Embed Size (px)
Citation preview

This article was downloaded by: [University of Connecticut]On: 10 October 2014, At: 09:04Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number:1072954 Registered office: Mortimer House, 37-41 Mortimer Street,London W1T 3JH, UK
Journal of Plant NutritionPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/lpla20
Photosystem II and cellularmembrane stabilityevaluation in hexaploidwheat seedlings under saltstress conditionsVeli Cengiz Özalp a , Hüseyin Avni Öktem a ,Syed Mohammed Saqlan Naqvi b & Meral Yücel aa Department of Biology , Middle East TechnicalUniversity , 06531, Ankara, Turkeyb Tissue Culture Lab NARC , PO NIH, Islamabad,PakistanPublished online: 21 Nov 2008.
To cite this article: Veli Cengiz Özalp , Hüseyin Avni Öktem , Syed MohammedSaqlan Naqvi & Meral Yücel (2000) Photosystem II and cellular membrane stabilityevaluation in hexaploid wheat seedlings under salt stress conditions, Journal ofPlant Nutrition, 23:2, 275-283, DOI: 10.1080/01904160009382014
To link to this article: http://dx.doi.org/10.1080/01904160009382014
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of allthe information (the “Content”) contained in the publications on ourplatform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy,completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views ofthe authors, and are not the views of or endorsed by Taylor & Francis.The accuracy of the Content should not be relied upon and should be

independently verified with primary sources of information. Taylor andFrancis shall not be liable for any losses, actions, claims, proceedings,demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, inrelation to or arising out of the use of the Content.
This article may be used for research, teaching, and private studypurposes. Any substantial or systematic reproduction, redistribution,reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of accessand use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

JOURNAL OF PLANT NUTRITION, 23(2), 275-283 (2000)
Photosystem II and Cellular MembraneStability Evaluation in Hexaploid WheatSeedlings Under Salt Stress Conditions
Veli Cengiz Özalp,a Hüseyin Avni Öktem,a Syed MohammedSaqlan Naqvi,b and Meral Yücela
aMiddle East Technical University, Department of Biology, 06531 Ankara,TurkeybTissue Culture Lab NARC, PO NIH, Islamabad, Pakistan
ABSTRACT
Salinity limits crop production in large areas of the world. The application ofin vitro Photosystem II (PS-II) activity measurements to screen hexaploidwheat (Triticum aestivum, L.) genotypes for NaCl tolerance has beeninvestigated by comparing their responses under stress and control (no addedNaCl) conditions. One of the four cultivars used in the study was 'Kharchia'known for its high salt tolerance. Wheat seedlings were grown hydroponicallyin environmental chambers and treated with a range of NaCl concentrations(0.034 M, 0.17 M, 0.68 M, or 3.42 M) over a 1, 3, and 5-day period. The salttreatments were started in the appropriate time so that they were all ten-day-old during harvest. Cellular membrane stability (CMS) as measured by aconductivity method and PS-II activity values were affected adversely byNaCl concentration and duration of treatment. Both methods clearly distinguish
275
Copyright © 2000 by Marcel Dekker, Inc. www.dekker.com
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

276 ÖZALP ET AL.
between salt-sensitive and salt-tolerant genotypes. Statistical analysis showedthat PS-II activity and CMS measurements are well correlated (r=0.7589)suggesting that PS-II activity would be used as an additional screening methodbesides CMS to evaluate salt tolerance of wheat.
INTRODUCTION
Salt stress is one of the major environmental stresses affecting agriculturalproduction in arid regions of the world. Salinity that results from increasing saltconcentrations in soil and irrigation water has been unavoidable problem in thearid lands (Epstein, 1980). Wheat genotypes differ in response to NaCl stress(Ashraf, 1988). Therefore, physiological screening processes are needed to evaluatethe salt tolerance of plants to be used in breeding program (Blum, 1985).
Cellular membrane stability evaluations are the most commonly employedphysiological screening methodology used for a number of plant species, includingwheat varieties, for environmental stresses (Martineau, 1979). Among otherscreening methods, dry mass measurements are simple but a labor intensivescreening methodology. Chlorophyll a fluorescence has been used to assessphotosynthetic reactions of a variety of stresses on crop plants (Lichtenthaler,1988). The potential of utilizing quenching kinetics of chlorophyll a fluorescencefor salt tolerance screening was investigated in various plants including hexaploidwheat varieties (Larcher, 1990; Krishbaraj, 1993). Recently Photosystem II (PS-II)activity measurements on isolated thylakoid membranes from heat stress treatedwheat revealed promising results that such measurements can be also used toevaluate stress tolerance of a plant (Yücel et. al., 1992). Compared to fluorescencestudies, the PS-II method is a more through assessment of PS-II reaction centersince it gives the rate of the whole PS-II complex.
The main objective of this work was to evaluate the applicability of PS-II activitymeasurements as a screening method for salt tolerance of wheat cultivars. For thatpurpose, correlation between cellular membrane stability of root tissue and PS-IIactivity of hexaploid wheat seedlings under salt stress conditions was investigated.
MATERIALS AND METHODS
Plant Material and Growth of Seedlings
Indian origin Triticum aestivum 'Kharchia' has been used as standard since itwas confirmed previously as salt tolerant cultivar (Diaz et al., 1995). The otherthree genotypes are selected randomly from the most cultivated genotypes ofTurkish agriculture ('Bezostaya, 'Gerek', and 'Tosun').
Four cultivars of hexaploid wheat were grown hydroponically from seeds at23°C±3°C in a controlled-environmental chamber in half-strength Hoagland solution(HoaglandandArnon, 1938). Seedlings were illuminated with4 fluorescent lamps(2,300 lm each) at 50 cm above the containers in a photoperiod of 16 hours light and
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

HEXAPLOID WHEAT SEEDLINGS UNDER SALT STRESS 277
8 hours dark. The humidity inside the chamber was kept at 65% with humiditycontroller with a humidifier.
Salt Treatments
Seedlings were grown in half-strength Hoagland solutions. Seedlings were salt-shocked by changing the solution in the container with the treatment solutionswhich were half-strength Hoagland solutions with a final concentration of NaCl at0.02% (0.034 M), 1% (0.17 M), 4% (0.68 M) or20% (3.42 M). The controlplants wereleft in the half-strength Hoagland solution for 10 days. The timing of salt treatmentwas adjusted so that seedlings were 10 days old during harvest. The durations ofsalt treatment were 1,3 or 5 days. The leaf and root samples were used immediatelyfor PS-II activity and conductivity experiments. For each salt application andduration, four measurements from four different containers were taken.
Measurement of Cellular Membrane Stability
Cellular Membrane Stability of root tissue was measured with the ion leakagemethod of Marsh et al. (1985). Root tissue collected from four seedlings grown inthe same container were washed with distilled water and placed in plastic tubeswith 15 mL of distilled water with a known conductivity value. This value wassubtracted from the further readings. Tubes were incubated in a temperaturecontrolled chamber at 10°C overnight and measured for conductivity (Cl). Thisprocedure was repeated for control and salt-stressed plants in the same way.Subsequent control and treatment tubes were frozen at -70°C for 10 hours to allowcomplete ion release. After the tubes were kept overnight at room temperature,second conductivity (C2) values was measured. The cell membrane stability ofroot tissues was calculated, and CMS was expressed as percent of conductivity ofsalt-stressed plant compared to the control plant according to the following formula:
CMS=[l-(Cl/C2)/(Ccl/Cc2)]xlOO
where:
Cl is the first conductivity measurement of treated plants.C2 is the second conductivity measurement of treated plants.Ccl is the first conductivity measurement of control plants.Cc2 is the first conductivity measurement of control plants.
PS-n Activity
Thylakoid membrane fragments from treated and control seedling were isolatedand assayed for PS-II activities as described in Chetti et al. (1988). Five gm of leaftissue were homogenized in grinding medium (0.33 Msorbitol, 50 mM NaCl, 5 mMMgClj, 1 mM ethylenediaminetetraacetic acid (EDTA), 0.1% bovine serum albumin(BSA), and 50 mM N-(2-hydroxyethyl)piperazine-N'-(2-ethanesulfonic acid)
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

278 ÖZALP ET AL.
(HEPES) KOHbufferpH=7.0) for 10 seconds. The debris was removed by filtratingthe homogenate through cheese-cloth. The chloroplast-rich pellet was collectedby centrifuge at 6,000 g and solubilized in suspension medium (0.1. Msorbitol, 50mM MgClj, 50 mM HEPES KOH buffer pH=7.0). The thylakoid membrane fragmentswere collected by centrifugation at 12,000 g for 10 min. and resuspended in 100 uLof suspension medium. The biochemical assay used was based on themeasurements of electron transfer efficiency from water to dichloropheno-lindophenol (DCPIP) over PS-II complex spectrophotometrically. Thylakoidmembrane fragments equivalent to 50 ng chlorophyll as determined according toArnon (1949) were added in 2 mL of assay solution (40 mM Tricine KOH pH=8.1,5mM MgCl2,2 mM KjHPO,, 10 mM KC1,30 uM DCPIP) in dark and placed in aspectrophotometer cell with a cooling jacket maintained at 25 °C. The reaction wasstarted by turning on a cool light source. The decreases in absorbance at 595 nmwere recorded for 60 seconds. The initial phase of absorbance versus time curvewas used as the indication of rate of PS-II complex. The PS-II values were evaluatedas the percentage of control values.
Data Analysis
All experiments for CMS and PS-II measurements were repeated four times.Treatment responses were determined by analysis of variance. Differences amongmeans were tested by least significant differences (LSD) (P<0.05). Spearman rankcorrelation coefficient was calculated for reductions in photosynthetic activity vs.retention of cellular membrane stability with SPSS for Windows.
RESULTS
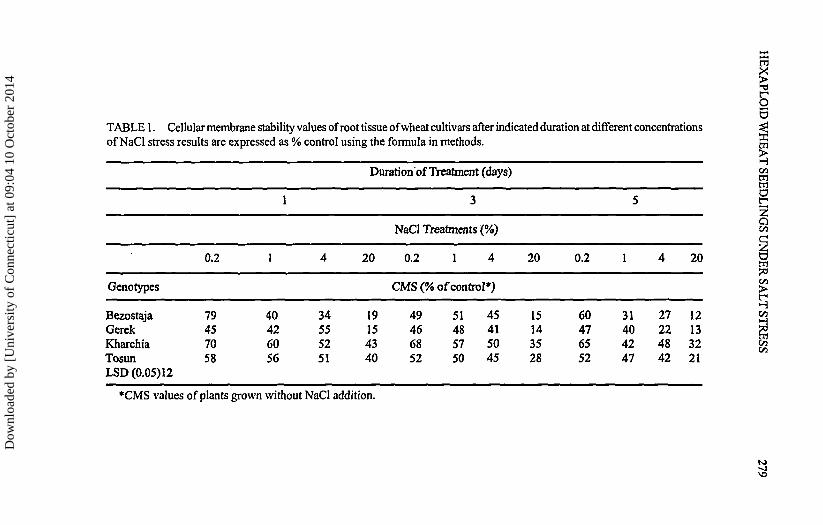
Cellular Membrane Stability
The cellular membrane stability was measured with the conductivity methodand was calculated as the proportion of conductivity retained after stress as thepercentage of control root tissue (Table 1). The overall analysis of data show thatthe four genotypes can be separated into two group based on the CMS values atthe indicated salt treatments. 'Bezostaya' and'Gerek' can be labeled as salt sensitivecompared to 'Tosun' and 'Kharchia' as the salt tolerant genotypes. For eachgenotype, the CMS values increased with the increasing percentage of NaCl andwith the duration of treatment from 1 day to 5 days. The 50% injury measured byconductivity method was used as an index of salt stress for comparative purposes.The extreme salt concentration (20% NaCl) treatment of seedlings caused morethan 50% injury for all the genotypes at all the duration of stresses applied. Similarlythe high salt concentration (4% NaCl) caused more than 50% injury at most of thetreatments but with a lower degree of injury than the 20% treatment. In 0.2 and 1 %treatments, 'Gerek' and 'Bezostaya' show near or more than 50% injury for most ofthe treatments whereas 'Tosun' and 'Kharchia' showed an injury value below50%.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4
Xa
TABLE 1. Cellular membrane stability values of root tissue of wheat cultivars after indicated duration at different concentrationsof NaCl stress results are expressed as % control using the formula in methods.
Genotypes
BezostajaGerekKharchiaTosunLSD (0.05)12
0,2
79457058
1
1
40426056
4
34555251
Duration of Treatment (days)
20
19154340
3
NaCl Treatments (%)
0.2 1 4
CMS (% of control»)
49466852
51485750
45415045
20
15143528
0.2
60476552
5
1
31404247
4
27224842
20
12133221
GO
3Q
om
on
oo
Ionin
"CMS values of plants grown without NaCl addition.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4
ooo
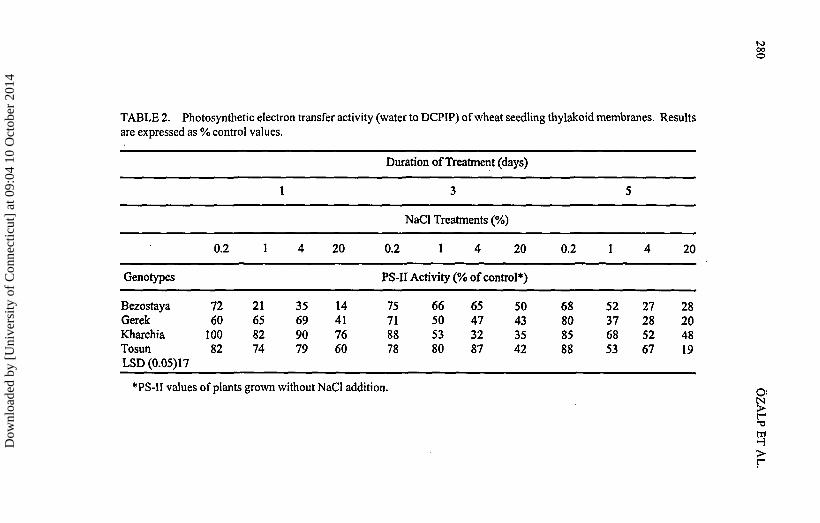
TABLE 2. Photosynthetic electron transfer activity (water to DCPIP) of wheat seedling thylakoid membranes. Resultsare expressed as % control values.
Genotypes
0.2
1
1 4 20
Duration of Treatment (days)
3
NaCl Treatments (%)
0.2 1 4 20
PS-II Activity (% of control*)
0.2
5
1 4 20
BezostayaGerekKharchiaTosunLSD (0.05)17
7260
10082
21658274
35699079
14417660
75718878
66505380
65473287
50433542
68808588
52376853
27285267
28204819
*PS-II values of plants grown without NaCl addition.
>t - 1
• •a
w
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

HEXAPLOID WHEAT SEEDLINGS UNDER SALT STRESS 281
Photosynthetic Rates
The photosynthetic rates were expressed as the percentage of DCPBP reductionrates compared to control plants (Table 2). The rates changed significantly betweendifferent NaCl treatments and between the durations for all the genotypes. Foreach genotype, the PS-II values decreased with the increasing percentage of NaCland with the duration of treatment from 1 day to 5 days. The extreme saltconcentration 20% NaCl treatment of seednngs caused more than 50% PS-II activitydecrease for all the genotypes at all the duration of stresses applied, except for the1 day treatment for 'Kharchia' and 'Tosun'. Similarly the high salt concentration of4% caused more than 50% injury at most of the treatments but with a lower decreasecompared to the 20% treatment. In 0.2 and 1% treatments, all genotypes kept morethan 50% of their PS-II activities.
Photosynthetic rates of seedlings differed among four genotypes. The overallanalysis of all data shows that the photosynthetic activities of 'Kharchia' and'Tosun' are tolerant to salt stress whereas 'Bezostaya' and 'Gerek' are sensitiveunder salt stress.
DISCUSSION
Cellular Membrane Stability evaluation has been a commonly used physiologicalscreening method for environmental stresses including salt stress (Blum, 1985).On the other hand, photosynthesis, particularly PS-II activity center was shown tobe the most sensitive process under environmental stresses (Harding et al., 1990;Yüceletal., 1992). Furthermore, PS-II activity has been thought to be representativeof the vegetative growth potential under stress conditions. Therefore, in a screeningprogram PS-II activity can be used to assess the tolerance of wheat varieties undersalt stress conditions.
In this study, we have shown a good correlation (r=7589) of high PS-II activityand high CMS. Although the photosynthetic responses of seedlings under saltstress did not totally correspond with the ranking of CMS, the LSD analysis groupsthe data into two statistically significant groups. In both tests 'Kharchia' and'Tosun' appeared to be tolerant whereas 'Bezostaya' and 'Gerek' were sensitive.It has been demonstrated that the high PS-II activity under salt stress indicates apotential for high yield productivity, and that the high CMS indicates the potentialfor long yield stability (Blum, 1985). Therefore, since the results of this researchshow a correlation between CMS and PS-II activity, these methods might be usedtogether in rapid screening programs to achieve a better assessment of the cultivarsunder study.
ACKNOWLEDGMENTS
We thank to Assoc. Prof. Dr. Fahri Atay from Transitional Zone Agricultural ResearchInstitute in Eskisehir and Prof. Dr. Ekrem Kûn from the University of Ankara for
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

282 ÖZALP ET AL.
providing the wheat seeds. This study was supported by Scientific and TechnicalResearch Council of Turkey (TÜBITAK) Project no. TBGAG 31 DPT.
REFERENCES
Arnon, D.I. 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Betavulgaris. Plant Physiol. 24:1-15.
Ashraf, M. and T. McNeilly. 1988. Variability in salt tolerance of nine spring wheatcultivars. J. Agron. Crop. Sci. 160:14-21.
Blum, A. 1985. Breeding Crop Varieties for Stress Environments. CRC Crit. R. Plant 2-199.
Chetti, M.B. and P.S. Nobel. 1988. Recovery of photosynthetic reactions after hightemperature treatments of a heat tolerant cactus. Photosyn. Res. 18:277-286.
Diaz, D.L., M. Carillo-Laguna, S. Rajaram, and A. Mujeeb-Kazi. 1995. Rapid in vitroscreening of some salt tolerant bread wheats. Cereal Res. Commun. 23 4):383-389.
Epstein, E. 1980. Responses of plants in saline environments, pp. 7-21. In: D.W. Rains,R.C. Valentine, and A. Hollaender (eds.), Genetic Engineering of Osmoregulation. PlenumPress, New York, NY.
Harding, S.A., J.A. Guikema, and G.M. Paulsen. 1990. Photosynthetic decline from hightemperature stress during maturation of wheat. I. Interaction with senescence processes.Plant Physiol. 92:648-653.
Hoagland, D.R. and D.I. Arnon. 1938. The water culture method for growing plantswithout soil. California Agric. Exp. Sta. Circ. 347.
Krishbaraj, S., B.T. Mawson, E.C. Yeung, and T.E. Thorpe. 1993. Utilization of inductionand quenching kinetics of chlorophyll a fluorescence for in vivo salinity screening studiesin wheat (Triticum aestivum vars. Kharchia-65 and Fielder). Can. J. Bot. 71:87-92.
Larcher, W., J. Wagner, and A. Thammathaworn. 1990. Effects of superimposed temperaturestress on in vivo chlorophyll fluorescence of Vigna unguiculata under saline stress. J.Plant Physiol. 1136:92-102.
Lichtenthaler, H.K. and V. Rinderle. 1988. The role of chlorophyll fluorescence in thedetection of stress conditions in plants. CRC Crit. Rev. Anal. Chem. 136:92-102.
Marsh, L.E., D.W. Davis, and P.H. Li. 1985. Selection and inheritance of heat tolerance inthe common bean by use of conductivity. J. Am. Soc. Hort. Sci. 110:680-683.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4

HEXAPLOID WHEAT SEEDLINGS UNDER SALT STRESS 283
Martineau, J.R. 1979. Temperature tolerance in soybeans. II. Evaluations of segregatingpopulations for membrane thermostability. Crop. Sci. 19:79-89.
Yücel, M., J. Burke, and H.T. Nguyen. 1992. Inhibition and recovery of Photosystem IIfollowing exposure of wheat to heat shock. Environ. Exp. Bot. 32(2): 125-135.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
9:04
10
Oct
ober
201
4





























