Embed Size (px)
Citation preview

Phylogenetic Utility of Elongation Factor-1 a! in Noctuoidea (Insecta: Lepidoptera): The Limits of Synonymous Substitution
Andrew Mitchell, * Soowon Cho, * Jerome C. Regier, t Charles Mitter, * Robert W. Poole, $ and Marcus Matthews5 *Department of Entomology, and TCenter for Agricultural Biotechnology, University of Maryland; $Systematic Entomology Laboratory, USDA-ARS, % National Museum of Natural History; and $Division of Entomology, CSIRO, Canberra, Australia
To test its phylogenetic utility, nucleotide sequence variation in a 1,240-bp fragment of the elongation factor-la (W-la) gene was examined in 49 moth species representing the major groups of the superfamily Noctuoidea. Both parsimony and distance analyses supported the monophyly of nearly all groups for which there are clear morpho- logical synapomorphies. Clades of subfamily rank and lower, probably mid-Tertiary and younger, were strongly supported. The third codon position contains 88% of variable sites, and approaches saturation at -20% sequence divergence, possibly due to among-site rate heterogeneity and composition bias; higher divergences occur only in association with shifts in composition. Surprisingly, the few nonsynonymous changes appear no more phylogenet- ically reliable than synonymous changes. Signal strength for basal divergences is weak and fails to improve with character weighting; thus, dense taxon sampling is probably needed for strong inference from EF-lcu regarding deeper splits in Noctuoidea (probably early Tertiary). EF-lcu synonymous changes show promise for phylogeny reconstruction within Noctuidae and other groups of Tertiary age.
Introduction
In the quest for concordance among multiple, in- dependent lines of evidence (Miyamoto and Cracraft 1991; Avise 1994), molecular systematists are beginning to exploit the nuclear genome. Although several protein- encoding nuclear genes have recently been identified as having potential utility for molecular systematics (Fried- lander, Regier, and Mitter 1992, 1994; Palumbi and Ba- ker 1994; Slade, Moritz, and Heideman 1994; Gupta 1995; Waters 1995; Friedlander et al. 1996; Orti and Meyer 1996; Fang et al. 1997), for none of these has the temporal range of utility been characterized ade- quately. Recently, Cho et al. (1995) investigated the util- ity of the nuclear gene encoding elongation factor-la (EF-la) protein for resolving relationships within a sub- family of noctuid moths.
In this study, we further characterized the utility of EF-ICX by extending the observations of Cho et al. (1995) to a broader sample across the superfamily Noc- tuoidea. The phylogenetic utility of the gene, patterns of substitution, and potential sources of homoplasy were evaluated by comparison of the gene trees to indepen- dently derived phylogenies based on morphology. Fi- nally, we examined the implications of our results for unsolved problems in the phylogeny of Noctuoidea.
Materials and Methods Specimens Examined
The species sampled, their taxonomic affinities, and their geographic sources are listed in table 1, along with GenBank accession numbers for their EF-la sequences. Forty of the 49 sequences are new to this study, with the remaining nine from Cho et al. (1995). Taxon sam-
Key words: molecular systematics, synonymous substitutions, Noctuoidea, Noctuidae.
Address for correspondence and reprints: Andrew Mitchell, De- partment of Entomology, University of Maryland, College Park, Mary- land 20742. E-mail: [email protected].
Mol. Biol. Evol. 14(4):381-390. 1997 0 1997 by the Society for Molecular Biology and Evolution. ISSN: 0737-4038
pling was most dense (35 species) in “trifine” Noctui- dae (see below), which include the subfamily Heliothi- nae, studied by Cho et al. (1995). The remaining 14 species were chosen to span the diversity of the super- family. Our sample included all of the 4 traditional noc- tuoid families, 15 of the 18 noctuid subfamilies, and 24 of the 38 noctuid tribes found in North America (Fran- clement and Todd 1983).
Field-collected adult moths were either frozen at - 80°C or placed in 100% ethanol at -20 to 0°C and transferred to -80°C after 2-6 months. The wings were kept as dried vouchers at the Department of Entomol- ogy, University of Maryland at College Park. In most cases, abdomens were kept at -8O”C, both as tissue vouchers and to facilitate subsequent species identifi- cation.
Comparative Framework
Phylogenetic relationships within Noctuoidea have been problematic (Kitching 1984), but recent work has identified a number of probable monophyletic groups, for which Poole (1995) and/or Kitching and Rawlins (1997) cite at least one synapomorphy. We will refer to these as “concordance groups.” We sampled multiple representatives from most such groups, and used recov- ery of concordance groups as one gauge of phylogenetic utility for EF-la. Figure 1 illustrates recent hypotheses of noctuoid relationships examined in this study; con- cordance groups are indicated by asterisks. Figure lA, redrawn from Weller et al. (1994), shows a widely ac- cepted hypothesis of relationships among families of Noctuoidea. Within Noctuidae, recent reviews support two groups, “trifines” and “quadrifines,” of which only the former is monophyletic (Poole 1995; Speidel and Naumann 1995; Kitching and Rawlins 1997). Poole (1995) used a broad definition of trifines, and combined the “true cutworms” of Lafontaine (1993) into an ex- panded subfamily Noctuinae s.l., containing most of the trifine species (fig. 1B). Kitching and Rawlins (1997) used a more restricted definition of trifines, excluding
381
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019

382 Mitchell et al.
Table 1 Taxa Sampled
Higher Taxon Species Sourcea (voucher #) GenBank
Accession #
Notodontidae Notodontinae ............. Phalerinae ................ Nystaleinae ............... Heterocampinae ...........
Arctiidae
Lithosiinae ............... Arctiinae .................
Lymantriidae . . . . . . . . . . . . . . .
“Quadrifine” Noctuidae
Euteliinae ................ Nolinae .................. Sarrothripinae ............. Catocalinae ............... Herminiinae ..............
“Trifine” Noctuidae
Eustrotiinae .............. Acontiinae ...............
Condicinae ............... Pantheinae ...............
Oncocnemidinae .......... Plusiinae .................
Acronictinae ..............
Raphiinae ................ Agaristinae ...............
Stiriinae .................
Amphipyrinae. ............ Cuculliinae ............... Psaphidinae ..............
Noctuinae s.1. “Caradrinini” ...........
Hadenini ...............
Xylenini S.S. ............
Noctuini. ...............
Heliothinae . . . . . . . . . . . . . . .
Furcula cinerea Maryland, College Park (43) U85665 Datana perspicua Maryland, Beltsville (44) II85666 Symmerista albifrons Maryland, Beltsville (172) U85667 Schizura sp. Maryland, Beltsville (16) U85668
Hypoprepia miniata Estigmene acreab
West Virginia, Mathias (79) Maryland, Beltsville (45) Maryland, College Park (115) Maryland, University Park (155) Lab. colony, University of Maryland (92) Maryland, Clarksville (56)
U85669 U85670
Hyphantria cunea Lymantria dispar Dasychira sp.
U8567 1 U85672 U85673
Paectes pygmaea Maryland, Clarksville (67) U85674 Meganola sp. Maryland, Clarksville (25) U85675 Baileya levitans Maryland, Clarksville (170) U85676 Catocala ultronia Maryland, Clarksville (22) U85677 Palthis sp. Maryland, College Park (78) U85678
Thioptera nigrofimbria Spragueia leo Tarachidia candefacta Condica videns Charadra deridensb
Maryland, Beltsville (26) Maryland, Fort Meade (157) Maryland, Beltsville (12) Maryland, Rockville (47) Maryland, Beltsville (168) West Virginia, Mathias (163) Maryland, Beltsville (165) Maryland, Beltsville (169) Texas, El Paso (58) Maryland, Beltsville (15) Lab. colony, University of Maryland (92) Maryland, Beltsville (166) Maryland, Clarksville (21) Maryland, Fort Meade (158) Maryland, Clarksville (10) Maryland, Clarksville ( 150) Texas (84) Texas, El Paso (126) Maryland, Fort Meade (162) Texas (156) West Virginia, Mathias (154) Texas, Tyler County (59)
U85679 U85680 II8568 1 U85682 U85683
Panthea furcillab U85684
Oncocnemis obscurata Anagrapha falcifera Trichoplusia ni” Acronicta sp. Polygrammate hebraeicum Raphia abrupta Eudryas grata Psychomorpha epimenis Stiria rugifrons Basilodes chrysopiti Amphipyra pyramidoides Cucullia convexipennis Psaphida resumens Feralia major
U85685 U85686 U20140 U85687 U85688 U85689 U85690 U8569 1 U85692 U20125 U85693 U85694 II85695 U85696
Spodoptera frugiperdabc
Elaphria grata Nedra ramulosa Orthodes crenulata Lacinipolia renigera Lithophane hemina Anathix ralla Anicla infecta Agrotis ipsilonb
Adisura bellaC Helicoverpa zeaC Schinia arcigerac Pyrrhia exprimensC Eutricopis nexilis” Heliothodes diminutivusC
Lab. colony, University of Maryland (94) Lab. colony, USDA, Georgia (175) Maryland, College Park (23) Maryland, Rockville (27) Maryland, Rockville (55) Maryland, Rockville (24) West Virginia, Mathias (15 1) Maryland, Clarksville (152) Maryland, College Park (160) Maryland, College Park (8) Maryland, Clarksville (159) MALI, Mourdiah (199) Lab. colony, USDA, Mississippi (87) Texas, Houston (118) Maryland (38) California, Sonora Pass (116) California, Cobb Mountain (117)
U20139
U85697 U85698 U85699 U85700 U85701 U85702 U85703 U85704
U20123 U20136 U20138 U20137 U20126 U20130
a All taxa collected in the U.S.A., except Adisuru bella (collected in Mali). b Two specimens are listed. The first was used to discover the entire 1,240-bp
variation. c Published sequences (Cho et al. 1995).
sequence; the was sequenced (400-l ,200 bp) to assess intraspecific
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019

A
EF-la Synonymous Changes Track Noctuoid Phylogeny 383
C Oenosandridae
Doidae
Notodontidae *
Arctiidae*
Lymantriidae*
Noctuidae
Acontiinae S.S. *
Amphipyrinae S.S.
Eriopinae
Noctuini * - Xylenini s.s?
Antitypini
Hadenini Apameini
“Caradrininl’_
1 r Psaphidinae*
Noctuidae
Noctuinae s.1. *
Lymantriidae* Pantheidae
-i!
Acronictinae Bryophilinae Raphiinae Herminiinae Q Catocalinae Q Calpinae Q Hypeninae Q Aganainae Cocytiinae : Strepsimaniinae Q
Euteliinae Q Stictopterinae Q
Plusiinae * Agaristinae * Eucocytiinae Acontiinae Eustrotiinae Bagisarinae Cuculliinae Dilobinae Condicinae
t
Agaristinae * Heliothinae* / Stiriinae* Acronictinae “Trifines” * Heliothinae * Pantheinae Glottulinae Bryophilinae Hadeninae Oncocnemidinae Ufeinae
*J Cuculliinae S.S. Noctuinae *
FIG. l.-Recent views on phylogeny of the Noctuoidea. Taxa not sampled in this study are in italics. Asterisks denote “concordance groups,” defended by morphological synapomorphies. A, Relationships among families, after Weller et al. (1994). B, Relationships among redefined subfamilies of “Mine” Noctuidae, modified from Poole (1995). “Caradrinini” refers to a heterogeneous set of genera removed from Amphipyrinae by Poole (1995) but not named by him. C, Relationships among families and within Noctuidae, after Kitching and Rawlins (1997). “Q” denotes taxa traditionally regarded as “quadrifine” noctuids.
Acronictinae and associated taxa from this group, and excluding subfamilies Nolinae and Pantheinae from Noctuidae (fig. 1C).
Laboratory Protocols
Methods of DNA extraction, PCR amplification, and DNA sequencing, and EF-Icu primer sequences, fol- lowed Cho et al. (1995). A nested PCR approach was frequently used to obtain purer templates: the initial am- plification used primers M3 and rcM4, yielding a 1,286-bp fragment, while subsequent amplifications used internal primers to yield double-stranded DNA fragments of less than 600 bp in length. All new se- quences were generated using an Applied Biosystems 373A Stretch DNA sequencer. For each taxon, 1,240 bp of the EF-Ia coding region, representing 89% of the total, were sequenced. This region corresponds to nu- cleotides 89-1328 of the Bombyx mori IF-la cDNA sequence (Kamiie et al. 1993; GenBank accession num- ber D13338). Both DNA strands were sequenced. No introns were detected.
For five species, the W-la! gene was sequenced from a second specimen to assess intraspecific variation (see table 1). Only one variable site was observed, a
synonymous difference (C or T at a third codon posi- tion) in the 510-bp comparison within Spodopteru fru- giperda.
Data Analyses
Sequence chromatograms were checked for accu- racy of base-calling using the editor TED, distributed with the software program STADEN (Staden 1992). Se- quences were assembled and aligned manually using the Genetic Data Environment software package (GDE 2.2, Smith et al. 1994). Alignments did not require gaps.
Calculation of uncorrected pairwise divergences and all parsimony analyses were performed using a test version of PAUP* (PAUP 4.0d26; D. Swofford, per- sonal communication) for Macintosh PowerPC. Each parsimony analysis implemented a heuristic search, us- ing TBR branch swapping, with random-addition se- quence for taxa, and 1,000 replications per search. The Notodontidae were used as the outgroup, as there is strong morphological evidence for their basal position in the Noctuoidea relative to the other taxa sampled (Miller 1991). Both decay indices (Bremer 1988; Don- oghue et al. 1992) and bootstrap proportions in 1,000
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019

384 Mitchell et al.
bootstrap replications were used to evaluate support for nodes.
Additional parsimony analyses applied unequal weighting schemes, involving codon position, transi- tions versus transversions, and synonymous versus non- synonymous substitutions. A range of differential weights was applied, including 2 X , 4X, and 10X up- weighting of the partition of interest; for example, first : second : third codon positions were weighted 2: 1: 1, 4: 1: 1, 10: 1: 1, 1:2: 1, etc. Third codon positions and synon- ymous substitutions were also analyzed alone. The “synonymous change” data set was determined by ex- cluding from the original data set all sites in codons for which an amino acid substitution occurred in any taxon.
MacClade (version 3; Maddison and Maddison 1992) was used to produce charts for character analysis and to derive summary statistics. Codon usage and base composition by codon position were determined using the XNIP program in the STADEN package (Staden 1992). Chi-square tests for homogeneity of base com- position among taxa were performed using PAUP* (PAUP 4.0d29; D. Swofford, personal communication). Among-site heterogeneity in evolutionary rate was test- ed by chi-square goodness-of-fit to the Poisson distri- bution for number of changes per site on the most par- simonious tree (Tamura and Nei 1993), with cells of expected frequency less than five being combined.
Neighbor-joining trees (Saitou and Nei 1987) were calculated for comparison based on the Kimura two-pa- rameter model (Kimura 1980) and on the log determi- nant (LogDet) distance, which corrects for unequal base composition (Lockhart et al. 1994), as programmed in PAUP* (PAUP 4.0d29; D. Swofford, personal com- munication). Following Lockhart et al. (1994), a quali- tative test was also performed to detect whether a bias in base composition at third codon positions could be affecting the topology of the parsimony tree, i.e., a neighbor-joining tree was constructed from Euclidean distances of third codon position base proportions, using the program NTSYS-pc (Rohlf 1990), and compared to distance trees derived from the original data.
Results and Discussion Gene Trees and Concordance with Morphology
Equally weighted parsimony analysis of all substi- tutions produced a single most-parsimonious (mp) tree (fig. 2). Tree statistics on the mp tree for all sites, in- dividual codon positions, and amino acids are given in table 2.
The mp tree (fig. 2) recovered 14 of 15 concor- dance groups with generally strong support, indicating that EF-Ia! contains much information about noctuoid phylogeny. Support was especially robust at the tribal and subfamily levels, for which 7 of 10 concordance groups have bootstrap values of 95%-100%. Recovery of the few morphologically supported relationships above the subfamily level indicates that EF-la carries information about older divergences as well, although the latter are weakly supported.
Signal strength decreased with hierarchical depth on the mp tree; of the 24 nodes subtending multiple subfamilies, 23 had bootstrap support less than 50%. Consistent with lesser support for deeper nodes, some topologies a few steps longer than the mp tree were considerably different from the mp tree. For example, among trees one step longer (15 total), three types of departure from the mp tree were discernible: (1) rear- rangements involving nearby noctuids only; (2) out- group species (Notodontidae) not monophyletic with re- spect to noctuids; and (3) the Arctiidae, Lymantriidae, and associated “quadrifine” noctuids falling within tri- fines, rendering the latter paraphyletic. Trees two steps longer (160 total) included placement of three of the four notodontids as sister group to the noctuid subfamily Heliothinae. There were 1,272 trees that were three steps longer.
Results of neighbor-joining (nj) analysis under the Kimura two-parameter distance (Kimura 1980) were similar to those of parsimony analysis (fig. 3; thick lines identify groups recovered by both methods). The nj tree supports one less concordance group (Arctiidae), with similarly high bootstrap values for shallower nodes. At deeper levels, the nj and mp trees differ only in nodes weakly supported under parsimony. Like the mp tree, the nj tree shows weak support for groupings above sub- family rank, except that the monophyly of Arctiidae + Lymantriidae has a bootstrap value of 80% on the nj tree.
Sequence Variability
Variable sites were distributed uniformly across broad regions of the 1,240 bp, with only the first ap- proximately 100 sites being more conserved (plot not shown; see fig. 3 in Cho et al. 1995). Heterogeneity in variability among codon positions is striking (table 2). Third codon positions alone account for 94% of the mp tree length and 88% of all variable sites, while first po- sitions contribute 8% of variable sites and second po- sitions contribute only 4%. Table 2 shows that second codon positions are also the least homoplasious data set (retention index [RI] = 0.52), with other sites, and ami- no acids, about equal (RI = 0.42-0.44). Most of the few amino acids that are free to change do so too rapidly to be phylogenetically informative. In contrast, Cho et al. (1995) mapped amino acids onto their mp tree based on all nucleotide sites, with RI = 1.
Heterogeneity of evolutionary rate is also marked within codon positions (fig. 4). At third codon positions, for example, the null hypothesis of a Poisson distribu- tion for numbers of steps on the mp tree is strongly rejected (chi-square = 1,733, df = 9, P < 0.001). The mean number of steps per character for third positions is 4.35, but 79 sites (= 19.1% of total) are invariant, while two sites contribute 17 steps each. The distribu- tions for both first and second positions, and for amino acids, are even more strongly right-skewed.
Saturation of Substitutions Approach to saturation (Mindell and Honeycutt
1990) in synonymous changes is suggested by the sim-
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019
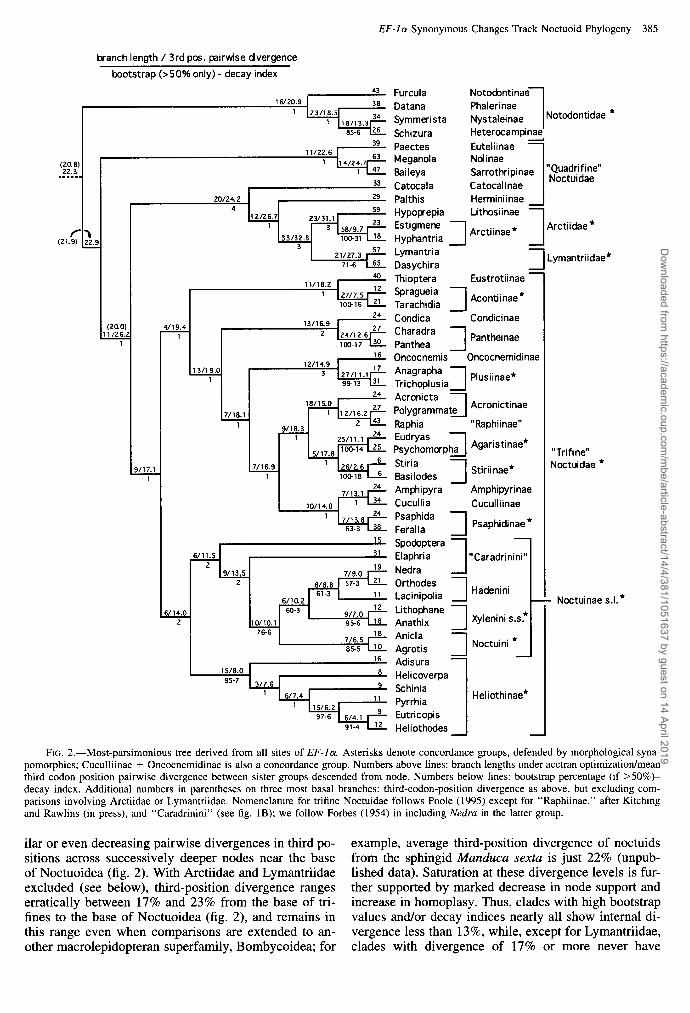
EF-la Synonymous Changes Track Noctuoid Phylogeny 385
branch length / 3rd pos. pairwise dvergence
bootstrap (>50% only) - decay index
1
v17.1 1
I/19.4 1 ,3,,;.oc$!
24
7/1&l 1 9/18.3
1 2Wll.l 24
S/17.8 100-14 (25
1 6 7/16.9
1 24
1on4.0 1 24
Furcula Notocbntinae Datana Phalerinae Symmerista Nystaleinae 1 Notodontidae *
Schizura Heterocampinae
Paectes Euteliinae - Meganola Nolinae Baileya Catocala
Sarrothripinae ‘~~~$itdin$” Catocalinae
Palthis Herminiinae 1 Hypoprepia Lithosiinae 1 Estigmene Hyphantria 7
Arctiinae * Arctiidae *
Lymantria Dasychira 7
Lymantriidae*
Thicptera Eustrotiinae - Spraguei a Tarachidia 7
Acontiinae l
Condica Condicinae Charadra Panthea 7
Pantheinae
Oncocnemis Oncocnemidinae Anagrapha Trichoplusia 7
Plusiinae*
Raphia “Raphiinae” Eudryas Psychommpha Agaristinae*
7 Stiria Basilodes 7
Stiriinae*
Amphipyra Amphipyrinae Cucullia Cuculliinae Psaphida Feralia 7
Psaphidinae *
Spodoptera Elaphria 1 “Caradrinini” Nedra Orthodes Lacinipdia 7
Hadenini
Lithophane Anathix 7
Xylenini s.s.”
Anicla Agrotis 7
Noctuini l -
Adisura Helicoverpa 1 Schinia Pyrrhia Eutricopis Heliothodes
Heliothinae*
“Trifine” Noctuidae l
L Noctuinae s.l. *
FIG. 2.-Most-parsimonious tree derived from all sites of EF-la. Asterisks denote concordance groups, defended by morphological syna- pomorphies; Cuculliinae + Oncocnemidinae is also a concordance group. Numbers above lines: branch lengths under acctran optimization/mean third codon position pairwise divergence between sister groups descended from node. Numbers below lines: bootstrap percentage (if >50%)- decay index. Additional numbers in parentheses on three most basal branches: third-codon-position divergence as above, but excluding com- parisons involving Arctiidae or Lymantriidae. Nomenclature for trifine Noctuidae follows Poole (1995) except for “Raphiinae,” after Kitching and Rawlins (in press), and “Caradrinini” (see fig. 1B); we follow Forbes (1954) in including Nedru in the latter group.
ilar or even decreasing pairwise divergences in third po- example, average third-position divergence of noctuids sitions across successively deeper nodes near the base from the sphingid A4unduca sexta is just 22% (unpub- of Noctuoidea (fig. 2). With Arctiidae and Lymantriidae lished data). Saturation at these divergence levels is fur- excluded (see below), third-position divergence ranges ther supported by marked decrease in node support and erratically between 17% and 23% from the base of tri- increase in homoplasy. Thus, clades with high bootstrap fines to the base of Noctuoidea (fig. 2), and remains in values and/or decay indices nearly all show internal di- this range even when comparisons are extended to an- vergence less than 13%, while, except for Lymantriidae, other macrolepidopteran superfamily, Bombycoidea; for clades with divergence of 17% or more never have
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019
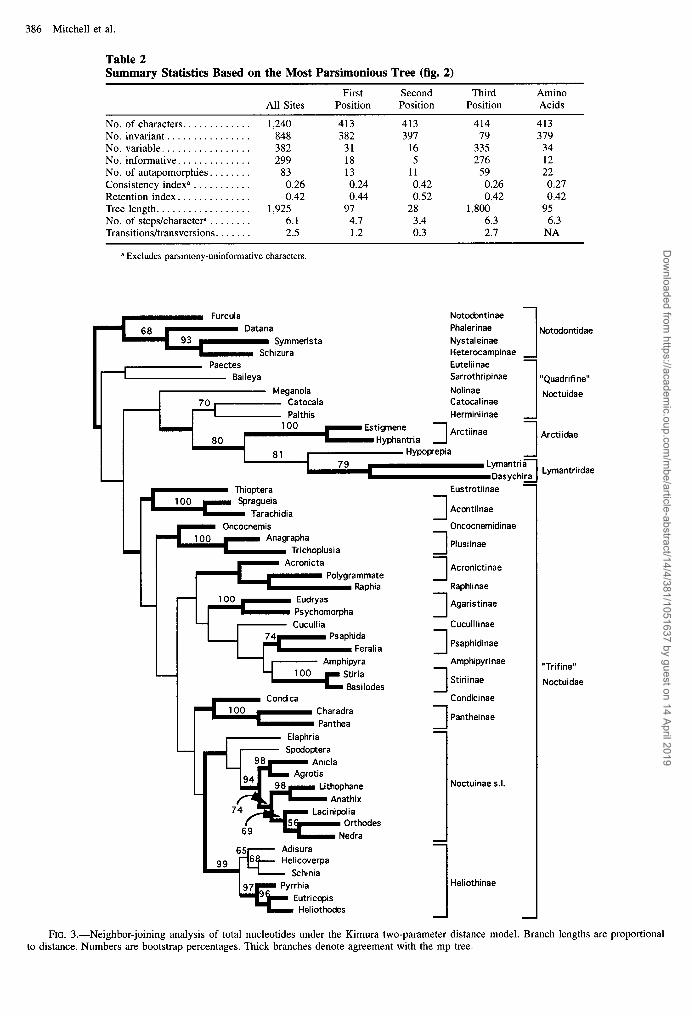
386 Mitchell et al.
Table 2 Summary Statistics Based on the Most Parsimonious Tree (fig. 2)
First Second Third Amino All Sites Position Position Position Acids
No. of characters. ............ 1,240 413 413 414 413 No. invariant. ............... 848 382 397 79 379 No. variable. ................ 382 31 16 335 34 No. informative. ............. 299 18 5 276 12 No. of autapomorphies ........ 83 13 11 59 22 Consistency indexa ........... 0.26 0.24 0.42 0.26 0.27 Retention index. ............. 0.42 0.44 0.52 0.42 0.42 Tree length. ................. 1,925 97 28 1,800 95 No. of steps/charactep ........ 6.1 4.7 3.4 6.3 6.3 Transitions/transversions ....... 2.5 1.2 0.3 2.7 NA
a Excludes parsimony-uninformative characters.
Notodontinae Phalerinae Nystaleinae Heterocampinae Euteliinae Sarrothripinae
Nolinae Catocalinae Herminiinae
II ” 80
Eustrotiinae
1 Acontiinae
Oncocnemidinae
1 Plusiinae
-
1 , loo 7 ~~~~~~orpha ] Agaristinae
- Cucullia
I- Condica
Cuculliinae
1 Psaphidinae
Amphipyrinae
1 Stiriinae
Condicinae
1 Pantheinae
Noctuinae s.1.
Heliothinae
lotodontidae
‘Quadrifine”
Noctuidae
Arctiidae
Lymantriidae
“Trifine”
Noctuidae
FIG. 3.-Neighbor-joining analysis of total nucleotides under the Kimura two-parameter distance model. Branch lengths are proportional to distance. Numbers are bootstrap percentages. Thick branches denote agreement with the mp tree.
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019
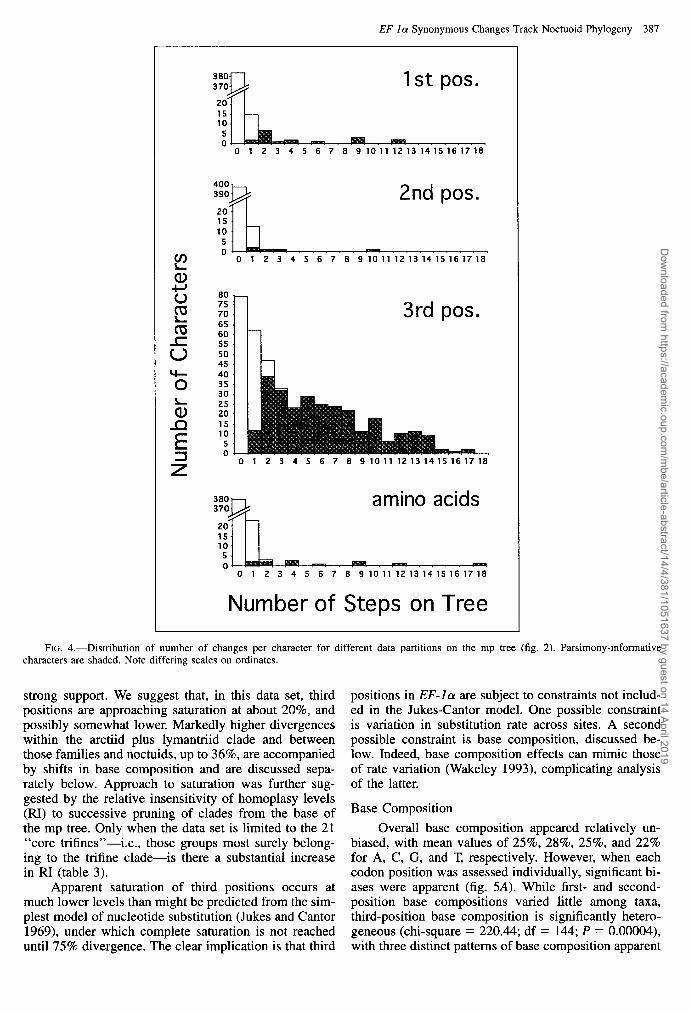
EF-la Synonymous Changes Track Noctuoid Phylogeny 387
1st pos.
0 1 2 3 4 5 6 7 8 9101112131415161718
2nd pos.
o- “““. -. . . .‘. . ( 0 1 2 3 4 5 6 7 8 9101112131415161718
80 I---, 75 70 65
II-l
3rd pos. 60 55
2 - 0 1 2 3 4 5 6 7 8 9 101112131415161718
380 370 /(
P amino acids
20 15 10 m
0 1 2 3 4 5 6 7 8 9101112131415161718
Number of Steps on Tree
FIG. 4.-Distribution of number of changes per character for different data partitions on the mp tree (fig. 2). Parsimony-informative characters are shaded. Note differing scales on ordinates.
strong support. We suggest that, in this data set, third positions are approaching saturation at about 20%, and possibly somewhat lower. Markedly higher divergences within the arctiid plus lymantriid clade and between those families and noctuids, up to 36%, are accompanied by shifts in base composition and are discussed sepa- rately below. Approach to saturation was further sug- gested by the relative insensitivity of homoplasy levels (RI) to successive pruning of clades from the base of the mp tree. Only when the data set is limited to the 21 “core trifines”-i.e., those groups most surely belong- ing to the trifine clade-is there a substantial increase in RI (table 3).
Apparent saturation of third positions occurs at much lower levels than might be predicted from the sim- plest model of nucleotide substitution (Jukes and Cantor 1969), under which complete saturation is not reached until 75% divergence. The clear implication is that third
positions in EF-Ia are subject to constraints not includ- ed in the Jukes-Cantor model. One possible constraint is variation in substitution rate across sites. A second possible constraint is base composition, discussed be- low. Indeed, base composition effects can mimic those of rate variation (Wakeley 1993), complicating analysis of the latter.
Base Composition
Overall base composition appeared relatively un- biased, with mean values of 25%, 28%, 25%, and 22% for A, C, G, and T, respectively. However, when each codon position was assessed individually, significant bi- ases were apparent (fig. 5A). While first- and second- position base compositions varied little among taxa, third-position base composition is significantly hetero- geneous (chi-square = 220.44; df = 144; P = 0.00004), with three distinct patterns of base composition apparent
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019

388 Mitchell et al.
Table 3 Effect on Retention Index of Pruning Clades from Base of Most Parsimonious Tree (fig. 2)
TAXA INCLUDED
Alltaxa ......................... All except Notodontidae ........... Noctuidae ....................... Trifines ......................... “Core trifines”a .................. Noctuinae s.1. and Heliothinae ...... Noctuinae s.1. .................... Heliothinae ......................
RETENTION INDEX
No. OF First Second Third All TAXA LENGTH Position Position Position Sites
49 1,925 0.44 0.52 0.42 0.42 45 1,727 0.48 0.55 0.43 0.43 40 1,364 0.45 0.65 0.44 0.44 35 1,082 0.49 0.69 0.47 0.47 21 536 0.58 0.80 0.57 0.57 15 327 0.68 1.00 0.60 0.60 9 211 0.56 1.00 0.43 0.47 6 95 1.00 1.00 0.63 0.67
a “Core Wines” denotes groups most certainly placed in the Wine clade, namely, the “Wine” clade in figure 2 minus Eustrotiinae, Acontiinae, Plusiinae, Acronictinae, Pantheinae, and Raphiinae.
(fig. 5B). However, this composition heterogeneity ap- pears to have little effect on our phylogeny estimate. The nj tree based on third-position base proportions alone (not shown) had only two nodes in common with the mp tree (those defining Acontiinae and Stiriinae). Moreover, the nj tree (using third positions only) based on the LogDet distance (Lockhart et al. 1994), which corrects for unequal base composition, showed only mi- nor differences from that based on the Kimura two-pa- rameter distance, and the positions of taxa with distinct base compositions were unchanged (not shown).
We hypothesize that a shift in base composition indicates change or relaxation of some (unknown) con- straint(s) on sequence evolution, causing or allowing the affected lineage to undergo unusual sequence diver- gence as well. If change in these constraints is rare or slow, there should commonly be saturation in local regions of the phylogeny, at divergence levels much lower than would comparisons. The
be inferred from broad taxonomic nature of constraints on base com-
position is unclear (Perna and Kocher 1995), and their relationship to sequence divergence is probably com- plex, as illustrated by our data for the arctiid Estigmene acrea. This species shows the elevated divergence char- acteristic of the arctiid plus lymantriid clade, but lacks the unusual base proportions.
Taxon Sampling
One approach to extending the range of utility for EF-la! might be increased taxon sampling density (Hillis 1996, but see Kim 1996). The decreased tree stability observed at deeper levels could, in part, be a reflection of the parallel decrease in sampling density. For trifine Noctuidae apart from Heliothinae, for example, we took at most two taxa each from a set of widely divergent tribes and subfamilies. To sample the entire Noctuoidea at the same proportional representation with which He- liothinae were sampled by Cho et al. (1995) would re- quire up to 3,000 species, but even just doubling the current sample might appreciably increase tree stability.
FIG. 5.-A, Mean nucleotide base composition for all taxa, by codon position. Bar brackets = ranges. B, Mean third-codon-position base composition, for each of three composition-homogeneous groups. Bar brackets = ranges.
1 st pos. All taxa
A C G T A C G T A C G T
2nd pos. All taxa
3rd pos.
Lymantriidae and Arctiidae except Estigmene
A C G T
50
40
3rd pos.
Schizura
30
20
10
r-i Lkh
“ACGT
50 3rd pos.
All others 40
30
20
10
0 h A C G T
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019

EF-la Synonymous Changes Track Noctuoid Phylogeny 389
Weighted Parsimony
As phylogenetic signal in the gene weakens for deeper nodes, we investigated whether preferential weighting of the (theoretically) more conserved char- acters could amplify the signal. We applied a wide range of differential weights to data partitions, including downweighting of sites undergoing only synonymous change, but found no improvement in support for deeper nodes. This is not surprising, given that essentially all of the phylogenetic signal lies in synonymous change; analysis of “synonymous sites” alone gave a single tree which differed from the mp tree for all sites only in the placement of the Plusiinae.
Range of Phylogenetic Utility of EF-la Synonymous Substitutions
While it is difficult to assign absolute dates to noc- tuoid divergences because of the sparse fossil record for lepidopteran families in comparison to other insects, there is an arctiid known from the Eocene (Lutetian, 42.1-50 MYA; Labandeira 1994). Thus noctuoids arose in the early Tertiary at latest, and the apparently derived position of Arctiidae lends credence to suggestions that the superfamily may have arisen even earlier (Kitching and Rawlins 1997). This suggests that the range of util- ity of synonymous substitutions in EF-Ia may extend over much of the Tertiary, at least for Lepidoptera. EF- Icu will clearly be highly useful for further study within subfamilies of Noctuoidea. With increased taxon sam- pling, and in combination with other genes, it should contribute to understanding of higher-level noctuoid re- lationships as well. EF-lcu is also proving useful within other groups of insects, including the ant tribe Attini (Hymenoptera: Formicidae; T. Schultz, personal com- munication) and genera, tribes, and subfamilies of hal- ictid bees (Hymenoptera: Halictidae; B. Danforth, per- sonal communication).
We thank T. Friedlander, Q. Fang, B. Wiegmann, I. Kitching, and J. Miller for helpful discussion, and D. Swofford for providing a test version of PAUP*4. For help in obtaining essential specimens we are indebted to generous colleagues, including R. Denno, E. Knudson, J. Lafontaine, D. Miller, R. Robertson, and N. Spencer. We are grateful for financial support from the U.S. De- partment of Agriculture-NRICGP, the National Science Foundation, the Maryland Agricultural Experiment Sta- tion, the Center for Agricultural Biotechnology, Univer- sity of Maryland Biotechnology Institute, and the Uni- versity of Maryland at College Park Graduate School.
LITERATURE CITED
AVISE, J. C. 1994. Molecular markers, natural history and evo- lution. Chapman and Hall, New York.
BREMER, K. 1988. The limits of amino-acid sequence data in angiosperm phylogeny reconstruction. Evolution 42:795- 803.
Phylogeny of Noctuoidea
Our data support the tentative findings of Weller et al. (1994), based on an analysis of the mitochondrial ND1 gene and nuclear 28s rDNA, that the Noctuidae are paraphyletic, with a subset of “quadrifine” subfam- ilies, including Catocalinae, being more closely related to Arctiidae and Lymantriidae. Analysis of dopa decar- boxyhse (DDC) sequences in a similar set of taxa also gives this result (Q. Fang, personal communication). Ev- idence is found for Kitching and Rawlins’ postulate of close relationship between Arctiidae and Lymantriidae, but not for their placements of Nolinae as sister group of Arctiidae and of Pantheinae as sister group to Ly- mantriidae.
CHO, S., A. MITCHELL, J. C. REGIER, C. MITTER, R. W. POOLE, T. I? FRIEDLANDER, and S. ZHAO. 1995. A highly conserved nuclear gene for low-level phylogenetics: Elongation Fuc- tor-I a recovers morphology-based tree for heliothine moths. Mol. Biol. Evol. 12:650-656.
DONOGHUE, M. J., R. G. OLMSTEAD, J. E SMITH, and J. D. PALMER. 1992. Phylogenetic relationships of Dipsacales based on rbcL sequences. Ann. MO. Bot. Gard. 79:333-345.
FANG, Q., S. CHO, J. C. REGIER, C. MITTER, M. MATTHEWS, R. W. POOLE, T I? FRIEDLANDER, and S. ZHAO. 1997. A new nuclear gene for insect phylogenetics: dopa decarbox- yluse is informative of relationships within Heliothinae (Lepidoptera: Noctuidae). Syst. Biol. (in press).
FORBES, W. T M. 1954. Lepidoptera of New York and neigh- boring states. Part 3. Noctuidae. Mem. Cornell Univ. Agric. Exp. Sta. 329: l-433.
Within Noctuidae, our results support, albeit weak- ly, a broadly defined “trifine” clade that includes taxa whose alliance with trifines has been uncertain, such as Pantheinae, Acontiinae s.s., Acronictinae, and the aber- rant genus Ruphiu. Within trifines, our data support a broad version of the “true cutworm” clade, the redefi- ned Noctuinae s.1. of Poole (1995). “Caradrinini” is of
FRANCLEMONT, J. G., and E. L. TODD. 1983. Noctuidae. Pp. 120-159 in R. W. HODGES, ed. Check list of the Lepidop- tera of America north of Mexico. The Wedge Entomologi- cal Research Foundation, Washington, D.C.
FRIEDLANDER, T P., J. C. REGIER, and C. MITTER. 1992. Nu- clear gene sequences for higher-level phylogenetic analysis: 14 promising candidates. Syst. Biol. 41:483-489.
-. 1994. Phylogenetic information content of five nuclear gene sequences in animals: initial assessment of character sets from concordance and divergence studies. Syst. Biol. 43:5 1 l-525.
FRIEDLANDER, T F?, J. C. REGIER, C. MITTER, and D. L. WAG- NER. 1996. A nuclear gene for higher level phylogenetics: phosphoenolpyruvate carboxykinase tracks Mesozoic-age divergences within Lepidoptera (Insecta). Mol. Biol. Evol. 13:594-604.
uncertain composition, and is only tenuously attached GUPTA, R. S. 1995. Phylogenetic analysis of the 90kD heat here, but support for the remainder of this assemblage shock family of protein sequences and an examination of
is reasonably strong. Our data suggest, although only weakly, that the sister group to the Noctuinae s.1. is the Heliothinae. This hypothesis, if confirmed bv further study, would unite the two most important pest-contain- ing noctuoid subfamilies, both rich in species with po- lyphagous, herb-feeding larvae. In contrast, the remain- ing trifines, with some clear exceptions, such as Plusi- inae, are predominantly oligophagous.
Acknowledgments
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019

390 Mitchell et al.
the relationship among animals, plants, and fungi species. Mol. Biol. Evol. 12: 1063-1073.
HILLIS, D. M. 1996. Inferring complex phylogenies. Nature 383:130-131.
JUKES, T. H., and C. R. CANTOR. 1969. Evolution of protein molecules. Pp. 21-132 in H. N. MUNRO, ed. Mammalian protein metabolism. Academic Press, New York.
KAMIIE, K., H. TAIRA, H. OOURA, S. MATSUMOTO, S. EJIRI, and T. KATSUMATA. 1993. Nucleotide sequencing of the cDNA encoding silk gland elongation factor-l alpha. Nu- cleic Acids Res. 211742.
KIM, J. 1996. General inconsistency conditions for maximum parsimony: effects of branch lengths and increasing num- bers of taxa. Syst. Biol. 45:363-374.
KIMURA, M. 1980. A simple method for estimating evolution- ary rate of base substitution through comparative studies of nucleotide sequences. J. Mol. Evol. 16: 11 l-120.
KITCHING, I. J. 1984. An historical review of the higher clas- sification of the Noctuidae (Lepidoptera). Bull. Br. Mus. Nat. Hist. (Entomol.) 49: 153-234.
KITCHING, I. J., and J. E. RAWLINS. 1997. Noctuoidea. Chapter 19 in N. F? KRISTENSEN, ed. Handbook of zoology. Lepi- doptera. Vol 1: Systematics and evolution. W. de Gruyter, Berlin (in press).
LABANDEIRA, C. C. 1994. A compendium of fossil insect fam- ilies. Milwaukee Public Museum Contributions in Biology and Geology, no. 88. Milwaukee Public Museum, Milwau- kee.
LAFONTAINE, J. D. 1993. Cutworm systematics: confusions and solutions. Mem. Entomol. Sot. Can. 165:189-196.
LOCKHART, I? J., M. A. STEEL, M. D. HENDY, and D. PENNY. 1994. Recovering evolutionary trees under a more realistic model of sequence evolution. Mol. Biol. Evol. 11:605-612.
MADDISON, W. l?, and D. R. MADDISON. 1992. MacClade: anal- ysis of phylogeny and character evolution. Version 3. Sin- auer, Sunderland, Mass.
MILLER, J. S. 1991. Cladistics and classification of the Noto- dontidae (Lepidoptera: Noctuoidea) based on larval and adult morphology. Bull. Am. Mus. Nat. Hist. 204:1-230.
MINDELL, D. I?, and R. L. HONEYCUT~. 1990. Ribosomal RNA in vertebrates: evolution and phylogenetic applications. Annu. Rev. Ecol. Syst. 21541-566.
MIYAMOTO, M. M., and J. CRACRAFT. 1991. Phylogenetic in- ference, DNA sequence analysis, and the future of molec- ular systematics. Pp. 3-17 in M. M. MIYAMOTO and J. CRA- CRAFT, eds. Phylogenetic analysis of DNA sequences. Ox- ford University Press, New York.
ORT~, G., and A. MEYER. 1996. Molecular evolution of epen- dymin and the phylogenetic resolution of early divergences among euteleost fishes. Mol. Biol. Evol. 13:556-574.
PALUMBI, S., and C. S. BAKER. 1994. Contrasting population structure from nuclear intron sequences and mtDNA of humpback whales. Mol. Biol. Evol. 11:426-435.
PERNA, N. T., and T D. KOCHER. 1995. Patterns of nucleotide composition at fourfold degenerate sites of animal mito- chondrial genomes. J. Mol. Evol. 41:353-358.
POOLE, R. W. 1995. Noctuoidea. Noctuidae (part). Cuculliinae, Stiriinae, Psaphidinae (part). Moths of America North of Mexico, Fascicle 26.1. The Wedge Entomological Research Foundation, Washington, D.C.
ROHLF, E J. 1990. NTSYS-pc. Numerical taxonomy and mul- tivariate analysis system. Version 1.60. Exeter Software, New York.
SAITOU, N., and M. NEI. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406-425.
SLADE, R. W., C. MORITZ, and A. HEIDEMAN. 1994. Multiple nuclear-gene phylogenies: application to pinnipeds and comparison with a mitochondrial DNA gene phylogeny. Mol. Biol. Evol. 11:341-356.
SMITH, S. W., R. OVERBEEK, C. R. WOESE, W. GILBERT, and I? M. GILLEVET. 1994. The genetic data environment and expandable GUI for multiple sequence analysis. Comput. Appl. Biosci. 10:671-675.
SPEIDEL, W., and C. M. NAUMANN. 1995. Further morpholog- ical characters for a phylogenetic classification of the Noc- tuidae (Lepidoptera). Beitr. Entomol. 45: 119-135.
STADEN, R. 1992. Staden package. MRC Laboratory of Mo- lecular Biology, Cambridge, England.
TAMURA, K., and M. NEI. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochon- drial DNA in humans and chimpanzees. Mol. Biol. Evol. 10:512-526.
WAKELEY, J. 1993. Substitution rate variation among sites in hypervariable region 1 of human mitochondrial DNA. J. Mol. Evol. 37:6 13-623.
WATERS, E. R. 1995. An evaluation of the usefulness of the small heat shock genes for phylogenetic analysis in plants. Ann. MO. Bot. Gard. 82:278-295.
WELLER, S. J., D. I? PASHLEY, J. A. MARTIN, and J. L. CON- STABLE. 1994. Phylogeny of noctuoid moths and the utility of combining independent nuclear and mitochondrial genes. Syst. Biol. 43: 194-211.
RODNEY L. HONEYCUTT, reviewing editor
Accepted December 11, 1996
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/4/381/1051637 by guest on 14 April 2019