Embed Size (px)
Citation preview

BIOCHIMICA ET BIOPHYSICA ACTA 22I
BBA 25926
PHOSPHORUS COMPOUNDS IN T H E HUMAN ERYTHROCYTE
GRANT R. BARTLETT
Laboratory for Comparative Biochemistry, San Diego, Calif. (U.S.A.)
(Received August 2Ist, I967)
SUMMARY
I. A procedure using column chromatography on Dowex I formate with gradient elution by ammonium formate buffer is recommended for the isolation of the water-soluble phosphate compounds of the human erythrocyte. The elution positions of many known compounds of biological interest were determined.
2. Metabolic turnover values for the various water-soluble phosphate compounds. present in freshly collected human erythrocytes were measured by the extent of incorporation of ~2p and 14C after a brief incubation of blood with 32p~ or uniformly 14C-labeled glucose.
3. After incubation of the human erythrocytes with [14C61 glucose, the metabolic intermediates were found to contain 14C in the following order of decreasing concen- tration: glucose 6-phosphate, fructose 6-phosphate, glucose 1,6-diphosphate UDPG ~ fructose 1,6-diphosphate ~ mannose 1,6-diphosphate ~ monophospho- glycerate, 2,3-diphosphoglycerate. No 14C was found in the ribose portion of any of the ribonucleotides.
4. After incubation of the erythrocytes with s2p~ the following order of de- creasing 32p concentration was found in the various phosphate groups of the cell:: ATP (P-3), ATP (P-2), ADP (P-2), GTP (~P) , APPX ( ~ P ) ~ glucose 6-phosphate, fructose 6-phosphate ~ fructose 1,6-diphosphate (P-l) ~ triose phosphate ~ fruc- tose L6-diphosphate (P-6) ~ glucose 1,6-diphosphate (P-6) ~ mannose 1,6-diphos- phate (P-6) ~ glucose 1,6-diphosphate (P-I) ~ mannose 1,6-diphosphate (P-I) UDPG (P-2) ~ monophosphoglycerate ~ 2,3-diphosphoglycerate (P-2), 2,3-diphos- phoglycerate (P-I). There was no incorporation of 32p into the phosphate attached to ribose in any of the nucleotides. No ~2p was found in NAD + of NADP +. No 32p. entered into two phosphorus compounds tentatively identified as inorganic poly- phosphates.
5. An individual with polycythemia vera was given ~2Pi intravenously Io mil~ before withdrawal of a sample of blood. The erythrocytes exposed to this incubation in vivo with 3~p showed the same relative specific activities of their phosphate groups a~ did human erythrocytes incubated in vitro.
INTRODUCTION
The author has reported previously on water-soluble phosphate compounds which were isolated by ion-exchange column chromatography from human ery-
Biochim. Biophys. Acta, 156 (I968) 221-23o

222 Go R. BARTLETT
*hrocytes ~. The present communication extends this study and includes informatiot? about the metabolic turnover rates of the red cell phosphates based on their incorpo- ration of asp or 1~C from labeled P1 or glucose.
Companion papers about the human erythroeyte include: (i) the identification of glucose and mannose diphosphates2; (if) the metabolism of purine ribonucleoside a i; and (iii) the metabolism of purine deoxyribonucleoside4.
MATERIALS AND METHODS
Chemicals Most phosphate compounds were obtained from commerciai sources and further
purified by ion-exchange chromatography. Glucose-x,6-P 2 and mannose-I,6-P~ were isolated from outdated blood bank erythrocytes 2. Ribose-I,5-P2 was separated from erythrocytes which had been incubated with inosine ~. Octulose-I,8-P~ was isolated from erythrocytes which had been incubated with purine ribonucleoside and was also prepared by treatment of dihydroxyacetone-P and ribose-5-P with muscle aldolase '~. Octulose-8-P was prepared by acid hydrolysis of octulose-I,8-P2. 5-Deoxyoctulose- 1,8-P 2 was isolated from erythrocytes which had been incubated with purine deoxy- ribonucleoside and was made enzymatically by the action of muscle atdolase on dihydroxyacetone-P and deoxyribose-5-P (ref. 4). Sedoheptulose-7-P, sedoheptulose- 1,7-P ~, ribulose-5-P and xyiulose-5-P were prepared by the methods of Ho~EcKzI~>. Inorganic tri- and tetra-, recta- and polyphosphates were gifts of J. R. VAN WAZER, Monsanto Chemical Co.
Isolation of e1,ythrocyte phosphates Blood was collected from healthy human donors into heparin (o.I mg/ml),
centrifuged in the cold for 30 rain at about 5000 A g and the surface layer of white cells removed. After determination of the hematocrit, a measured vo!ume of the packed erythrocytes was added slowly to 2 vol. of cold Io % trich[oroacetie acid. The centrifuged precipitate was thoroughly suspended in tile same volume of 5 % trichloro- acetic acid and recentrifuged. Trieh!oroacetie acid was removed from the combined solutions by extraction with about 2 vol. of ether 4 times and the residual e~her removed by evaporation with N~. The extract was dituted 5-fold and passed through a column of Dowex I-X8 formate (Ioo-325 wet mesh which was eluted ax room temperature at o.5-I.o ml/min by a linear gradient increase in ammonium formate (formic acid-ammonium formate 4: I, v/v). Columns I cm ~ 2o-3o cm were used for extracts of 5o ml of erythrocytes or less and columns I. 5 cm ~ 3o-4o cm for extracts from 5o to 3o0 ml of cells. Fractions of Io to 2o m1 (in 2o rain were collected auto- matically, and elution vol. of 2 to Io 1 were used depending on the volume of red ceiis used and degree of resolution desired. Elution with other ammonmm formate mixtures or with formic acid will be described ill the text in connection with the purification o~ individual compounds which were recovered from the first chromatographic separation and the isolation of individual phosphate groups for ~2p assay. Additional information about the chromatographic procedures and analytical methods is given in previous reports ~, a o
To identify the compounds which were eluted from tile chromatographic columns, each fraction was analysed first for total phospi~orus 7 and for ultraviolet
Biochim. Biophys. Asia, 156 (1968) 221-23o

ERYTHROCYTE PHOSPHATES 22~
absorption at 260 m/~. A prerequisite for identification was that the position of elution coincides with that which has been found with known compounds.
Isot@e experiments For isotope studies, freshly coilected blood was incubated with ~2Pi for 5 min at
37 ° or with E~4C6jglucose for Io rain just prior to the separation and extraction of the erythrocytes as described above. To measure the radioactivity, aliquots of the elution fractions were evaporated to dryness on planchets (all formate removed) and counted in a methane gas flow detector in the proportiona~ region under conditions which recorded approx. 5o % of the total disintegrations of the ~4C or 32p.
A nalyticaZ procedures The following special tests were performed to identify the individual compounds :
NAD +, fluorescence s and glyceraldehyde-3-P dehydrogenase; NADP+, fluorescence and glucose-6-P dehydrogenase; glucose-6-P, anthrone ~ and glucose-6-P dehydro- genase; fructose-6-P, carbazole i° and P-glucose isomerase plus glucose-6-P dehydro~ genase; dihydroxyacetone-P, Ioo ° carbazole a and glycerol-P dehydrogenase; glyceraldehyde-3-P, glyceraldehyde-3-P dehydrogenase; UDPG, UDPG dehydro- genase; glucose-L6-P 2, and mannose-I,6-P2, anthrone, phosphoglucomutase and hydrolysis to the monophosphates~; fructose-I,6-P2, carbazole and aldolase; mono- P-glycerate and 2,3-P2-glycerate, chromotropic acid ~ and acid resistant phosphate; P-enolpyruvate, dinitrophenylhydrazine ~2 and pyruvate kinase plus lactate dehydro~ genase; P1 (ref. I3).
Nucleotides were identified by their ultraviolet absorption spectra at pH 2, 7 and I I , after evaporation of aliquots of column eluates to dryness in a vacuum oven a~ 8o ° for at least 8 h to remove all traces of ammonium formate. The nucleotides were also identified by the ratio of purine to ribose (orcinol) and phosphorus. ADP and ATP were further identified with myokinase, hexokinase and glucose-6-P dehydro- genase.
The enzymatic methods were essentially according to BERGMEYER I4.
RESULTS
Chromatography of known phosphate compounds Formate buffers of different concentration and pH were tested for the ion-
exchange separation of known carbohydrate intermediates and nueieotides using columns of Dowex I-X8 formate. The best separation was achieved by elution -with a linear gradient increase from o to 5 M of an ammomum formate buffer which was prepared by mixing 4 parts of 5 M formic acid with I part of 5 M ammonium formate. This mixture gave a pH of approx. 3.o at I M. Table I lists the positions of elution which were found by this procedure for a number of water-soluble phosphate compounds of biological interest, if the compounds expected to be separated were limited to a lower range of resin binding strength, then the concentration of am- monium formate was reduced co give a better spread of the phosphate compounds throughout the total elution volume. The 5 M ammonium formate buffer was used as the eluant for the first chromatographic separation in most of the erythrocyte studies in order to be certain that very strongly acidic compounds such as the inorganic
Biochim. Biophys. Act~, _r56 Iz968) 22z-23o

224 G .R . BARTLETT
T A B L E I
ELUTION PATTERN OF PHOSPHATE COMPOUNDS
Approx. IO/*moles each of known compounds were chromatographed using I cm × 20 cm columns of Dowex I -X8 formate (lOO-325 wet mesh) which were eluted wi th 41 of linear gradient o to 5 M a m m o n i u m formate (formic a c i d - a m m o n i u m formate, 4:1 v/v) at a flow rate of o. 5 to o.8 inl/min, collecting 20 min fractions. The figures give the center of the elution posit ion as the percent of the tota l elufion volume.
Compound Elution position Compound Elution position
CMP o.5 A D P 2 i NAD + 5 2-P-Glycerate 24 Octulose-8-P 6 3 -P-Glycerate 24 AMP 7 Octulose-i,8-P2 26 Sedoheptulose-7-P 7 5-Deoxyoctulose-I,8-P~ 26 Glucose-I-P 7.5 Glncose-i,6-P~ 26 Glucose-6-P 8 Mannose- I, 6-Pu 27.3 F r u c t o s e - I - P 8 Sedoheptulose-I,7-P2 27.5 Fruc tose-6-P 8. 5 U D P 28. 5 ~z-Glycerol-P 8. 7 Fructose-I ,6-P~ 29 Deoxyribose-5-P 8.7 Ribose- I, 5-P~ 3 ° Ribose-5-P 9 CTP 3 ° -Ribulose-5-P 9.5 I D P 33 Xylulose-5-P 9.7 PPi 34 Glyceraldehyde-3-P I o 5 -P-Ribose- 1 -P-P 35 Dihydroxyace tone-P I I GDP 36 5 -Deoxyxylulose- I -P I I UDP-GlcUA 37 UMP I 1.5 P -Eno lpy ruva t e 38 NADP÷ 12 dATP 38 Pl 12. 5 ATP 38.5 ,CDP 13. 5 UTP 49 I M P 14 2,3-P2-Glycerate 5 ° Xao -5 ' -P 15. 5 I T P 52 6-P-Gluconate 18 Ado-5'-P-P-P-P 53 GMP 18. 5 GTP 54 UDP-GlcNAc 19 Tripoly-Pi 60 d A D P 20. 5 Trimeta-Pi 7 ° ~JDP-Glc 21 Tet rameta-Pi 72
Inosi tol -P 6 IOO
polyphosphates and inositol-P 6 would be eluted. Nucleotides which eluted in this system mixed with each other or with non-nucleotide phosphates could almost always be separated by rechromatography with formate at a different pH. In regard to the reproducibility of the results shown in Table I, the elution position of the nucleotldes relative to the other phosphates was in some cases extremely Sensitive to pH. Given a constant pH, the relative elution positions were reproducible. However, the values for elution position as percent of total elution volume were influenced by the length of the resin column, the anion load and the rate of elution.
Phosphates of the human erythroeyte Figs. I and 2 show typical phosphorus elution patterns which were obtained by
ion-exchange chromatography of trichloroacetic acid extracts of erythrocytes from freshly collected human blood. The erythrocytes used for the experiment of Fig. I were from a young healthy male without hematological problems. The donor for the s tudy illustrated in Fig. 2 was a middle aged male who had polycythemia vera but was
Biochim. Biophys. Acta, 156 (1968) 221-23o
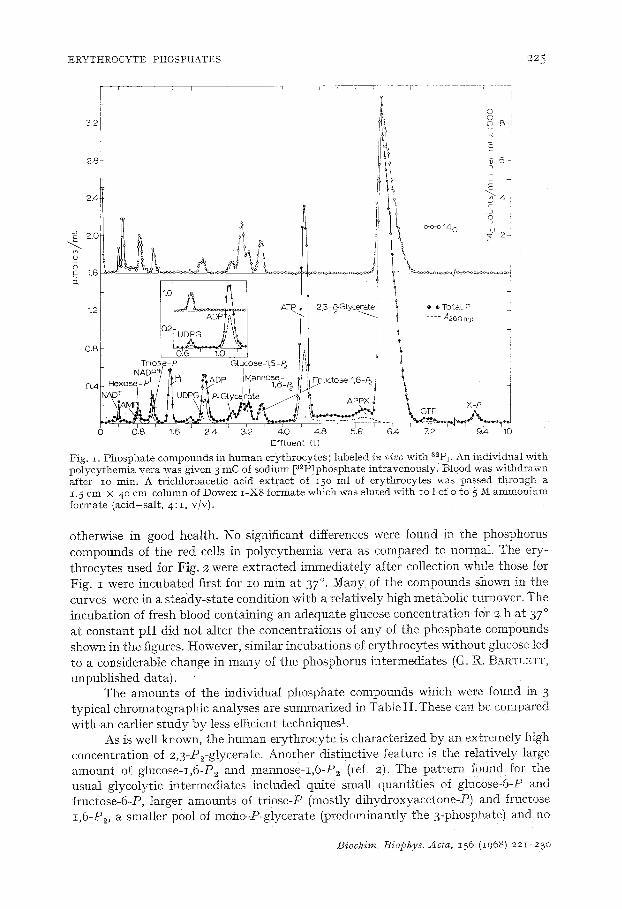
ERYTHROCYTE PHOSPHATES 22~
3.2F r L
2.8 t
2.4
..• 2.0
E t6 ::k
i I -T - - 7
1 © B] O × !
E
o~c-o 14 C ~u 2
1.0 i - - t - - ~ - ~ ~ A T ~ , 1.2
/ t 1 0.8
Tr iose-P GLucohe-1,6-P2 !I* ' NADP ÷ I J ,,/
0 4 .o o,o b
0 0.8 1.e 2.4 3.2 4.0 4.8 5.g 6,4 Z2 9.4 10 EffLuent (k)
Fig. z. Phospha te compounds in h u m a n erythrocytes ; labeled i~ vivo with ~2Pi. An individual wi th polycythemia vera was given 3 mC of sodium [a2PJphosphate intravenously. B10od was wi thdrawn after io rain. A trichloroacetic acid extract of z50 ml of erythrocytes was passed th rough a 1.5 cm X 4 ° cm column of Dowex I -X8 formate which was Muted with io 1 of o to 5 M ammon ium formate (acid-sal t , 4 : I , v/v).
otherwise in good health. No significant differences were found in the phosphorus compounds of the red cells in polycythemia vera as compared to normat. The ery- throcytes used for Fig. 2 were extracted immediately after collection while those for Fig. i were incubated first for zo rain at 37 °. Many of the compounds shown in the curves were in a steady-state condition with a relatively high metabolic turnover. The incubation of fresh blood containing an adequate glucose concentration for 2 h at 37 ° at constant pH did not alter the concentrations of any of the phosphate compounds shown in the figures. However, similar incubations of erythrocytes without glucose led to a considerable change in many of the phosphorus intermediates (G. R. BARTLETT, unpublished data).
The amounts of the individual phosphate compounds which were found in 3 typical chromatographic analyses are summarized in Table I I .These can be compared with all earlier s tudy by less efficient techniques 1.
As is well known, the human erythrocyte is characterized by an extremely high concentration of 2,3-P2-glycerate. Another distinctive feature is the relatively large amount of glucose-I,6-P2 and mannose-z,6-P2 (ref. 2). The pattern found for the usual glycolytic intermediates included quite small quantities of glucose-6-P and fructose-6-P, larger amounts of triose-P (mostly dihydroxyaeetone-P) and fructose z,6-P2, a smaller pool of mono-P-glycerate (predominantly the 3-phosphate) and no
Biochi*m. ]3iophys. Acts, i56 (I968) 221-23o
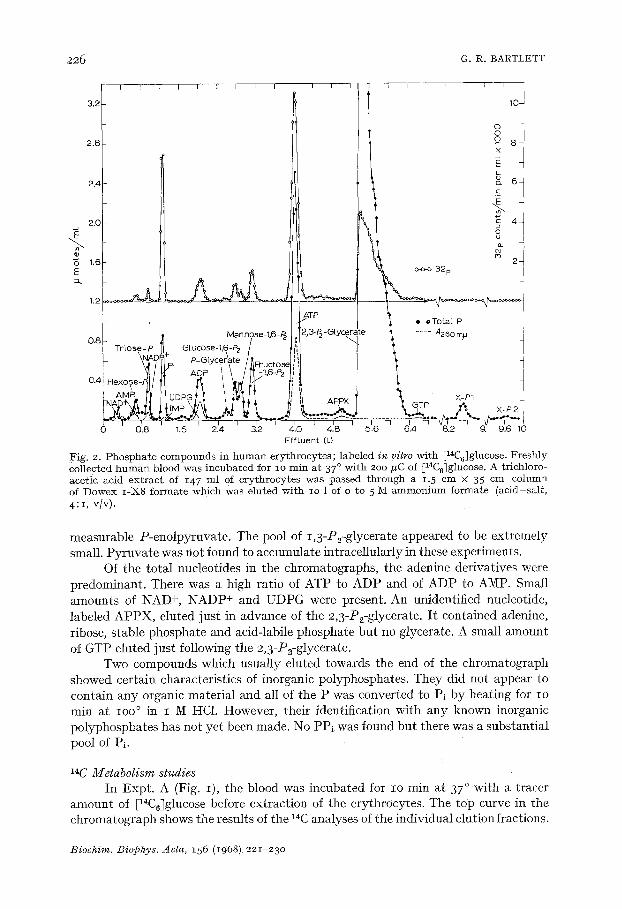
226 C. R. BARTLETT
3.2 0
I I tit i 2.4 c _
1,6 ~, o-o-o 3 2 p 2
1.2
I- I f " 1 l , - . Total P fl
A P P X X -P1
O 0 . 8 1.6 2 . 4 3.2 4 . 0 - - I ~ ~ ~ I i I t I ~ t ~8 5 [ 6 i 61.4 ~ 5. v ~2 i g. 9 "6 v ~ 10
E f f L u e n t (L)
Fig. 2. Phospha te compounds in h u m a n erythrocytes; labeled in vitro with [14C6]glucose. Freshly collected h u m a n blood was incubated for io min at 37 ° wi th 2oo ffC of [14C6]glucose. A trichloro- acetic acid ext rac t of 147 ml of erythrocytes was passed th rough a 1.5 cm × 35 cm column of Dowex I-X8 formate which was eluted wi th lO 1 of o to 5 M a m m o n i u m formate (acid-sal t , 4: i, v/v).
measurable P-enolpymvate . The pool of 1,3-P2-glycerate appeared to be extremely small. Pyruvate was not found to accumulate intracellularly in these experiments.
Of the total nucleotides in the chromatographs, the adenine derivatives were predominant. There was a high ratio of ATP to ADP and of ADP to AMP. Small amounts of NAD +, NADP+ and UDPG were present. An unidentified nucleotide, labeled APPX, eluted just in advance of the 2,3-P2-glycerate. I t contained adenine, ribose, stable phosphate and acid-labile phosphate but no glycerate. A small amount of GTP etuted just following the 2,3-P2-glycerate.
Two compounds which usually eluted towards the end of the chromatograph showed certain characteristics of inorganic polyphosphates. They did not appear to contain any organic material and all of the P was converted to P1 by heating for Io min at IOO ° in I M HC1. However, their identification with any known inorganic polyphosphates has not yet been made. No PP~ was found but there was a substantial pool of Pi.
14C Metabolism studies In Expt. A (Fig. I), the blood was incubated for zo rain at 37 ° with a tracer
amount of [14C6~glucose before extraction of the erythrocytes. The top curve in the chromatograph shows the results of the 14C analyses of the individual elution fractions.
Biochim. Biophys. Acta, 156 (1968)221-23o
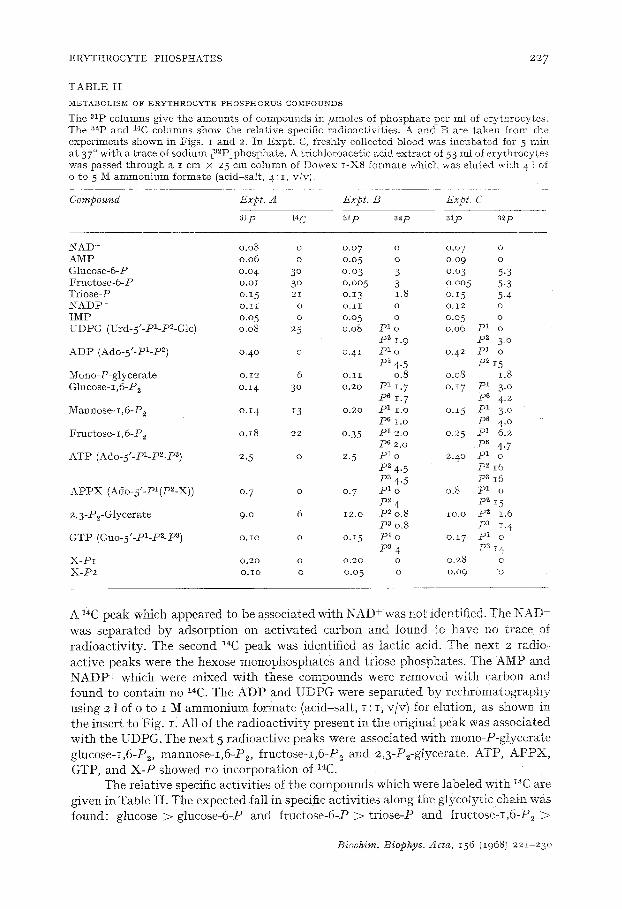
E R Y T H R O C Y T E P H O S P H A T E S 2 2 7
TABLE I I
M~TABOLISNI OF ERYTHIIOCYTE PILIOS]PI-IORUS COMPOUNDS
T h e a l p c o l u m n s g i v e t h e a m o u n t s of c o m p o u n d s i n # m o l e s of p h o s p h a t e p e r mI of e r y t h r o c y t e s = T h e a2p a n d 1~C c o l u m n s s h o w t h e r e l a t i v e spec i f i c r a d i o a c t i v i t i e s . A a n d B a r e t a k e n f r o m t h e e x p e r i m e n t s s h o w n i n F i g s . I a n d 2. I n E x p t . C, f r e s h l y c o l l e c t e d b l o o d w a s i n c u b a t e d f o r 5 r a i n a t 37 ° w i t h a t r a c e o f s o d i u m [ a 2 P ] p h o s p h a t e . A t r i c h l o r o a c e t i c a c i d e x t r a c t of 53 m l o f e r y t h r o c y t e s w a s p a s s e d t h r o u g h a I c m × 25 c m c o l u m n of D o w e x I - X 8 f o r m a t e w h i c h w a s e l u t e d w i t h 4 1 of o t o 5 M a m m o n i u m f o r m a t e ( a c i d - s a I t , 4 : i , v / v ) .
Compound Expt. A Expt. B Expt. C
alp 14 C alp 82p asp ~zp
N A D + 0 .08 o 0 .07 o o .o 7 o A M P 0 . 0 6 o 0 .05 o o .o9 o G l u c o s e - 6 - P o .o 4 3 ° o. o 3 3 o .o3 5 .3 F r u c t o s e - 6 - P o . o I 3 ° o - o o 5 3 o . o o 5 5.3 T r i o s e - P o . i 5 21 o .13 1.8 o . I 5 5 .4 N A D P + o . I I o o . I I o o . i 2 o I M P o .o 5 o 0 .05 o 0 .05 o U D P G (Urd-5"-P1-P~-Glc) o .o8 25 0 .o8 p 1 o o .o0 p 1 o
p 2 I -9 p 2 3. 0 A D P ( A d o - 5 ' - P I - P ~) 0 .40 o o . 4 i p 1 o 0 .42 p 1 o
p2 4-5 p2 i 5 M o n o - P - g l y c e r a t e o. 12 6 o . i I o .8 o .o8 1.8 G l u c o s e - i , 6 - P 2 o.x 4 3 ° 0 .20 p 1 iT. 7 o . i 7 p l 3~ o
p 6 1. 7 p 6 4. 2 M a n n o s e - r , 6 - P z o . i 4 13 o .2o p 1 I . o o . i 5 p 1 3. 0
p 6 I . o P~ 4.0 F r u c t o s e - I , 6 - P ~ o . i 8 22 o .35 p 1 2 .0 0 .25 p l 6 .2
p s 2 .0 p s 4.7 A T P (Ado-5'-Pi-p2-P 3) 2. 5 o 2. 5 P a o 2 .4o p 1 o
iD2 4.5 P~ I 6 p a 4 .5 P a i 6
APPX (Ado-5'-PI(P2-X)) 0. 7 o 0. 7 p1 o 0.8 p1 o P"~ 4 p2 z5
2 , 3 - P ~ - O l y c e r a t e 9 .0 6 12 .o P~ 0 .8 i o . o p 2 1.6 p3 0.8 pa i. 4
GTP (Guo-5'-PI-P2-P a) o.Io o o.15 pl o o.17 p1 o
P~ 4 pa 14 X-HI o.2o o o.2o o o.28 o
X - P 2 O.lO o 0 .05 o 0 .09 o
A l~C peak which appea red to be associa ted wi th N A D + was not identif ied. The N A D was sepa ra t ed b y adsorp t ion on a c t i v a t e d carbon and found "co have no t race of r ad ioac t iv i ty . The second 14C peak was ident if ied as lac t ic acid. The next 2 radio- ac t ive peaks were the hexose monophospha t e s and triose phosphates. The AMP and N A D P + which were mixed wi th these compounds were removed with carbon and found to conta in no 1~C. The A D P and U D P G were s epa ra t ed b y r e c h r o m a t o g r a p h y using 2 ] of o to I M a m m o n i u m formate (acid-sal t , I : I, VlW for etution, as ShOWn in the inser t to Fig. I . Al l of the r a d i o a c t i v i t y present in the original peak was associa ted wi th the UDPG. The nex t 5 r ad ioac t ive peaks were associa ted wi th mono-P-g!yce ra te g lucose- i ,6-P2, mannose -x ,6 -P> f ruc tose - I ,6 -P 2 and 2,3-P2-glycerate. ATP, A P P X , GTP, and X - P showed no incorpora t ion of 14C.
The re la t ive specific ac t iv i t ies of the compounds which were labe led wi th '4C are given in Table I I . The expec ted fa!l in specific ac t iv i t ies along the g lycoly t ie chain was found: glucose > glucose-6-P and f ruc tose-6-P > t r iose -P and f ructose-I ,6-P2 >
Biochim. Biop~ys. Acta, I 5 6 ( I968) 221 2 3 0

228 G. R. BARTLETT
mono-P-glycerate and 2,3-P~-glycerate > lactate.The radioactive results indicated a relatively high rate of movement of carbon from glucose-6-P through the glucose- 1,6-P 2 and mannose-i ,6-P 2 into UDPG. No trace of radioactivity entered into any of tile nucleotides.
a2p Metabolism studies In Expt. B (Fig. 2) the donor was given 3 mC of sodium [~2Plphosphate intra-
venously IO min before withdrawal of a sample of blood which was cooled during collection and then immediately processed as usual. The top curve in the chromato- graph shows the results of the a2p assays of the individual elution fractious.
In Expt. C, blood was withdrawn from a healthy young male and incubated for exactly 5 rain at 37 ° with a tracer amount of sodium [a2P]phosphate. The blood was then cooled quickly and the red cells extracted for chromatographic analysis.
Table I I shows the relative specific activities of the various phosphate groups of the compounds isolated from the erythrocytes in Expts. B and C.
The following procedures were employed to separate the individual phosphate groups for isotope assay. AMP and NADP+ were removed from the hexose monophos- phates and triose phosphates by adsorption on activated carbon followed by elution with 2 M ammonia in 40 % ethanol. UDPG was separated from ADP by rechromato- graphy as described above. The following compounds were hydrolyzed by heating for 30 rain at IOO ° in o.I M HC1 and the phosphate products were isolated by chromato- graphy on I cm x 20 cm columns of Dowex I-X8 formate which were eluted with I 1 of o to 2 M formic acid: ADP-+ ribose-5-P and Pi; UDPG ~ UMP and Pi; glucose- 1,6-P2-+ glucose-6-P and Pi; mannose-i ,6-P~-~ mannose-6-P and Pi; fructose-i,6- P=-~ fructose-6-P and Pi. The Pi was derived from the terminal phosphate of the ADP, the phosphate attached to glucose in UDPG and the phosphate in the I position of the sugar diphosphates. ATP was treated with glucose and hexokinase, the resulting glucose-6-P and ADP heated for 30 rain at IOO ° in o.I M HC1 and the products, ribose-5-P, glucose-6-P and Pi, isolated in the same chromatographic system. The 2,3-P2-glycerate was hydrolyzed by heating for 6h in I M acetic acid. This resulted in the splitting to PI of about 1/2 of the phosphate in the 2 position without any hydro- lysis of the phosphate in the 3 position and without any exchange of phosphate between the 2 and 3 positions*. The products, P~, 3-P-glycerate and 2,3-P2-glycerate, were isolated by chromatography on a I cm × 20 cm column of Dowex I-X8 formate which was Muted with I 1 of o to I M ammonium formate. The APPX and the GTP were hydrolyzed by heating for 30 rain in o.I M HC1 and the products, Pi (from all labile phosphate) and ribose-5-P, were isolated by the formic acid elution system, described above.
The results of the isotope labeling experiments shown in Table I I can be summarized as follows. The acid-labile phosphate groups of ADP, ATP, A P P X and GTP all showed close to the same specific activity which was significantly higher than any other phosphate group present in tile compounds isolated. Tile following com- pounds showed a decreasing concentration of asp: glucose-6-P and fructose-6-P > triose-P and fructose-i ,6-P 2 > mono-P-glycerate and 2,3-P2-glycerate. The 2 phos- phate groups of glucose-I,6-P~, mannose-I,6-P~ or fructose-I,6-P 2 showed only minor differences in specific activity. There was a relatively high specific activity in the
* O. i°t. BARTLETT, unpubl ished data.
Biochim. Biophys. Acta, I56 (1968) 221-23o

ERYTHROCYTE PHOSPHATES 22~
glucose-i,6-Ps and in the phosphate attached to glucose in UDPG and a lesser amount of a~p in the mannose-I,6-P2. No trace of radioactivity was found in the phosphate attached to ribose in AMP, ADP, ATP, APPX, GTP or UDPG. NAD+ and NADP + also showed no incorporation of a2p, and no radioactivity was found in the unknown compounds labeled X-P.
DISCUSSION
This paper has described an ion-exchange chromatographic procedure for the separation of a number of water-soluble phosphate compounds of biological interest and has applied the method to the isolation of such compounds from freshly coliected human erythroeytes and to a s tudy of their metabolic turnover as measured by the incorporation of 3~p from 32PI and of ~C from El~C,!gIucose.
In the experiment with [14C61glucose, the expected order of decreasing isotope concentration along the chain of glycolytic intermediates was found. There was evidence for an active shunt of carbon from glucose-6-P through giucose-I,6-P 2 and mannose-I ,6-P 2 to UDPG. No glucose carbon entered into the ribose part of any of the nucleotides.
Following the incubation of the erythrocytes, in vivo or in vitro, with a2p, the acid-labile phosphate groups of the nucleotides, ADP, ATP, APPX and GTP, showed essentially the same specific radioactivity, which was significantly higher than in any other phosphate group of the compounds isolated.The rapid equilibration of the labile phosphates of the nucleotides could have been produced bv the action of nucleotide phosphotransferases. The results are compatible with an initial uptake of the 32Pi during the oxidation of glyceraldehyde-3-P followed by transfer of the phosphorus through the carboxyl phosphate of 1,3-P2-giycerate to ADP, a mechanism suggested in previous reports 1~-~7, Subsequent phosphorytation by ATP of glucose and fruetose-6-P with the further movement of this phosphate into fructose-6-P, mannose-I .6-P 2, glucose-I,6-P~, UDPG and triose-P would account for the substantial drop m specific activity between the labile phosphate of the nucleotides and the phosphate in the various hexose phosphates and triose phosphates. The conversion of the phosphate in the 3 position of the triose-P zo the 3 position of mono-P-glycerate and its subsequent equilibration with the 2 phosphate groups of 2,3-P2-glycerare and with the phosphate of 2-P-glycerate in the phosphogiycerate mutase reaction wouId explain the large drop in specific activity at this stage.
The contamination of intracellular Pi with small amounts of very high specific activity extracellular Pi makes it difficult to determine which one is the preferred substrate for the first step of incorporation into organic phosphate.
There was no movement of a2p into the phosphate attached ~o ribose in any of the nucleotides. The stability of this bond agrees with the finding that no ~4C from glucose entered the nucleotide ribose. NAD + and NADP + proved ~o be very stable metabolically in the sense that they showed no trace of incorporation of ~4C or a~p.
The labeling with sap confirmed the results with ~*C which indicated an active shuttle from glucose-6-P to UDPG.
A comparison of the results obtained when the erythrocytes were exposed to 3"Pi in the circulation with the incubation in vitro shows that the metabolism under the 2 circumstances was almost identical.
Biochim. Biophys. Acfa. i56 (i968) 221-23o

230 G. R. BARTLETT
T h e p r e s e n t s t u d y conf i rms a n d e x t e n d s p r ev ious r epor t s b y BARTLETT 15,
GERLACH, I~'LECKENSTEIN AND GROSS is a n d YOSHIKAWA et al. 19 that following a brief i n c u b a t i o n of f r e sh ly co l l ec ted h u m a n e r y t h r o c y t e s w i t h 32Pi, t he labi le p h o s p h a t e
g roups of A T P a n d A D P s h o w e d a p p r o x i m a t e l y t h e s a m e a m o u n t of i n c o r p o r a t i o n of
32p w h i c h was s u b s t a n t i a l l y h ighe r t h a n t h e hexose p h o s p h a t e s wh ich in t u r n was
m u c h h i g h e r t h a n the 2 ,3 -P2-g lycera te .
ACKNOWLEDGEMENTS
Th i s w o r k was s u p p o r t e d b y G r a n t H E 6950 f r o m t h e N a t i o n a l H e a r t I n s t i t u t e ,
U . S. Pub l i c H e a l t h Service , a n d b y G r a n t D A - M D - 4 9 - I 9 3 f r o m t h e U.S. A r m y
Medica l R e s e a r c h a n d D e v e l o p m e n t C o m m a n d , D e p a r t m e n t of t h e A r m y .
REFERENCES
i G. R. 13ARTLETT, J. Biol. Chem., 234 (1959) 449- 2 G. R. BARTLETT, Bioehim. Biophys. Aeta, 156 (1968) 231. 3 G. R. ~AI~a:LW, TT AND G. BUCOLO, Biochim. Biophys. Acta, 156 (1968) 240. 4 G. R. 13ARTLETT, Biochim. Biophys, Acta, 156 (1968) 254. 5 B. L. HORECI~R, in S. P. COLOWICK AND N. O. t~_APLAN, Methods in Enzymology, Vol. 3,
Academic Press, New York, 1957, p. 193, 195. 6 G. R. BARTLETT, J . Biol. Chem., 234 (1959) 459. 7 G. R. ]BARTLETT, J. Biol. Chem., 234 (1959) 466. 8 N. LEVlTAS, J. ROBINSON, F. ROSEN, J. W. HuFF AND W. G. PERLZW~m, J. Biol. Chem., 167
(1947) 169. 9 R. DREYWOOD, Ind. Eng. Gl~em. Anal. ed., 18 (1946) 499.
IO Z. DISCHE AND E. BORiSNFR~tlND, J. Biol. Chem., 192 (1951) 583 . I I G. R. t3ARTLX~ra:, J. Biol. Chem., 234 (1959) 469 . 12 G. D. LF, Biochem. ,[., 33 (1939) 249. 13 C. H. FISI<E AND Y. SUBBAROW, 3 r. Biol. Chem., 66 (1925) 375. 14 H. U. BERGMEYER, Methods of Enzymatic Analysis, Academic Press, New York, 1963. 15 G. R. BARTLETT, Ann. N. Y. Acad. Sei., 75 (1958) IiO. 16 S. L. SC~IRIER, jr. Clin. Invest., 42 (1963) 756. 17 G. RONQUIST AND G. AGREN, Nature, 209 (1966) lO9O. 18 E. GERLACH, A. FLECKEI~Sa'a~IN AI~D E. GROSS, Arch. Ges. Physiol., 266 (1958) 528. 19 H. YOSIIIKAWA, M. NAKAO, K. MIYAMOTO AND IV[. TATIBANA, Jr. Biochem. Tokyo, 47 (196°) 635.
Biochim. Biophys. Acta, 156 (1968) 221-23o





























