Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 2’70, No. 1, April, pp. 42-49,1989
Phosphatidylinositol Modulates the Response of Calmodulin-Dependent Phosphatase to Calmodulin’
SHENGLI HUANG,2 DENNIS MERAT, AND WA1 YIU CHEUNG3
Department of Biochemistry, St. Jude Children’s Research Hospital, 3.92 North Lmderdak, Memphis, Tennessee 38101
Received August 3,1988, and in revised form November 15,1933
Phosphatidylinositol (PtdIns) and many other phospholipids activated calmodulin (CaM)-dependent phosphatase in the presence or absence of Ca2+, and the stimulation was more pronounced in the presence of Ca2+. In addition, PtdIns modulated the re- sponse of phosphatase to CaM: at low and nonstimulatory concentrations (~70 PM),
PtdIns augmented the activity of phosphatase by a submaximum concentration of CaM, giving a synergistic effect; and at high concentrations (>lOO PM), PtdIns suppressed the synergistic effect. Kinetic experiments indicated that PtdIns (both nonstimulatory and stimulatory concentrations) increased the affinity of phosphatase for CaM. In addition to the CaM regulatory site, phosphatase appears to have two PtdIns regulatory sites: a high-affinity site the occupation of which does not stimulate enzyme activity, and a low- affinity site the occupation of which stimulates enzyme activity in the absence of CaM and inhibits it in the presence of CaM. Modulating the response of phosphatase to CaM is not unique to PtdIns, and was observed with other phospholipids, including some that did not stimulate the enzyme. This raises the possibility that certain phospholipids may regulate phosphatase in two ways: (i) direct activation of the enzyme and (ii) modulation of its response to CaM. o 198s Academic press, I,,~.
CaM4 as a major intracellular receptor for Ca2+ regulates many Ca2+-dependent enzymes and plays an important role in a wide spectrum of cellular functions (1, 2). Stimulation of a cell increases its intracel- lular concentration of Ca2+, which binds to CaM and exposes a hydrophobic region that is believed to interact with the target enzyme. We and others have shown that
’ This work was supported by Grants CA 21765 and GM 36734 from the National Institutes of Health, and by American Lebanese Syrian Associated Charities
(ALSAC). * On leave from the Cancer Institute, Chinese Acad-
emy of Medical Sciences, Beijing, People’s Republic of China.
3 To whom correspondence should be addressed. ’ Abbreviations used: CaM, calmodulin; PtdIns,
phosphatidylinositol; EGTA ethylene glycol bis(,T-
aminoethyl ether) N,N’-tetraacetic acid.
certain phospholipids stimulate phospho- diesterase (3, 4) to a level comparable to that of CaM. Subsequently several groups of investigators have noted that phospho- lipids also stimulate other CaM-dependent enzymes such as Ca2+-ATPase (5, 6) and adenylate cyclase (7). Politino and King (8) recently reported that the effect of certain phospholipids can either stimulate or inhibit CaM-dependent phosphatase de- pending on the substrate.
In view of the increasing interest on the role of phospholipids in regulating cellular processes, we have investigated further the effect of phospholipids on the activity of phosphatase. The present study reveals that phospholipids not only stimulate phosphatase activity but also modulate the response of the enzyme to CaM, an obser- vation that appears not to have been re- ported before.
0603-9861/89 $3.00 42 Copyright 0 1989 by Academic Press, Inc. All rights of reproduction in any form reserved.
MODULATION OF CALCINEURIN PHOSPHATASE 43
MATERIALS AND METHODS
Materids. CaM-dependent phosphatase (9) and CaM (10) were purified to homogeneity from bovine brain. Casein was phosphorylated with [y-32PjATP using the catalytic subunit of CAMP-dependent pro- tein kinase (11) purified from bovine heart (12). Ca- sein was purchased from Sigma; [T-~P]ATP (sp act 2900-3000 Ci/mmol ATP) from New England Nuclear; phosphatidylinositol (a M, 740 was used to estimate its concentration), lysophosphatidylinositol (M, 618). phosphatidylcholine (M, ‘787), and lysophos- phatidylcholine (Mr 508) from Serdary Labs; and phosphatidylserine (M, 733), lysophosphatidylserine (M, 569), phosphatidylethanolamine (M, 744), lyso-
phosphatidylethanolamine (M, 498), and phosphati- dylglycerol (Mr 775) from Avanti.
Assay of phosphtase. Phosphatase activity was measured at 30°C essentially as before (13). The reac- tion mixture (50 ~1) contained 50 mM Tris-HCl (pH 7.0), 0.5 mM dithiothreitol, 0.1 mM CaClr, 1 mg/ml bo- vine serum albumin, 15 pM [=P]casein, and 12.5 or 25 nM phosphatase. CaM or phospholipids, where added, are given in the figure or table legends. Assays were usually run for 15 min such that the substrate de- phosphorylated was less than 15%. Specific activity is defined as nanomoles of 32P released per milligram protein per minute.
Preparation of phpholipid Aliquots of phospho- lipids in chloroform or methanol were dried under a stream of nitrogen and were taken up in a suspension of 10 mM Tris-HCl (pH 7.0) by sonieating for 20 s at 0°C. The samples were used fresh. Since sonication causes phospholipids to form vesicular structures, the concentrations of the phospholipids listed hence- forth are not strictly true concentrations and should be understood with this fact in mind.
RESULTS
Stimulation of CaM-Dependent Phosphatase bg Phospholipids
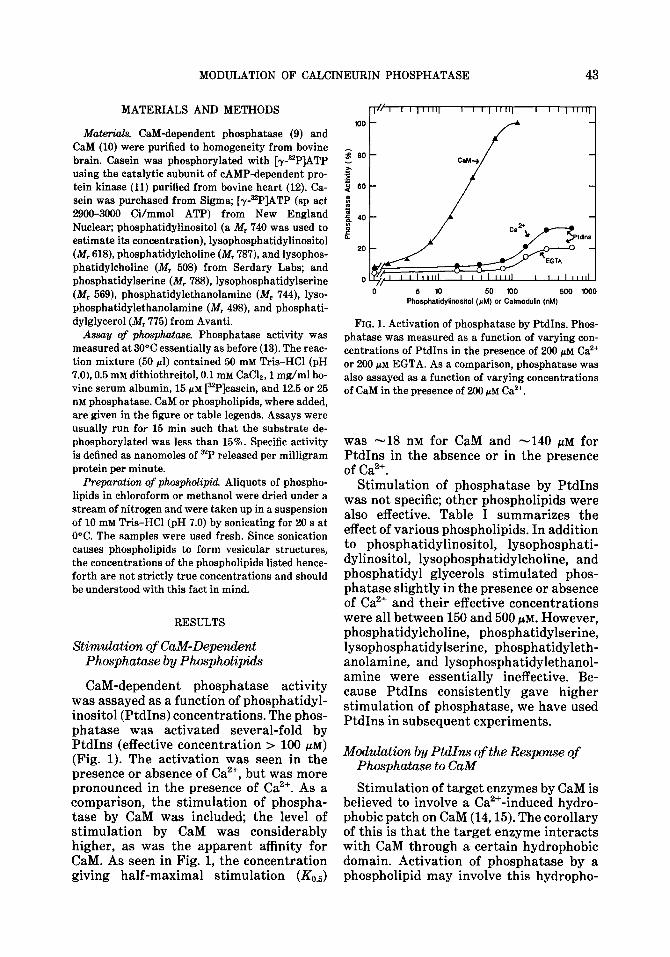
CaM-dependent phosphatase activity was assayed as a function of phosphatidyl- inositol (PtdIns) concentrations. The phos- phatase was activated several-fold by PtdIns (effective concentration > 100 PM) (Fig. 1). The activation was seen in the presence or absence of Ca2+, but was more pronounced in the presence of Ca2+. As a comparison, the stimulation of phospha- tase by CaM was included; the level of stimulation by CaM was considerably higher, as was the apparent affinity for CaM. As seen in Fig. 1, the concentration giving half-maximal stimulation (&,J
5 10 50 100 500 1000 Phosphatidylinositol (CM) or Calmodulin (nM)
FIG. 1. Activation of phosphatase by PtdIns. Phos- phatase was measured as a function of varying con- centrations of PtdIns in the presence of 200 pM Ca2+ or 200 pM EGTA. As a comparison, phosphatase was also assayed as a function of varying concentrations of CaM in the presence of 200 pM Ca’+.
was -18 nM for CaM and -140 PM for PtdIns in the absence or in the presence of Ca2+.
Stimulation of phosphatase by PtdIns was not specific; other phospholipids were also effective. Table I summarizes the effect of various phospholipids. In addition to phosphatidylinositol, lysophosphati- dylinositol, lysophosphatidylcholine, and phosphatidyl glycerols stimulated phos- phatase slightly in the presence or absence of Ca2’ and their effective concentrations were all between 150 and 500 PM. However, phosphatidylcholine, phosphatidylserine, lysophosphatidylserine, phosphatidyleth- anolamine, and lysophosphatidylethanol- amine were essentially ineffective. Be- cause PtdIns consistently gave higher stimulation of phosphatase, we have used PtdIns in subsequent experiments.
Moduluticm by PtdIns of the Resporwe of Phosphatase to CaM
Stimulation of target enzymes by CaM is believed to involve a Ca2+-induced hydro- phobic patch on CaM (14,15). The corollary of this is that the target enzyme interacts with CaM through a certain hydrophobic domain. Activation of phosphatase by a phospholipid may involve this hydropho-
44 HUANG, MERAT, AND CHEUNG
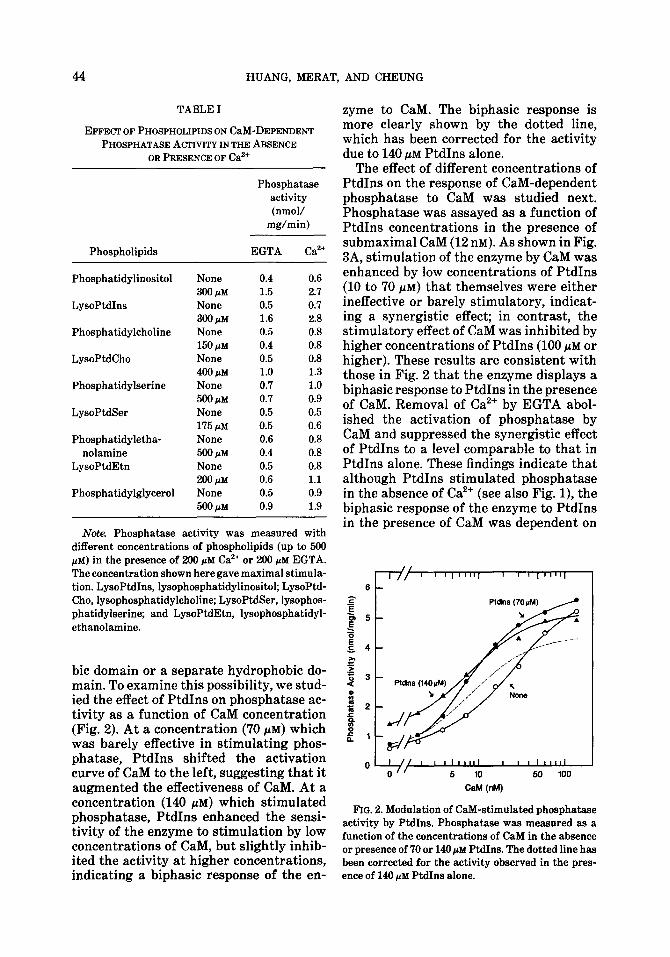
TABLE I
EFFECT OF PHOSPHOLIPIDS ON Cal&DEPENDENT PHOSPHATASE ACTIVITY IN THE ABSENCE
OR PRESENCE OF Caa’
Phosphatase activity (nmol/
mg/min)
Phospholipids EGTA Ca*+
Phosphatidylinositol None 0.4 0.6 300 pM 1.5 2.7
LysoPtdIns None 0.5 0.7 300 pM 1.6 2.8
Phosphatidylcholine None 0.5 0.8 150 pM 0.4 0.8
LysoPtdCho None 0.5 0.8 400 @VI 1.0 1.3
Phosphatidylserine None 0.7 1.0 mm 0.7 0.9
LysoPtdSer None 0.5 0.5 175 pM 0.5 0.6
Phosphatidyletha- None 0.6 0.8 nolamine 500 PM 0.4 0.8
LysoPtdEtn None 0.5 0.8 200 pM 0.6 1.1
Phosphatidylglycerol None 0.5 0.9 mm 0.9 1.9
Note. Phosphatase activity was measured with different concentrations of phospholipids (up to 500 PM) in the presence of 200 PM Ca2+ or 200 PM EGTA. The concentration shown here gave maximal stimula- tion. LysoPtdIns, lysophosphatidylinositol; LysoPtd- Cho, lysophosphatidylcholine; LysoPtdSer, lysophos- phatidylserine; and LysoPtdEtn, lysophosphatidyl- ethanolamine.
bit domain or a separate hydrophobic do- main. To examine this possibility, we stud- ied the effect of PtdIns on phosphatase ac- tivity as a function of CaM concentration (Fig. 2). At a concentration (70 PM) which was barely effective in stimulating phos- phatase, PtdIns shifted the activation curve of CaM to the left, suggesting that it augmented the effectiveness of CaM. At a concentration (140 f&M) which stimulated phosphatase, PtdIns enhanced the sensi- tivity of the enzyme to stimulation by low concentrations of CaM, but slightly inhib- ited the activity at higher concentrations, indicating a biphasic response of the en-
zyme to CaM. The biphasic response is more clearly shown by the dotted line, which has been corrected for the activity due to 140 PM PtdIns alone.
The effect of different concentrations of PtdIns on the response of CaM-dependent phosphatase to CaM was studied next. Phosphatase was assayed as a function of PtdIns concentrations in the presence of submaximal CaM (12 DM). As shown in Fig. 3A, stimulation of the enzyme by CaM was enhanced by low concentrations of PtdIns (10 to ‘70 PM) that themselves were either ineffective or barely stimulatory, indicat- ing a synergistic effect; in contrast, the stimulatory effect of CaM was inhibited by higher concentrations of PtdIns (100 PM or higher). These results are consistent with those in Fig. 2 that the enzyme displays a biphasic response to PtdIns in the presence of CaM. Removal of Ca2+ by EGTA abol- ished the activation of phosphatase by CaM and suppressed the synergistic effect of PtdIns to a level comparable to that in PtdIns alone. These findings indicate that although PtdIns stimulated phosphatase in the absence of Ca2+ (see also Fig. l), the biphasic response of the enzyme to PtdIns in the presence of CaM was dependent on
FIG. 2. Modulation of CaM-stimulated phosphatase activity by PtdIns. Phosphatase was measured as a function of the concentrations of CaM in the absence or presence of 70 or 140 CM PtdIns. The dotted line has been corrected for the activity observed in the pres- ence of 140 prd PtdIns alone.
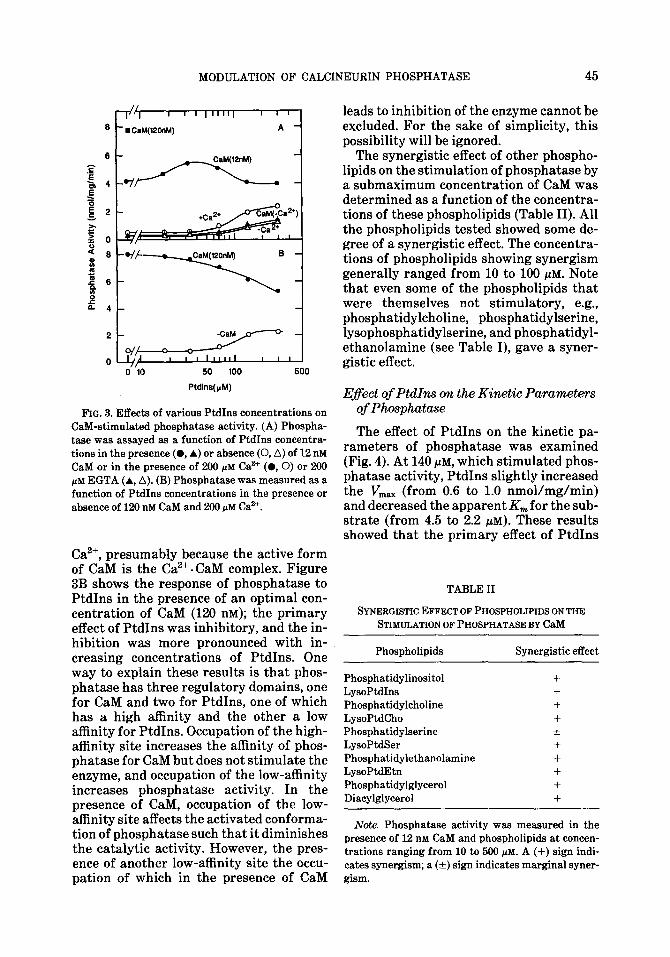
MODULATION OF CALCINEURIN PHOSPHATASE 45
50 100
Ptdlns(p.4)
FIG. 3. Effects of various PtdIns concentrations on CaM-stimulated phosphatase activity. (A) Phospha- tase was assayed as a function of PtdIns concentra-
tions in the presence (0, A) or absence (0, A) of 12 n&i CaM or in the presence of 200 PM Car’ (0,O) or 200 PM EGTA (A, A). (B) Phosphatase was measured as a
function of PtdIns concentrations in the presence or absence of 120 nM CaM and 200 PM Ca”.
Ca’+, presumably because the active form of CaM is the Ca”. CaM complex. Figure 3B shows the response of phosphatase to PtdIns in the presence of an optimal con- centration of CaM (120 nM); the primary effect of PtdIns was inhibitory, and the in- hibition was more pronounced with in- creasing concentrations of PtdIns. One way to explain these results is that phos- phatase has three regulatory domains, one for CaM and two for PtdIns, one of which has a high affinity and the other a low affinity for PtdIns. Occupation of the high- affinity site increases the affinity of phos- phatase for CaM but does not stimulate the enzyme, and occupation of the low-affinity increases phosphatase activity. In the presence of CaM, occupation of the low- affinity site affects the activated conforma- tion of phosphatase such that it diminishes the catalytic activity. However, the pres- ence of another low-affinity site the occu- pation of which in the presence of CaM
leads to inhibition of the enzyme cannot be excluded. For the sake of simplicity, this possibility will be ignored.
The synergistic effect of other phospho- lipids on the stimulation of phosphatase by a submaximum concentration of CaM was determined as a function of the concentra- tions of these phospholipids (Table II). All the phospholipids tested showed some de- gree of a synergistic effect. The concentra- tions of phospholipids showing synergism generally ranged from 10 to 100 PM. Note that even some of the phospholipids that were themselves not stimulatory, e.g., phosphatidylcholine, phosphatidylserine, lysophosphatidylserine, and phosphatidyl- ethanolamine (see Table I), gave a syner- gistic effect.
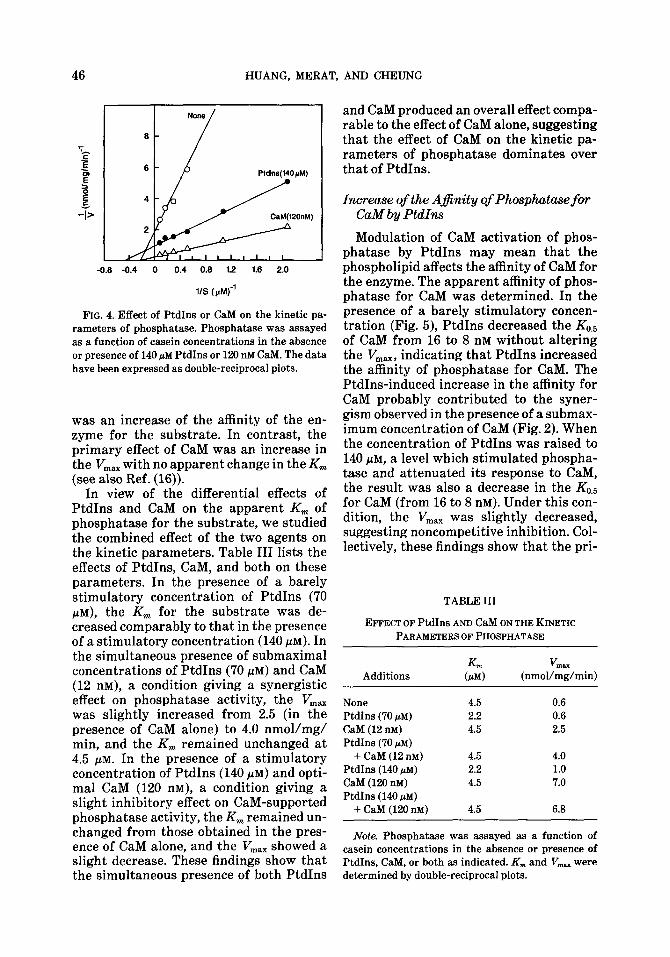
Eflect of PtdIns on the Kinetic Parameters of Phosphatase
The effect of PtdIns on the kinetic pa- rameters of phosphatase was examined (Fig. 4). At 140 /IM, which stimulated phos- phatase activity, PtdIns slightly increased the V,,,, (from 0.6 to 1.0 nmol/mg/min) and decreased the apparent K,,, for the sub- strate (from 4.5 to 2.2 PM). These results showed that the primary effect of PtdIns
TABLE II
SYNERGISTICEFFECTOFPHOSPHOLIPIDSONTHE STIMULATIONOFPHOSPHATASEBYC~M
Phospholipids Synergistic effect
Phosphatidylinositol +
LysoPMIns +
Phosphatidylcholine +
LysoPtdCho + Pbosphatidylserine + LysoPtdSer +
Pbospbatidylethanolamine + LysoPtdEtn + Phosphatidylglycerol +
Diacylglycerol +
Note. Phosphatase activity was measured in the presence of 12 nM CaM and phospholipids at concen-
trations ranging from 10 to 500 PM. A (+) sign indi- cates synergism; a (k) sign indicates marginal syner- gism.
HUANG, MERAT, AND CHEUNG
-0.8 -0.4 0 0.4 0.6 1.2 1.6 2.0
11s tph4 j'
FIG. 4. Effect of PtdIns or CaM on the kinetic pa- rameters of phosphatase. Phosphatase was assayed as a function of casein concentrations in the absence or presence of 140 pM PtdIns or 120 nM CaM. The data have been expressed as double-reciprocal plots.
was an increase of the affinity of the en- zyme for the substrate. In contrast, the primary effect of CaM was an increase in the V,,, with no apparent change in the Km (see also Ref. (16)).
In view of the differential effects of PtdIns and CaM on the apparent Km of phosphatase for the substrate, we studied the combined effect of the two agents on the kinetic parameters. Table III lists the effects of PtdIns, CaM, and both on these parameters. In the presence of a barely stimulatory concentration of PtdIns (70 FM), the Km for the substrate was de- creased comparably to that in the presence of a stimulatory concentration (140 PM). In the simultaneous presence of submaximal concentrations of PtdIns (70 PM) and CaM (12 nM), a condition giving a synergistic effect on phosphatase activity, the V,,, was slightly increased from 2.5 (in the presence of CaM alone) to 4.0 nmol/mg/ min, and the Km remained unchanged at 4.5 PM. In the presence of a stimulatory concentration of PtdIns (140 PM) and opti- mal CaM (120 nM), a condition giving a slight inhibitory effect on CaM-supported phosphatase activity, the Km remained un- changed from those obtained in the pres- ence of CaM alone, and the V,, showed a slight decrease. These findings show that the simultaneous presence of both PtdIns
and CaM produced an overall effect compa- rable to the effect of CaM alone, suggesting that the effect of CaM on the kinetic pa- rameters of phosphatase dominates over that of PtdIns.
Increase of the Afinity of Phosphatase fm CaM by PtdIns
Modulation of CaM activation of phos- phatase by PtdIns may mean that the phospholipid affects the affinity of CaM for the enzyme. The apparent affinity of phos- phatase for CaM was determined. In the presence of a barely stimulatory concen- tration (Fig. 5), PtdIns decreased the Ko, of CaM from 16 to 8 nM without altering the Vi,,,, indicating that PtdIns increased the affinity of phosphatase for CaM. The PtdIns-induced increase in the affinity for CaM probably contributed to the syner- gism observed in the presence of a submax- imum concentration of CaM (Fig. 2). When the concentration of PtdIns was raised to 140 PM, a level which stimulated phospha- tase and attenuated its response to CaM, the result was also a decrease in the Ko.5 for CaM (from 16 to 8 nM). Under this con- dition, the V,,, was slightly decreased, suggesting noncompetitive inhibition. Col- lectively, these findings show that the pri-
TABLE III
EFFECT OF PtdIns AND CaM ON THE KINETIC PARAMETERS OF PHOSPHATASE
Additions
None PtdInS (70 pM)
CaM (12 nM) PtdIns (70 pM)
+ CaM (12 nM) PtdIns (140 pM) CaM (120 nM) PtdIns (140 pM)
+ CaM (120 nM)
KWl V
(PM) (nmol/&/min)
4.5 0.6
2.2 0.6 4.5 2.5
4.5 4.0 2.2 1.0 4.5 7.0
4.5 6.8
Note. Phosphatase was assayed as a function of casein concentrations in the absence or presence of PtdIns, CaM, or both as indicated. K,,, and V,,, were determined by double-reciprocal plots.
MODULATION OF CALCINEURIN PHOSPHATASE 47
-0.1 -0.05 0 0.05 0.1 0.15 0.2 0.25
l/CaM (nM)-’
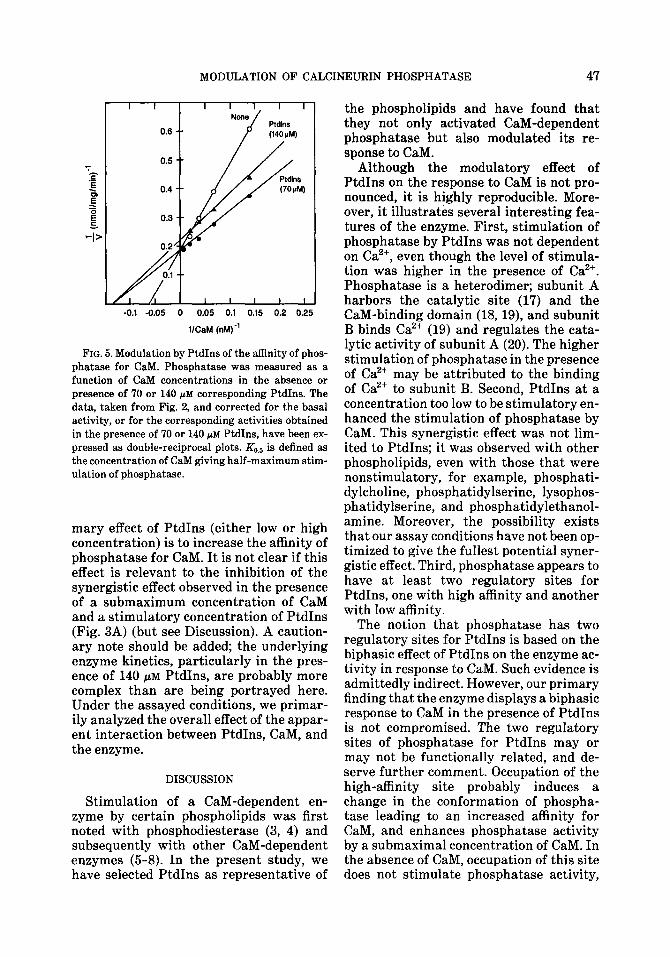
FIG. 5. Modulation by PtdIns of the affinity of phos- phatase for CaM. Phosphatase was measured as a
function of CaM concentrations in the absence or presence of 70 or 140 PM corresponding PtdIns. The data, taken from Fig. 2, and corrected for the basal
activity, or for the corresponding activities obtained in the presence of 70 or 140 pM PtdIns, have been ex- pressed as double-reciprocal plots. && is defined as
the concentration of CaM giving half-maximum stim- ulation of phosphatase.
mary effect of PtdIns (either low or high concentration) is to increase the affinity of phosphatase for CaM. It is not clear if this effect is relevant to the inhibition of the synergistic effect observed in the presence of a submaximum concentration of CaM and a stimulatory concentration of PtdIns (Fig. 3A) (but see Discussion). A caution- ary note should be added; the underlying enzyme kinetics, particularly in the pres- ence of 140 PM PtdIns, are probably more complex than are being portrayed here. Under the assayed conditions, we primar- ily analyzed the overall effect of the appar- ent interaction between PtdIns, CaM, and the enzyme.
DISCUSSION
Stimulation of a CaM-dependent en- zyme by certain phospholipids was first noted with phosphodiesterase (3, 4) and subsequently with other CaM-dependent enzymes (5-8). In the present study, we have selected PtdIns as representative of
the phospholipids and have found that they not only activated CaM-dependent phosphatase but also modulated its re- sponse to CaM.
Although the modulatory effect of PtdIns on the response to CaM is not pro- nounced, it is highly reproducible. More- over, it illustrates several interesting fea- tures of the enzyme. First, stimulation of phosphatase by PtdIns was not dependent on Ca’+, even though the level of stimula- tion was higher in the presence of Ca”. Phosphatase is a heterodimer; subunit A harbors the catalytic site (17) and the CaM-binding domain (l&19), and subunit B binds Ca2+ (19) and regulates the cata- lytic activity of subunit A (20). The higher stimulation of phosphatase in the presence of Ca2+ may be attributed to the binding of Ca2+ to subunit B. Second, PtdIns at a concentration too low to be stimulatory en- hanced the stimulation of phosphatase by CaM. This synergistic effect was not lim- ited to PtdIns; it was observed with other phospholipids, even with those that were nonstimulatory, for example, phosphati- dylcholine, phosphatidylserine, lysophos- phatidylserine, and phosphatidylethanol- amine. Moreover, the possibility exists that our assay conditions have not been op- timized to give the fullest potential syner- gistic effect. Third, phosphatase appears to have at least two regulatory sites for PtdIns, one with high affinity and another with low affinity.
The notion that phosphatase has two regulatory sites for PtdIns is based on the biphasic effect of PtdIns on the enzyme ac- tivity in response to CaM. Such evidence is admittedly indirect. However, our primary finding that the enzyme displays a biphasic response to CaM in the presence of PtdIns is not compromised. The two regulatory sites of phosphatase for PtdIns may or may not be functionally related, and de- serve further comment. Occupation of the high-affinity site probably induces a change in the conformation of phospha- tase leading to an increased affinity for CaM, and enhances phosphatase activity by a submaximal concentration of CaM. In the absence of CaM, occupation of this site does not stimulate phosphatase activity,

48 HUANG, MERAT, AND CHEUNG
even though it enhances the affinity of the enzyme for the substrate. On the other hand, occupation of the low-affinity site stimulates the enzyme in the absence of CaM and inhibits the enzyme in the pres- ence of CaM. It is not clear why occupation of both PtdIns sites suppresses the syner- gistic effect of CaM. One possibility is that excess PtdIns competes with CaM for the CaM-regulatory site on phosphatase, ren- dering the enzyme apparently less respon- sive or even unresponsive to CaM. Such an explanation is not satisfactory; increasing the concentration of CaM did not amelio- rate the inhibition, but accentuated it. An- other possibility is that excess PtdIns in- terferes with the utility of the substrate. This possibility also appears unlikely; a comparable high concentration of PtdIns did not inhibit phosphatase activity in the absence of CaM (see Fig. 1). A third possi- bility is that the occupation of both PtdIns regulatory sites in the presence of CaM may lead to a new conformation of phos- phatase such that its activity is sup- pressed, even though this conformation may still display an increased affinity for CaM.
The apparent increase of the affinity be- tween phosphatase and CaM by PtdIns has been considered from the modulatory effect of PtdIns on phosphatase. It is con- ceivable that PtdIns may also interact with and bring about a conformation change on CaM such that its affinity for the enzyme is enhanced. Such a notion appears compatible with the finding described herein and also with that of Johnson (21) who noted that binding of one hydrophobic ligand to CaM increased its affinity for the binding of another ligand.
In bovine brain, the distribution of CaM- dependent phosphatase is half soluble and half particulate. The studies carried out here used phosphatase prepared from the soluble fraction. The subunit structure and the biochemical properties of the particu- late enzyme that have been examined ap- pear similar to those of the soluble enzyme (22, 23). Phospholipids are associated pri- marily with membranes, and the findings described here may be equally applicable to the particulate enzyme.
Although the concentration of CaM is generally high in most tissues (1, 2), it is not known if it is uniformly accessible to its target enzyme(s). The sensitivity of phosphatase to PtdIns (or other phospho- lipids) may prime the enzyme to be more responsive to a low concentration of CaM under certain conditions and to prevent ex- cess stimulation by high concentrations of CaM under other conditions. Such consid- erations, however, should be taken with reservation; it is difficult to ascertain the local cellular concentrations of CaM, PtdIns, or other phospholipids and there- fore difficult to define whether the findings reported here are physiologically relevant. Nevertheless, the modulatory effect of PtdIns on phosphatase in the presence of CaM is not an isolated observation; a quali- tatively similar effect has been observed with a CaM-dependent phosphodiester- ase.5 It appears worthwhile in future stud- ies to establish whether PtdIns or perhaps other phospholipids exert a similar modu- latory effect on other CaM-dependent en- zymes.
In summary, our findings suggest that phosphatase potentially has multiple reg- ulatory sites: one for CaM and two for PtdIns, one of high affinity and another of low affinity. Occupation of these sites by PtdIns modulates the response of the en- zyme to CaM, and the nature of response depends on whether one or two sites are oc- cupied and on the concentration of CaM and PtdIns. In addition, phosphatase has a separate and distinct regulatory site for subunit B (20). The possession of multiple regulatory sites adds another layer of com- plexity in understanding the regulatory properties of this CaM-dependent enzyme.
ACKNOWLEDGMENTS
We are grateful to Mary Margaret Dulaney and Su- zanne Wilmoth for preparation of calmodulin and calmodulin-dependent phosphatase.
5 S. Huang and W. Y. Cheung, unpublished experi- ment.

MODULATION OF CALCINEURIN PHOSPHATASE 49
REFERENCES
1. CHEUNG, W. Y. (1380) Science 207.19-n. 2. MANALAN, A. S., AND KLEE, C. B. (1984) Adv. Cy-
clic Nucleotide Protein Phosphor&&on Res. 18.227-278.
3. PICHARD, A., AND CHEIJNG, W. Y. (1977) J. Biol C’hem 252.4872-48’75.
4. WOLFF, D. J., AND BROSTROM, C. 0. (1976) Arch Biochem Biophys. 173,720-731.
5. NIGGLI, V., ADUNGAH, E. S., AND CARAFOLI, E. (1981) J. Bid Chem 256,8588-8592.
6. CARAFOLI, E. (1984) Fed Proc 43,3005-3010. 7. YEAGER, R. E., HEIDEMAN, W., OLWIN, B. B., KEL-
LER, C. H., SCHMIDT, J. W., SHATLUCK, R. L., AND STORM, D. R. (1985) .I. Neurochem. 44,818- 824.
8. POLITINO, M., AND KING, M. M. (1987) J. Bid Chem 262,10103-10113.
3. TALLANT, E. A., WALLACE, R. W., AND CHEUNG,
W. Y. (1988) in Methods in Enzymology (Means, A. R., and O’Malley, B. W., Eds.), Vol. 102, pp. 244-256, Academic Press, San Diego.
10. GOPALAKRISHNA, R., AND ANDERSON, W. B. (1982) Biochem, Biophys. Res. Commun 104,830~8.36.
11. KHANDELWAL, R. L., VANDENHEEDE, J. R., AND
KREBS, E. G. (1976) J. Bid Chem 251, 4850- 4858.
12. REIMANN, E. M., AND BEHAM, R. A. (1983) in Methods in Enzymology (Corbin, J. D., and Hardman, J. G., Eds.), Vol. 33, pp. 51-55, Aca- demic Press, San Diego.
13. TALLANT, E. A., ANIZI CHEUNG, W. Y. (1983) Bie chemistly 22,3630-3635.
14. LAPORTE, D. C., WIERMAN, B. M., AND STORM, D. R. (1980) Biochemistry 19,3814-3819.
15. TANAKA, T., AND HIDAKA, H. (1980) J. Biol Chem, 255,11078-11080.
16. TALLANT, E. A., AND CHEUNG, W. Y. (1984) Arch. Biochxm. Biophys. 232,269-279.
17. WINKLER, M. A., MERAT, D. L., TALLANT, E. A., HAWKINS, S., AND CHEUNG, W. Y. (1984) Proc Natl Ad Sci USA 81,3054-8058.
18. WANG, J. H., AND DESAI, R. (1977) J. Biol Chem 252,4175-4188.
19. KLEE, C. B., CROUCH, T. H., AND KRINKS, M. H. (1979) Proc Nat1 Acad Sci. USA 76,6270-6273.
20. MERAT, D. L., Hu, 2. Y., CARTER, T. E., AND CHEUNG, W. Y. (1985) J. Bid Chew. 260,11053- 11053.
21. JOHNSON, J. D. (1983) Biochem, Biophys Res. Commun. 112,787-793.
22. INGEBRITSEN, T. S., STEWART, A. A., AND COHEN, P. (1983) Eur. J. Biochem. 132,237-X)7.
23. PALLEN, C. J., VALENTINE, K. A., WANG, J. H., AND HOLLENBERG, M. D. (1985) Biochemistry 24, 4727-4730.


![Buparlisib, another step in the quest to cure ......PIP3 – Phosphatidylinositol 3,4,5 trisphosphate [PI(3,4,5)P3] PTEN – Phosphatase and tensin homologue RAEB – Refractory anemia](https://img.pdfslide.us/doc/110x75/5f3382fe83ae4a328035e0d8/buparlisib-another-step-in-the-quest-to-cure-pip3-a-phosphatidylinositol.jpg)


























