Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 82, pp. 3164-3168, May 1985Biochemistry
Phosphorylation of DNA topoisomerase II by casein kinase II:Modulation of eukaryotic topoisomerase II activity in vitro
(supercoiled DNA/phosphoserine/alkaline phosphatase/Drosophila melanogaster)
PAT ACKERMAN*, CLAIBORNE V. C. GLOVERt, AND NEIL OSHEROFF*t*Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, TN 37232; and tDepartment of Biochemistry, University of Georgia,Athens, GA 30602
Communicated by Stanley Cohen, January 18, 1985
ABSTRACT The phosphorylation of Drosophilamelanogaster DNA topoisomerase II by purified casein kinaseII was characterized in vitro. Under the conditions used, thekinase incorporated a maximum of 2-3 molecules of phosphateper homodimer oftopoisomerase II. No autophosphorylation ofthe topoisomerase was observed. The only amino acid residuemodified by casein kinase II was serine. Apparent Km and Vm..values for the phosphorylation reaction were 0.4 ,.Mtopoisomerase II and 3.3 ,umol of phosphate incorporated permin per mg of kinase, respectively. Phosphorylation stimulatedthe DNA relaxation activity of topoisomerase II by 3-fold overthat of the dephosphorylated enzyme, and the effects ofmodification could be reversed by treatment with alkalinephosphatase. Therefore, this study demonstrates that post-translational enzymatic modifications can be used to modulatethe interaction between topoisomerase II and DNA.
The topology of DNA has a profound influence on how itsgenetic information is regenerated, rearranged, and ex-pressed in vivo (1, 2). Consequently, the enzymes that affectthe topological structure of nucleic acids play a crucial rolein controlling the cellular functions ofDNA (1-4). One classof enzymes, the type II topoisomerases, catalyzes changes inthe topological state of nucleic acids by passing one intactDNA helix through a transient double-stranded break madein a second helix (5-7). These ubiquitous enzymes areessential for the viability of eukaryotic cells (8-10) and areinvolved in many aspects of DNA metabolism (1-4), includ-ing replication' (11-17), repair (18), transcription (13), andchromosome segregation (9, 10).
Despite the importance of type II topoisomerases toeukaryotic organisms, little information exists concerning thephysiological regulation of these enzymes. Although theiractivity is stimulated >10-fold by cell proliferation (19-21, §),nothing is known about the events that control this increasein activity. Clearly, before the cellular functions of topoisom-erase II can be completely described, the factors that regulateits activity must be well understood.
Recently, the type I topoisomerase from Novikoff hepa-toma cells was shown to exist in vivo as a phosphoprotein(22). Moreover, when purified, the enzyme was found to besensitive to its state of phosphorylation, with higher levels ofincorporation yielding increased rates of activity in vitro (23,24). This suggests that post-translational modification mayplay a role in the regulation of topoisomerases. With this inmind, the effects of phosphorylation on the activity oftopoisomerase II have been examined.
This paper describes the interaction between topoisomer-ase II and casein kinase II from Drosophila melanogaster.While topoisomerase II showed no autophosphorylationunder the conditions used, kinase-mediated phosphorylation
of the enzyme stimulated its DNA relaxation activity. Fur-thermore, the effects of phosphorylation could be reversedby treatment with alkaline phosphatase. This study, there-fore, demonstrates that interactions between topoisomeraseII and its nucleic acid substrates can be modulated bypost-translational modification of the enzyme.
MATERIALS AND METHODSTris phosphorylated amino acid standards, and protein mo-lecular weight standards were obtained from Sigma; tri-chloroacetic acid and triethanolamine were from Fisher;alkaline phosphatase (calf intestine, enzyme immunoassaygrade) was from Boehringer Mannheim; NaDodSO4 wasfrom MCB Chemical (Norwood, OH); ATP was from P-LBiochemicals; [y-32P]ATP (2900 Ci/mmol; 1 Ci = 37 GBq)was from New England Nuclear. All other chemicals wereanalytical reagent grade. DNA topoisomerase II and caseinkinase II were purified from 6- to 18-hr-old embryos of D.melanogaster by the procedures of Shelton et al. (25) andGlover et al. (26), respectively.
Casein kinase II assays were performed by a modificationof the procedure of Glover et al. (26). Unless stated other-wise, phosphorylation reactions contained 3 nM caseinkinase II/35 nM topoisomerase II/30 ,uM [y-32P]ATP (1-1.25Ci/mmol) in a total of 30 ,ul of kinase buffer (50mM Tris HCl,pH 8.5/75 mM NaCl/10 mM MgCl2). Incubations were for 5min at 25°C. The effect of topoisomerase II concentration onthe kinetics of phosphorylation was studied over a range of20-110 nM. Reactions were stopped by trichloroacetic acidprecipitation onto Whatman GF/C filters (26). Phosphoryl-ation was quantitated by liquid scintillation using a BeckmanLS-7500 liquid scintillation counter (open window) andAmersham aqueous counting scintillant.
Unless stated otherwise, dephosphorylation reactions con-tained 35 nM topoisomerase II and 5 units of alkalinephosphatase (supplied by the manufacturer in 3 M NaCl/1 mM ZnCl2/1 mM MgCl2/1 mM triethanolamine, pH 7.6,storage buffer) in a total of 22.5 A1 of kinase buffer. Incuba-tions were at 30°C for 15 min. The final ZnCl2 concentration,which was carried along with the phosphatase in thesereactions was -33 AM. An equivalent amount of alkalinephosphatase storage buffer replaced the phosphatase incontrol reactions to compensate for the slight (<10%) inhibi-tion of topoisomerase II activity by this concentration ofZnCl2 (unpublished data).Topoisomerase II activity was assessed by the steady-state
DNA relaxation assay of Osheroff et al. (27), using 0.8 nMenzyme and 5.3 nM negatively supercoiled bacterial plasmidpBR322 DNA (28). Samples were electrophoresed on 1.0%
tTo whom reprint requests should be addressed.§Sahai, B. M. & Kowalski, D., 12th International Congress ofBiochemistry, Aug. 15-21, 1982, Perth, Western Australia, abstr.POS-003-44.
3164
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 82 (1985) 3165
agarose (MCB Chemical) gels, and the relaxation ofsupercoiled plasmid molecules was quantitated by scanningdensitometry as described (27).
Protein gel electrophoresis was carried out on 10%acrylamide gels as described by Laemmli (29). Prior toelectrophoresis, samples were dissolved in 1 vol of 250 mMTris HCl, pH 6.8/8% NaDodSO4/20% 2-mercaptoetha-nol/40% (vol/vol) glycerol/0.004% bromophenol blue, andheated at 95°C for 2 min. Gels were stained in 0.05%Coomassie brilliant blue R-250 (Bio-Rad) in 25% isopropan-ol/10% acetic acid and destained by continuous diffusion in25% isopropanol/10% acetic acid. [32P]Phosphate incorpo-ration in dried polyacrylamide gels was visualized byautoradiography, using Kodak XAR film with a DuPontLightning Plus screen.
Phosphorylated topoisomerase II samples were preparedfor partial acid hydrolysis as described by Glover et al. (26).Hydrolysis was in 6 M HCl under reduced pressure at 110°Cfor 2 hr. The hydrosylate was recovered and mixed withphosphorylated amino acid standards (P-Ser, P-Thr, P-Tyr)and analyzed by two-dimensional electrophoresis at pH 1.9and pH 3.5 on Brinkmann Polygram Cel 300 thin layercellulose plates as described by Hunter and Sefton (30).Standards were located by ninhydrin staining and[32P]phosphate was visualized by autoradiography as de-scribed above.
RESULTSPhosphorylation ofDNA Topoisomerase II by Casein Kinase
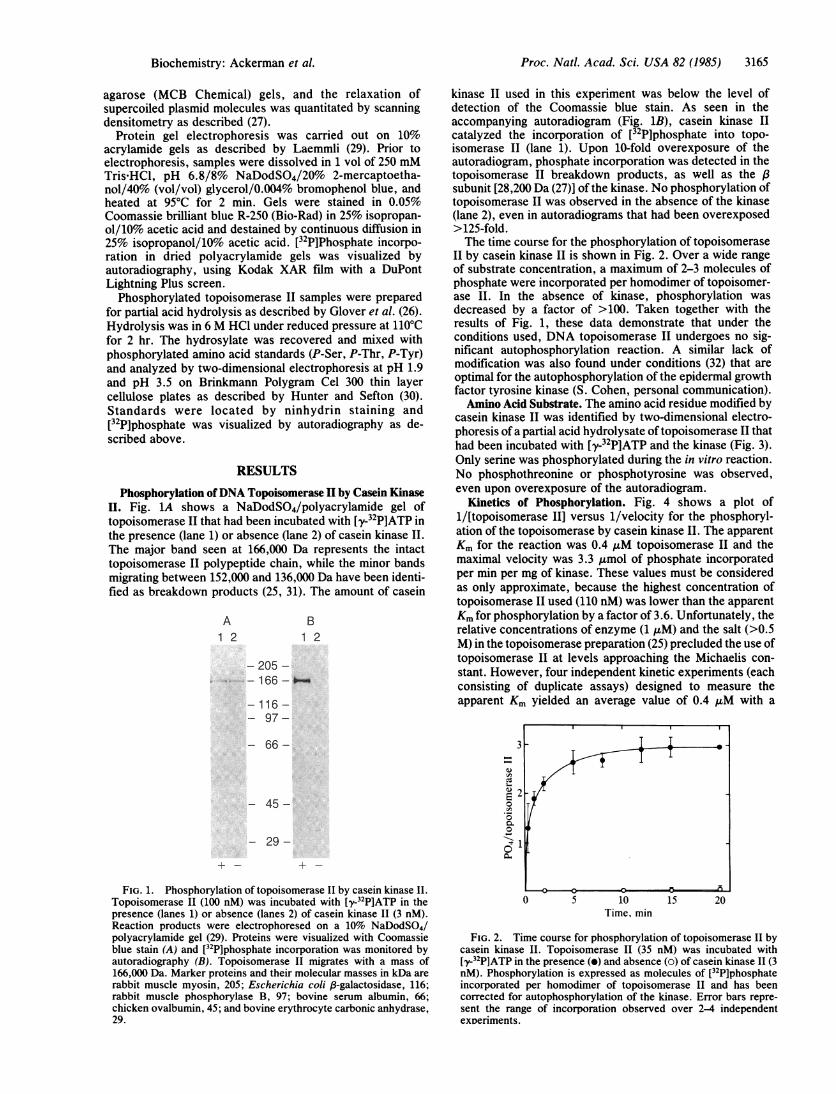
II. Fig. 1A shows a NaDodSO4/polyacrylamide gel oftopoisomerase II that had been incubated with [y-32PIATP inthe presence (lane 1) or absence (lane 2) of casein kinase II.The major band seen at 166,000 Da represents the intacttopoisomerase II polypeptide chain, while the minor bandsmigrating between 152,000 and 136,000 Da have been identi-fied as breakdown products (25, 31). The amount of casein
A1 2
B1 2
- 205 --1 66 - _--
-116 -97-
- 66-
- 45 -
- 29-
FIG. 1. Phosphorylation of topoisomerase II by casein kinase II.Topoisomerase 11 (100 nM) was incubated with [y-32P]ATP in thepresence (lanes 1) or absence (lanes 2) of casein kinase II (3 nM).Reaction products were electrophoresed on a 10% NaDodSO4/polyacrylamide gel (29). Proteins were visualized with Coomassieblue stain (A) and [32P]phosphate incorporation was monitored byautoradiography (B). Topoisomerase II migrates with a mass of166,000 Da. Marker proteins and their molecular masses in kDa arerabbit muscle myosin, 205; Escherichia coli /3-galactosidase, 116;rabbit muscle phosphorylase B, 97; bovine serum albumin, 66;chicken ovalbumin, 45; and bovine erythrocyte carbonic anhydrase,29.
kinase II used in this experiment was below the level ofdetection of the Coomassie blue stain. As seen in theaccompanying autoradiogram (Fig. 1B), casein kinase IIcatalyzed the incorporation of [32P]phosphate into topo-isomerase II (lane 1). Upon 10-fold overexposure of theautoradiogram, phosphate incorporation was detected in thetopoisomerase II breakdown products, as well as the /3subunit [28,200 Da (27)] of the kinase. No phosphorylation oftopoisomerase II was observed in the absence of the kinase(lane 2), even in autoradiograms that had been overexposed>125-fold.The time course for the phosphorylation of topoisomerase
II by casein kinase II is shown in Fig. 2. Over a wide rangeof substrate concentration, a maximum of 2-3 molecules ofphosphate were incorporated per homodimer of topoisomer-ase II. In the absence of kinase, phosphorylation wasdecreased by a factor of >100. Taken together with theresults of Fig. 1, these data demonstrate that under theconditions used, DNA topoisomerase II undergoes no sig-nificant autophosphorylation reaction. A similar lack ofmodification was also found under conditions (32) that areoptimal for the autophosphorylation of the epidermal growthfactor tyrosine kinase (S. Cohen, personal communication).Amino Acid Substrate. The amino acid residue modified by
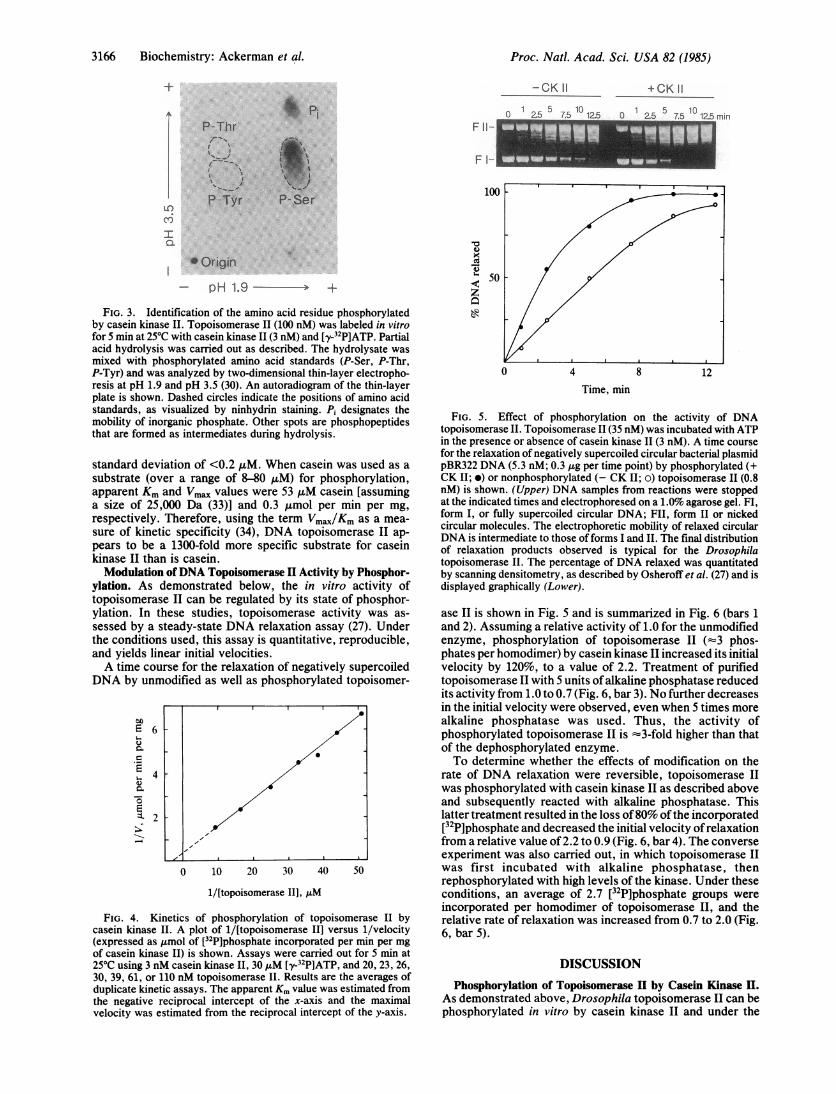
casein kinase II was identified by two-dimensional electro-phoresis of a partial acid hydrolysate of topoisomerase II thathad been incubated with [y-32P]ATP and the kinase (Fig. 3).Only serine was phosphorylated during the in vitro reaction.No phosphothreonine or phosphotyrosine was observed,even upon overexposure of the autoradiogram.
Kinetics of Phosphorylation. Fig. 4 shows a plot of1/[topoisomerase II] versus 1/velocity for the phosphoryl-ation of the topoisomerase by casein kinase II. The apparentKm for the reaction was 0.4 ,uM topoisomerase II and themaximal velocity was 3.3 ,umol of phosphate incorporatedper min per mg of kinase. These values must be consideredas only approximate, because the highest concentration oftopoisomerase II used (110 nM) was lower than the apparentKm for phosphorylation by a factor of 3.6. Unfortunately, therelative concentrations of enzyme (1 ,M) and the salt (>0.5M) in the topoisomerase preparation (25) precluded the use oftopoisomerase II at levels approaching the Michaelis con-stant. However, four independent kinetic experiments (eachconsisting of duplicate assays) designed to measure theapparent Km yielded an average value of 0.4 ,uM with a
10Time, min
FIG. 2. Time course for phosphorylation of topoisomerase II bycasein kinase II. Topoisomerase II (35 nM) was incubated with[y-32P]ATP in the presence (o) and absence (o) of casein kinase 11 (3nM). Phosphorylation is expressed as molecules of [32P]phosphateincorporated per homodimer of topoisomerase II and has beencorrected for autophosphorylation of the kinase. Error bars repre-sent the range of incorporation observed over 2-4 independentexneriments.
Biochemistry: Ackerman et al.
3166 Biochemistry: Ackerman et al.
+ -CK II1 2.5 7.5 102.5
P- Thri s*.
I
I T
P--Tyr
+CK II
O1
C -I 7 a- 10 1 ; min
F II-
F I-
10P- Ser
I
Origin- pH 1.9 -- +
FIG. 3. Identification of the amino acid residue phosphorylatedby casein kinase II. Topoisomerase II (100 nM) was labeled in vitrofor 5 min at 250C with casein kinase 11(3 nM) and [y-32P]ATP. Partialacid hydrolysis was carried out as described. The hydrolysate wasmixed with phosphorylated amino acid standards (P-Ser, P-Thr,P-Tyr) and was analyzed by two-dimensional thin-layer electropho-resis at pH 1.9 and pH 3.5 (30). An autoradiogram of the thin-layerplate is shown. Dashed circles indicate the positions of amino acidstandards, as visualized by ninhydrin staining. Pi designates themobility of inorganic phosphate. Other spots are phosphopeptidesthat are formed as intermediates during hydrolysis.
standard deviation of <0.2 1AM. When casein was used as asubstrate (over a range of 8-80 AuM) for phosphorylation,apparent Km and Vmax values were 53 AM casein [assuminga size of 25,000 Da (33)] and 0.3 umol per min per mg,respectively. Therefore, using the term Vmax/Km as a mea-sure of kinetic specificity (34), DNA topoisomerase II ap-pears to be a 1300-fold more specific substrate for caseinkinase II than is casein.Modulation ofDNA Topoisomerase II Activity by Phosphor-
ylation. As demonstrated below, the in vitro activity oftopoisomerase II can be regulated by its state of phosphor-ylation. In these studies, topoisomerase activity was as-sessed by a steady-state DNA relaxation assay (27). Underthe conditions used, this assay is quantitative, reproducible,and yields linear initial velocities.A time course for the relaxation of negatively supercoiled
DNA by unmodified as well as phosphorylated topoisomer-
E 6
._
E
& 4
a.:-21-
0 10 20 30 40 50
1/[topoisomerase II], AM
FIG. 4. Kinetics of phosphorylation of topoisomerase II bycasein kinase II. A plot of 1/[topoisomerase II] versus 1/velocity(expressed as ,umol of [32P]phosphate incorporated per min per mgof casein kinase II) is shown. Assays were carried out for 5 min at250C using 3 nM casein kinase II, 30 AM [y-32P]ATP, and 20, 23, 26,30, 39, 61, or 110 nM topoisomerase II. Results are the averages ofduplicate kinetic assays. The apparent Km value was estimated fromthe negative reciprocal intercept of the x-axis and the maximalvelocity was estimated from the reciprocal intercept of the y-axis.
0
x
50z
0 4 8 12Time, min
FIG. 5. Effect of phosphorylation on the activity of DNAtopoisomerase II. Topoisomerase II (35 nM) was incubated with ATPin the presence or absence of casein kinase II (3 nM). A time coursefor the relaxation of negatively supercoiled circular bacterial plasmidpBR322 DNA (5.3 nM; 0.3 gg per time point) by phosphorylated (+CK II; *) or nonphosphorylated (- CK II; o) topoisomerase 11 (0.8nM) is shown. (Upper) DNA samples from reactions were stoppedat the indicated times and electrophoresed on a 1.0% agarose gel. FI,form I, or fully supercoiled circular DNA; FII, form II or nickedcircular molecules. The electrophoretic mobility of relaxed circularDNA is intermediate to those offorms I and II. The final distributionof relaxation products observed is typical for the Drosophilatopoisomerase II. The percentage of DNA relaxed was quantitatedby scanning densitometry, as described by Osheroff et al. (27) and isdisplayed graphically (Lower).
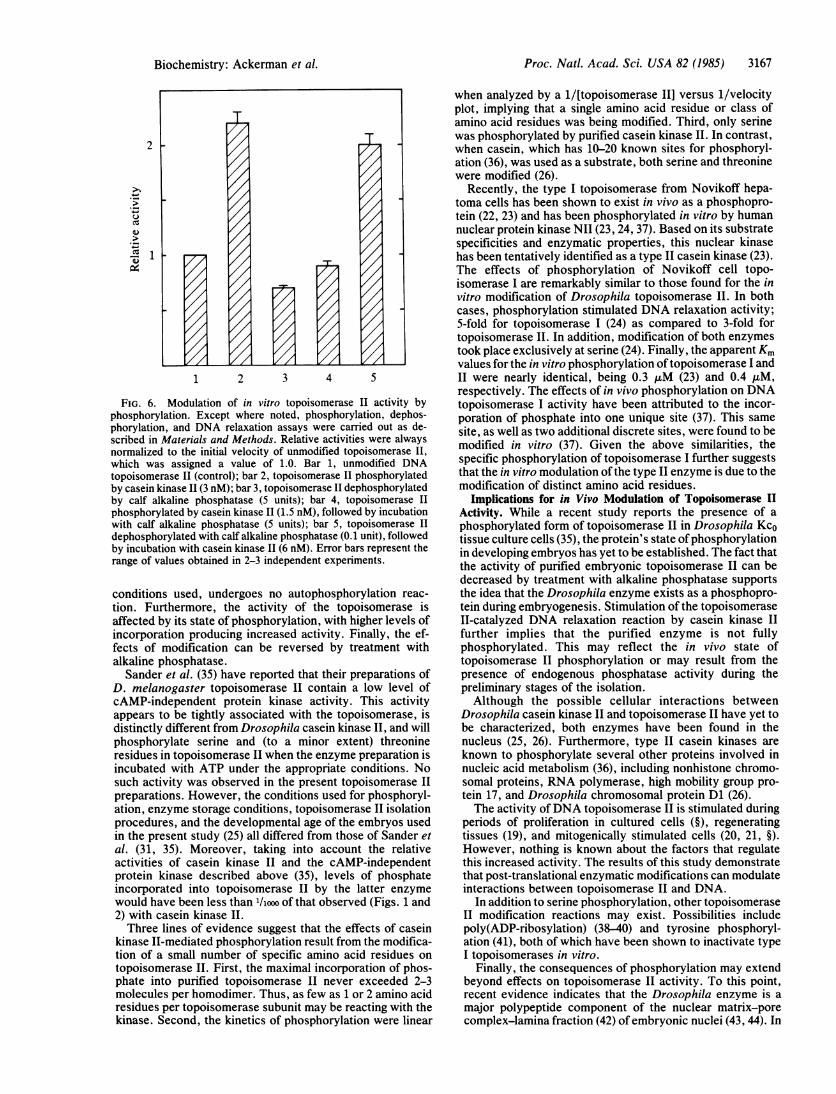
ase II is shown in Fig. 5 and is summarized in Fig. 6 (bars 1and 2). Assuming a relative activity of 1.0 for the unmodifiedenzyme, phosphorylation of topoisomerase II (-3 phos-phates per homodimer) by casein kinase II increased its initialvelocity by 120%, to a value of 2.2. Treatment of purifiedtopoisomerase II with 5 units ofalkaline phosphatase reducedits activity from 1.0 to 0.7 (Fig. 6, bar 3). No further decreasesin the initial velocity were observed, even when 5 times morealkaline phosphatase was used. Thus, the activity ofphosphorylated topoisomerase II is -3-fold higher than thatof the dephosphorylated enzyme.To determine whether the effects of modification on the
rate of DNA relaxation were reversible, topoisomerase IIwas phosphorylated with casein kinase II as described aboveand subsequently reacted with alkaline phosphatase. Thislatter treatment resulted in the loss of80% ofthe incorporated[32P]phosphate and decreased the initial velocity ofrelaxationfrom a relative value of 2.2 to 0.9 (Fig. 6, bar 4). The converseexperiment was also carried out, in which topoisomerase IIwas first incubated with alkaline phosphatase, thenrephosphorylated with high levels of the kinase. Under theseconditions, an average of 2.7 [32P]phosphate groups wereincorporated per homodimer of topoisomerase II, and therelative rate of relaxation was increased from 0.7 to 2.0 (Fig.6, bar 5).
DISCUSSIONPhosphorylation of Topoisomerase II by Casein Kinase II.
As demonstrated above, Drosophila topoisomerase II can bephosphorylated in vitro by casein kinase II and under the
Proc. Natl. Acad. Sci. USA 82 (1985)
F.ld le-i v e-u I.Q Z.Z)
f:: :: :,..
It'l
Proc. Natl. Acad. Sci. USA 82 (1985) 3167
2
1 2 3 4 5
FIG. 6. Modulation of in vitro topoisomerase II activity byphosphorylation. Except where noted, phosphorylation, dephos-phorylation, and DNA relaxation assays were carried out as de-scribed in Materials and Methods. Relative activities were alwaysnormalized to the initial velocity of unmodified topoisomerase II,
which was assigned a value of 1.0. Bar 1, unmodified DNAtopoisomerase II (control); bar 2, topoisomerase II phosphorylatedby casein kinase II (3 nM); bar 3, topoisomerase II dephosphorylatedby calf alkaline phosphatase (5 units); bar 4, topoisomerase II
phosphorylated by casein kinase II (1.5 nM), followed by incubationwith calf alkaline phosphatase (5 units); bar 5, topoisomerase IIdephosphorylated with calf alkaline phosphatase (0.1 unit), followedby incubation with casein kinase II (6 nM). Error bars represent therange of values obtained in 2-3 independent experiments.
conditions used, undergoes no autophosphorylation reac-
tion. Furthermore, the activity of the topoisomerase isaffected by its state of phosphorylation, with higher levels ofincorporation producing increased activity. Finally, the ef-fects of modification can be reversed by treatment withalkaline phosphatase.
Sander et al. (35) have reported that their preparations ofD. melanogaster topoisomerase II contain a low level ofcAMP-independent protein kinase activity. This activityappears to be tightly associated with the topoisomerase, isdistinctly different from Drosophila casein kinase II, and willphosphorylate serine and (to a minor extent) threonineresidues in topoisomerase II when the enzyme preparation isincubated with ATP under the appropriate conditions. Nosuch activity was observed in the present topoisomerase IIpreparations. However, the conditions used for phosphoryl-ation, enzyme storage conditions, topoisomerase II isolationprocedures, and the developmental age of the embryos usedin the present study (25) all differed from those of Sander etal. (31, 35). Moreover, taking into account the relativeactivities of casein kinase II and the cAMP-independentprotein kinase described above (35), levels of phosphateincorporated into topoisomerase II by the latter enzymewould have been less than 'liooo of that observed (Figs. 1 and2) with casein kinase II.
Three lines of evidence suggest that the effects of caseinkinase II-mediated phosphorylation result from the modifica-tion of a small number of specific amino acid residues on
topoisomerase II. First, the maximal incorporation of phos-phate into purified topoisomerase II never exceeded 2-3molecules per homodimer. Thus, as few as 1 or 2 amino acidresidues per topoisomerase subunit may be reacting with thekinase. Second, the kinetics of phosphorylation were linear
when analyzed by a 1/[topoisomerase II] versus 1/velocityplot, implying that a single amino acid residue or class ofamino acid residues was being modified. Third, only serinewas phosphorylated by purified casein kinase II. In contrast,when casein, which has 10-20 known sites for phosphoryl-ation (36), was used as a substrate, both serine and threoninewere modified (26).
Recently, the type I topoisomerase from Novikoff hepa-toma cells has been shown to exist in vivo as a phosphopro-tein (22, 23) and has been phosphorylated in vitro by humannuclear protein kinase NIl (23, 24, 37). Based on its substratespecificities and enzymatic properties, this nuclear kinasehas been tentatively identified as a type II casein kinase (23).The effects of phosphorylation of Novikoff cell topo-isomerase I are remarkably similar to those found for the invitro modification of Drosophila topoisomerase II. In bothcases, phosphorylation stimulated DNA relaxation activity;5-fold for topoisomerase I (24) as compared to 3-fold fortopoisomerase II. In addition, modification of both enzymestook place exclusively at serine (24). Finally, the apparent Kmvalues for the in vitro phosphorylation oftopoisomerase I andII were nearly identical, being 0.3 AuM (23) and 0.4 AuM,respectively. The effects of in vivo phosphorylation on DNAtopoisomerase I activity have been attributed to the incor-poration of phosphate into one unique site (37). This samesite, as well as two additional discrete sites, were found to bemodified in vitro (37). Given the above similarities, thespecific phosphorylation of topoisomerase I further suggeststhat the in vitro modulation of the type II enzyme is due to themodification of distinct amino acid residues.
Implications for in Vivo Modulation of Topoisomerase IIActivity. While a recent study reports the presence of aphosphorylated form of topoisomerase II in Drosophila KcOtissue culture cells (35), the protein's state ofphosphorylationin developing embryos has yet to be established. The fact thatthe activity of purified embryonic topoisomerase II can bedecreased by treatment with alkaline phosphatase supportsthe idea that the Drosophila enzyme exists as a phosphopro-tein during embryogenesis. Stimulation of the topoisomeraseII-catalyzed DNA relaxation reaction by casein kinase IIfurther implies that the purified enzyme is not fullyphosphorylated. This may reflect the in vivo state oftopoisomerase II phosphorylation or may result from thepresence of endogenous phosphatase activity during thepreliminary stages of the isolation.Although the possible cellular interactions between
Drosophila casein kinase II and topoisomerase II have yet tobe characterized, both enzymes have been found in thenucleus (25, 26). Furthermore, type II casein kinases areknown to phosphorylate several other proteins involved innucleic acid metabolism (36), including nonhistone chromo-somal proteins, RNA polymerase, high mobility group pro-tein 17, and Drosophila chromosomal protein D1 (26).The activity ofDNA topoisomerase II is stimulated during
periods of proliferation in cultured cells (§), regeneratingtissues (19), and mitogenically stimulated cells (20, 21, §).However, nothing is known about the factors that regulatethis increased activity. The results of this study demonstratethat post-translational enzymatic modifications can modulateinteractions between topoisomerase II and DNA.
In addition to serine phosphorylation, other topoisomeraseII modification reactions may exist. Possibilities includepoly(ADP-ribosylation) (38-40) and tyrosine phosphoryl-ation (41), both of which have been shown to inactivate typeI topoisomerases in vitro.
Finally, the consequences of phosphorylation may extendbeyond effects on topoisomerase II activity. To this point,recent evidence indicates that the Drosophila enzyme is amajor polypeptide component of the nuclear matrix-porecomplex-lamina fraction (42) of embryonic nuclei (43, 44). In
T
Biochemistry: Ackerman et al.

3168 Biochemistry: Ackerman et al.
mammalian cells, phosphorylation of the lamins increases 3-to 6-fold during mitotic solubilization of the nuclear lamina(45, 46). Therefore, phosphorylation may also affect theintracellular localization of type II DNA topoisomerases ineukaryotic organisms.
We are most grateful to Dr. Jai Dev Dasgupta for his assistance indetermining the amino acid residue modified by casein kinase II, toDr. David Gene Morgan for his help in running polyacrylamide gels,to Dr. Stanley Cohen for many stimulating discussions and sugges-tions, and to Susan Heaver for the preparation of this manuscript.This work was supported by National Institutes of Health GrantGM-33944, Basil O'Connor Starter Research Grant No. 5-472 fromthe March ofDimes Birth Defects Foundation, and American CancerSociety Institutional Research Grant IN-25W (to N.O.) and NationalInstitutes of Health Grant GM-33237 (to C.V.C.G.). P.A. was atrainee under U.S. Public Health Service Grant ES-07028.
1. Gellert, M. (1981) Annu. Rev. Biochem. 50, 953-960.2. Gellert, M. (1981) Enzymes 14, 345-366.3. Cozzarelli, N. R. (1980) Cell 22, 327-328.4. Wang, J. C. (1982) in Nucleases, eds. Linn, S. M. & Roberts,
R. J. (Cold Spring Harbor Laboratory, Cold Spring Harbor,NY), pp. 41-57.
5. Brown, P. 0. & Cozzarelli, N. R. (1979) Science 206,1081-1083.
6. Mizzuchi, K., Fisher, L. M., O'Dea, M. H. & Gellert, M.(1980) Proc. Natl. Acad. Sci. USA 77, 1847-1851.
7. Liu, L. F., Liu, C.-C. & Alberts, B. M. (1980) Cell 19, 697-707.8. Goto, T. & Wang, J. C. (1984) Cell 36, 1073-1080.9. DiNardo, S., Voelkel, K. & Sternglantz, R. (1984) Proc. Nail.
Acad. Sci. USA 81, 2616-2620.10. Uemura, T. & Yanagida, M. (1984) EMBO J. 3, 1737-1744.11. Mattern, M. R. & Painter, R. B. (1979) Biochim. Biophys. Acta
563, 306-312.12. Sundin, 0. & Varshavsky, A. (1981) Cell 25, 659-669.13. Nishio, A. & Uyeki, E. M. (1982) Biochem. Biophys. Res.
Commun. 106, 1448-1455.14. Speck, W. T., Rosenkranz, P. G. & Rosenkranz, H. S. (1982)
Mutat. Res. 104, 125-130.15. Englund, P. T., Hajduk, S. L. & Marini, J. C. (1982) Annu.
Rev. Biochem. 51, 695-726.16. Castora, F. J., Vissering, F. F. & Simpson, M. V. (1983)
Biochim. Biophys. Acta 740, 417-427.17. Shlomai, J. & Zadok, A. (1983) Nucleic Acids Res. 11,
4019-4034.18. Mattern, M. R., Paone, R. F. & Day, R. S. (1982) Biochim.
Biophys. Acta 697, 6-13.19. Duguet, M., Lavenot, C., Harper, F., Mirambeau, G. &
DeRecondo, A. M. (1983) Nucleic Acids Res. 11, 1059-1075.20. Miskimins, R., Miskimins, W. K., Bernstein, H. & Shimizu,
N. (1983) Exp. Cell Res. 146, 53-62.
21. Taudou, G., Mirambeau, G., Lavenot, C., der Garabedian, A.,Vermeersch, J. & Duguet, M. (1984) FEBS Lett. 176, 431-435.
22. Durban, E., Roll, D., Beckner, G. & Busch, H. (1981) CancerRes. 41, 537-545.
23. Mills, J. S., Busch, H. & Durban, E. (1982) Biochem. Biophys.Res. Commun. 109, 1222-1227.
24. Durban, E., Mills, J. S., Roll, D. & Busch, H. (1983) Biochem.Biophys. Res. Commun. 111, 897-905.
25. Shelton, E. R., Osheroff, N. & Brutlag, D. L. (1983) J. Biol.Chem. 258, 9530-9535.
26. Glover, C. V. C., Shelton, E. R. & Brutlag, D. L. (1983) J.Biol. Chem. 258, 3258-3265.
27. Osheroff, N., Shelton, E. R. & Brutlag, D. L. (1983) J. Biol.Chem. 258, 9536-9543.
28. Bolivar, F., Rodriguez, R., Greene, P. J., Betlach, M.,Heyneker, H. L., Boyer, H. W., Crosa, J. & Falkow, S. (1977)Gene 2, 95-113.
29. Laemmli, U. K. (1970) Nature (London) 227, 680-685.30. Hunter, T. & Sefton, B. M. (1980) Proc. Natl. Acad. Sci. USA
77, 1311-1315.31. Sander, M. & Hsieh, T.-S. (1983) J. Biol. Chem. 258,
8421-8424.32. Cohen, S., Carpenter, G. & King, L., Jr. (1980) J. Biol. Chem.
255, 4834-4842.33. Jenness, R. (1970) in Milk Proteins-Chemistry and Molecular
Biology, ed. McKenzie, H. A. (Academic, New York), pp.17-43.
34. Bender, M. L. & Kezdy, F. J. (1965) Annu. Rev. Biochem. 34,49-76.
35. Sander, M., Nolan, J. M. & Hsieh, T.-S. (1984) Proc. Natl.Acad. Sci. USA 81, 6938-6942.
36. Hathaway, G. M. & Traugh, J. A. (1982) Curr. Top. Cell.Regul. 21, 101-127.
37. Durban, E., Goodenough, M. & Busch, H. (1984) Fed. Proc.Fed. Am. Soc. Exp. Biol. 43, 1542 (abstr.).
38. Jongstra-Bilen, J., Ittel, M.-E., Nidergan$, C., Vosberg, H.-P.& Mandel, P. (1983) Eur. J. Biochem. 136, 391-3%9.
39. Ferro, A. M., Higgins, N. P. & Olivera, B. M. (1983) J. Biol.Chem. 258, 6000-6003.
40. Ferro, A. M. & Olivera, B. M. (1984) J. Biol. Chem. 259,547-554.
41. Tse-Dinh, Y.-C., Wong, T. W. & Goldberg, A. R. (1984)Nature (London) 312, 785-786.
42. Fisher, P. A., Berrios, M. & Blobel, G. (1982) J. Cell Biol. 92,674-686.
43. Berrios, M., Osheroff, N. & Fisher, P. (1984) J. Cell Biol. 99,128a (abstr.).
44. Halligan, B. D., Small, D., Vogelstein, B., Hsieh, T.-S. & Liu,L. F. (1984) J. Cell Biol. 99, 128a (abstr.).
45. Gerace, L. & Blobel, G. (1980) Cell 19, 277-287.46. Gerace, L. & Blobel, G. (1981) Cold Spring Harbor Symp.
Quant. Biol. 46, 967-978.
Proc. Natl. Acad. Sci. USA 82 (1985)





























