern study areaor against distance in the southern study area.For further evaluation of possible trends,quadratic polynomials were fitted in the data,and the significance of quadratic terms wastested.
Results and discussion
Continent-scale phylogeography in northerncottid fishesOn the continental scale, the phylogeographicpatterns emerging from the mtDNA data wererather different among the three northerncottid complexes studied but some subdivi-sions were familiar from other fish or evenfrom terrestrial species (III and IV). In theC. poecilopus complex, the compositemtDNA haplotypes fell into four main line-ages (Fig. 4). Apart from a deeply divergentbasal Far Eastern branch (6 % sequence di-vergence), the most prominent division wasobserved within Europe (Fig. 5A). The NorthEuropean and Siberian mtDNA lineagesformed a widely distributed Euro-Siberianclade that differed from the Carpathian (Cen-tral Europe) lineage by over 4 % sequencedivergence (III).
The mtDNA haplotypes of the EuropeanC. gobio, Siberian C. sibiricus and NorthAmerican C. ricei clustered into a well-sup-ported monophyletic group in the molecularphylogeny (Fig. 4). Thus, the mtDNA dataagree with morphological evidence, whichhas recognized the gobio group amongHolarctic Cottus (Nybelin 1958; McAllister& Lindsey 1961; Koli 1969). In contrast tothe well-defined hierarchy of C. poecilopuslineages, the branching order within the C.gobio group, however, remained practicallyunresolved. Moreover, the Siberian andAmerican lineages appeared inseparablefrom a bunch of European C. gobio diversity(III), which comprises seven deep mtDNAlineages identified in earlier studies (Engl-brecht et al. 2000; Volckaert et al. 2002).
The divergences between the NorthAmerican and Eurasian lineages of the C.gobio group were only about one third ofthose observed across the Eurasian range in

8 Kontula
Fig. 4 General view on relation-ships among cottoid taxa ex-amined in I–V based on cyto-chrome b gene sequences. Thetree represents the topology andbranch lengths recovered fromME analysis (distances using themodel GTR+I+Γ). Numbers indi-cate bootstrap percentage sup-port values (values below 60 %are not shown). For more de-tailed analyses based on largerdata sets, and for MP or MLtrees, see the original papers.The taxa Scorpaena, Sebastesand Sebasticus represent an-other suborder, Scorpaenoidei(sequences from GeneBank;AF030711, AF030713 and AF-030768, respectively).

9Phylogeography and evolution of freshwater cottid fishes
the C. poecilopus complex. Yet, the Euro-Si-berian divergences were notably similar inboth Cottus complexes (1–2 %; Fig. 5), andindicated isolation since the Pliocene or Mid-dle Pleistocene (III). In terms of sequencedivergence, the observed Euro-Siberian re-lationships have actually been closer in otherstudied fishes. For instance, the divergencebetween the genetically closest European andSiberian haplotypes was only 0–0.5 % in theruffe Gymnocephalus cernuus (Stepien et al.1998), perch (Nesbø et al. 1999) and Arcticcharr Salvelinus alpinus (Brunner et al.2001). The difference between cottids andthese species may reflect weaker dispersalcapacities of cottids, or a higher potential ofsurviving in local refugia. In all, still only afraction of potential diversity of Asian Cottuscan be considered examined; as indicated bya study of similarly distributed Thymallus,the great Asian rivers may harbour much un-noticed phylogeographic diversity (Koskinenet al. 2003).
The principal split in the mtDNA geneal-ogy of the Myoxocephalus quadricornis com-plex separated the North American lacustrinedeepwater sculpin (M. q. thompsonii) fromthe Arctic marine and North European land-locked populations of the fourhorn sculpin
(M. q. quadricornis; Fig. 4; IV). This find-ing, both in respect of the pattern and of thegenetic divergence, is comparable to theEurasian-Beringian vs. continental NorthAmerican divisions e.g. in the Coregonuslavaretus whitefish complex (Bernatchez &Dodson 1994), and in the burbot Lota lota(Van Houdt et al. 2003) (Fig. 6). In the M.quadricornis complex, the North Americancontinental invasion appears to have takenplace several glaciation cycles ago in theEarly to Middle Pleistocene. In all, such es-timates of Myoxocephalus divergence arewell in line with traditional views on the ori-gin of the glacial relict element (e.g.Segerstråle 1962; Dadswell 1974), but devi-ate from the recent, molecular-based resultson glacial relict crustaceans, which haveshown much deeper subdivisions (Väinöläet al. 1994; 2001; Väinölä & Varvio 1989).
European cottid phylogeographyIn Europe, our analyses on Cottus poecilopusand C. gobio revealed various phylogeo-graphic subdivisions, some of which werecongruent among species. The deepestmtDNA divergences were found within C.poecilopus. The North European and Car-pathian lineages, which represent the disjunct
Fig. 5 The mtDNA lineage relationships in the Cottus poecilopus (A) and C. gobio (B) complexes, witha focus on European diversity. The trees are based on combined data set of cytochrome b and ATPase8 and 6 genes and represent parts of the mtDNA phylogeny presented in the comparative phylogeographicstudy of the northern cottids (III). Branches with strong support (BP = 90 % or higher) are indicated bythicker line.

10 Kontula
sub-ranges (Fig. 2A), appear to have divergedalready in the Pliocene or even Late Miocene(3–8 Myr ago; III). Such “intraspecific” di-vergences are rare, but not totally unparal-leled among other European fishes (cf.Durand et al. 1999). It should be noted thatat the time of this population split the ances-tral C. poecilopus may not have been livinganywhere near the Carpathians or NorthernEurope. If the ancestral C. poecilopus wascold-adapted as the modern one, the Plioceneclimate of Central Europe may have been toowarm for it (Haywood et al. 2000).
The mtDNA data also showed finerphylogeographic structuring, probably ofPleistocene origin, within both European
lineages of C. poecilopus (Fig. 5A; III). Inthe Carpathians, the haplotype relationshipsimplied a more recent connection over themountain range (i.e. Danube + Vistula vs.Dnestr) than through the lower reaches of therivers draining to the Black Sea. In NorthernEurope, on the other hand, the NorthFennoscandian haplotypes made a clusterdistinct from those found in Southern Fenno-scandia and Denmark (Figs. 5A and 7, seebelow).
The genetic diversity and population his-tory of the bullhead C. gobio has been thor-oughly studied using allozymes, mtDNA andmicrosatellites (Englbrecht et al. 2000;Hänfling et al. 2002; Volckaert et al. 2002;
Fig. 6 The principal phylogeographic subdivisions observed on a Holarctic scalein the Coregonus lavaretus whitefish complex (Bernatchez & Dodson 1994), inthe burbot Lota lota (Van Houdt et al. 2003), and in the Myoxocephalus quadri-cornis complex (IV). The mtDNA divergence between the major lineages is closeto 1 % in all three taxa.

11Phylogeography and evolution of freshwater cottid fishes
I–III). The mtDNA analyses have identifiedseven main lineages, labelled I–VII. The iden-tity of the main lineage found in eastern NorthEurope (E lineage of study I, in Finland, NERussia and Estonia), however, has not beenunambiguously resolved in the earlier stud-ies (cf. Englbrecht et al. 2000 and Volckaertet al. 2002). It was now named VIII to high-light its distinctive status (III). It is not clearwhether the major split within the EuropeanC. gobio divides the eastern lineage VIII fromthe other lineages (Figs. 4 and 5B; III), orwhether the principal division is between thecluster I + VIII (Danubian + eastern NorthEuropean lineages) and the more westernlineages (Volckaert et al. 2002). In any event,the first subdivision probably dates back toEarly Pleistocene – Late Pliocene ages, andis of the same magnitude as observed amongC. gobio, C. sibiricus and C. ricei on muchlarger geographical scale (Fig. 5B).
Postglacial colonization of Northern EuropeNorthern Europe was entirely glaciated untilapproximately 15 000 years ago, and theaquatic and terrestrial habitats were recol-onized postglacially. As to C. gobio, most ofthe Fennoscandian freshwater range seemsto have been invaded during the AncylusLake stage of the Baltic basin. The currentdistribution roughly corresponds to thedrainages accessible from that freshwaterlake (Koli 1969; Nybelin 1969). NorthernEurope was actually colonized from threedifferent directions by the distinct western,southeastern and eastern lineages (Fig. 7 andI). The North European western (I-W) andsoutheastern (II-SE) haplogroups of C. gobiorepresent the Danubian lineage (lineage I),whereas the lineage VIII probably stems froma stock that survived in or near the easternice-lakes in NW Russia (I, III). The princi-pal pattern in Fennoscandia is a strong east-west subdivision, which has even previouslybeen recognized in morphology (e.g. Koli
1969; Nybelin 1969). This distinction repre-sents a common biogeographic pattern inFenno-scandia; molecular data indicate dis-tinct eastern and western origins in the At-lantic salmon (Koljonen et al. 1999), gray-ling Thymallus thymallus (Koskinen et al.2000), a freshwater amphipod Gammaruslacustris (Vainio & Väinölä 2003), and inseveral mammals (Taberlet & Bouvet 1994;Jaarola et al. 1999).
In North European C. poecilopus, thenorthern and eastern haplotypes also clus-tered into an eastern mtDNA sub-lineage(NEur-E in Fig. 5A). Apart from indicatingtwo sources of postglacial colonization (seealso Andreasson 1972), this pattern also sug-gests glacial survival in northern CentralEurope, conceivably even between theScandinavian ice sheet and the northernCarpathian mountain slopes (cf. Hänfling etal. 2002).
Fig. 7 Mitochondrial DNA haplogroup distributionsof C. gobio and C. poecilopus in Northern Europe(circles and squares, respectively; I and III). Thearrows represent proposed routes of postglacialrecolonization for C. gobio. The hachured lineshows the maximum extent of the Late Weich-selian glaciation (c. 18 kya) (proglacial lake sys-tems and rivers according to Kvasov (1979) andArkhipov et al. (1986)).
12 Kontula
Secondary contact zones between Cottusgobio lineagesNorth of the Baltic Sea, the current distribu-tions of the western and eastern C. gobiogenomes show a clear-cut west-east distinc-tion in the upstream parts of the rivers (Fig.3 and II). During the postglacial recol-onization of northern waters, the E lineage(VIII) has obviously followed the easterncoast of the Baltic basin (Ancylus Lake) andcolonized the freshwaters as far as RiverKalixälven. Correspondingly, the W lineage(I-W) has immigrated along the westerncoastline and reached River Råneälven on thewest coast of the Bothnian Bay. At some pointafter the initial contact in the north, the Wlineage appears to have continued more suc-cessfully east than the E lineage towardswest, as detected from the mixed composi-tion of populations along the northeasternBaltic coasts (Koli 1969; I). Koli (1969) sug-gested that the W lineage was more tolerantof saltwater and therefore got an advantagein recolonization of Baltic habitats after themost saline Litorina Sea stage of the Balticbasin.
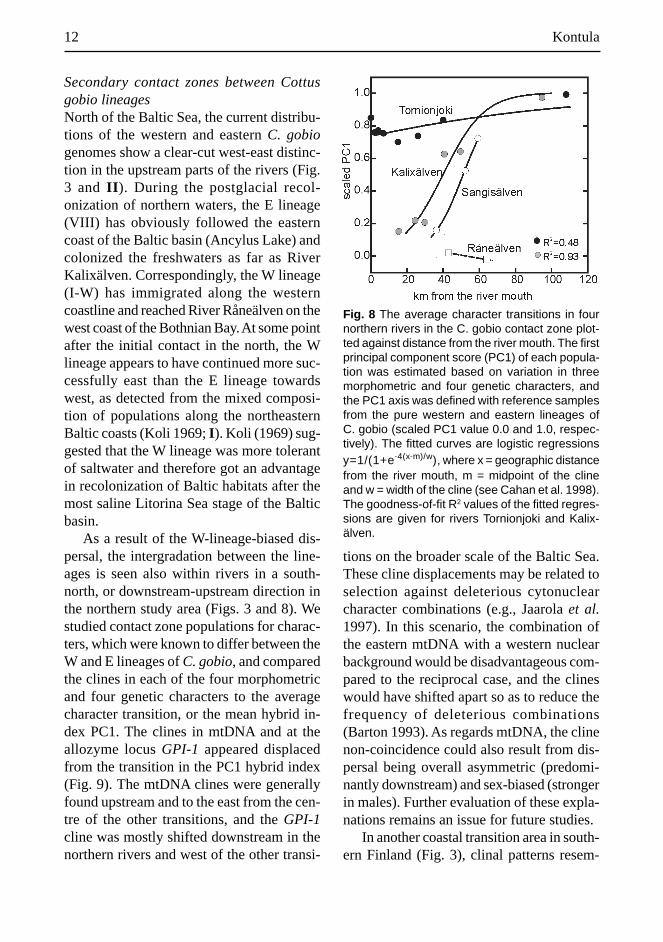
As a result of the W-lineage-biased dis-persal, the intergradation between the line-ages is seen also within rivers in a south-north, or downstream-upstream direction inthe northern study area (Figs. 3 and 8). Westudied contact zone populations for charac-ters, which were known to differ between theW and E lineages of C. gobio, and comparedthe clines in each of the four morphometricand four genetic characters to the averagecharacter transition, or the mean hybrid in-dex PC1. The clines in mtDNA and at theallozyme locus GPI-1 appeared displacedfrom the transition in the PC1 hybrid index(Fig. 9). The mtDNA clines were generallyfound upstream and to the east from the cen-tre of the other transitions, and the GPI-1cline was mostly shifted downstream in thenorthern rivers and west of the other transi-
tions on the broader scale of the Baltic Sea.These cline displacements may be related toselection against deleterious cytonuclearcharacter combinations (e.g., Jaarola et al.1997). In this scenario, the combination ofthe eastern mtDNA with a western nuclearbackground would be disadvantageous com-pared to the reciprocal case, and the clineswould have shifted apart so as to reduce thefrequency of deleterious combinations(Barton 1993). As regards mtDNA, the clinenon-coincidence could also result from dis-persal being overall asymmetric (predomi-nantly downstream) and sex-biased (strongerin males). Further evaluation of these expla-nations remains an issue for future studies.
In another coastal transition area in south-ern Finland (Fig. 3), clinal patterns resem-
Fig. 8 The average character transitions in fournorthern rivers in the C. gobio contact zone plot-ted against distance from the river mouth. The firstprincipal component score (PC1) of each popula-tion was estimated based on variation in threemorphometric and four genetic characters, andthe PC1 axis was defined with reference samplesfrom the pure western and eastern lineages ofC. gobio (scaled PC1 value 0.0 and 1.0, respec-tively). The fitted curves are logistic regressionsy=1/(1+e-4(x-m)/w), where x = geographic distancefrom the river mouth, m = midpoint of the clineand w = width of the cline (see Cahan et al. 1998).The goodness-of-fit R2 values of the fitted regres-sions are given for rivers Tornionjoki and Kalix-älven.

13Phylogeography and evolution of freshwater cottid fishes
bled those in the northern contact zone. How-ever, the southern area could not be inter-preted as a parallel of the northern one, aspopulation compositions could be explainedneither by in situ mixing of the eastern andwestern lineages (I-W and VIII), nor by mix-ing of the whole set of the three putativelypure North European lineages (I-W, II-SE orVIII).
Evolution of the Baikalian cottoid fishesOn the basis of mtDNA sequence data, wefound, for the first time, strong support forthe monophyly of the whole Baikalian cottoiddiversity (bootstrap support values in Fig. 4based on cyt-b only are fairly low, but for ananalysis with a larger data set, see V). In themolecular phylogeny, the Baikalian cottoidswere deeply nested within the Holarcticfreshwater Cottus, making the familyCottidae paraphyletic (see below). All but oneof the current Baikalian genera formed well-supported monophyletic groups, but the
branching order among them could not be re-solved completely. The short basal branchesin the phylogeny indicate rapid diversifica-tion early in the history of the species flock– a feature, which also compromised the in-ference about the major ecological leaps inthe Baikalian cottoid evolution, i.e., about thetransitions to pelagial and abyssal lifestyles.In any event, the rate of morphological evo-lution has been fast in Baikalian cottoids (seealso Kiril’chik et al. 1995; Kiril’chik &Slobodyanyuk 1997). The molecular diver-gences between Baikalian taxa are compar-able to those observed within the EurasianCottus species complexes (Fig. 4) but themorphological and ecological diversificationhas achieved quite different levels, as indi-cated by the assignment of Baikalian cottoidsinto three different families (Sideleva 2001).If the molecular calibrations are reliable, thediversification of the Baikalian cottoidsseems to have started at the earliest in theMiddle-to-Late Pliocene. As regards other
Fig. 9 Differences in character transitions in the northern contact zone (populations from all studiedrivers) between the eastern and western lineages of C. gobio (II). Frequencies of the eastern variants inmtDNA and at GPI-1 allozyme locus are plotted against scaled PC1 scores, which represent an aver-age character transition. The lines are fitted cubic polynomials y=x+2αx(1–x)+2βx(3x–2x2–1) forced topass through the points (0,0) and (1,1). Here, α represents twice the shift in the position of the transi-tion, and β is the decrease in the cline width below the average, both in units of average cline width (seeSzymura & Barton 1986). The goodness-of-fit R2 values are given for the curves.

14 Kontula
studied species flocks of Lake Baikal, themolluscan family Baicaliidae may be of simi-lar age with cottoids, whereas the flocks ofthe gammarid crustaceans and of the pulmo-nate Choanomphalus appear much older (forreview, see Sherbakov 1999).
Taxonomic remarksOur analyses on Cottus and Myoxocephalus,and on Baikalian cottoids have shown sev-eral cases, where molecular data disagreewith the conventional morphology-basedclassification, or otherwise indicate a needfor systematic revision. In the EurasianCottus poecilopus complex, the divergenceof the Far Eastern mtDNA lineage is excep-tionally high for an intraspecific comparison(c. 6 %), and probably deserves taxonomicrecognition (III). However, the previouslysuggested taxonomic units do not concur withobserved mtDNA clades (Fig. 4). For in-stance, Banarescu (1992) referred thepopulations of Far Eastern and Siberian“poecilopus” to C. szanaga Dybowski, butthe mtDNA phylogeny showed a clade com-prising the Siberian + North European C.poecilopus.
In European C. poecilopus, Witkowski(1984) has documented consistent morpho-logical differences between the northern andsouthern population groups, and he inter-preted these as a recent adaptive response tothe lake vs. river habitats of the populations.As shown by the deep mtDNA divergencewithin European C. poecilopus (Fig. 5A), thismorphological distinction may, however, berelated to remarkably old (c. 3 – 8 Myr)lineages warranting taxonomic recognition(III).
In the Cottus gobio complex, the distri-bution of the lineage VIII largely seems tocorrespond to that of a controversial EastEuropean bullhead (sub)species C. g.koshewnikowi Gratzianow, which differs
from the typical form in the length of the lat-eral line and in the prickling of skin (e.g, Berg1949; Nybelin 1958). Yet, as populations ofthe contact area in Northern Europe show alllevels of intergradation and genetic mixingbetween the eastern bullhead and the typicalform, a specific status has been denied, andeven subspecific treatment is difficult in thebroad overlap area (Koli 1969; I and II). In awider framework, the mtDNA divergence ofthis unit is still similar to the conventionallyrecognized vicariant species further east, C.sibiricus of Siberia and C. ricei of NorthAmerica (III). On the basis of mtDNA data,a higher than subspecies distinction for thelatter taxa does not seem warranted either.
As regards Myoxocephalus quadricorniscomplex, the morphological and genetic datafor geographically vicarious fourhorn (M. q.quadricornis) and deepwater (M. q. thomp-sonii) sculpins are congruent (Figs. 2C and6), and the recognition of the taxa in itselfshould not be challenged (IV). Yet, a fullspecific rank for these taxa, suggested e.g.by McAllister (1961; 1990), Bailey & Smith(1981), Parker (1988) and Eschmeyer (1998),would deviate from the practice recently pro-moted for similarly structured diversity inother Holarctic fish complexes (Bernatchez& Dodson 1994; Van Houdt et al. 2003).Therefore, we have preferred a subspeciesrather than species distinction, in accord withMcPhail & Lindsey (1970) and McAllister& Aniskowicz (1976). The generic affinitiesof the M. quadricornis complex also deservea comment. Several recent authors have refer-red the fourhorn and deepwater sculpins intotheir own genus Triglopsis Girard, 1852 (e.g.Fedorov 1986; Eschmeyer 1998). In themtDNA phylogeny, the M. quadricornis com-plex is deeply nested within the Myoxo-cephalus genus (Fig. 4 and IV), and accept-ing Triglopsis thus would make Myoxo-cephalus paraphyletic. Following McAllister

15Phylogeography and evolution of freshwater cottid fishes
(1961), Page & Burr (1991) and Kottelat(1997), we suggest that also the fourhorn anddeepwater sculpins should be retained inMyoxocephalus.
In the flock of Baikalian cottoids, the in-congruence between the molecular and sys-tematic subdivisions was also obvious. First,the suggested endemic families Abysso-cottidae and Comephoridae (e.g. Berg 1949;Taliev 1955; Sideleva 1982) were completelynested within Cottidae, or more specificallywithin the genus Cottus in the molecularphylogeny (Fig. 4 and V). Under phylo-genetic principles this indicates either a needfor further splitting of the family Cottidae orrecognition of the Baikalian diversity onlower taxonomic level. Second, as to thegroupings themselves, the genus Batracho-cottus has been placed within the subfamilyCottocomephorinae in Cottidae, whereas inthe mtDNA phylogeny it rather shows affin-ity with the taxa referred as abyssocottids.
Conclusions
In this thesis I studied the diversity and evo-lution of freshwater fishes in the suborderCottoidei, using mtDNA sequence data, allo-zymes or morphology, and material frommore than 40 taxa. Some important resultsand new, or at least rarely emphasized ideasemerging from the data are discussed below.
Much of the observed molecular diver-sity within cottid taxa appear to have its back-ground in the Pleistocene, probably in popu-lation subdivisions related to glacial events.However, this ‘genetic legacy of the Quater-nary ice ages’ (Hewitt 2000) does not accountfor all phylogeographic structuring observedwithin the northern cottid species complexes;the mtDNA analyses revealed also ancient,previously unnoticed subdivisions that ap-pear older than the Pleistocene (III).
It is also worth emphasizing that the ef-fects of Quaternary ice ages on biota appar-ently were more diverse than just devasta-tion of northern populations, or splitting thepopulations into isolated glacial refugia. Forcold-adapted species, such as the Alpinesculpin Cottus poecilopus, the glacial peri-ods could also have represented times ofrange expansions (Thienemann 1950, seealso Volckaert et al. 2002). Furthermore, gla-cial periods created freshwater connectionsacross present watersheds, e.g., vast ice-dammed lakes in central northern Russia(Arkhipov et al. 1995; Mangerud et al. 2001).Such, currently non-existing, connectionsacross central and western Palearctic prob-ably partly explain why subdivisions in manyspecies are deeper within Europe than be-tween Asian lineages and their European rela-tives. For instance, the European C. gobio“is one of the most highly structured fresh-water fish species” (Hänfling & Brandl1998), but on the Holarctic scale, the totaldivergence within the C. gobio complex ap-pears no greater than between neighbouringlineages in Europe (III).
The phylogeographic patterns among thestudied cottids are only partly congruent witheach other, but most of the identified evolu-tionary units find counterparts from other fishtaxa. Starting from the broadest scale, theEuro-Siberian clades of the C. gobio and C.poecilopus complexes deserve a notion (III).Similarly widespread, although somewhatyounger clades have been observed in vari-ous taxa from fish to rodents (e.g. Nesbø etal. 1999; Jaarola & Searle 2002). In Europe,one of the most commonly repeated phylo-geographic patterns appears to be the east-west subdivision around the Baltic Sea(Taberlet et al. 1998; Jaarola et al. 1999;Koskinen et al. 2000; I–III). The suture zonerecognized in northern Sweden on the basisof terrestrial phylogeography is notably close

16 Kontula
also to the observed contact zone in C. gobio.Yet, the role of the Baltic Sea is not ad-equately described as just a divider of post-glacial immigration routes. The large basinhas also enabled contact and mixing of dif-ferentiated units, and thus brought about new,and exceptionally high intraspecific geneticdiversity in these northern aquatic habitats.
The whole endemic diversity of theBaikalian cottoid species flock appearsmonophyletic from the mtDNA data (V). InBaikal, the cottoid radiation has produced 33currently recognized species with a widespectrum of morphologies. At similar timescales, the evolution of cottids in otherEurasian freshwater habitats could be ratherdescribed as morphological stasis; for in-stance, the Far Eastern and North EuropeanC. poecilopus have gained no obvious dif-ferences despite estimated divergence ofmore than 5 Myr. Such distinction in evolu-tionary rates reflects on one hand the powerof adaptive radiation related to invasions ofnew habitats (the oxygenated abyssal ofBaikal), and on the other hand, primarily sta-bilizing selection acting in the cottidpopulations that inhabit the shallow-waterenvironments of Eurasia. It is also noteworthythat the fish species radiation involvedparticularly cottoids, not other fishes of an-cient Baikalian faunas (e.g. Perca andRutilus; Sideleva 2000). The crucial differ-ence between an ancestral cottoid and otherfishes may have been the lack of swimmingbladder in cottoids, which could be seen as apreadap-tation for life in high-pressure envi-ronments.
Acknowledgements
First of all, I would like to thank my supervi-sor Risto Väinölä, who gave me the opportu-nity to start the PhD work in the “RelictGroup” in the last millennium. Ever since
Risto has always had time and energy to dis-sect my writings, which has undoubtedly im-proved the quality of the manuscripts. I amvery grateful to the former members of theRelict Group, Jouni Vainio, Jukka Palo andHannu Mäkinen, who have taught me every-thing I know about lab work. My warmestthanks also go to Sergei V. Kirilchik, withwhom we prepared the article about Bai-kalian cottoids, to Raisa Nikula for all thesupport she has kindly offered me during thisproject, and to Asta Audzijonyte, who hasalso helped me in many ways. Thank you allfor being such great colleagues, both on thegood days and on the days of the furiousläski-Queen.
I am indebted to more than 40 persons andseveral institutes for providing samples usedin this work. I thank particularly Sergei Ya.Slobodyanyuk and Marina E. Pavlova (Rus-sia), who provided most of the material forthe Baikal cottoid study, and Atso Romakka-niemi and Ari Saura from the Game and Fish-eries Research Institute (Finland), as well asIngemar Perä from the National Board ofFisheries (Sweden) for generously supplyingfish samples during the past few years.
I wish to thank the former and presentsequencing ladies of the MES lab, TupuKuismin and Anne Aronta for their pricelesscontribution in producing the sequence data.I thank also the other co-workers in the labfor creating such a pleasant, and sometimeseven hilarious working environment. I amgrateful to the Relict colleagues, and to LauriKoli, Craig Primmer, Kari Vepsäläinen andLotta Sundström for their helpful commentson the manuscripts. In the Elukka library,Sirkka-Liisa Nyeki deserves my sincere grati-tude for her excellent assistance in findingeven the weirdest literature needed in myresearch.
Back at the department Lotta Sundström,Ilkka Teräs and Hannu Pietiäinen have ar-ranged the practicalities related to the exami-

17Phylogeography and evolution of freshwater cottid fishes
nation of my thesis, and I thank them warmlyfor that. Jaakko Lumme and Maarit Jaarolaare gratefully acknowledged for the fast butthorough pre-examination of this thesis. Iwish to thank my friends Sanna Moisala andVappu Penttilä for their recent help — with-out their contribution the printed version ofthis thesis would not have come to exist. Spe-cial thanks also go to Minttu Hannonen, whohas been an invaluable advisor through thefinal stages of this project.
For several years, my friends and relativeshave had to listen to my rather pessimisticmonologues about this work. Thank you foryour patience and encouragement! Mymother Aino deserves special thanks for herassistance in the fieldwork; I would say thatcatching a bagful of Cottus by hand in +5º Cwater shows some attitude! Finally, I wantto give my greatest thanks to Eerika, my be-loved companion and best friend — she hassupported me fantastically throughout theproject but has also always been able to re-mind me that work is just a part of life, notthe whole.
The work was supported by grants fromthe Academy of Finland under the FinnishBiodiversity Research Program FIBRE, andby a grant from the Finnish Cultural Foun-dation.
References
Andreasson S (1972) Distribution of Cottuspoecilopus Heckel and C. gobio L. (Pis-ces) in Scandinavia. Zoologica Scripta,1, 69–78.
Arkhipov SA, Bespaly VG, Faustova MA,Glushkova OYu, Isaeva LL, Velichko AA(1986) Ice-sheet reconstructions. Quater-nary Science Reviews, 5, 475–483.
Arkhipov SA, Ehlers J, Johnson RG, WrightHEJ (1995) Glacial drainage towards theMediterranean during the Middle and
Late Pleistocene. Boreas, 24, 196–206.Avise JC (2000) Phylogeography: the his-
tory and formation of species. HarvardUniversity Press, Cambridge.
Avise JC, Arnold J, Ball RM, BerminghamE, Lamb T, Neigel JE, Reeb CA, SaundersNC (1987) Intraspecific phylogeography:the mitochondrial DNA bridge betweenpopulation genetics and systematics. An-nual Review of Ecology and Systematics,18, 489–522.
Bailey RM, Smith GR (1981) Origin andgeography of the fish fauna of the Lau-rentian Great Lakes basin. CanadianJournal of Fisheries and Aquatic Sci-ences, 38, 1539–1561.
Banarescu P (1990) Zoogeography of freshwaters, Vol. 1. AULA-Verlag, Wiesbaden.
Banarescu P (1992) Zoogeography of freshwaters, Vol. 2. AULA-Verlag, Wiesbaden.
Barton NH (1993) Why species and subspe-cies? Current Biology, 3, 797–799.
Berg LS (1949) Ryby presnykh vod SSSR isopredelnykh stran, vol. 3. IzdatelstvoAkademii Nauk SSSR, Moscow-Lenin-grad.
Bernatchez L (2001) The evolutionary his-tory of brown trout (Salmo trutta L.) in-ferred from phylogeographic, nestedclade, and mismatch analyses of mito-chondrial DNA variation. Evolution, 55,351–379.
Bernatchez L, Dodson JJ (1994) Phylogen-etic relationships among Palearctic andNearctic whitefish (Coregonus sp.) popu-lations as revealed by mitochondrial DNAvariation. Canadian Journal of Fisheriesand Aquatic Sciences, 51 (Suppl. 1), 240–251.
Bromham LD, Rambaut AE, Harvey PH(1996) Determinants of rate variation inDNA sequence evolution of mammals.Journal of Molecular Evolution, 43, 610–621.
Brunhoff C, Galbreath KE, Fedorov VB,

18 Kontula
Cook JA, Jaarola M (2003) Holarcticphylogeography of the root vole (Micro-tus oeconomus): implications for lateQuaternary biogeography of high lati-tudes. Molecular Ecology, 12, 957–968.
Brunner PC, Douglas MR, Osinov A, WilsonCC, Bernatchez L (2001) Holarctic phylo-geography of arctic charr (Salvelinusalpinus L.) inferred from mitochondrialDNA sequences. Evolution, 55, 573–586.
Cahan S, Helms KR, Rissing SW (1998) Anabrupt transition in colony behaviour inthe ant Messor pergandei. Animal Behav-iour, 55, 1583–1594.
Dadswell MJ (1974) Distribution, ecology,and postglacial dispersal of certain crus-taceans and fishes in eastern NorthAmerica. National Museum of NaturalSciences, Publications in Zoology, 11, 1–110.
Donner J (1995) The Quaternary History ofScandinavia. Cambridge UniversityPress, Cambridge.
Durand JD, Persat H, Bouvet Y (1999)Phylogeography and postglacial disper-sion of the chub (Leuciscus cephalus) inEurope. Molecular Ecology, 8, 989–997.
Ekman S (1953) Zoogeography of the Sea.Sidgwick & Jackson, London.
Englbrecht CC, Freyhof J, Nolte A, Rass-mann K, Schliewen U, Tautz D (2000)Phylogeography of the bullhead Cottusgobio (Pisces: Teleostei: Cottidae) sug-gests a pre-Pleistocene origin of the ma-jor central Euro-pean populations. Mo-lecular Ecology, 9, 709–722.
Eschmeyer WN (1998) Catalog of fishes.California Academy of Sciences, SanFrancisco.
Faustova MA (1984) Late Pleistocene gla-ciation of European USSR. In: Late Qua-ternary Environments of the Soviet Un-ion (ed. Velichko AA), pp. 3–12. Long-man, London.
Fedorov VV (1986) Cottidae. In: Fishes of
the North-eastern Atlantic and the Medi-terranean, vol. 3 (eds. Whitehead PJP,Bauchot M-L, Hureau J-C, Nielsen J,Tortonese E), pp. 1243–1260. UNESCO,Paris.
Froese R, Pauly D (eds) (2003) FishBase.World Wide Web electronic publication.www.fishbase.org, version 04 February2003.
Goloboff PA (1993–1997) NONA, Version1.5.1. Software distributed by the author,Tucuman, Argentina.
Hänfling B, Brandl R (1998) Genetic differ-entiation of the bullhead Cottus gobio L.across watersheds in Central Europe: evi-dence for two taxa. Heredity, 80, 110–117.
Hänfling B, Hellemans B, Volckaert FAM,Carvalho GR (2002) Late glacial historyof the cold-adapted freshwater fish Cottusgobio, revealed by microsatellites. Mo-lecular Ecology, 11, 1717–1729.
Hasegawa M, Iida Y, Yano T, Takaiwa F,Iwabuchi M (1985) Phylogenetic relation-ships among eukaryotic kingdoms in-ferred from ribosomal DNA sequences.Journal of Molecular Evolution, 22, 32–38.
Haywood AM, Sellwood BW, Valdes PJ(2000) Regional warming: Pliocene (3Ma) paleoclimate of Europe and the Medi-terranean. Geology, 28, 1063–1066.
Hewitt GM (1999) Post-glacial re-coloniza-tion of European biota. Biological Jour-nal of the Linnean Society, 68, 87–112.
Hewitt (2000) The genetic legacy of the Qua-ternary ice ages. Nature, 405, 907–913.
Houston J (1990) Status of the fourhornsculpin, Myoxocephalus quadricornis, inCanada. Canadian Field-Naturalist, 104,7–13.
Hunt DM, Fitzgibbon J, Slobodyanyuk SJ,Bowmaker JK, Dulai KS (1997) Molecu-lar evolution of the cottoid fish endemicto Lake Baikal deduced from nuclearDNA evidence. Molecular Phylogenetics

19Phylogeography and evolution of freshwater cottid fishes
and Evolution, 8, 415–422.Jaarola M, Searle JB (2002) Phylogeography
of field voles (Microtus agrestis) in Eura-sia inferred from mitochondrial DNA se-quences. Molecular Ecology, 11, 2613–2621.
Jaarola M, Tegelström H, Fredga K (1997) Acontact zone with noncoincident clines forsex-specific markers in the field vole (Mi-crotus agrestis). Evolution, 51, 241–249.
Jaarola M, Tegelström H, Fredga K (1999)Colonization history in Fennoscandian ro-dents. Biological Journal of the LinneanSociety, 68, 113–127.
Kiril’chik SV, Slobodyanyuk SYa, BelikovSI, Pavlova ME (1995) Phylogenetic re-latedness of 16 species of the Baikal LakeCottoidei bullhead fishes deduced frompartial nucleotide sequences of mtDNAcytochrome b genes. Molecular Biology,29, 471–476. Translated from Mole-kulyarnaya Biologiya, 29, 817–825.
Kiril’chik SV, Slobodyanyuk SYa (1997)Evolution of the cytochrome b gene frag-ment from mitochondrial DNA in someBaikalian and non-Baikalian Cottoideifishes. Molecular Biology, 31, 141–148.Translated from Molekulyarnaya Biolo-giya, 31, 168–175.
Koli L (1969) Geographical variation ofCottus gobio L. (Pisces, Cottidae) inNorthern Europe. Annales ZoologiciFennici, 6, 353–390.
Koljonen ML, Jansson H, Paaver T, VasinO, Koskiniemi J (1999) Phylogeographiclineages and differentiation pattern ofAtlantic salmon (Salmo salar) in the Bal-tic Sea with management implications.Canadian Journal of Fisheries andAquatic Sciences, 56, 1766–1780.
Koskinen MT, Ranta E, Piironen J, VeselovA, Titov S, Haugen TO, Nilsson J,Carlstein M, Primmer CR (2000) Geneticlineages and postglacial colonization ofgrayling (Thymallus thymallus, Salmo-
nidae) in Europe, as revealed by mito-chondrial DNA analyses. Molecular Ecol-ogy, 9, 1609–1624.
Koskinen MT, Knitzhin I, Primmer CR,Schlötterer C, Weiss S (2002) Mitochon-drial and nuclear DNA phylogeographyof Thymallus spp. (grayling) provides evi-dence of ice-age mediated environmentalperturbations in the world´s oldest bodyof freshwater, Lake Baikal. MolecularEcology, 11, 2599–2611.
Kottelat M (1997) European freshwaterfishes. Biologia, 52 (Suppl. 5), 1–271.
Kozhova OM, Izmest’eva LR (eds.) (1998).Lake Baikal. Evolution and biodiversity.Backhuys Publishers, Leiden.
Kruuk LEB (1997) Barriers to gene flow: aBombina (fire-bellied toad) hybrid zoneand multilocus cline theory. UnpublishedPh.D. Thesis, University of Edinburgh.
Kvasov DD (1979) The Late-Quaternary his-tory of large lakes and inland seas of east-ern Europe. Annales Academiae Scien-tiarum Fennicae, Series A, 127, 1–71.
Lewis PO, Zaykin D (2001) Genetic DataAnalysis: Computer program for theanalysis of allelic data. Version 1.1. Freeprogram distributed by the authors overthe internet from http://lewis.eeb.uconn.edu/lewishome/software.html.
Li W-H (1993) Unbiased estimation of therates of synonymous and nonsynonymoussubstitutions. Journal of Molecular Evo-lution, 36, 96–99.
Mangerud J, Astakhov V, Jakobsson M,Svendsen JI (2001) Huge ice-age lakes inRussia. Journal of Quaternary Science,16, 773–777.
Mats VD (1993) The structure and develop-ment of the Baikal rift depression. Earth-Science Reviews, 34, 81–118.
McAllister DE (1961) The origin and statusof the deepwater sculpin, Myoxocephalusthompsonii, a Nearctic glacial relict. Na-tional Museum of Canada Bulletin, 172,

20 Kontula
44–65.McAllister DE (1990) A list of the fishes of
Canada. National Museum of Natural Sci-ences Syllogeus, 64, 1–310.
McAllister DE, Aniskowicz J (1976) Verte-bral number in North American sculpinsof the Myoxocephalus quadricornis-com-plex. Journal of the Fisheries ResearchBoard of Canada, 33, 2792–2799.
McAllister DE, Lindsey CC (1961) System-atics of the freshwater sculpins (Cottus)of British Columbia. National Museum ofCanada Bulletin, 172, 66–88.
McPhail JD, Lindsey CC (1970) Freshwaterfishes of Northwestern Canada andAlaska. Fisheries Research Board ofCanada Bulletin 173.
Nei M, Kumar S (2000) Molecular evolu-tion and phylogenetics. Oxford Univer-sity Press, Oxford.
Nesbø CL, Fossheim T, Vollestad LA,Jakobsen KS (1999) Genetic divergenceand phylogeographic relationships amongEuropean perch (Perca fluviatilis) popula-tions reflect glacial refugia and postglacialcolonization. Molecular Ecology, 8,1387–1404.
Nilsson J, Gross R, Dove O, Jansson H, Kohl-mann K, Löytynoja A, Nielsen EE, PaaverT, Penttilä H, Primmer CR, Vasemägi A,Veselov A, Öst T, Lumme J (2001) Matri-linear phylogeography of Atlantic salmon(Salmo salar L.) in Europe and postglacialcolonization of the Baltic Sea area. Mo-lecular Ecology, 10, 89–102.
Nybelin O (1958) Några resultat av naturhis-toriska museets undersökningar översvenska sötvattenssimpor. GöteborgsMusei Årstryck, 1958, 32–56.
Nybelin O (1969) Om stensimpans, Cottusgobio L., och bergsimpans, C. poecilopusHeckel, utbredningshistoria i södra ochmellersta Sverige. Acta Regiae SocietatisScientiarum et Litterarum Gothobur-gensis, 4, 1–52.
Page LM, Burr BM (1991) A field guide tofreshwater fishes of North America northof Mexico. Houghton Mifflin Company,Boston.
Pamilo P, Bianchi NO (1993) Evolution ofthe Zfx and Zfy genes: rates and inter-dependence between the genes. Molecu-lar Biology and Evolution, 10, 271–281.
Parker BJ (1988) Status of the deepwatersculpin, Myoxocephalus thompsoni, inCanada. Canadian Field-Naturalist, 102,126–131.
Posada D, Crandall KA (1998) MODEL-TEST: testing the model of DNA substi-tution. Bioinformatics, 14, 817–818.
Raymond M, Rousset F (1995) GENEPOP(version 1.2): population genetics soft-ware for exact tests and ecumenism. Jour-nal of Heredity, 86, 248–249.
Rohwer S, Bermingham E, Wood C (2001)Plumage and mitochondrial DNA haplo-type variation across a moving hybridzone. Evolution, 55, 405–422.
Segerstråle SG (1962) The immigration andprehistory of the glacial relicts of Eurasiaand North America. A survey and discus-sion of modern views. Internationale Re-vue der gesamten Hydrobiologie, 47, 1–25.
Segerstråle SG (1982) The immigration ofglacial relicts into Northern Europe in thelight of recent geological research.Fennia, 160, 303–312.
Sherbakov DYu (1999) Molecular phylogen-etic studies on the origin of biodiversityin Lake Baikal. Trends in Ecology andEvolution, 14, 92–95.
Sideleva VG (1982) Seismosensornayasistema i ekologiya baikalskikh podka-menshchikovykh ryb (Cottoidei). Nauka,Novosibirsk.
Sideleva VG (1994) Speciation of endemicCottoidei in Lake Baikal. In: Speciationin ancient lakes (eds Martens K, God-deeris B, Coulter G). Arch. Hydrobiol.

21Phylogeography and evolution of freshwater cottid fishes
Beih. Ergebn. Limnol., 44, 441–450.Sideleva VG (2000) The ichtyofauna of Lake
Baikal, with special reference to its zoo-geographical relations. Advances in Eco-logical Research, 31, 81–96.
Sideleva VG (2001) List of fishes from LakeBaikal with descriptions of new taxa ofcottoid fishes. In New contributions tofreshwater fish research (eds PugachevON, Balushkin AV), Proceedings of thezoological institute, 287, 45–79, Zoologi-cal Institute RAS, St. Petersburg.
Sideleva VG (2002) Cottus poecilopusHeckel, 1836 – pestronogii podkamennik.In: Atlas presnovodnych ryb rosii (Atlasof Russian freshwater fishes) (ed Reshet-nikov IuS), pp. 166–168. Nauka, Moscow.
Stepien CA, Dillon AK, Chandler MD (1998)Genetic identity, phylogeography, andsystematics of ruffe Gymnocephalus inthe North American Great Lakes andEurasia. Journal of Great Lakes Research,24, 361–378.
Swofford DL (1998–2001) PAUP*: Phyloge-netic analysis using parsimony (and othermethods). Sinauer Associates, Sunder-land, MA.
Szymura JM, Barton NH (1986) Geneticanalysis of a hybrid zone between the fire-bellied toads, Bombina bombina and B.variegata, near Cracow in southern Po-land. Evolution, 40, 1141–1159.
Taberlet P, Bouvet J (1994) MitochondrialDNA polymorphism, phylogeography,and conservation genetics of the brownbear Ursus arctos in Europe. Proceedingsof the Royal Society of London B, 255,195–200.
Taberlet P, Fumagalli L, Wust-Saucy A-G,Cosson J-F (1998) Comparative phylo-geography and postglacial colonizationroutes in Europe. Molecular Ecology, 7,453–464.
Taliev DN (1955) Bychki-podkamenshchikiBaikala (Cottoidei). Izdatelstvo Akademii
Nauk SSSR, Moscow - Leningrad.Tamura K, Nei M (1993) Estimation of the
number of nucleotide substitutions in thecontrol region of mitochondrial DNA inhumans and chimpanzees. Molecular Bi-ology and Evolution, 10, 512–526.
Thienemann A (1950) Verbreitungsge-schichte der Süsswassertierwelt Europas.Die Binnengewässer 18, Stuttgart.
Timoshkin OA (1997) Biodiversity of Baikalfauna: state-of-the-art (preliminary analy-sis). In: New scope on boreal ecosystemsin East Siberia (eds Wada E, TimoshkinOA, Fujita N, Tanida K), DIWPA series,2, 35–76, Siberian Branch Russian Acad-emy of Sciences, Novosibirsk.
Vainio JK, Väinölä R (2003) Refugial racesand postglacial colonization history of thefreshwater amphipod Gammarus lacustrisin Northern Europe. Biological Journalof the Linnean Society, 79, 523–542.
Väinölä R, Varvio SL (1989) Molecular di-vergence and evolutionary relationshipsin Pontoporeia (Crustacea: Amphipoda).Canadian Journal of Fisheries andAquatic Sciences, 46, 1705–1713.
Väinölä R, Riddoch BJ, Ward RD, Jones RI(1994) Genetic zoogeography of theMysis relicta species group (Crustacea:Mysidacea) in northern Europe and NorthAmerica. Canadian Journal of Fisheriesand Aquatic Sciences, 51, 1490–1505.
Väinölä R, Vainio JK, Palo JU (2001)Phylogeography of “glacial relict” Gam-maracanthus (Crustacea, Amphi-poda)from boreal lakes and the Caspian andWhite seas. Canadian Journal of Fisher-ies and Aquatic Sciences, 58, 2247–2257.
Van Houdt JK, Hellemans B, Volckaert FAM(2003). Phylogenetic relationships amongPalearctic and Nearctic burbot (Lota lota):Pleistocene extinctions and recolo-nization. Molecular Phylogenetics andEvolution, in press.
Volckaert FAM, Hänfling B, Hellemans B,

22 Kontula
Carvalho GR (2002) Timing of the popu-lation dynamics of bullhead Cottus gobio(Teleostei: Cottidae) during the Pleisto-cene. Journal of Evolutionary Biology, 15,930–944.
Witkowski A (1979) A taxonomic study onfreshwater sculpins of genus CottusLinnaeus, 1758 (Cottus gobio L. andCottus poecilopus Heck.) in Poland. ActaUniversitatis Wratislaviensis, 458, 1–95.
Witkowski A (1984). Morphological variabil-ity in lake and river populations of Cottuspoecilopus Heckel, 1836 (Pisces: Cotti-lkdae). Acta Ichthyologica et Piscatoria,XIV, 43–57.
Yang Z (1994) Maximum likelihood phylo-genetic estimation from DNA sequenceswith variable rates over sites: approximatemethods. Journal of Molecular Evolution,39, 306–314.





























