Embed Size (px)
Citation preview

2013
Chronobiology International, 2013; 30(9): 1089–1100! Informa Healthcare USA, Inc.ISSN: 0742-0528 print / 1525-6073 onlineDOI: 10.3109/07420528.2013.800090
Photoperiod affects the diurnal rhythm of hippocampal neuronalmorphology of siberian hamsters
Tomoko Ikeno, Zachary M. Weil, and Randy J. Nelson
Department of Neuroscience, The Ohio State University Wexner Medical Center, Columbus, Ohio, USA
Individuals of many species can regulate their physiology, morphology, and behavior in response to annual changesof day length (photoperiod). In mammals, the photoperiodic signal is mediated by a change in the duration ofmelatonin, leading to alterations in gene expressions, neuronal circuits, and hormonal secretion. The hippocampus isone of the most plastic structures in the adult brain and hippocampal neuronal morphology displays photoperiod-induced differences. Because the hippocampus is important for emotional and cognitive behaviors, photoperiod-driven remodeling of hippocampal neurons is implicated in seasonal differences of affect, including seasonal affectivedisorder (SAD) in humans. Because neuronal architecture is also affected by the day-night cycle in several brain areas,we hypothesized that hippocampal neuronal morphology would display a diurnal rhythm and that day length wouldinfluence that rhythm. In the present study, we examined diurnal and seasonal differences in hippocampal neuronalmorphology, as well as mRNA expression of the neurotrophic factors (i.e., brain-derived neurotrophic factor [Bdnf],tropomyosin receptor kinase B [trkB; a receptor for BDNF], and vascular endothelial growth factor [Vegf]) and a circadianclock gene, Bmal1, in the hippocampus of Siberian hamsters. Diurnal rhythms in total length of dendrites, the numberof primary dendrites, dendritic complexity, and distance of the furthest intersection from the cell body were observedonly in long-day animals; however, diurnal rhythms in the number of branch points and mean length of segmentswere observed only in short-day animals. Spine density of dendrites displayed diurnal rhythmicity with different peaktimes between the CA1 and DG subregions and between long and short days. These results indicate that photoperiodaffects daily morphological changes of hippocampal neurons and the daily rhythm of spine density, suggesting thepossibility that photoperiod-induced adjustments of hippocampal neuronal dynamics might underlie seasonaldifference of affective responses. Bmal1 mRNA showed a diurnal rhythm and different expression levels between longand short days were observed. However, there were no strong effects of day length on Bdnf, trkb, and Vegf geneexpression, suggesting that these genes are not involved in the photoperiodic effects on hippocampal neurons.
Keywords: Animal models, BDNF, circadian clock, day length, diurnal rhythm, seasonal affective disorder, seasonality,Siberian hamsters, trkb, VEGF
INTRODUCTION
Many organisms are able to respond to annual cycles of
day length (photoperiod) by physiological, morpho-
logical, and behavioral changes in anticipation of
approaching seasons (Goldman, 2001). In response to
short day lengths, many small rodents cease reproduct-
ive activities to avoid producing offspring during the
winter when temperatures and food availability are
challenging. The photoperiodic response is mediated by
a change in the duration of nocturnal hormone, mela-
tonin (Pevet, 1988). Melatonin secretion from the pineal
gland is regulated by a circadian clock, which is an
internal daily timekeeping system, and is directly sup-
pressed by light (Goldman, 2001).
In addition to the reproductive traits, seasonal
changes of brain morphology (such as brain volume
and neuronal architecture) have also been reported for
individuals of some vertebrate species. In songbirds, for
example, the volumes of brain regions that control song
production increase dramatically in anticipation of the
breeding season (Brenowitz et al., 1997; Smith et al.,
1997; Tramontin et al., 2001). These volumetric changes
are mediated by increases in neuronal size, number,
and spacing (reviewed in Tramontin & Brenowitz, 2000).
In mammals, seasonal differences have been observed
in various brain regions, especially in the hippocampus,
which is one of the most plastic sites in adult mamma-
lian brains (Breedlove & Jordan, 2001). Wild-captured
rodents of many species display seasonal variation in
Correspondence: Tomoko Ikeno, Department of Neuroscience, The Ohio State University, Columbus, OH 43210, USA. Tel. +1 614688 4674; Fax: +1 614 292 3464; E-mail: [email protected]
Submitted March 6, 2013, Returned for revision April 12, 2013, Accepted April 24, 2013
1089
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

hippocampal volume; exposure to short days in the
laboratory reduces whole-brain and hippocampal
volume (Burger et al., 2013; Galea & McEwen, 1999;
Perrot-Sinal et al., 1998; Pyter et al., 2005). In white-
footed mice, Peromyscus leucopus, and Siberian ham-
sters, Phodopus sungorus, short days change neuronal
architecture, including soma size, dendritic complexity,
and dendritic spine density in the hippocampus
(Pyter et al., 2005; Workman et al., 2011).
The hippocampus has been considered a key area
for emotion and cognition (Small et al., 2011). The
hippocampus plays an inhibitory role in the regulation
of the hypothalamic-pituitary-adrenal (HPA) axis activ-
ity, which is the major neuroendocrine system that
regulates reactions to stressors and regulates several
biological and psychological processes, including mood
and emotions (Jacobson & Sapolsky, 1991). Reduction in
hippocampal volume, atrophy of hippocampal neurons,
altered neuronal morphology, or loss of hippocampal
dendritic spines is often observed in depressed patients
(Cole et al., 2011; Neumeister et al., 2005; Stockmeier
et al., 2004) and in nonhuman animals displaying
depressive-like responses (Hajszan et al., 2009, 2010;
Magarinos & McEwen 1995; Watanabe et al., 1992).
It has been suggested that photoperiodic changes
in hippocampal structure possibly underlie seasonal
affective disorder (SAD), which is a specific mood
disorder characterized by depression induced by shor-
tened day lengths in winter (Workman & Nelson, 2011).
In addition to seasonal changes, many processes,
ranging from gene expression to behavior, show daily
rhythms that are regulated by the circadian clock (Ko &
Takahashi, 2006). Diurnal rhythmicity is observed in
mood status both in humans (Adan & Sanchez-Turet,
2001; Owens et al., 2000) and in animal models of
affective disorders (Kaya et al., 2011). In mammals, a
master clock is located in the suprachiasmatic nucleus
(SCN) of the hypothalamus, where circadian clock genes
drive autonomous feedback loops to generate circadian
signals (Ko & Takahashi, 2006). In addition to the SCN,
circadian clock genes are also expressed throughout the
brain, including the hippocampus (Jilg et al., 2010). In
some tissues or cells, neurotrophic factors or their
receptors are implicated in depression; for example,
brain-derived neurotrophic factor (BDNF), tropomyosin
receptor kinase B (TrkB) (a receptor for BDNF and also
called neurotrophic tyrosine kinase receptor type 2
[Ntrk2]), and vascular endothelial growth factor (VEGF)
show circadian or diurnal rhythms in mRNA or protein
expressions (Blanco et al., 2011; Dolci et al., 2003;
Koyanagi et al., 2003; Schaaf et al., 2000). Because these
genes are essential for neuronal development and
survival, synaptic transmission and plasticity, or neuro-
genesis (Fournier & Duman, 2012; Yu & Chen, 2011) and
because the hippocampus expresses these genes (Dolci
et al., 2003; Shaaf et al., 2000; Wang et al., 2005), we
hypothesized that hippocampal neuronal architecture
also shows diurnal differences, as well as seasonal
changes, and that these neurotrophic factors contribute
to hippocampal neuronal plasticity.
In the present study, we examined the diurnal
differences of hippocampal neuronal morphology as
well as mRNA expression of neurotrophic genes, Bdnf,
trkb, and Vegf, and the circadian clock gene, Bmal1, in
the hippocampus of Siberian hamsters exposed to long
or short days. Individuals of this species exhibit many
seasonal adaptations such as a loss of body mass and
the cessation of reproduction during the winter that can
be produced by short-day exposure in the laboratory
(reviewed in Scherbarth & Steinlechner, 2010). Because
individuals of this species also respond to short days
with a depressive-like phenotype, Siberian hamsters
may serve as a potential animal model for SAD
(Prendergast & Nelson, 2005; Pyter & Nelson, 2006;
Workman & Nelson, 2011; Workman et al., 2011). We
compared the diurnal patterns between long days and
short days in Siberian hamsters to determine whether
day length affects hippocampal rhythmicity.
MATERIALS AND METHODS
AnimalsSiberian hamsters (Phodopus sungorus) used in this
study were bred in our colony at The Ohio State
University. Male hamsters were weaned during the
light phase at 21–24 d of age and immediately placed
into either short-day conditions (SD: 8 h light:16 h dark;
N¼ 35) or maintained in long-day conditions (LD: 16 h
light:8 h dark; lights off at 15:00 h Eastern Standard Time
[EST] in all cases; N¼ 37) and at a constant temperature
of 21� 2 �C and relative humidity of 50� 10%. Hamsters
were individually housed in polypropylene cages
(30� 15� 14 cm) and had ad libitum access to food
(Harlan Teklad Rodent Diet 8640; Indianapolis, IN, USA)
and filtered tap water. Hamsters were housed in their
respective photoperiods for 4 wks prior to experiments.
All procedures were approved by the Ohio State
University Institutional Animal Care and Use
Committee and comply with guidelines established
by the National Instituted of Health published in
Guide for the Care and Use of Laboratory Animals
(Institute of Laboratory Animal Resources (U.S.), 2011).
Tissue collection and processingAfter 4 wks of photoperiod exposure, hamsters were
anesthetized with isoflurane vapors at zeitgeber time
(ZT; hours after lights on) 0 (N¼ 7 in LD; 6 in SD), ZT4
(N¼ 5 in LD; 9 in SD), ZT8 (N¼ 5 in LD; 6 in SD), ZT12
(N¼ 5 in LD; 5 in SD), ZT16 (N¼ 5 in LD; 4 in SD), and
ZT20 (N¼ 10 in LD; 5 in SD), and body mass was
measured. Then, hamsters were rapidly decapitated,
and brains were removed, placed in RNAlater (Ambion,
Austin, TX, USA), and stored at 4 �C to maintain mRNA
integrity for gene expression analysis. Five brains
collected at each time point of ZT4, ZT12, and ZT20
were cut along the anterior-posterior axis by using a
1090 T. Ikeno et al.
Chronobiology International
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

razor blade immediately after removal from a skull,
the half of the brain was placed in RNAlater and
the other half was processed for Golgi impregnation
using the FD Rapid GolgiStain Kit (FD
NeuroTechnologies, Ellicott City, MD, USA) according
to the manufacturer’s instructions. Testes, epididy-
mides, gonadal fat pads, and seminal vesicles were
also removed and weighed.
cDNA cloningTotal RNA was isolated using Trizol (Life Technologies,
Carlsbad, CA, USA) from two brains according to the
manufacturer’s instructions. cDNA was synthesized
from total RNA with Moloney murine leukemia virus
(M-MLV) reverse transcriptase (Life Technologies) and
random primers (Life Technologies). Fragments of Bdnf,
trkb, Vegf, and 18 S ribosomal RNA genes were obtained
by polymerase chain reaction (PCR) using PCR
Supermix (Life Technologies) according to the manu-
facturer’s instruction. Primers used were bdnf-1 F
(50-ACT CTG GAG AGC GTG AAT GG-30) and bdnf-1 R
(50-TGT CCA ATA AAT AGA TTG TAG AAC CA-30) for
Bdnf, and 18 S-1 F (50-GTC TAA GTA CGC ACG GCC
GG-30) and 18 S-1 R (50-CAT GCA CCA CCA CCC ACG
GA-30) for 18 S rRNA. For trkb, the primary PCR was
performed using trkb-1 F (50-TCT CCW GGC ATC GTG
GCA TT-30) and trkb-1 R (50-CCA GGC CRG CCC ATG
AAG TG-30) and the secondary PCR was performed with
10% of the primary PCR product using trkb-2 F (50-CCT
AAC CTC CAN GTG GAG GAA GG-30) and trkb-1 R.
For Vegf, the primary PCR was performed using vegf-1 F
(50-GCC AGC ACA TAG GAG AGA TGA-30) and vegf-1 R
(50-GGG CAG AGC TGA GTG TTA GC-30) and the
secondary PCR was performed with 10% of the primary
PCR product using vegf-1 F and vegf-2 R (50-CGA TCN
GGG AGA GAG AGA TTG G-30). PCR products were
cloned into plasmids using pGEM-T Easy Vector System
(Promega, Fitchburg, WI, USA) and sequenced at the
Plant-Genomics Center at the Ohio State University. The
determined partial sequences of 595, 496, 350, and
1239 bp for Bdnf, trkb, Vegf, and 18 S rRNA, respectively,
showed high similarities of more than 90% to the
corresponding genes in Mus musculus. Accession num-
bers (DDBJ/GenBank/EMBL) are AB794380 for Bdnf,
AB794381 for trkb, AB794382 for Vegf, and AB794386 for
18 S rRNA.
Quantitative real-time PCRThe temporal expression patterns of Bdnf, trkb, Vegf,
and Bmal1 in the hippocampus were investigated. The
hippocampus was dissected from one brain hemisphere
by gently separating it from the overlying cortex and the
underlying thalamus, and then total RNA was isolated as
mentioned above. DNAs in RNA samples were digested
with deoxyribonuclease I, Amplification Grade (Life
Technologies). cDNAs were synthesized from 1 mg of
RNA using High Capacity cDNA Reverse Transcription
Kit (Applied Biosystems, Foster City, CA, USA). For
real-time PCR analysis, 1% of the cDNA was used at a
final concentration of 1�Power SYBR Green PCR
Master Mix (Life Technologies) and 0.05 mM of each
primer using a 7500 Fast Real-Time PCR System (Life
Technologies) according to the manufacturer’s instruc-
tions. Each reaction was performed in duplicates.
Primers used were bdnf-3 F (50-TTA CCT GGA TGC
CGC AAA C-30) and bdnf-3 R (50-GCG GGC TGG GTC
AGA GT-30) for Bdnf, trkb-3 F (50-CCA CTC CCC ACG
ATG CA-30) and trkb-3 R (50-TTC ATT CGT GTG TTT GA
AAC C-30) for trkb, vegf-3 F (50-GGG TGA GAT TCC GGC
AGA A-30) and vegf-3 R (50-TTC CAT GAG GGA CCA TGC
TT-30) for Vegf, bmal1-1 F (50-GGC AGC GAT GGC TGT
CA-030) and bmal1-1 R (50-TCC ACC CAG GCC TGC AT-
30), which were designed from the P. sungorus Bmal1
sequence (DDBJ/GenBank/EMBL, AY316534), for
Bmal1, and 18 S-3 F (50-AGA AAC GGC TAC CAC ATC
CAA-30) and 18 S-3 R (50-GGG TCG GGA GTG GGT AAT
TT-30) for 18 S rRNA. In all the reactions, the generation
of only a single expected amplicon was confirmed by
melting analysis. Each reaction was done in duplicate.
Quantification of cDNAs was performed by the standard
curve methodology.
Analysis of neuronal morphologyBrains were sliced at 100mm on a cryostat and counter-
stained with cresyl violet (Sigma-Aldrich, MO, USA).
Hippocampal cell morphology was assessed in the CA1
and dentate gyrus (DG) fields in the dorsal hippocampus
(Figure 1A). Sections were visualized using a Nikon E800
bright-field microscope and intact neurons were recon-
structed using Neurolucida software (MicroBrightField,
Burlington, VT, USA) with a 20� objective. Six repre-
sentative neurons were traced per area, from each
animal. The criteria for neuronal selection were (1)
neurons had to be fully impregnated; (2) dendrites could
not be truncated; and (3) for the CA1 region, neurons
had to be pyramidal cells with somata lying within the
pyramidal cell layer and for the DG, neurons had to be
granule cells with somas lying within the granule cell
layer (Figure 1A). One brain sample collected at ZT12 in
long-day conditions was removed from morphological
analysis of dendrites, because the sections of this
sample were in a poor state and it was impossible to
trace the entire neuron. The cell traces were analyzed
using the accompanying NeuroExplorer software
(MicroBrightField). Cell body perimeter and size, the
number of branch points, total length of dendrites,
length of dendritic segments (intervals between soma
origin and the first branch point, between two succes-
sive branch points, or between the last branch point and
the branch end), length of segments, and the number of
primary dendrites were calculated (see Figure 1B). Sholl
analyses were also conducted. This method is used for
measuring the complexity of neurons by counting the
number of dendritic intersections with concentric cir-
cles centered on the cell body (Figure 1B). In our
analyses, the distance between concentric Sholl circles
Rhythmicity in Hippocampal Neuronal Morphology 1091
! Informa Healthcare USA, Inc.
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.
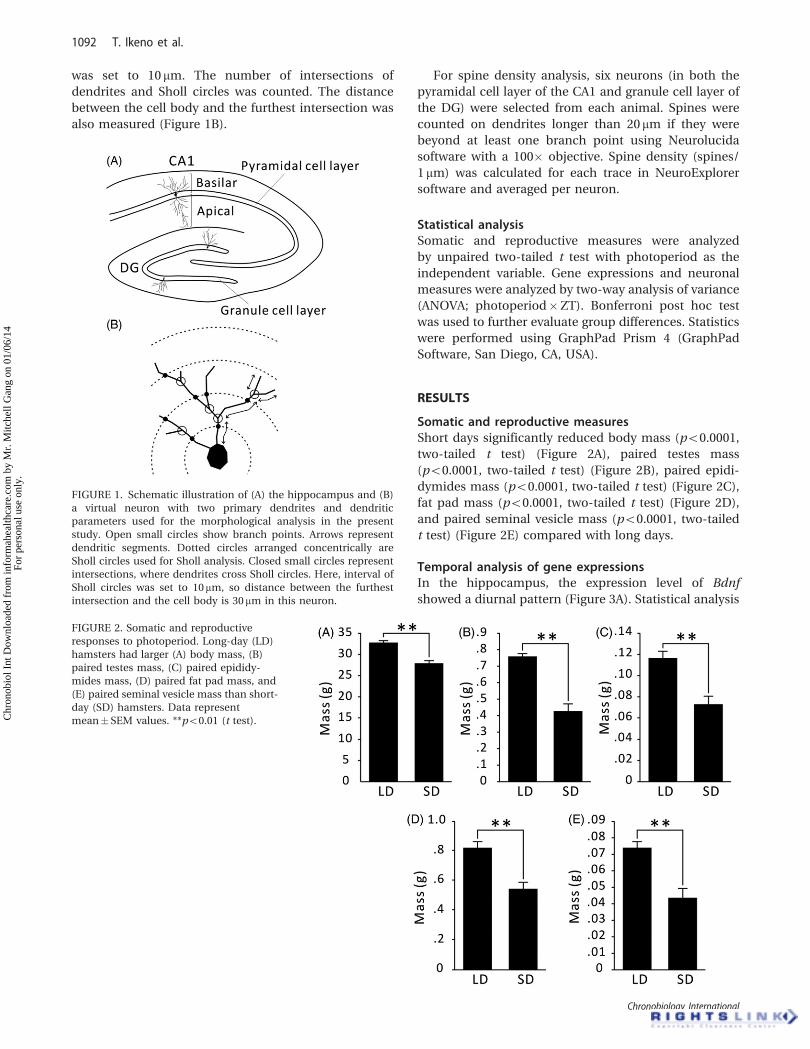
was set to 10 mm. The number of intersections of
dendrites and Sholl circles was counted. The distance
between the cell body and the furthest intersection was
also measured (Figure 1B).
For spine density analysis, six neurons (in both the
pyramidal cell layer of the CA1 and granule cell layer of
the DG) were selected from each animal. Spines were
counted on dendrites longer than 20 mm if they were
beyond at least one branch point using Neurolucida
software with a 100� objective. Spine density (spines/
1 mm) was calculated for each trace in NeuroExplorer
software and averaged per neuron.
Statistical analysisSomatic and reproductive measures were analyzed
by unpaired two-tailed t test with photoperiod as the
independent variable. Gene expressions and neuronal
measures were analyzed by two-way analysis of variance
(ANOVA; photoperiod�ZT). Bonferroni post hoc test
was used to further evaluate group differences. Statistics
were performed using GraphPad Prism 4 (GraphPad
Software, San Diego, CA, USA).
RESULTS
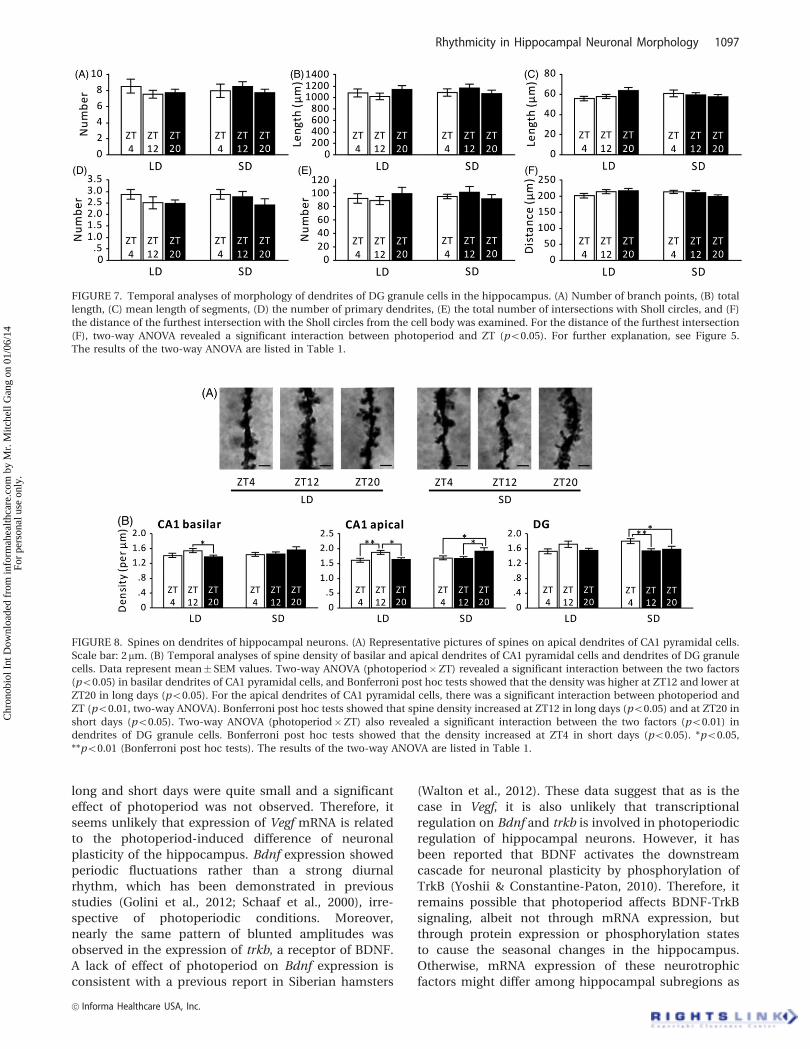
Somatic and reproductive measuresShort days significantly reduced body mass (p50.0001,
two-tailed t test) (Figure 2A), paired testes mass
(p50.0001, two-tailed t test) (Figure 2B), paired epidi-
dymides mass (p50.0001, two-tailed t test) (Figure 2C),
fat pad mass (p50.0001, two-tailed t test) (Figure 2D),
and paired seminal vesicle mass (p50.0001, two-tailed
t test) (Figure 2E) compared with long days.
Temporal analysis of gene expressionsIn the hippocampus, the expression level of Bdnf
showed a diurnal pattern (Figure 3A). Statistical analysis
FIGURE 2. Somatic and reproductive
responses to photoperiod. Long-day (LD)
hamsters had larger (A) body mass, (B)
paired testes mass, (C) paired epididy-
mides mass, (D) paired fat pad mass, and
(E) paired seminal vesicle mass than short-
day (SD) hamsters. Data represent
mean� SEM values. **p50.01 (t test).
FIGURE 1. Schematic illustration of (A) the hippocampus and (B)
a virtual neuron with two primary dendrites and dendritic
parameters used for the morphological analysis in the present
study. Open small circles show branch points. Arrows represent
dendritic segments. Dotted circles arranged concentrically are
Sholl circles used for Sholl analysis. Closed small circles represent
intersections, where dendrites cross Sholl circles. Here, interval of
Sholl circles was set to 10mm, so distance between the furthest
intersection and the cell body is 30mm in this neuron.
1092 T. Ikeno et al.
Chronobiology International
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

with two-way ANOVA (photoperiod�ZT) revealed a
significant main effect of ZT (p50.05). Further analysis
with Bonferroni post hoc tests showed that the Bdnf
mRNA abundance was higher at the beginning of the
light phase (ZT0) than that at the beginning of the dark
phase (ZT16) in long days (p50.05). A similar pattern
of fluctuation was observed in the trkb expression
level (Figure 3B); however, the effect of ZT was statis-
tically not significant (p¼ 0.054, two-way ANOVA
(photoperiod�ZT)). Although two-way ANOVA did not
detect any main effects of photoperiod and ZT on the
mRNA level of Vegf, Bonferroni post hoc tests showed
that the Vegf mRNA expression was higher at ZT0 than at
ZT16 in long days (p50.05) (Figure 3C). A clear diurnal
rhythm was observed in the expression of Bmal1 mRNA
(Figure 3D). Two-way ANOVA (photoperiod�ZT)
revealed significant main effects of photoperiod
(p50.05) and of ZT (p50.01). Further analysis with
Bonferroni post hoc tests showed that Bmal1 mRNA
expression was higher during the light phase (ZT0 and
ZT4) than at the beginning of the dark phase (ZT8) in
short days (p50.05). The results of the two-way ANOVA
are listed in Table 1.
Temporal analysis of cell body morphologyRepresentative tracings of hippocampal neurons are
shown in Figure 4A. Neither photoperiod nor ZT
nor an interaction between the two factors was
significant on the cell body perimeter or the cell
body area in CA1 pyramidal cells or DG granule cells
(p40.05, two-way ANOVA, for all cases) (Figure 4B,
C). The results of the two-way ANOVA are listed
in Table 1.
Temporal analysis of dendritic morphologyBasilar dendrites of pyramidal cells in the CA1 regionIn basilar dendrites of CA1 pyramidal cells, the num-
ber of branch points underwent a diurnal change
only in short days (Figure 5A). Two-way ANOVA
(photoperiod�ZT) revealed a significant main effect of
ZT (p50.01). Further analysis with Bonferroni post hoc
tests showed that the number of branch points signifi-
cantly increased at ZT20 in short days (p50.01).
Total length of dendrites increased in the dark phase
(Figure 5B). Two-way ANOVA (photoperiod�ZT)
revealed a significant main effect of ZT (p50.01) and
Bonferroni post hoc tests showed that length of den-
drites was significantly longer in the dark phase than
that at the light phase in long days (p50.05). Although a
similar apparent increase of dendritic length at ZT20
was observed in short days, it was not statistically
significant (p40.05, Bonferroni post hoc test). A signifi-
cant interaction between photoperiod and ZT was
detected in mean length of segments (p50.01, two-
way ANOVA) (Figure 5C). Bonferroni post hoc tests
showed that mean length of segments decreased at ZT20
only in short days (p50.05). Two-way ANOVA
(photoperiod�ZT) detected a significant main effect
of ZT (p50.05) in the number of primary dendrites
(Figure 5D). Bonferroni post hoc tests showed that
primary dendrites increased at ZT4 and decreased
at ZT12 only in long days (p50.05). A diurnal rhythm
was also observed in the total number of intersections
in the Sholl analysis (Figure 5E). Two-way ANOVA
(photoperiod�ZT) revealed a significant main effect of
ZT (p50.01) and Bonferroni post hoc tests showed that
intersections significantly increased at ZT20 in long days
(p50.05). Two-way ANOVA (photoperiod�ZT) revealed
FIGURE 3. Temporal expression patterns
of hippocampal genes in long days (closed
circles and solid line) and short days (open
circles and broken line). (A) For Bdnf there
was a main effect of ZT (p50.05, two-way
ANOVA [photoperiod�ZT]). Bonferroni
post hoc tests revealed that the Bdnf
mRNA expression was higher at ZT0 than
that at ZT16 in long days (p50.05). (B) For
trkb there were no effects of photoperiod
and ZT, and no interaction between the
two factors (p40.05, two-way ANOVA). (C)
For Vegf two-way ANOVA showed no main
effects of photoperiod and ZT, and no
interaction between the two factors
(p40.05). However, Bonferroni post hoc
tests revealed that the Vegf mRNA expres-
sion was higher at ZT0 than that at ZT16 in
long days (p50.05). (D) For bmal1 there
were main effects of photoperiod and ZT
(p50.05 and p50.01, respectively, two-
way ANOVA). Bonferroni post hoc tests
showed that bmal1 mRNA expression was
higher at ZT0 and ZT4 than that at ZT8 in
short days (p50.05). The results of the
two-way ANOVA are listed in Table 1.
Rhythmicity in Hippocampal Neuronal Morphology 1093
! Informa Healthcare USA, Inc.
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

a significant interaction between the two factors when
we analyzed the distance of the furthest intersection
with the Sholl circles from the cell body (p50.05)
(Figure 5F). The distance at ZT20 was longer than that at
ZT4 and that at ZT12 only in long days (p50.01 and
p50.05, respectively, Bonferroni post hoc tests). The
results of the two-way ANOVA are listed in Table 1.
Apical dendrites of pyramidal cells in the CA1 regionIn apical dendrites of CA1 pyramidal cells, neither
photoperiod nor ZT affected the number of branch
points (Figure 6A) or total length of dendrites (p40.05,
two-way ANOVA, for both cases) (Figure 6B). Two-way
ANOVA (photoperiod�ZT) revealed a significant inter-
action between the two factors in mean length of
segments (p50.01) (Figure 6C). Further analysis with
Bonferroni post hoc tests showed that the mean length
of segments at ZT20 was longer than that at ZT12 only in
long days (p50.05). The number of primary dendrites
displayed diurnal rhythms (Figure 6D). Two-way
ANOVA (photoperiod�ZT) detected a significant main
effect of ZT (p50.05), and Bonferroni post hoc tests
showed that branch points increased at ZT4 and
decreased at ZT20 only in long days (p50.01). The
total number of intersections with the Sholl circles was
not statistically significant (p40.05, two-way ANOVA)
(Figure 6E). The distance of the furthest intersection
with the Sholl circles from the cell body did not display
any diurnal changes (Figure 6F) (p40.05, two-way
ANOVA). The results of the two-way ANOVA are listed
in Table 1.
Dendrites of granule cells in the DG regionIn DG granule cells, the effect of photoperiod or ZT, or
an interaction between the two factors, was not signifi-
cant on most dendritic morphology examined here,
including the number of branch points (Figure 7A), total
length of dendrites (Figure 7B), mean length of
TABLE 1. Statistical results.
p values from the two-way ANOVA
Variable Photoperiod ZT Interaction
Gene expressions (Figure 3)
Bdnf 0.984 0.027* 0.781
trkb 0.370 0.054 0.863
Vegf 0.992 0.221 0.826
Bmal1 0.027* 0.009** 0.963
Cell body morphology (Figure 4)
CA1
Cell body perimeter 0.478 0.158 0.431
Cell body area 0.444 0.361 0.297
DG
Cell body perimeter 0.081 0.060 0.561
Cell body area 0.075 0.131 0.319
Dendritic morphology (Figures 5, 6, 7)
CA1 basilar
Number of branch points 0.339 0.005** 0.282
Total length of dendrites 0.260 0.003** 0.653
Mean length of segments 0.412 0.679 0.004**
Number of primary dendrites 0.369 0.029* 0.112
Total number of intersections 0.245 0.007** 0.692
Distance of the furthest intersection 0.457 0.126 0.018*
CA1 apical
Number of branch points 0.870 0.839 0.668
Total length of dendrites 0.715 0.584 0.422
Mean length of segments 0.744 0.808 0.004**
Number of primary dendrites 0.318 0.017* 0.321
Total number of intersections 0.838 0.295 0.406
Distance of the furthest intersection 0.260 0.256 0.552
DG
Number of branch points 0.660 0.841 0.497
Total length of dendrites 0.590 0.947 0.221
Mean length of segments 0.852 0.670 0.077
Number of primary dendrites 0.694 0.148 0.750
Total number of intersections 0.608 0.933 0.190
Distance of the furthest intersection 0.661 0.535 0.041
Spine density (Figure 8)
CA1 basilar 0.268 0.331 0.021*
CA1 apical 0.298 0.076 0.001**
DG 0.356 0.318 0.003**
Significant differences: *p50.05 and **p50.01.
1094 T. Ikeno et al.
Chronobiology International
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

segments (Figure 7C), number of primary dendrites
(Figure 7D), and total number of intersections
(Figure 7E) (p40.05, two-way ANOVA, for all cases).
A significant interaction between photoperiod and ZT
was detected only in the distance of the furthest
intersection with the Sholl circles from the cell body
(p50.05, two-way ANOVA) (Figure 7F). The results of
the two-way ANOVA are listed in Table 1.
Temporal analysis of spine densityBasilar dendrites of pyramidal cells in the CA1 regionRepresentative pictures of spines on dendrites are shown
in Figure 8A. The spine density of basilar dendrites
of CA1 pyramidal cells showed a diurnal rhythm
(Figure 8B). Two-way ANOVA (photoperiod�ZT)
revealed a significant interaction between the two factors
(p50.05) and Bonferroni post hoc tests showed that
the density was elevated at ZT12 and decreased at ZT20
in long days (p50.05). The results of the two-way
ANOVA are listed in Table 1.
Apical dendrites of pyramidal cells in the CA1 regionA significant interaction between photoperiod and
ZT was detected in the spine density in apical dendrites
of CA1 pyramidal cells (p50.01, two-way ANOVA)
(Figure 8B). Bonferroni post hoc tests showed that the
spine density was increased at ZT12 compared with that
at ZT4 and that at ZT20 in long days (p50.01 and
p50.05, respectively), but was elevated at ZT20 in short
days (p50.05). The results of the two-way ANOVA are
listed in Table 1.
Dendrites of granule cells in the DG regionTwo-way ANOVA (photoperiod�ZT) revealed a signifi-
cant interaction between the two factors in the spine
density of dendrites of DG granule cells (p50.01)
(Figure 8B). Further analysis with Bonferroni post hoc
tests detected significant differences among ZTs only in
short days: spine density was higher in the light phase
than that at ZT12 and that at ZT20 (p50.01 and p50.05,
respectively). The results of the two-way ANOVA are
listed in Table 1.
DISCUSSION
The present study demonstrated that Bmal1 expression
in the hippocampus displays a diurnal rhythm, con-
firming previous reports showing the rhythmic expres-
sion of circadian clock genes (e.g., Golini et al., 2012;
Jilg et al., 2010). Notably, Bmal1 mRNA was highly
expressed with elevated amplitude in short days
compared with long days. The effects of photoperiod
on expression levels of multiple circadian clock genes
have been demonstrated in the pars tuberalis, where the
FIGURE 4. Neuronal morphology of hip-
pocampal neurons. (A) Representative
tracings of CA1 pyramidal cells and DG
granule cells. Scale bar: 100 mm. Temporal
analyses of (B) cell body perimeter and (C)
cell body area of CA1 pyramidal cells and
DG granule cells. Data represent
mean� SEM values. Two-way ANOVA
(photoperiod�ZT) detected no main
effects of photoperiod and ZT, and no
interaction between the two factors for
both cell body perimeter and cell body
area (p40.05, for all cases). The results of
the two-way ANOVA are listed in Table 1.
Rhythmicity in Hippocampal Neuronal Morphology 1095
! Informa Healthcare USA, Inc.
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

seasonal signal of melatonin is decoded, of several
seasonal mammals including Siberian hamsters; it has
thus been proposed that regulation of circadian clock
genes may play a key role for decoding of melatonin
signal (Johnston et al., 2005; Lincoln et al., 2002;
Messager et al., 1999, 2000). Therefore, different expres-
sion levels of Bmal1 revealed here suggest a possibil-
ity that photoperiodic information in the form of
hormonal or other signals might be processed also in
the hippocampus via expression patterns of circadian
clock genes.
VEGF and BDNF are the most prevalent neurotrophic
factors, which are thought to stimulate neuronal
reorganization in the adult brain (Fournier & Duman,
2012; Yu & Chen, 2011). In contrast to the short-day
elevation of Bmal1 amplitude, the diurnal oscillation of
Vegf expression was detectable only in long days.
However, the different expression patterns between
FIGURE 5. Temporal analyses of morphology of basilar dendrites of CA1 pyramidal cells in the hippocampus. Data represent mean�SEM
values. (A) For the number of branch points, two-way ANOVA (photoperiod�ZT) revealed a main effect of ZT (p50.01). Bonferroni post
hoc tests showed that the number of branch points increased at ZT20 in short days (p50.01). (B) For total length of dendrites, two-way
ANOVA (photoperiod�ZT) revealed a main effect of ZT (p50.01), and Bonferroni post hoc tests revealed that length of dendrites increased
at ZT20 in long days (p50.05). (C) For mean length of segments, a significant interaction between photoperiod and ZT was detected by
two-way ANOVA (p50.01). Bonferroni post hoc tests showed that mean length of segments decreased at ZT20 in short days (p50.05). (D)
For the number of primary dendrites, there was a main effect of ZT (p50.01, two-way ANOVA (photoperiod�ZT)). Bonferroni post hoc
tests showed that the number of primary dendrites was elevated at ZT4 in long days (p50.01). (E) For the total number of intersections with
the Sholl circles, there was a main effect of ZT (p50.01, two-way ANOVA [photoperiod�ZT]), and Bonferroni post hoc tests showed that
the number of intersections increased at ZT20 in long days (p50.05). (F) For the distance of the furthest intersection with the Sholl circles
from the cell body, two-way ANOVA (photoperiod�ZT) revealed a significant interaction between the two factors (p50.05). Bonferroni
post hoc tests showed that the distance increased at ZT20 compared with those at ZT4 and ZT12 (p50.01 and p50.05, respectively).
*p50.05, **p50.01 (Bonferroni post hoc tests). The results of the two-way ANOVA are listed in Table 1.
FIGURE 6. Temporal analyses of morphology of apical dendrites of CA1 pyramidal cells in the hippocampus. (A) Number of branch points,
(B) total length, (C) mean length of segments, (D) the number of primary dendrites, (E) the total number of intersections with Sholl circles,
and (F) the distance of the furthest intersection with the Sholl circles from the cell body was examined. For mean length of segments (D), a
significant interaction between photoperiod and ZT was detected by two-way ANOVA (p50.01). Bonferroni post hoc tests showed that
mean length of segments increased at ZT20 in long days (p50.05). For the number of primary dendrites (D), two-way ANOVA
(photoperiod�ZT) revealed a main effect of ZT (p50.01), and Bonferroni post hoc tests showed that the number of primary dendrites
increased at ZT4 compared with that at ZT20 in long days (p50.01). For further explanation, see Figure 5. The results of the two-way
ANOVA are listed in Table 1.
1096 T. Ikeno et al.
Chronobiology International
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.
long and short days were quite small and a significant
effect of photoperiod was not observed. Therefore, it
seems unlikely that expression of Vegf mRNA is related
to the photoperiod-induced difference of neuronal
plasticity of the hippocampus. Bdnf expression showed
periodic fluctuations rather than a strong diurnal
rhythm, which has been demonstrated in previous
studies (Golini et al., 2012; Schaaf et al., 2000), irre-
spective of photoperiodic conditions. Moreover,
nearly the same pattern of blunted amplitudes was
observed in the expression of trkb, a receptor of BDNF.
A lack of effect of photoperiod on Bdnf expression is
consistent with a previous report in Siberian hamsters
(Walton et al., 2012). These data suggest that as is the
case in Vegf, it is also unlikely that transcriptional
regulation on Bdnf and trkb is involved in photoperiodic
regulation of hippocampal neurons. However, it has
been reported that BDNF activates the downstream
cascade for neuronal plasticity by phosphorylation of
TrkB (Yoshii & Constantine-Paton, 2010). Therefore, it
remains possible that photoperiod affects BDNF-TrkB
signaling, albeit not through mRNA expression, but
through protein expression or phosphorylation states
to cause the seasonal changes in the hippocampus.
Otherwise, mRNA expression of these neurotrophic
factors might differ among hippocampal subregions as
FIGURE 8. Spines on dendrites of hippocampal neurons. (A) Representative pictures of spines on apical dendrites of CA1 pyramidal cells.
Scale bar: 2 mm. (B) Temporal analyses of spine density of basilar and apical dendrites of CA1 pyramidal cells and dendrites of DG granule
cells. Data represent mean� SEM values. Two-way ANOVA (photoperiod�ZT) revealed a significant interaction between the two factors
(p50.05) in basilar dendrites of CA1 pyramidal cells, and Bonferroni post hoc tests showed that the density was higher at ZT12 and lower at
ZT20 in long days (p50.05). For the apical dendrites of CA1 pyramidal cells, there was a significant interaction between photoperiod and
ZT (p50.01, two-way ANOVA). Bonferroni post hoc tests showed that spine density increased at ZT12 in long days (p50.05) and at ZT20 in
short days (p50.05). Two-way ANOVA (photoperiod�ZT) also revealed a significant interaction between the two factors (p50.01) in
dendrites of DG granule cells. Bonferroni post hoc tests showed that the density increased at ZT4 in short days (p50.05). *p50.05,
**p50.01 (Bonferroni post hoc tests). The results of the two-way ANOVA are listed in Table 1.
FIGURE 7. Temporal analyses of morphology of dendrites of DG granule cells in the hippocampus. (A) Number of branch points, (B) total
length, (C) mean length of segments, (D) the number of primary dendrites, (E) the total number of intersections with Sholl circles, and (F)
the distance of the furthest intersection with the Sholl circles from the cell body was examined. For the distance of the furthest intersection
(F), two-way ANOVA revealed a significant interaction between photoperiod and ZT (p50.05). For further explanation, see Figure 5.
The results of the two-way ANOVA are listed in Table 1.
Rhythmicity in Hippocampal Neuronal Morphology 1097
! Informa Healthcare USA, Inc.
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

observed in the morphological change of neurons
and, therefore, we could not detect clear diurnal or
photoperiodic changes because we analyzed the whole
hippocampus. It would be useful to compare the
expression patterns of neurotrophic factors among the
hippocampal subregions in the future.
The adult brain has a remarkable capacity for struc-
tural plasticity. To cope with dramatic seasonal envir-
onmental changes, reversible remodeling of neural
circuits occurs in response to photoperiod or even
within a shorter period within a single day (Holtmaat
et al., 2005; Magarinos et al., 2006: Perez-Cruz et al.,
2009; Popov et al., 1992; Pyter et al., 2005; Workman
et al., 2011). Here we demonstrated that hippocampal
neuronal morphology displays a diurnal rhythm in a
subregion-specific manner; moreover, photoperiod
affects the morphological changes differently. Many
parameters of dendrite morphology in CA1 neurons
showed diurnal changes both in long and short days.
Although the pattern of diurnal changes is similar in
basilar and apical dendrites, these changes are only
significant mainly in the basilar dendrites, suggesting
that basilar dendrites are more flexible to daily envir-
onmental fluctuations than apical dendrites. Our results
are consistent with the report that diurnal difference in
the dendritic architecture are more pronounced in the
basilar dendrites than apical dendrites of layer III
pyramidal neurons in the rat infralimbic cortex (Perez-
Cruz et al., 2009). In contrast to CA1 neurons, DG
neurons did not display diurnal morphological changes
in either long or short days. Our results suggest that the
effects of time of day and photoperiod are different
among subregions in the hippocampus. Hippocampal
subregions are functionally and molecularly distinct and
are organized in an interconnecting complex circuit
(Small et al., 2011). Because the CA1 region provides the
main hippocampal outflow (van Strien et al., 2009), the
diurnal change of dendritic structure in CA1 neurons
might contribute to the diurnal variation of hippocam-
pal function, including learning and memory
(Valentinuzzi et al., 2004, 2008).
Overt rhythmicity of most dendritic parameters in
CA1 neurons is observed only in long days or only in
short days. These results suggest that photoperiod
affects the diurnal morphological change differently.
In short days, the number of branches increases at ZT20,
indicating that they produce new branches around this
time. Addition of branch points along the dendrite
would cause shortening of an interval between succes-
sive branch points, resulting in a decrease of segment
length, as observed at ZT20 in short days. However,
generation of new branches does not lead to a signifi-
cant increase of dendritic length at ZT20. One possible
interpretation is that the newly produced branches are
too short to increase total dendritic length. This also
could explain that an increase of branches does not
affect the number of intersections with Sholl circles.
No rhythmicity in the furthest intersection distance
suggests that the covered area by dendrites does not
vary during a day. Overall, these data indicate that CA1
neurons increase their morphological complexity with
increased branches mainly in basilar dendrites during
the latter half of the dark phase in short days. In long
days, however, total dendritic length, intersections with
Sholl circles, and the furthest intersection distance
increases at ZT20, suggesting that dendrites become
longer and cover a larger area during the dark phase of
long days. One would expect that increases of these
parameters are attributed to addition of new branches.
However, this is unlikely because the number of branch
points does not significantly change during long days.
Another possibility might be that the changes of total
dendritic length, intersections, and the furthest inter-
section distance are derived from extension of dendrites.
However, this possibility can be dismissed because
segment length at ZT20 was not significantly affected in
long days. Otherwise, the two processes described above
would occur at the same time. In this case, even though
segment length is extended to lead an increase of total
dendritic length at ZT20, a slight, but not significant,
increase of branch points would shorten intervals
between two successive branch points, and hence
there would be no net change in mean segment
length. Although it remains unclear what morphological
changes CA1 neurons undergo at the present, our
present results suggest photoperiod affects diurnal
changes of morphological parameters in CA1 neurons.
In addition to elevated morphological characteristics
during the dark phase, CA1 neurons have more primary
dendrites at ZT4 in long, but not, short days. It has been
reported that most inhibitory synapses in CA1 neurons
are distributed on proximal primary dendrites (Megıas
et al., 2001). Therefore, the morphological changes
revealed here suggest that inhibition of CA1 neurons
increases at morning only in long days.
The present study also demonstrated a diurnal
change of spine density in both CA1 and DG regions.
Different patterns were again observed between long
and short days. In the CA1 region, where rhythmicity is
more robust in apical dendrites than in basilar den-
drites, spine density increases during the latter half of
the light phase in long days but it increases during the
latter half of the dark phase in short days. In the DG
region, dendrites have dense spines during the light
phase, but the diurnal difference is apparent only in
short days. Spines increase the surface area of dendrites,
which increases the excitatory synaptic density and
number of connections between neurons (Sorra &
Harris, 2000). However, spine density and dendritic
complexity showed different peak time in long days,
suggesting that these dendritic properties have distinct
functions and are regulated by different mechanisms.
Taken together, these data indicate that photoperiod
induces a different diurnal change of dendritic pattern-
ing and a different peak time of spine density in a
subregion-specific manner. Hippocampal neuronal
1098 T. Ikeno et al.
Chronobiology International
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

morphology has been suggested to play an important
role in depressive-like behaviors (Bedrosian et al., 2011;
Workman et al., 2011). Our present results are consistent
with the hypothesis that photoperiod-induced adjust-
ments of hippocampal neuronal dynamics might under-
lie seasonal difference of affective responses.
DECLARATION OF INTEREST
T.I. was supported by a JSPS Postdoctoral Fellowshipsfor Research Abroad.
The authors report no conflicts of interest. Theauthors alone are responsible for the content andwriting of the paper.
REFERENCES
Adan A, Sanchez-Turet M. (2001). Gender differences in diurnal
variations of subjective activation and mood. Chronobiol Int,
18, 491–502.
Bedrosian TA, Fonken LK, Walton JC, et al. (2011). Dim light at
night provokes depression-like behaviors and reduces CA1
dendritic spine density in female hamsters.
Psychoneuroendocrinology, 36, 1062–9.
Blanco AF, Garcıa AL, Inda AM, Errecalde AL. (2011). Vascular
endothelial growth factor expression along a circadian time
span in intact adult mice liver. Biol Rhythm Res, 42, 141–6.
Breedlove SM, Jordan CL. (2001). The increasingly plastic,
hormone-responsive adult brain. Proc Natl Acad Sci U S A,
98, 2956–7.
Brenowitz EA, Margoliash D, Nordeen KW. (1997). An introduction
to birdsong and the avian song system. J Neurobiol, 33,
495–500.
Burger DK, Saucier JM, Iwaniuk AN, Saucier DM. (2013). Seasonal
and sex differences in the hippocampus of a wild rodent. Behav
Brain Res, 236, 131–8.
Cole J, Costafreda SG, McGuffin P, Fu CH. (2011). Hippocampal
atrophy in first episode depression: a meta-analysis of magnetic
resonance imaging studies. J Affect Disord, 134, 483–7.
Dolci C, Montaruli A, Roveda E, et al. (2003). Circadian variations
in expression of the trkB receptor in adult rat hippocampus.
Brain Res, 994, 67–72.
Fournier NM, Duman RS. (2012). Role of vascular endothelial
growth factor in adult hippocampal neurogenesis: implications
for the pathophysiology and treatment of depression. Behav
Brain Res, 227, 440–9.
Galea LA, McEwen BS. (1999). Sex and seasonal differences in the
rate of cell proliferation in the dentate gyrus of adult wild
meadow voles. Neuroscience, 89, 955–64.
Goldman BD. (2001). Mammalian photoperiodic system: formal
properties and neuroendocrine mechanisms of photoperiodic
time measurement. J Biol Rhythms, 16, 283–301.
Golini RS, Delgado SM, Navigatore Fonzo LS, et al. (2012). Daily
patterns of clock and cognition-related factors are modified in
the hippocampus of vitamin A-deficient rats. Hippocampus, 22,
1720–32.
Hajszan T, Dow A, Warner-Schmidt JL, et al. (2009). Remodeling of
hippocampal spine synapses in the rat learned helplessness
model of depression. Biol Psychiatry, 65, 392–400.
Hajszan T, Szigeti-Buck K, Sallam NL, et al. (2010). Effects of
estradiol on learned helplessness and associated remodeling of
hippocampal spine synapses in female rats. Biol Psychiatry, 67,
168–74.
Holtmaat AJ, Trachtenberg JT, Wilbrecht L, et al. (2005). Transient
and persistent dendritic spines in the neocortex in vivo.
Neuron, 45, 279–91.
Institute of Laboratory Animal Resources (U.S.). (2011). Guide for
the care and use of laboratory animals. 8th ed. Washington, DC:
National Academy Press, 220p.
Jacobson L, Sapolsky R. (1991). The role of the hippocampus in
feedback regulation of the hypothalamic-pituitary-adrenocor-
tical axis. Endocr Rev, 12, 118–34.
Jilg A, Lesny S, Peruzki N, et al. (2010). Temporal dynamics of
mouse hippocampal clock gene expression support memory
processing. Hippocampus, 20, 377–88.
Johnston JD, Ebling FJ, Hazlerigg DG. (2005). Photoperiod regu-
lates multiple gene expression in the suprachiasmatic nuclei
and pars tuberalis of the Siberian hamster (Phodopus sungorus).
Eur J Neurosci, 21, 2967–74.
Karakas� A, Cos� kun H, Kaya A, et al. (2011). The effects of the
intraamygdalar melatonin injections on the anxiety like behav-
ior and the spatial memory performance in male Wistar rats.
Behav Brain Res, 222, 141–50.
Kaya A, Karakas� A, Cos� kun H. (2011). The effects of time of the day
and the pinealectomy on anxiety-like behaviour in male Wister
rats. Biol Rhythm Res, 42, 367–83.
Ko CH, Takahashi JS. (2006). Molecular components of the
mammalian circadian clock. Hum Mol Genet, 15, R271–7.
Koyanagi S, Kuramoto Y, Nakagawa H, et al. (2003). A molecular
mechanism regulating circadian expression of vascular endo-
thelial growth factor in tumor cells. Cancer Res, 63, 7277–83.
Lincoln G, Messager S, Andersson H, Hazlerigg D. (2002). Temporal
expression of seven clock genes in the suprachiasmatic nucleus
and the pars tuberalis of the sheep: evidence for an internal
coincidence timer. Proc Natl Acad Sci U S A, 99, 13890–5.
Magarinos AM, McEwen BS. (1995). Stress-induced atrophy of
apical dendrites of hippocampal CA3c neurons: involvement of
glucocorticoid secretion and excitatory amino acid receptors.
Neuroscience, 69, 89–98.
Magarinos AM, McEwen BS, Saboureau M, Pevet P. (2006). Rapid
and reversible changes in intrahippocampal connectivity
during the course of hibernation in European hamsters. Proc
Natl Acad Sci U S A, 103, 18775–80.
Messager S, Ross AW, Barrett P, Morgan PJ. (1999). Decoding
photoperiodic time through Per1 and ICER gene amplitude.
Proc Natl Acad Sci U S A, 96, 9938–43.
Messager S, Hazlerigg DG, Mercer JG, Morgan PJ. (2000).
Photoperiod differentially regulates the expression of Per1 and
ICER in the pars tuberalis and the suprachiasmatic nucleus of
the Siberian hamster. Eur J Neurosci, 12, 2865–70.
Neumeister A, Wood S, Bonne O, et al. (2005). Reduced
hippocampal volume in unmedicated, remitted patients with
major depression versus control subjects. Biol Psychiatry, 57,
935–7.
Owens DS, Macdonald I, Tucker P, et al. (2000). Diurnal variations
in the mood and performance of highly practised young women
living under strictly controlled conditions. Br J Psychol, 91,
41–60.
Perez-Cruz C, Simon M, Flugge G, et al. (2009). Diurnal rhythm and
stress regulate dendritic architecture and spine density of
pyramidal neurons in the rat infralimbic cortex. Behav Brain
Res, 205, 406–13.
Perrot-Sinal TS, Kavaliers M, Ossenkopp KP. (1998). Spatial
learning and hippocampal volume in male deer mice: relations
to age, testosterone and adrenal gland weight. Neuroscience,
86, 1089–99.
Pevet P. (1988). The role of the pineal gland in the photoperiodic
control of reproduction in different hamster species. Reprod
Nutr Dev, 28, 443–58.
Popov VI, Bocharova LS, Bragin AG. (1992). Repeated changes of
dendritic morphology in the hippocampus of ground squirrels
in the course of hibernation. Neuroscience, 48, 45–51.
Prendergast BJ, Nelson RJ. (2005). Affective responses to changes
in day length in Siberian hamsters (Phodopus sungorus).
Psychoneuroendocrinology, 30, 438–52.
Rhythmicity in Hippocampal Neuronal Morphology 1099
! Informa Healthcare USA, Inc.
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.

Pyter LM, Nelson RJ. (2006). Enduring effects of photoperiod on
affective behaviors in Siberian hamsters (Phodopus sungorus).
Behav Neurosci, 120, 125–34.
Pyter LM, Reader BF, Nelson RJ. (2005). Short photoperiods impair
spatial learning and alter hippocampal dendritic morphology
in adult male white-footed mice (Peromyscus leucopus).
J Neurosci, 25, 4521–6.
Schaaf MJ, Duurland R, de Kloet ER, Vreugdenhil E. (2000).
Circadian variation in BDNF mRNA expression in the rat
hippocampus. Brain Res Mol Brain Res, 75, 342–4.
Scherbarth F, Steinlechner S. (2010). Endocrine mechanisms
of seasonal adaptation in small mammals: from early
results to present understanding. J Comp Physiol B, 180,
935–52.
Small SA, Schobel SA, Buxton RB, et al. (2011). A pathophysio-
logical framework of hippocampal dysfunction in ageing and
disease. Nat Rev Neurosci, 12, 585–601.
Smith GT, Brenowitz EA, Beecher MD, Wingfield JC. (1997).
Seasonal changes in testosterone, neural attributes of song
control nuclei, and song structure in wild songbirds. J Neurosci,
17, 6001–10.
Sorra KE, Harris KM. (2000). Overview on the structure, compos-
ition, function, development, and plasticity of hippocampal
dendritic spines. Hippocampus, 10, 501–11.
Stockmeier CA, Mahajan GJ, Konick LC, et al. (2004). Cellular
changes in the postmortem hippocampus in major depression.
Biol Psychiatry, 56, 640–50.
Tramontin AD, Brenowitz EA. (2000). Seasonal plasticity in the
adult brain. Trends Neurosci, 23, 251–8.
Tramontin AD, Perfito N, Wingfield JC, Brenowitz EA. (2001).
Seasonal growth of song control nuclei precedes seasonal
reproductive development in wild adult song sparrows. Gen
Comp Endocrinol, 122, 1–9.
Valentinuzzi VS, Menna-Barreto L, Xavier GF. (2004). Effect of
circadian phase on performance of rats in the Morris water
maze task. J Biol Rhythms, 19, 312–24.
Valentinuzzi VS, Neto SP, Carneiro BT, et al. (2008). Memory for
time of training modulates performance on a place condition-
ing task in marmosets. Neurobiol Learn Mem, 89, 604–7.
van Strien NM, Cappaert NL, Witter MP. (2009). The anatomy of
memory: an interactive overview of the parahippocampal-
hippocampal network. Nat Rev Neurosci, 10, 272–82.
Walton JC, Grier AJ, Weil ZM, Nelson RJ. (2012). Photoperiod and
stress regulation of corticosteroid receptor, brain-derived
neurotrophic factor, and glucose transporter GLUT3 mRNA in
the hippocampus of male Siberian hamsters (Phodopus
sungorus). Neuroscience, 213, 106–11.
Wang WY, Dong JH, Liu X, et al. (2005). Vascular endothelial
growth factor and its receptor Flk-1 are expressed in the
hippocampus following entorhinal deafferentation.
Neuroscience, 134, 1167–78.
Watanabe Y, Gould E, Cameron HA, et al. (1992). Phenytoin
prevents stress- and corticosterone-induced atrophy of CA3
pyramidal neurons. Hippocampus, 2, 431–5.
Workman JL, Nelson RJ. (2011). Potential animal models of
seasonal affective disorder. Neurosci Biobehav Rev, 35, 669–79.
Workman JL, Manny N, Walton JC, Nelson RJ. (2011). Short day
lengths alter stress and depressive-like responses, and hippo-
campal morphology in Siberian hamsters. Horm Behav, 60,
520–8.
Yoshii A, Constantine-Paton M. (2010). Postsynaptic BDNF-TrkB
signaling in synapse maturation, plasticity, and disease. Dev
Neurobiol, 70, 304–22.
Yu H, Chen ZY. (2011). The role of BDNF in depression on the basis
of its location in the neural circuitry. Acta Pharmacol Sin, 32,
3–11.
1100 T. Ikeno et al.
Chronobiology International
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
r. M
itche
ll G
ang
on 0
1/06
/14
For
pers
onal
use
onl
y.



![EARLY FLOWERING3 Redundancy Fine-Tunes Photoperiod … · EARLY FLOWERING3 Redundancy Fine-Tunes Photoperiod Sensitivity1[OPEN] Andrew J.S. Rubenach, Valérie Hecht, Jacqueline K](https://img.pdfslide.us/doc/110x75/5f70a7e86c02c415f04ab3da/early-flowering3-redundancy-fine-tunes-photoperiod-early-flowering3-redundancy-fine-tunes.jpg)

























