Embed Size (px)
Citation preview

Eur. J. Biochem. 86, 301 -310 (1978)
Phosphorylation in vivo of Proteins Associated with Heterogenous Nuclear RNA in HeLa Cell Nuclei
Jean-Marie BLANCHARD, Claude BRUNEL, and Philippe JEANTEUR
Laboratoire de Biologie Moleculaire, Universite des Sciences et Techniques du Languedoc, Montpellier, and Laboratoire de Biochimie, Centre Paul Lamarque, Hopital St-Eloi, Montpellier
(Received July 27/December 7, 1977)
As a step towards understanding the role of nuclear ribonucleoprotein particles (hnRNA . proteins) in the nucleocytoplasmic transfer of heterogenous nuclear RNA sequences destined to become the cytoplasmic messenger RNA, the phosphorylation in vivo of the protein moiety of these hnRNA . proteins from HeLa cells has been studied.
After exposure of HeLa cells to [32P]orthophosphate in vivo, purified hnRNA . proteins were found to contain radioactivity covalently bound to proteins in the form of serine and threonine phosphate esters. The association of these phosphoproteins with hnRNA was demonstrated by sedimentation analysis in sucrose gradients, isopycnic banding in cesium chloride density gradients and chromatography on oligo(dT)-cellulose.
The pattern of radioactive phosphorylated species has been analysed by electrophoresis on sodium dodecylsulphate/polyacrylaniide slab gels followed by autoradiography. Although some hot- trichloroacetic-acid-resistant "P radioactivity was present throughout the gel, the highest amount of label was found associated with four discrete bands of molecular weights 28000, 30000, 37000 and 52000.
Incorporation of 32P into hnRNA . proteins could be detected after labeling periods as short as 15 min, the same species being labeled after 1 and 24-h labeling times. Although all the above species appear to be present in hnRNA . proteins of different size classes, as resolved by sedimenta- tion through sucrose gradients, their relative proportion is not reproducibly constant.
Exposure of hnRNA . proteins to conditions of increasing ionic strength led to the ready disso- ciation of the 28000-M, species at variance with other labeled species which appeared to be more firmly bound to hnRNA than the bulk of proteins. Upon treatment with pancreatic and TI ribonucleases, the 28 000-M, species was also preferentially released while the 37000-M, one was the main phosphorylated species remaining associated with core structures containing an RNA component resistant to nucleases which is now under further characterization.
The association of hnRNA with proteins in the form of hnRNA . proteins is now well established in a wide variety of eucaryotic systems. Working with HeLa cells, we have shown that the protein moiety of hnRNA . protein is made up of a complex array of polypeptides when analysed by polyacrylamide gel electrophoresis either in one dimension in the pres- ence of sodium dodecylsulphate [I j or in two dimen- sions [2j. This heterogeneity, already observed in rat liver by Niessing and Sekeris [3] and rat brain by Matringe and Jacob [4], was confirmed in HeLa cells by Pederson [ 5 ] .
In a search for enzymatic activities which might be in some way involved in the nucleocytoplasmic
Abbreviations. hnRNA, heterogenous nuclear RNA ; hnRNA . proteins, heterogenous nuclear ribonucleoprotein particles.
transfer of these hnRNA sequences destined to be- come cytoplasmic mRNA, we have become interested in phosphorylation reactions at the level of hnRNA . proteins, the more so as phosphorylated proteins have been found in nearly all classes of nuclear proteins [6- lo]. In this context we have previously shown that hnRNA . proteins, extracted from HeLa nuclei either by the technique of Samarina [ I l l or by sonication, contained and endogenous protein kinase activity [12,13].
Assuming that phosphorylation of a specific pro- tein would affect its function, we have been interested in determining which polypeptides could be subjected to such a regulation. Should the endogenous phos- phorylation which we have observed in isolated hnRNA . proteins [12,13j reflect a specific spatial arrangement of the kinase and its substrates within

302 Phosphorylation of Proteins from hnRNA . Proteins
the particles, it would be expected to mimic the pattern of phosphorylated species observed in hnRNA . pro- teins labeled in vivo with 32P. A detailed analysis of protein phosphorylation in hnRNA . proteins was therefore undertaken under conditions in vivo. This paper reports the results of such a study, a preliminary account of which has been already published [14].
MATERIALS AND METHODS
Materials
All materials were the same as those described in a preceding paper [13].
Histones were prepared from HeLa cells according to the procedure described by Bhorjee and Pederson [151.
Cell Growth and Labeling Conditions
Procedures for growth in suspension culture, collection and washing of HeLa cells (S3 strain) have been described previously [ 11. Washed cell pellets were always used immediately.
Standard labeling conditions were as follows : for short labeling times, cells were centrifuged and re- suspended to a concentration of 2 x lo6 cells/ml in medium containing dialysed calf serum, incubated for 90 min then exposed to 100 pCi/ml H332P04 for the indicated time. In the case of long labeling time (24 h) cells were resuspended to a concentration of 3 x lo5 cells/ml in a medium containing 0.01 mM phosphate and exposed to 10 pCi/ml H332P04. Label- ing conditions with [3H]uridine in the presence or in absence of actinomycin D have been described prev- iously [13].
Preparation of hnRNA . Proteins
hnRNA . proteins were purified as described on a preceding paper [13] using the sonication procedure of Kisch and Pederson [16]. In some cases the nuclear extract, in which the final centrifugation step through a triple sucrose cushion had been omitted, was used as such.
Characterization of Phosphoamino Acids
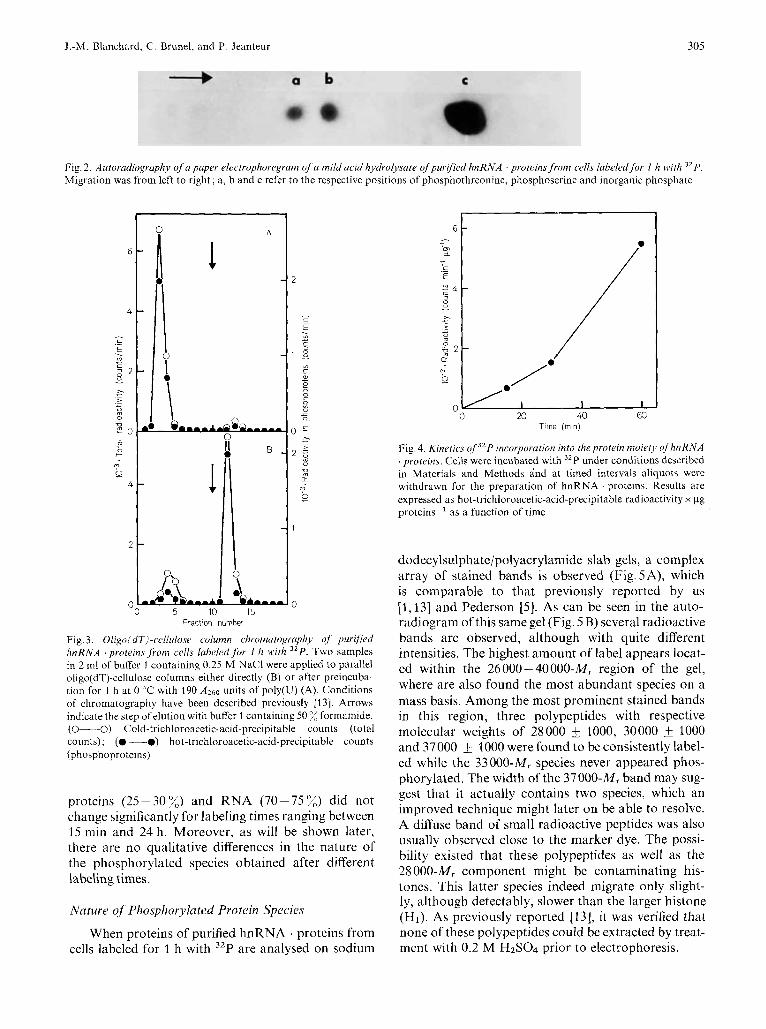
400 yg 32P-labeled hnRNA . proteins (expressed as protein) were exposed for 20 min to boiling 10% trichloroacetic acid in order to hydrolyse labeled nucleic acids. After centrifugation the pellet was wash- ed with 10% trichloroacetic acid, hydrolysed in 3 M HCl and the hydrolysate subjected to high-voltage paper electrophoresis as previously described [I31 to characterize phosphoserine and phosphothreonine.
Sucrose Density Gradient Analysis
Samples of nuclear extract or purified hnRNA . proteins were adjusted to about 10% sucrose (w/v) by dilution with buffer 1 (10 mM Tris-HC1 pH 7.4, (25 "C); 10 mM NaCl; 1.5 mM MgC12). 0.4-ml ali- quots were then layered on top of a 12-ml 15-30% (w/v) sucrose gradient in buffer 1 and centrifuged at 40000 rev./min in the SW41 rotor of the Spinco L5-75 ultracentrifuge for the time indicated in the legends. 0.4-ml fractions were collected from the bottom of the tubes by means of a peristaltic pump and two separate 5O-yl aliquots of each fraction were precipitated with trichloroacetic acid and analysed for total and phosphoprotein 32P counts as indicated below.
Oligo (dT) -cellulose Column Chromatography
32P-labeled hnRNA . proteins were prepared by the standard procedure except that the three sucrose cushions were in binding buffer (buffer 1 containing 0.25 M NaCI). 5-ml oligo(dT)cellulose columns (0.9 x 8 cm) equilibrated with the same buffer were loaded with 2-ml samples containing 6 x lo5 counts/min, washed with 15 ml buffer and eluted with 50 % formamide in regular buffer 1. Flow rate was 10 ml/h and l-ml fractions were collected. 50-$ aliquots of each fraction were used for determination of precip- itable 32P radioactivity.
Sodium Dodecylsulphate/Polyacrylamide Gel Electrophoresis
This was performed as previously described [13]. 60 pg hnRNA . proteins were routinely used in all experiments. In a few instances RNA was removed prior to electrophoresis either by the urea/LiCl method [17] or by chloroform/isoamyl alcohol ex- traction [18].
Isopycnic Banding of hnRNA ' Proteins in CsCl Density Gradients
The samples to be analysed were fixed by the addi- tion of glutaraldehyde according to Baltimore and Huang [19], then included in a 5-ml preformed CsCl gradient (1.2- 1.6 g/cm3) containing 0.5 % Brij 35 and centrifuged for 18 h at 35000 rev./min in the IEC rotor SB-283 at 4 "C. Gradients were collected into 4-drop fractions by puncturing the bottom of the tubes and the density of every third fraction was deter- mined by refractometry. Each fraction was then assay- ed for 32P radioactivity insoluble in cold or hot tri- chloroacetic acid as described below.

J.-M. Blanchard, C. Brunel, and P. Jeanteur 303
Miscellaneous
Protein concentrations were determined according to Linn and Lehman [20] or by the amido black method of Kuno and Kihara [21] both using bovine serum albumin as a standard. Radioactive samples were precipitated by 10 % trichloroacetic acid in the pres- ence of carrier bovine serum albumin for 10min at 0 "C (cold trichloroacetic acid) for determination of total 32P radioactivity, or eventually treated for 20 min at 90 "C (hot trichloroacetic acid) for phosphoproteins determination. Precipitates were collected on What- man GFjC glass filters and counted in an Intertech- nique SL 30 scintillation spectrometer using 5 ml of a scintillation fluid containing 5 g PPO and 0.3 g dimethyl POPOPjl toluene.
RESULTS
Presence o j Phosphoryluted Proteins in hnRNA . Proteins
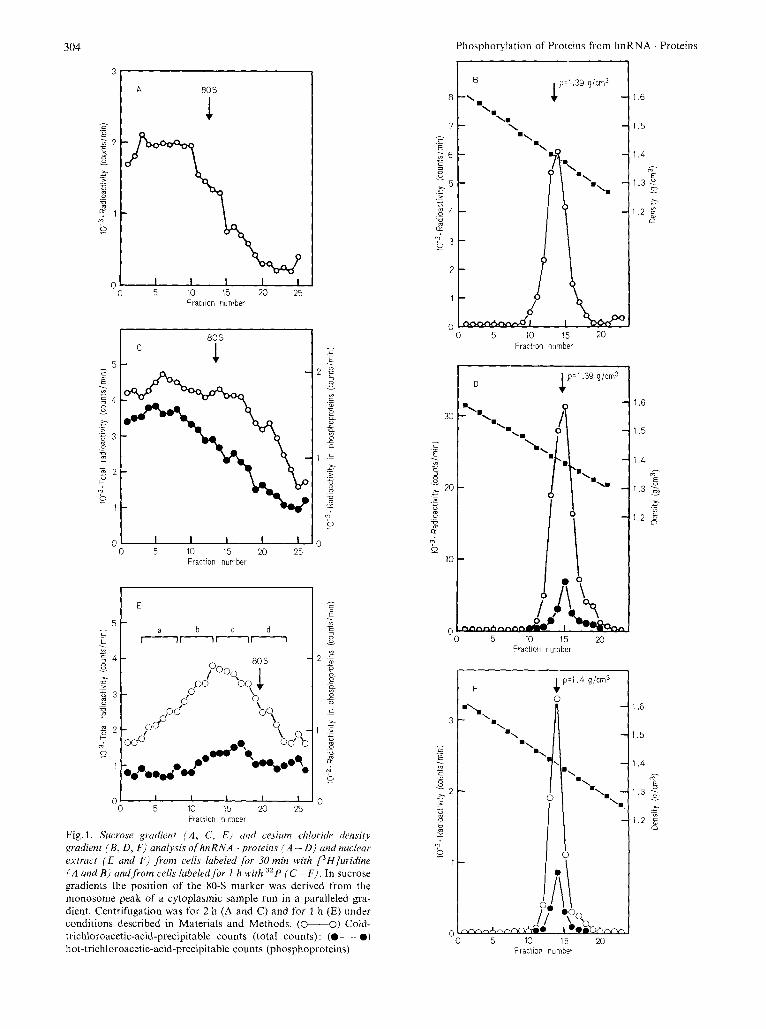
Typical patterns of sedimentation and buoyant density of purified hnRNA . proteins from cells labeled for 30 min with [3H]uridine in the absence of actino- mycin are presented in Fig. 1 A and B respectively. As previously observed [ l , 5,131, particles are hetero- genous in size from 30 S to above 200 S, and homo- genous in buoyant density (1.39 g/cm3). The same profiles (not shown) are observed in the presence of low doses (0.04 pgjml) of actinomycin D to prevent labeling of rRNA precursors [22].
When hnRNA . proteins were purified from cells labeled for 1 h with 32P in the absence of actinomycin D, respectively 95 - 98 % and 20- 25 7; of cold-acid- insoluble 32P radioactivity was resistant to further treatment with either a chloroformjmethanol mixture (to extract phospholipids) [23] of hot trichloroacetic acid indicating the presence of phosphoproteins in addition to labeled nucleic acids. Over 98% of this labeled material was sensitive to alkaline hydrolysis (0.5 M NaOH for 16 h at 37 "C), excluding the pres- ence of significant amounts of labeled DNA. This result confirms the long-term labeling experiment with [3 Hlthymidine reported in the preceding paper [ 131. Profiles of cold-acid-precipitable 32P radioactivity were similar to those obtained with E3H]uridine- labeled material both in sedimentation (Fig. 1 C) and buoyant density (Fig. 1 D). The distribution of phos- phoproteins (hot-trichloroacetic-acid-resistant radio- activity) follows the same patterns under both experi- mental conditions, suggesting that they are associated with hnRNA. Radioactive phosphoserine and phos- phothreonine were characterized by high-voltage pa- per electrophoresis of a mild acid hydrolysate (Fig. 2). A crude nuclear extract not purified through the three sucrose cushions but freed of nucleoli and most of
the chromatin by centrifugation, was also analysed so as to encompass soluble proteins. The majority (at least 90 %) of phosphoproteins comigrate with la- beled RNA in structures having sedimentation coeffi- cients larger than 30 S (Fig. 1 E). The relative amount of label in phosphoproteins as compared to RNA seems to increase with the size of the particles. However, as will be shown later (see below and Fig. 6), the same labeled species are found throughout the gradient. Around 10 "/, of the hot-trichloroacetic- acid resistant radioactivity behaved as soluble phos- phoproteins and did not enter the gradient. As for purified hnRNA . proteins, all the radioactivity of this nuclear extract banded in CsCl within a density range characteristic of hnRNA . proteins (1.38 - 1.40 g/cm3) (Fig. 1 F). The 10 o/, soluble phosphoproteins observed in sucrose gradient and not in CsCl gradient could therefore be due to some degradation occurring during centrifugation of particles untreated with formalde- hyde. This point was not further investigated. The ab- sence of radioactive material in the heavier region ex- cludes a significant contamination by preribosomes which contain, for this labeling time (1 h) in the absence of actinomycin D, comparable amounts of label in rRNA precursors to those in hnRNA [24]. The same observation with a 24-h-labeled nuclear extract (not shown) eliminates the presences of cytoplasmic ribo- somes. For all further experiments, hnRNA . proteins were purified through three sucrose cushions as de- scribed in Materials and Methods in order to get rid of soluble proteins and possibly remaining chromatin debris.
When purified "P-labeled hnRNA . proteins are chromatographed on oligo(dT)-cellulose in buffer 1 containing 0.25 M NaCI, between 80% and 90% of the radioactive material was retained by the column and eluted from it with 50% formamide in buffer 1. The bound fraction still contained 80-90% of the phosphorylated proteins as can be shown in Fig. 3 B. This high level of retention (80 - 90 %) is discrepant with the reported proportion of polyadenylated hnRNA molecules [1,26]. This point, already noted by other workers [25] as well as ourselves 1131 has been already discussed in a preceding paper [13]. The involvement of the poly(A) sequence in binding hnRNA . proteins to oligo(dT)-cellulose is therefore not clear. However, as already noted [13], when hnRNA proteins are preincubated with an excess of poly(U) before being applied to oligo(dT)-cellulose, essentially all the radioactive material is recovered in the flow-through (Fig. 3A).
Incorporation of 32P into the protein moiety of hnRNA . proteins is very rapid. The kinetic experi- ment reported in Fig. 4 shows that a significant amount of label is recovered as phosphoproteins for labeling times as short as 15 min (and probably even shorter). The percentage of radioactivity incorporated into
304
80 s l A 1
Fraction number
0 0 5 10 15 20 25
Fraction numbet
I I
b C d nnnn
I I I I ' J o 0 5 10 15 20 25
Fraction number
Fig. 1. Sucrose gi-adieni ( A , C, E ) cind cesium chloridi, densitj gradient (B, D , F ) analysis of hnRNA 'proteins ( A - 0) andnuclear extruct ( E und F ) from cells labeled for 30 min with L3Hjuridine ( A and B) andfrom cells lubeledfor I h with 3zP ( C - F ) . In sucrose gradients the position of the 80-S marker was derived from the monosome peak of a cytoplasmic sample run in a paralleled gra- dient. Centrifugation was for 2 h (A and C) and for 1 h (E) under conditions described in Materials and Methods. (0-0) Cold- trichloroacetic-acid-precipitable counts (total counts): (.--.) hot-trichloroacetic-acid-precipitable counts (phosphoproteins)
D
0 5 10 15 M Fraction number
F
0 0 5 10 15 20
Fraction number
1.6
1.5
1.4 - D
5 1.3 3
1.2 g > y1
- .-
1 .6
1.5
1.4 - 0 E
1.3 5 - > I -
1.2 2 d
J.-M. Blanchard, C. Brunel, and P. Jednteur 305
Fig. 2. Autorudiogruphy qf apuper electrophoregram of a mildacid hydrolysare ofpurified hnRNA proteins from cells labeledfiw I h with 32P. Migration was from left to right; a, h and c refer to the respective positions of phosphothreonine, phosphoserine and inorganic phosphate
2
- c .- E . - m
c 2
18 Y) c a,
a r
- - e
a a
0:
2 ; - - 0 n -
I9 ? 0
1
0
Fraction number
Fig. 3. Oligo(dT)-ccllulose column chromulogruph.v o j purified hnRNA . proteins ,from cells labeled for I h with 32P. Two samples in 2 ml o f buffer 1 containing 0.25 M NaCl were applied to parallel oligo(dT)-cellulose columns either directly (B) or after preincuha- tion for 1 h at 0 'C with 190 Az60 units of poly(U) (A). Conditions of chromatography have been described previously [I 31. Arrows indicate the step of elution with buffer 1 containing 50 formamide. (0--0) Cold-trichloroacetic-acid-precipitahle counts (total counts); (0 --0) hot-trichloroacetic-acid-precipitable counts (phosphoproteins)
proteins (25-30%) and RNA (70-75%) did not change significantly for labeling times ranging between 15 min and 24 h. Moreover, as will be shown later, there are no qualitative differences in the nature of the phosphorylated species obtained after different labeling times.
Nature of Phosphorylated Protein Species
When proteins of purified hnRNA . proteins from cells labeled for 1 h with 32P are analysed on sodium
Fig. 4. Kinetics o f j L P incorporation into theprotein moiety q fhnRNA . proteins. Cells were incubated with 32P under conditions described in Materials and Methods i nd at timed intervals aliquots were withdrawn for the preparation of hnRNA . proteins. Results are expressed as hot-trichloroacetic-acid-precipitahle radioactivity x pg proteins-' as a function of time
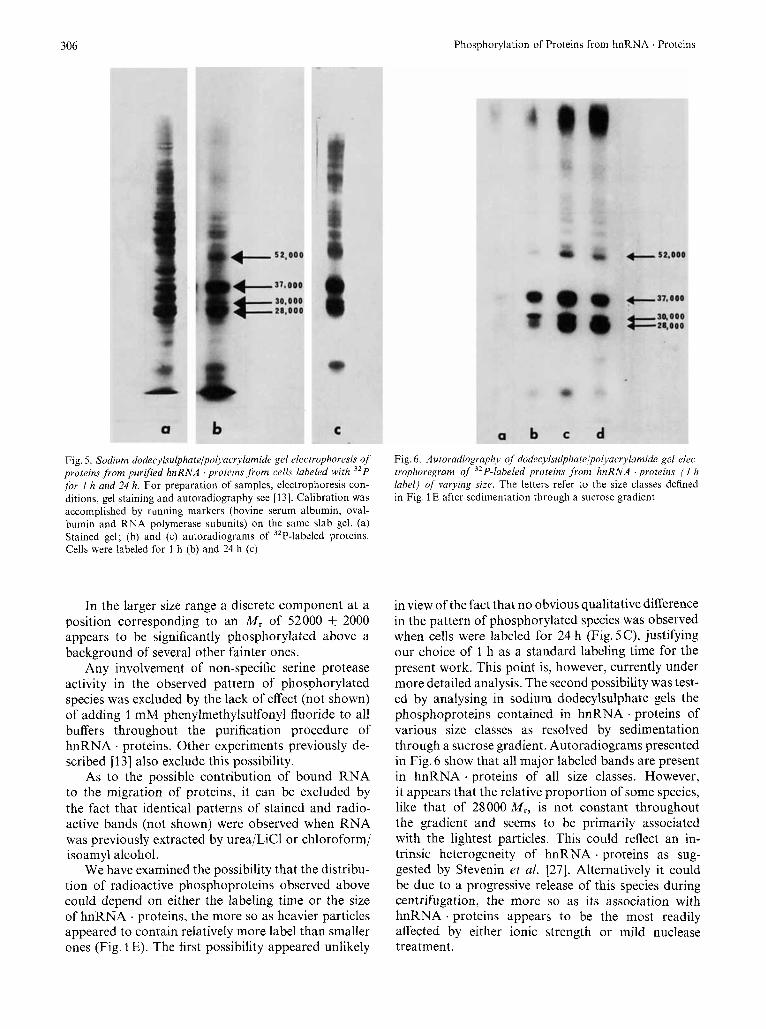
dodecylsulphate/polyacrylamide slab gels, a complex array of stained bands is observed (Fig. 5A), which is comparable to that previously reported by us [ I , 131 and Pederson [5] . As can be seen in the auto- radiogram of this same gel (Fig. 5 B) several radioactive bands are observed, although with quite different intensities. The highest amount of label appears locat- ed within the 26000 -40000-MI region of the gel, where are also found the most abundant species on a mass basis. Among the most prominent stained bands in this region, three polypeptides with respective molecular weights of 28000 +_ 1000, 30000 & 1000 and 37000 t- 1000 were found to be consistently label- ed while the 33000-MI species never appeared phos- phorylated. The width of the 37000-M, band may sug- gest that it actually contains two species, which an improved technique might later on be able to resolve. A diffuse band of small radioactive peptides was also usually observed close to the marker dye. The possi- bility existed that these polypeptides as well as the 28 000-M, component might be contaminating his- tones. This latter species indeed migrate only slight- ly, although detectably, slower than the larger histone (HI). As previously reported [13], it was verified that none of these polypeptides could be extracted by treat- ment with 0.2 M H2S04 prior to electrophoresis.
306 Phosphorylation of Proteins from hnRNA . Proteins
Fig. 5. Sodium dodecylsulphateli~olyacrylamide gel electrophoresis of proteins from purified hnRNA proteins .from cells labeled with 32P for 1 h and 24 h. For preparation of samples, electrophoresis con- ditions, gel staining and autoradiography see [13]. Calibration was accomplished by running markers (bovine serum albumin, oval- bumin and RNA polymerase subunits) on the same slab gel. (a) Stained gel; (b) and (c) autoradiograms of 32P-labeled proteins. Cells were labeled for 3 h (b) and 24 h (c)
In the larger size range a discrete component at a position corresponding to an M , of 52000 & 2000 appears to be significantly phosphorylated above a background of several other fainter ones.
Any involvement of non-specific serine protease activity in the observed pattern of phosphorylated species was excluded by the lack of effect (not shown) of adding 1 mM phenylmethylsulfonyl fluoride to all buffers throughout the purification procedure of hnRNA . proteins. Other experiments previously de- scribed I131 also exclude this possibility.
As to the possible contribution of bound RNA to the migration of proteins, it can be excluded by the fact that identical patterns of stained and radio- active bands (not shown) were observed when RNA was previously extracted by urea/LiCl or chloroform/ isoamyl alcohol.
We have examined the possibility that the distribu- tion of radioactive phosphoproteins observed above could depend on either the labeling time or the size of hnRNA . proteins, the more so as heavier particles appeared to contain relatively more label than smaller ones (Fig. 1 E). The first possibility appeared unlikely
Fig. 6 . Autoradiography of dodecylsulphatelpolyacrylamide gel elec- trophoregram of 32P-labeled proteins from hnRNA ‘proteins ( I h label) of varying size. The letters refer to the size classes defined in Fig. 1 E after sedimentation through a sucrose gradient
in view of the fact that no obvious qualitative difference in the pattern of phosphorylated species was observed when cells were labeled for 24 h (Fig. 5C), justifying our choice of 1 h as a standard labeling time for the present work. This point is, however, currently under more detailed analysis. The second possibility was test- ed by analysing in sodium dodecylsulphate gels the phosphoproteins contained in hnRNA . proteins of various size classes as resolved by sedimentation through a sucrose gradient. Autoradiograms presented in Fig. 6 show that all major labeled bands are present in hnRNA . proteins of all size classes. However, it appears that the relative proportion of some species, like that of 28000 M,, is not constant throughout the gradient and seems to be primarily associated with the lightest particles. This could reflect an in- trinsic heterogeneity of hnRNA . proteins as sug- gested by Stevenin et al. [27]. Alternatively it could be due to a progressive release of this species during centrifugation, the more so as its association with hnRNA . proteins appears to be the most readily affected by either ionic strength or mild nuclease treatment.
.I.-M. Blanchard. C. Brunel. and P. Jeanteur 307
r c 1 I 2 3
1
1 B
- I
1 D
-0 10 2 0 0 10 20 Fraction number
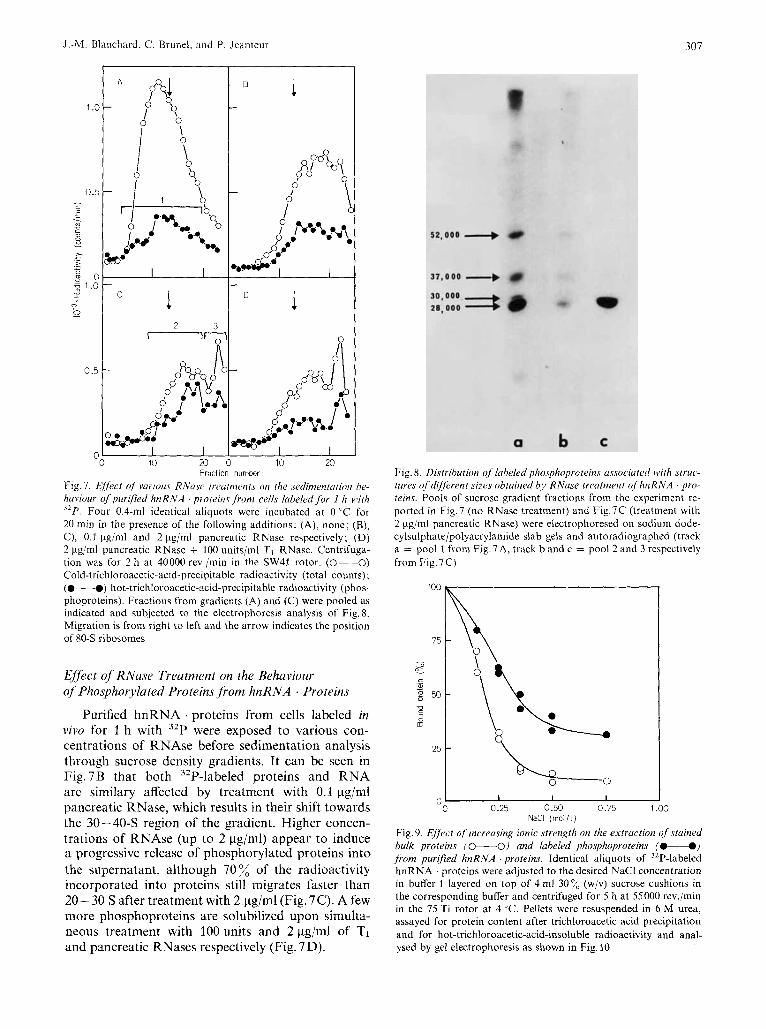
Fig. 7. Eiyect of vurious RNasc treatnzents on the .sedimmtariotr be- haviour of purified hnRNA . proteins from cells labeled fbr 1 h with 32P. Four 0.4-ml identical aliquots were incubated at 0 "C for 20 min in the presence of the following additions: (A), none; (B), C), 0.1 Fg/ml and 2 Fg/ml pancreatic RNase respectively; (D) 2 pg/ml pancreatic RNase + 100 units/ml TI RNase. Centrifuga- tion was for 2 h at 40000 rev./min in the SW41 rotor. (0--0) Cold-trichloroacetic-acid-precipitable radioactivity (total counts); (@-@) hot-trichloroacetic-acid-precipi~able radioactivity (phos- phoproteins). Fractions from gradients (A) and (C) were pooled as indicated and subjected to the electrophoresis analysis of Fig. 8. Migration is from right to left and the arrow indicates the position of 80-S ribosomes
Effect of RNuse Treatment on the Behaviour of Phosphorylated Proteins from hnRNA . Proteins
Purified hnRNA . proteins from cells labeled in vivo for 1 h with 32P were exposed to various con- centrations of RNAse before sedimentation analysis through sucrose density gradients. It can be seen in Fig.7B that both 32P-labeled proteins and RNA are similary affected by treatment with 0.1 pg/ml pancreatic RNase, which results in their shift towards the 30-40-S region of the gradient. Higher concen- trations of RNAse (up to 2 pg/ml) appear to induce a progressive release of phosphorylated proteins into the supernatant, although 70 % of the radioactivity incorporated into proteins still migrates faster than 20- 30 S after treatment with 2 pg/ml (Fig. 7C). A few more phosphoproteins are solubilized upon simulta- neous treatment with 100units and 2pg/ml of TI and pancreatic RNases respectively (Fig. 7 D).
Fig. 8. Distribution of labeled phosphoproteins ussociared with struc- tures of different .sizes obtained by RNuse treutrnent o fhnRNA pro- teins. Pools of sucrose gradient fractions from the experiment re- ported in Fig.7 (no RNase treatment) and Fig.7C (treatment with 2 pg/ml pancreatic RNase) were electrophoresed on sodium dode- cylsulphate/polyacrylamide slab gels and autoradiographed (track a = pool 1 from Fig.7A, track band c = pool 2 and 3 respectively from Fig. 7C)
100
75
I
c 6 - g 50
2 CE
25
n 1 I I Y
0 0.25 0.50 0.75 1 .oo NaCl ( rno l i l )
Fig. 9. E f j k t of increasing ionic strength on the extraction of stained bulk proteins (0-0) and labeled phosphoproteins (@d) from purified hnRNA . proteins. Identical aliquots of 32P-labeled hnRNA . proteins were adjusted to the desired NaCl concentration in buffer 1 layered on top of 4 ml 30 % (w/v) sucrose cushions in the corresponding buffer and centrifuged for 5 h at 55000 rev./min in the 75 Ti rotor at 4 "C. Pellets were resuspended in 6 M urea, assayed for protein content after trichloroacetic acid precipitation and for hot-trichloroacetic-acid-insoluble radioactivity and anal- ysed by gel electrophoresis as shown in Fig. I0
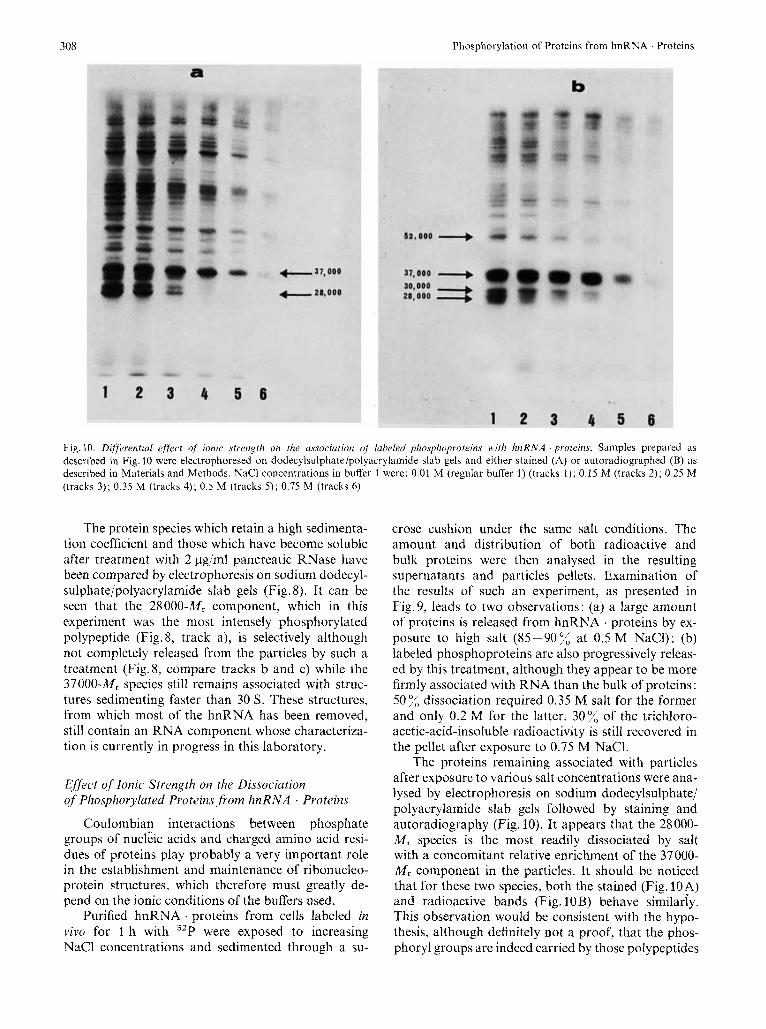
308 Phosphorylation of Proteins from hnRNA . Proteins
Fig. 10. Differential effrcl of ionic strength on the u.ssociutioiz uf labeled phosphoprotein.~ 1vith hnRNA . proteins. Samples prepared as described in Fig. 10 were electrophoresed on dodecylsulphate/polyacrylamide slab gels and either stained (A) or autoradiographed (B) as described in Materials and Methods. NaCl concentrations in buffer 1 were: 0.01 M (regular buffer 1) (tracks 1 ) ; 0.15 M (tracks 2); 0.25 M (tracks 3); 0.35 M (tracks 4); 0.5 M (tracks 5); 0.75 M (tracks 6)
The protein species which retain a high sedimenta- tion coefficient and those which have become soluble after treatment with 2 pg/ml pancreatic RNase have been compared by electrophoresis on sodium dodecyl- sulphate/polyacrylamide slab gels (Fig. 8). It can be seen that the 28000-M, component, which in this experiment was the most intensely phosphorylated polypeptide (Fig. 8, track a), is selectively although not completely released from the particles by such a treatment (Fig. 8, compare tracks b and c) while the 37000-M, species still remains associated with struc- tures sedinienting faster than 30 S. These structures, from which most of the hnRNA has been removed, still contain an RNA component whose characteriza- tion is currently in progress in this laboratory.
Effect of Ionic Strength on the Dissociation of Phosphoryluted Proteins.from hnRNA . Proteins
Coulombian interactions between phosphate groups of nucleic acids and charged amino acid resi- dues of proteins play probably a very important role in the establishment and maintenance of ribonucleo- protein structures, which therefore must greatly de- pend on the ionic conditions of the buffers used.
Purified hnRNA . proteins from cells labeled in vivo for 1 h with 32P were exposed to increasing NaCl concentrations and sedimented through a su-
crose cushion under the same salt conditions. The amount and distribution of both radioactive and bulk proteins were then analysed in the resulting supernatants and particles pellets. Examination of the results of such an experiment, as presented in Fig.9, leads to two observations: (a) a large amount of proteins is released from hnRNA . proteins by ex- posure to high salt (85-90% at 0.5 M NaCI); (b) labeled phosphoproteins are also progressively releas- ed by this treatment, although they appear to be more firmly associated with RNA than the bulk of proteins: 50 % dissociation required 0.35 M salt for the former and only 0.2 M for the latter. 30% of the trichloro- acetic-acid-insoluble radioactivity is still recovered in the pellet after exposure to 0.75 M NaCl.
The proteins remaining associated with particles after exposure to various salt concentrations were ana- lysed by electrophoresis on sodium dodecylsulphate/ polyacrylamide slab gels followed by staining and autoradiography (Fig. 10). It appears that the 28000- M , species is the most readily dissociated by salt with a concomitant relative enrichment of the 37000- M , component in the particles. It should be noticed that for these two species, both the stained (Fig. 10A) and radioactive bands (Fig. 10B) behave similariy. This observation would be consistent with the hypo- thesis, although definitely not a proof, that the phos- phoryl groups are indeed carried by those polypeptides

J.-M. Blanchard, C. Brunel, and P. Jeanteur 309
which are among the major constituents of hnRNA proteins.
DISCUSSION The data presented in this paper show that some
discrete protein species associated with hnRNA in the form of hnRNA . proteins from HeLa cells can be labeled in vivo after a brief exposure of cells to [32P]orthopliosphate. The highest amount of label is found associated with polypeptides in the 28 000- 40000-M, range, where are also found the most abun- dant bands on a mass basis. In this region at least three species of MI respectively 28000, 30000 and 37000 are labeled, the first containing the highest amount of label. In the larger size range a discrete 52000-MI species stands out of a background of several other slightly labeled ones. It should be noted that a protein of similar size, which seems to be common to most cytoplasmic mRNA . protein is also phos- phorylated [28,29].
A very similar overall pattern has been observed by Gallinaro-Matringe et al. with rat brain nuclear particles [lo]. The pattern of phosphorylated species does not seem to depend on the time of labeling. All these phosphorylated species are found in all hnRNA . proteins size classes, although in different proportions.
As pointed out by Bhorjee and Pederson [14], the possibility of incomplete elimination of RNA from the proteins could have led to: (a) interference of labeled RNA with putative bands of phosphory- lated proteins, and (b) alteration of their migration and therefore of their size estimation through their attachment to RNA fragments. The first objection was eliminated by routinely subjecting our gels to hot trichloroacetic acid before autoradiography. The ef- fectiveness of such a treatment was confirmed by check- ing the absence of radioactivity in individual protein bands from [3H]uridine-labeled hnRNA . proteins. The second one appeared unlikely, although not de- finitely excluded in view of the absence of any detect- able change in the migration of all labeled species after positive removal of RNA.
Phosphoproteins appear on the whole to be more firmly bound to hnRNA than the bulk of proteins, as shown by their relative dissociation by increasing ionic strength. These results are in excellent agree- ment with those previously reported with rat brain nuclear particles [lo]. According to these authors, 0.25 M and 0.7 M salt solubilize respectively 5 5 % and 87 % of total proteins as compared to 23 % and 65% of label. However, in contrast to the overall behaviour of phosphoproteins, which appear to be more tightly bound to hnRNA, the 28 000-M, species is most readily dissociated with either salt or mild RNase treatment.
We have reported in a preceding paper [13] that endogenous phosphorylation of hnRNA . proteins incubated in vitro with [Y-~~PIATP resulted in label being incorporated into the same 28 000 and 37 000-M, species labeled in vivo. However, neither the 30000 nor the 52000-M, components appeared to be labeled to any comparable level as observed in the present work. This may reflect the fact that these species are already present in the particles at their maximum level of phosphorylation and cannot therefore be further phosphorylated.
The data in this paper are part of the doctoral thesis of J . M. Blanchard. We are grateful to D r M. Jacob for stimulating discus- sion and criticism throughout this work. Thanks are due to Mrs A. Vie for expert technical assistance. This work was supported by grants from Centre National de la Recherche Scientifique, Institut National de la SuntL: et de la Recherche M&dicale, Diligution GPnh- rule it Iu Rrcherche Scientifique et Technique, the Ligue Nutinnale Contie li. Cancer and The Fondation pour la Recherche Midicule.
REFERENCES 1.
2.
3.
4.
5 . 6.
7.
8.
9.
30.
11.
12.
13.
14.
15.
16. 17.
18.
29.
20.
21.
22. 23.
Ducamp, Ch. & Jeanteur, Ph. (1973) Biorhimie (Paris) 55,
Brunel, C., Blanchard, J . M., Ducamp-Guiinont, Ch. & Jean-
Niessing, J. & Sekeris, C. E. (1971) Biochim. Biophyr. Acict,
Matringe, H. &Jacob, M. (1972) Biochimie (Paris) 54, 1169-
Pederson, T. (1974) J . Mol. Bid. 83, 163-183. Karn, J., Johnson, E. M., Widali, G. & Allfrey, V. G. (1974)
J . B i d . Chem. 249, 667 - 677. Kleinsmith, L. J . (1974) in Acidic Proteins of the Nucleus
(Cameron, I. L. & Jeter, J. R., eds) pp. 103- 135, Academic Press, New York.
Olson, M. 0. J., Ezrailson, E. G., Guetzow, K. & Busch, I f . (1975) J . Mol. Bid. 97. 631-619.
Rubin, S. & Rosen, 0. M. (1975) Annu. Rev. Biochem. 44,
Gallinaro-Matringe, H., Stevenin, J. & Jacob, M. (1975) Biu-
Samarina, 0. P., Krichewskaya, A. & Georgiev, G. P. (1966)
Blanchard, J. M., Ducamp, Ch. & Jeanteur, Ph. (1975) Nuture
Blanchard, J. M., Brunel, C. & Jeanteur, Ph. (1977) E w . J .
Blanchard, J. M., Brunel, C. & Jeanteur, Ph. (1977) Biothcm.
Bhorjee, J. S. & Pederson, T. (1972) Proc. Null Acad. Sci.
Kish, V. M. & Pederson, T. (1975) J . Mol. Bid . 95, 227-238. Kaltschmidt, E. & Wittmann, H. G. (1972) Biochimie iPari.r)
Chilina, R. A., Chang, M., Ives, D. 13. & Koestner, A. (1976)
Baltimore, D. & IHuang, A . S. (1970) J . Mol. Biol. 47, 263-
Linn, S. & Lehman, I . R. (1965) J . B i d . Chrm. 240, 1287-
Kuno, Ii. & Kihara, H. K. (1967) Narure (Lond.) 2I5, 974-
Perry, R. P. & Kelley, D . E. (1968) J . Mol. Bid. 35, 37-59. Egly, J . M., Johnson, B. C., Stricker, C., Mandel, P. & Kempf,
1235-1243.
teur, Ph. (1975) A h t r . F d x Meet. no. 281.
247, 391 -403.
I1 78.
831 - 887.
chemistry, 14, 2547 - 2554.
Nature (Lond.) 210, 1319-1322.
(Lond.) 253,467 - 468.
Biochem. 79, 117-131.
Soc. Trans. 5,610- 671.
U.S.A. 69, 3345-3349.
54,167- 175.
Analyt. Biochem. 72,552 - 565.
273.
1293.
975.
J . (1972) FEBS Left . 22, 181-384.

310 J.-H. Blanchard, C. Brunel, and P. Jeanteur: Phosphorylation of Proteins from hnRNA Proteins
24. Warner, J. R. & Soeiro, R. (1967) Proc. Nut1 Acud. Sci. U . S . A . 27. Stevenin, J., Gallinaro-Matringe, H., Gattoni, R. & Jacob. M.
25. Kumar, A. & Pederson, T. (1975) J. Mol. Bioi. 96, 353-365. 28. Morel, C., Gander, E. S., Herzberg, M., Dubocher, J. & 26. Jelinek, W., Adesnik, M., Salditt, M., Sheiness, D., Wall, R. ,
Molloy, G., Philipson, L. & Darnell, J. E. (1973) J . Mol. 29. Auerbach, S. & Pederson, T. (1975) Biochem. Bzophys. Res. B id . 75, 515-532.
58,1984-1990. (1977) Eur. J . Biochem. 74, 589-602.
Scherrer, K . (1973) Eur. J . Biochem. 36, 455-464.
Commun. 63, 149- 156.
J.-M. Blanchard, The Rockefeller University, 66th Street and York Avenue, New York City, New York, U.S.A. 10021
C. Brunel and P. Jeanteur, Laboratoire de Biologie Moleculaire, Universiti. des Sciences et Techniques du Languedoc, Place Eugene Bataillon, F-34060 Montpellier-Cedex, France





























