Embed Size (px)
Citation preview

DISTINGUISHING AMONG CAUSAL EXPLANATIONS IN BIOLOGY: MECHANICAL AND TOPOLOGICAL EXPLANATIONS
Philippe Huneman. IHPST (CNRS/ Université Paris I Sorbonne)

Models of explanation in philosophy of science
- Causal explanations (Salmon, 1984)- Unifying explanations (Friedman 1963,
Kitcher 1969)- The two models are rather two poles
(combinations are possible (eg Strevens 2004))

Often causal explanations mean : identify processes or mechanisms) displaying the phenomena. (Salmon 1984; Wimsatt 1982; Craver
Machamer Darden 2000, Craver 2007 In this view the role of counterfactuals is
downplayed
Very accurate concerning many aspects of neurosciences (Craver 2007), molecular biology (Darden 2003)

Here : in addition, define and consider « topological explanations »- Present in ecology, evolutionary biology,
social sciences, etc.- Do not pinpoint mechanisms- Influential with the rise of network science,
and the inflation of « neutral systems » (Hubbell, UNTB/ Stadler and Schuster, etc.)
- Are a kind of causal explanations in a counterfactual sense

OUTLINEI. Topological explanations: nature and role
I. The nature of topological explanationsII. The pervasiveness of topological explanations in
community ecologyIII. Topological vs mechanistic explanations
II. Mechanical and topological explanations reconsidered.- disjunct, combined
I. Disjunct explanations : the case of robustness in living systems
II. Neutral spaces, neutral networksIII. Combining mechanism and topological explanations
III. A general view of causal explanations in biology

TOPOLOGICAL EXPLANATIONS
Part I.

THE NATURE OF TOPOLOGICAL
EXPLANATIONS
I.A.
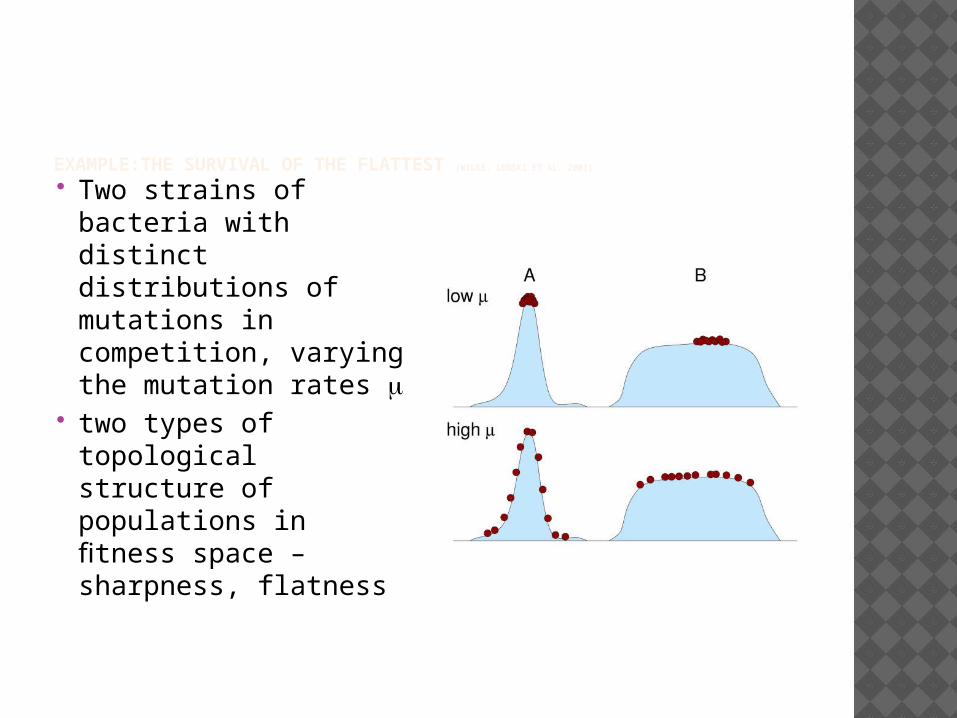
EXAMPLE:THE SURVIVAL OF THE FLATTEST (WILKE, LENSKI ET AL. 2001)
Two strains of bacteria with distinct distributions of mutations in competition, varying the mutation rates m
two types of topological structure of populations in fitness space – sharpness, flatness

The probability that a mutation displays a large drop in fitness as compared to the current alleles is clearly a consequence of the shape of the curve, because it only depends on the fitness distribution of possible mutations. And this probability is what determines the evolutionary fate of each strain

WHAT IS A TOPOLOGICAL EXPLANATION 1. Topological properties: System S, properties Ti, related to some
space: how parts of the system are located regarding one another, etc. The “parts” can be parts of a more abstract space:
Phase spaceCommunity ecology : parts are species/ cluster
of species in a (foodweb) graph

S has elements, parts, features or capacities, moments of its regular behavior, or of the set of its possible behaviors, which are likely to be represented in a graph, a network or a variety S’ in a space E.
S’ will have topological properties Ti, namely, properties which specify its invariance under some continuous transformations, and which will determine equivalence classes between all structures S* homotopic to S’. Or, if S is a graph, you can specify some properties of S’ (e.g. connexity; cyclicity, etc.) which will define an equivalence class, and distinguish S’ from other graphs S* not having those properties.

S has topological properties in virtue of its relation to S’ and its elements and relations
Eg. Lenski’s bacteria strains have the properties of “flatness” and “sharpness” in virtue of their essential relations to their mutations distribution.

Two consequences S realizes the topological properties Ti, and
many Sj can be said to be equivalent because they realize the same Ti (for example, belonging to the same equivalence class defined by homotopic paths in some space)
Ti have many consequences, especially, they may constrain the possible transformations of S’, for example because all continuous transformations should lead to some homotopic structure S’’.

2. Topological explanationan explanation in which a feature, a trait,
a property or an outcome X of a system S is explained by the fact that it possesses specific topological properties Ti.

TOPOLOGY AND PROCESSES The topological explanatory relation implies that
whatever possible process Bj occurs to S that involves some or all elements or parts of S, no Bj is sufficient to account for X, but the simple fact that S realizes Ti entails as a consequence the fact that S has X.
It may be that a given X causally results from some Bj, but that what explains X is not the Bj itself;
eg. causal process Bk also produces X, but that the topological properties Ti which constrain in the same way all processes Bi, entail that whatever the process Bj, the outcome will be of type X -> the causal process Bj itself does not make any difference (conditionally on the fact of Ti) to the outcome and is therefore not explanatory.

--> Explanation of X goes like a relation of entailment between topological properties Ti and X or features of X, and not like the
display of a mechanism from which X would be a temporal outcome.
Phrase « Topological explanations »: frequent in maths; in quantum physics (Arcos & Pereira 2007) – but about already mathematical entities (spin etc.)
Yet in biology, ecology etc. those are pervasive,

THE PERVASIVENESS OF TOPOLOGICAL EXPLANATIONS
IN COMMUNITY ECOLOGY
I.B.

ECOLOGY: THE DIVERSITY-STABILITY HYPOTHESIS« The more diverse a community is, the
more stable it is in its composition »Classical until the 70s
May 1974: mathematically it’s not the case
Thus: Need to define diversity (functional ? Number of species ? ) and stability (of composition ? Constancy of a biological property (eg biomass); persistence (no species get extinct); resilience

-> Various modes of the hypothesis:Tilman 1996 (constancy of biomass); etc.
Basically, May’s model assumed random connections between species; but this is not realistic.
Hence: exploring various consequences of specific connectivity schemes upon kinds of stability

NETWORKS OF CONNECTIONS APPROACHVariables: the number of species and
their average number of links,
the number of connections realized between species as compared to the number of possible connections (connectance),
the distribution of the connections between species.
Solé and Montoya 2001; Dunne et al. 2002; Mc Cann et al. 1998; Montoya and Solé 2006
Many ecological interactions (competition, predation, mutualism); some graphs are drawn without considering even the nature of the causal relation (Montoya and Solé 2006)

Network with 2 hubs and lots of poorly connected species (464).
Deleting one species: chances to delete only an isolated node (hence not changing the pattern) are 232 times higher than chances to alter the pattern
S : the communityS’: the network of interactionsT: property of having two hubs.T entails that the probability of
altering the pattern with one species extinction is very low-> stability

IMPORTANT NETWORKSScale-free networks : distribution of
connections varies according to a power law – hence, few big hubs, many slightly connected nodesProduction rule : preferential atachment (rich
get richer) (P(n) ~ n)
Small worlds (two nodes are always close, high level of clustering); Production rule – adding random links
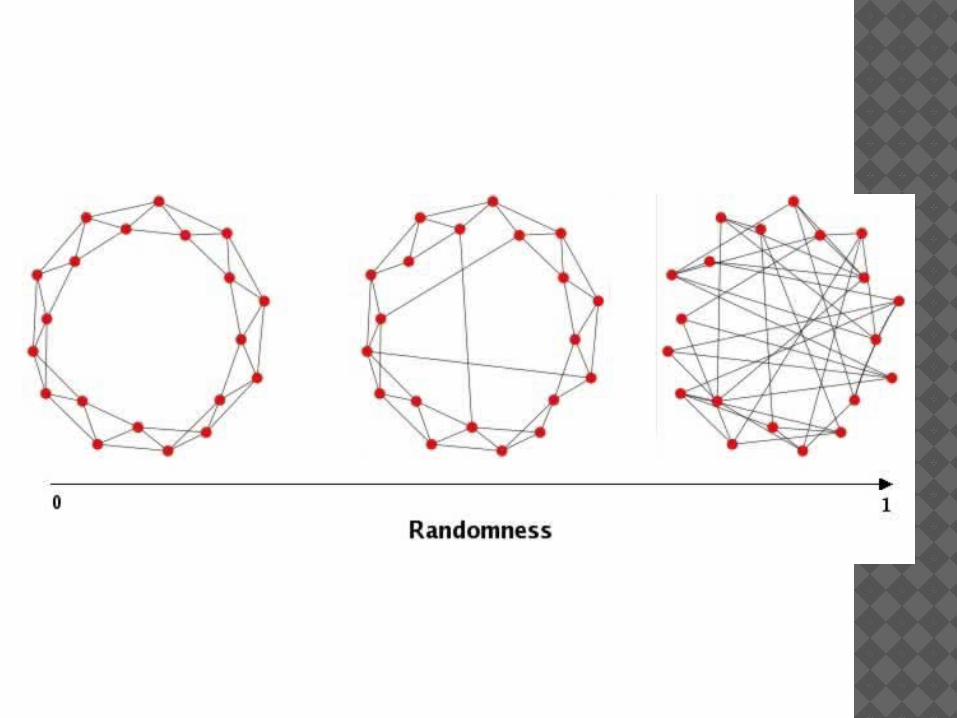

Scale free networks - deleting randomly a species has many chances to affect poorly connected species, hence the stability of the community through conservation of quite all the links Internet has the same property (Solé et al.
2001); also some financial systems (Levin et al. 2008)
Many ecological networks are often not exactly random but almost (truncated scale free networks) – hence the consequence of stability

Small world networks – disconnect two species A & B (by deleting an intermediary one between them ) then there exists a cluster where A is where there is a species closely connected to B.
S’ the interaction networkT: « Being a small work »Entails stability (robustness viz. Species
extinction)

These are topological explanations : they deduce the stability property from the topological property of the network of connections, without considering the dynamics, the causal interactions between species, etc. In both cases, if S1 has lots of preying species and
S2 lots of parasitic species but S’1=S’2, the same explanation for stability holds.
Stating the formal property – e.g., the relative number of small and large hubs – immediately yields the result, no matter what are all the temporal mechanisms which can happen in the community.

TOPOLOGICAL VS MECHANISTIC
EXPLANATIONS
I.C

TOPOLOGICAL AND MECHANISTIC EXPLANATIONS A mechanism is defined by entities and
activities (MDC 2000); here no specific activities and entities are identified.
For ex. in the « survival of the flattest » what’s explanatory is the topology of the mutations, not the mechanisms (the same in both cases)

COMMUNITY STABILITYTOPOLOGICAL EPXLANATIONS OF STABILITY CONTRAST WITH THE EXPLANATIONS WHICH WOULD IDENTIFY A RESTORATION MECHANISM
Mechanistic explanation Topological explanation
A ->B->C->D
A ->B- / DC extinct, B decreases
in frquency, D increases
Vacant niche appearsOther species C’
replaces CA->B-> C’ -> D
The food web is a scale free network
Probability that a random species extinction alters the community structure is therefore low

Mechanisms It i s in virtue of X’s y-ing,
that Y and Z will for example be triggered to do J and K, and then make the system S likely to be -f ing as a result of combined J and K.
The explanandum of a mechanistic explanation is determined by stating the “set-up” and “termination” conditions. The chronology of sequences through which entities act is crucial to explain the production of this termination condition
Topological explanationsThere are no specific activities
listed: no matter what the species do, whether they prey or not, whether they y or they f, and on whom etc. Stability rather occurs because of some network property of S’.
Another difference = the temporal sequence between events in topological explanations is irrelevant, since switching the order of
events does not affect the explanandum

Suppose that two systems S1 and S2 have the same associated shape S’ in abstract space but that the relata of links in S1 are J and K, and in S2 are J2 and K2, with their distinct associated activities. Yet the topological properties of S’, realized identically by S1 and S2, will not be affected by this difference of activities and properties; therefore the identification of mechanisms in S1 and S2 is not relevant for explaining their outcomes

MECHANICAL AND TOPOLOGICAL EXPLANATIONS RECONSIDERED.-
DISJUNCT, COMBINED
Part II.

DISJUNCT EXPLANATIONS : THE CASE OF ROBUSTNESS
IN LIVING SYSTEMS
2.A.

ROBUSTNESSA capacity to keep some parameters stable in the face of
changing variables. All modes of stability mentioned above in ecology satisfy
this characterization.
• Increasing researches about robustness in biology (Kitano 2005, de Visser 2002, Wagner Robustness and evolvability in living systems)
Two more specific characterizations: a. “level” definition, in the sense that change in low level
variables doesn’t involve change in high level variables – for instance changing the identity of several species does not affect some general properties like biomass or abundance pattern.
b. “functional“ definition: robust are the systems able to maintain (some of) their functions in the face of perturbations.

CLASSICAL KINDS OF ROBUSTNESS
Homeostasis (Cannon – Physiology) is an important kind of physiological robustness
« Canalisation » (Waddington) means robustness of development against genetic noise

IMAGES OF CANALISATION (WADDINGTON)
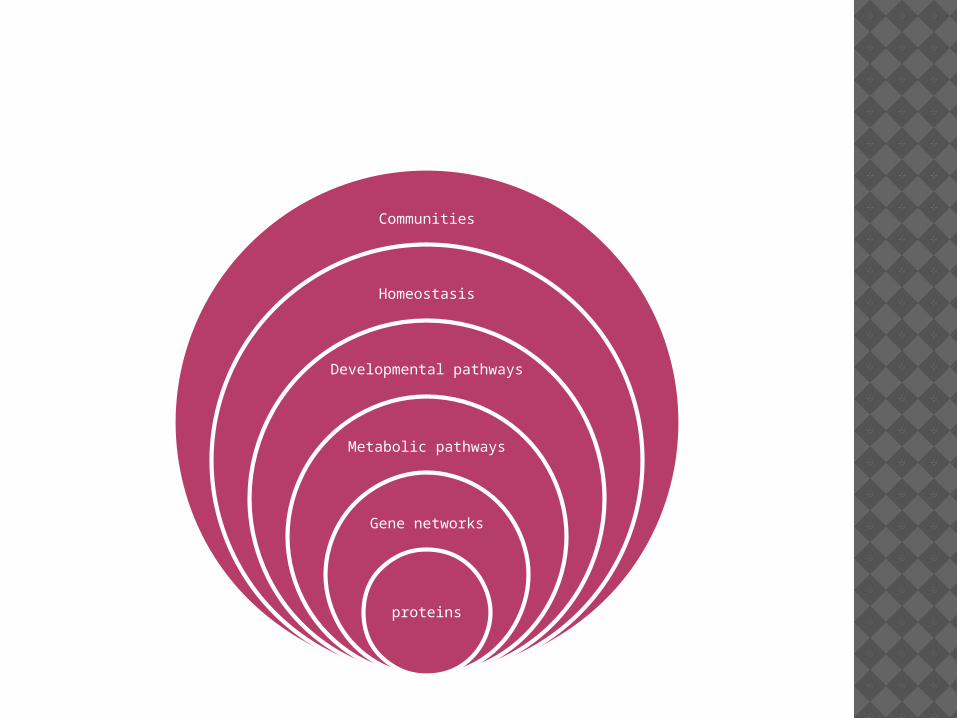
Communities
Homeostasis
Developmental pathways
Metabolic pathways
Gene networks
proteins

ROBUSTNESS OF GENETIC CODE Redundancy : many codons yield the same amino
acid-> often, changing A to C, G to T etc., does not
change the functional product
In general: Redundancy is an explanation of robustness (and also of evolvability)
Other kinds of robustness do not involve redundancy but « distributed robustness » (most of them, Wagner 2005b)
-> an example of distibuted robustness (at the gene level): gene regulation network of Endo16 (sea urchins)

2 TYPES OF ROBUSTNESS ISSUES a. proximate :
how is it ensured? (-> why does distributed robustness occurs?)?
About a .Many kinds of explanations; they can be partitioned into mechanical vs. topological explanations
b. evolutionary –b1), why has it
evolved? b2) what role may it
play in evolution
About b1. Articulating genetic and environmental robustness.(What has been selected for ?)

Mechanisms• Feedback control
(physiological homeostasis; lactose operon)
• Weak linkage (Gerhardt and Kirschner 2005)
• DNA repair systems (-> fctof sex (Michod and Bernstein 1996))
Topology• Modularity• Redundancy• Neutral spaces (Stadler,
Schuster et al.; Wagner 2005)
• Small world topologies• « aristocratic networks »
(laszlo barabasi)

DNA REPAIR The fact of diploidy and the mechanisms
of recombination are such that if DNA is altered, another copy is intact which can be a template for DNA repair. In this sense robustness of DNA is produced by this ability to copy on the basis of a second haplotype as a template.

MODULARITY(Modularity is a highly clustered network)
It reduces the effect of perturbations because those won’t break the whole systems, since they have chances to concentrate upon only one module, the others being likely to still function.
An explanation for evolvability (Altenberg and Wagner 1996)
ROBUSTNESS OF PROTEINS IN THE FACE OF RECOMBINATION.

WHY ARE TOPOLOGICAL EXPLANATIONS OF ROBUSTNESS PERVASIVE AT ALL LEVELS ? Many topological properties : defined by
invariance regarding to deformation Stability : reaction viz. perturbation; in
the S’ associated to system S , a « perturbation » is a « deformation »
Therefore, topological equivalence classes (ie shapes S* which are invariant trough continuous deformation) determine kinds of stabilities
Hence topological properties are often related to robustness properties

NEUTRAL SPACES, NEUTRAL NETWORKS
2.B
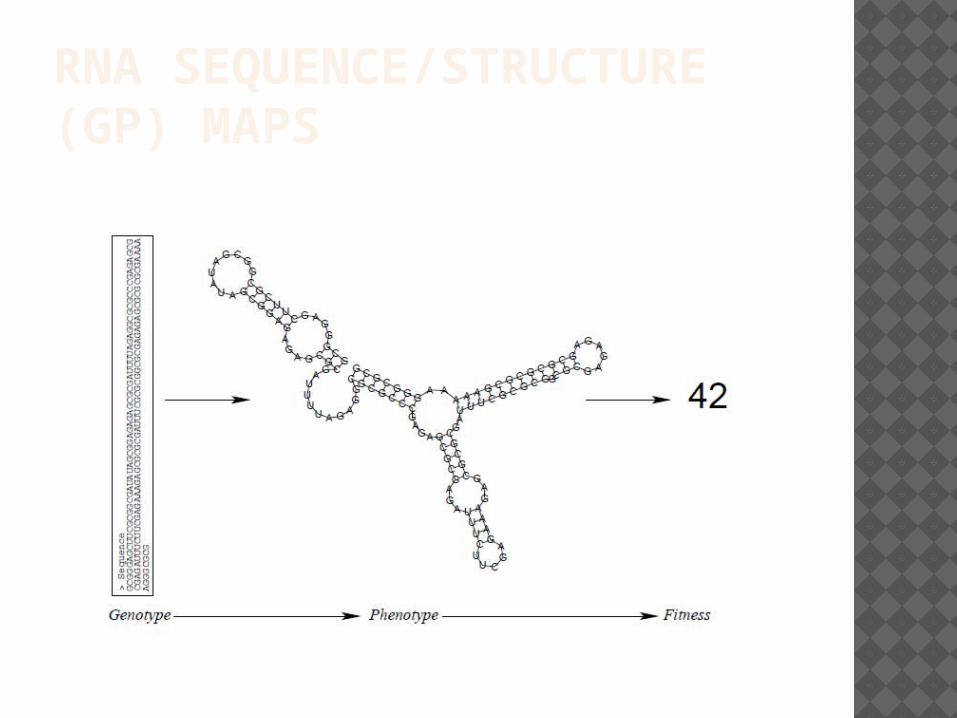
RNA SEQUENCE/STRUCTURE (GP) MAPS

Many different sequences, due to the laws of chemistries and the requirements of minimizing free energy, fold into the same spatial structure (ie, with the same molecules at surface sites, hence functionally behaving similarly).
The space of structures is such that few structures are reached by several large sets of RNA sequences, whereas many structures are reached only by a few sequences.

« NEUTRAL NETWORKS » Defining a distance between two sequences
G’G’ as the number of mutations needed to get from G to G’
Defining the set G1 of sequences such that f(H) = f (G) and d (G, H) = 1; same operation with each H in G1 etc.
Gives a neutral network – all Hs in it are accessible from G by a series of mutations without changing the phenotype
Stadler and Stadler, 2001; Otalloni and Stadler, 2007; Van Nimwegen, Huynen and Crutchfield, 1996; Stadler, Fontana and Schuster 2001, Fontana et al., 1992
Wagner 2005 Gavrilets 2004 (neutral tunnels)


Explains mutational robustness: if G is deep in a neutral network, mutations will not alter the phenotypes
Explains also evolution from low to high fitness with drift (through a neutral network)
Genetic robustness of some genotypes is the topological structure of the GP map, and the specific position of the genotype x in this structure. Once again, no mechanism of gene-protein interaction has to be hypothesized in order to understand such robustness.
And those neutral spaces allow biologists to formulate issues about the evolution of robustness (the more robust is a system, the larger are the neutral spaces in it) and the roles of robustness in evolution.

Neutral spaces are also likely to explain : the robustness of the genetic code (by devising a
space of alternative genetic codes (Wagner, 2005, 198),
as well as the robustness of gene expression (like in the case of Endo 16). This latter case is easily described by a space of DNA,
which maps each regulatory DNA regions onto a specific expression. Some genes like Endo 16 will be regulated by many alternative regulatory regions, and one can define the set A of regulatory DNA regions expressing the same gene. Very likely, A can be partitioned into several neutral spaces, and a very robust gene (regarding its expression) like Endo 16, will have a large neutral regulatory space associated.

COMBINING MECHANISM AND TOPOLOGICAL
EXPLANATIONS
2.C.

1. THE CONTINUUM VIEWNot any relation is relevant in topological
explanations (in the graphs)In ecological explanations here : consider
only ecological interactions (preying, parasiting etc); not « bigger than », etc.
The topological explanations take a subset of all relations and consider them as relevant, abstracting away from their causal nature;
The mechanistic explanation considers the causal nature of these causal relations (but sometimes equate them)

2. ARTICULATING TOPOLOGY AND EXPLANATIONS AS ANSWERING DIFFERENT QUESTIONS
1. Why has this ecosystem stability properties ? Because S’ is a scale free network.
topological explanation2. « Why is S’ a scale free network ? » Because
preying is built up according to a preferential attachemnt rule .
mechanistic explanation3. « Why are many ecosystems having scale
free foodwebs? » -> general properties of the preying and parasitism relations
causal, possibly mechanistic, explanation

A complete explanation (here: of stability of ecosystems) as a sequential explanation (involving mechanisms and topologies).

3. TWO TYPES OF TOPOLOGICAL EXPLANATIONS Pure Topological Explanations
(examples given)
Constraining Topological explanations – there are causal mechanisms but the topological properties are constraining their action and therefore the explanations
Example:Evolution-of-cooperation research (Nowak 2005;
Pacheco and Santos 2006)
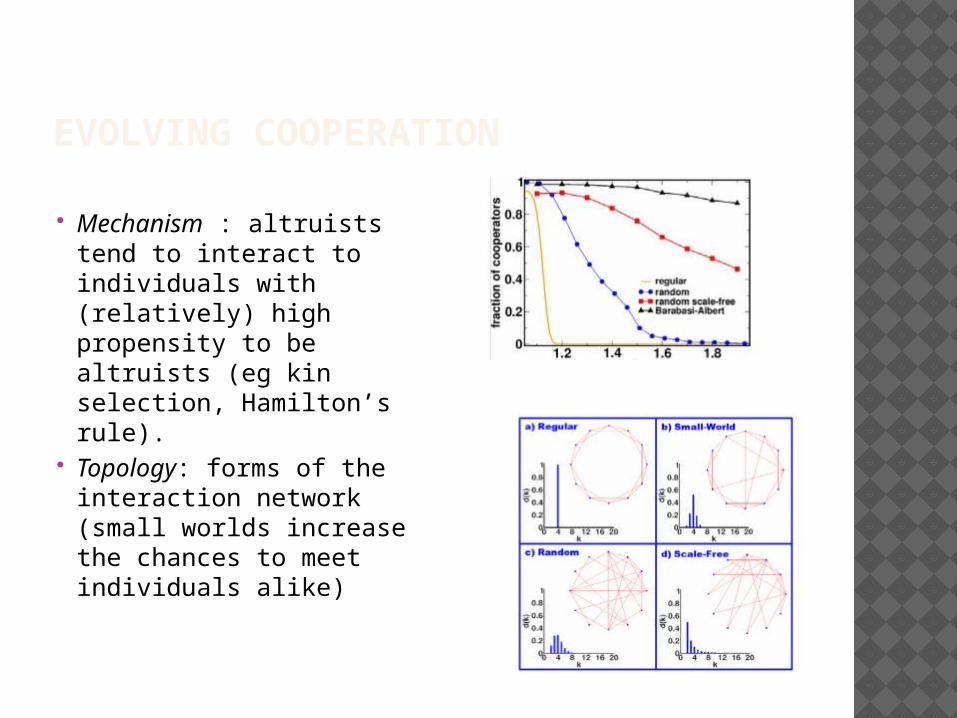
EVOLVING COOPERATION
Mechanism : altruists tend to interact to individuals with (relatively) high propensity to be altruists (eg kin selection, Hamilton’s rule).
Topology: forms of the interaction network (small worlds increase the chances to meet individuals alike)
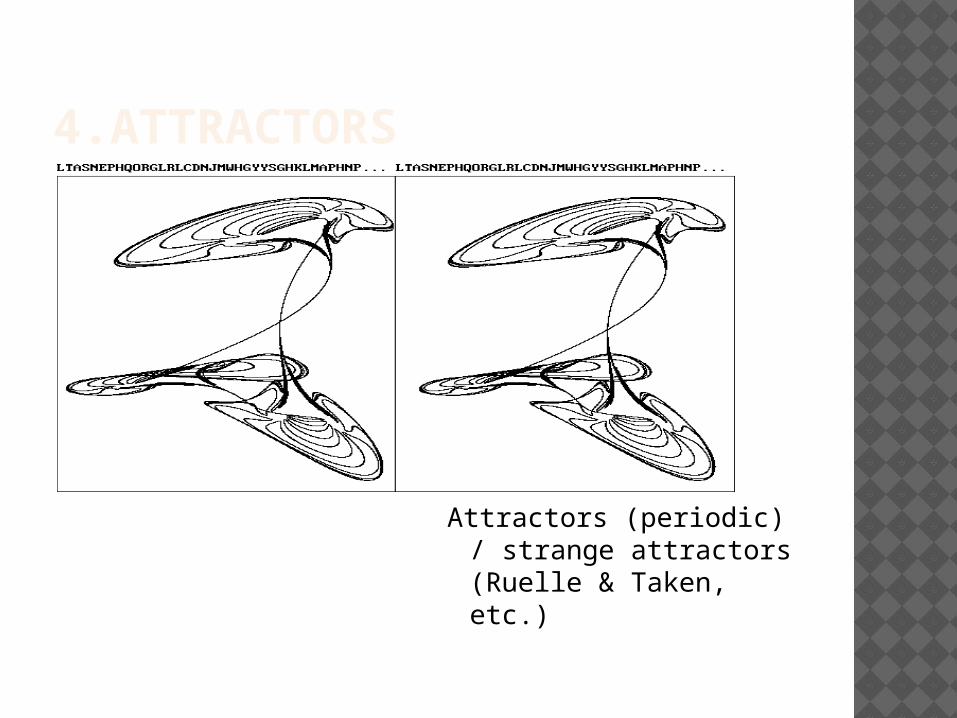
4.ATTRACTORS
Attractors (periodic) / strange attractors (Ruelle & Taken, etc.)

Once attractors are found in the phase space
Then topological properties of the attractor may explain some of the trajectories
Suppose a state space of a system with an attractor. Given only its topological properties one can deduce many things: the time before getting into chaotic regime, the chances of ending up in one rather than in another, the possible bifurcations (Feigenbaum 1978), etc. So those explanations do not consider the mechanical processes proper to the system.
-> Is it a pure topological explanation ?

No : such explanation elaborates on mechanical explanations and causal knowledge used to construct the phase space and the attractor
In a second step, then, studying the topology of the attractor may support conclusions about new properties of the trajectories of the system
So attractor studies define another kind of explanation, which somehow mixes topological and mechanical kinds of explanations (it’s a « secondary topological explanation »?)

5. COMBINING MECHANISMS AND TOPOLOGICAL EXPLANATIONS IN INTERLEVEL STUDIES
- Mechanistic explanations: Intralevel : causal relationsInterlevel : constitutive relations (Craver 2007)
“there is a temptation to say that the activation of cyclic GMP phosphodiesterase, which catalyzes the conversion of cyclic GMP to 5c/-GMP, causes rod cells to hyperpolarize, which in turn causes the eye to transduce light into neural activity. But the activation of cyclic GMP phosphodiesterase is part of the activity of depolarization, which is part of the eye’s transduction of light” (2006, p. 15)
S system; Mi subsystems; Nj, parts of subsystems Mi, causing Mi’s outcome. But they contribute to constitute S’s activity.
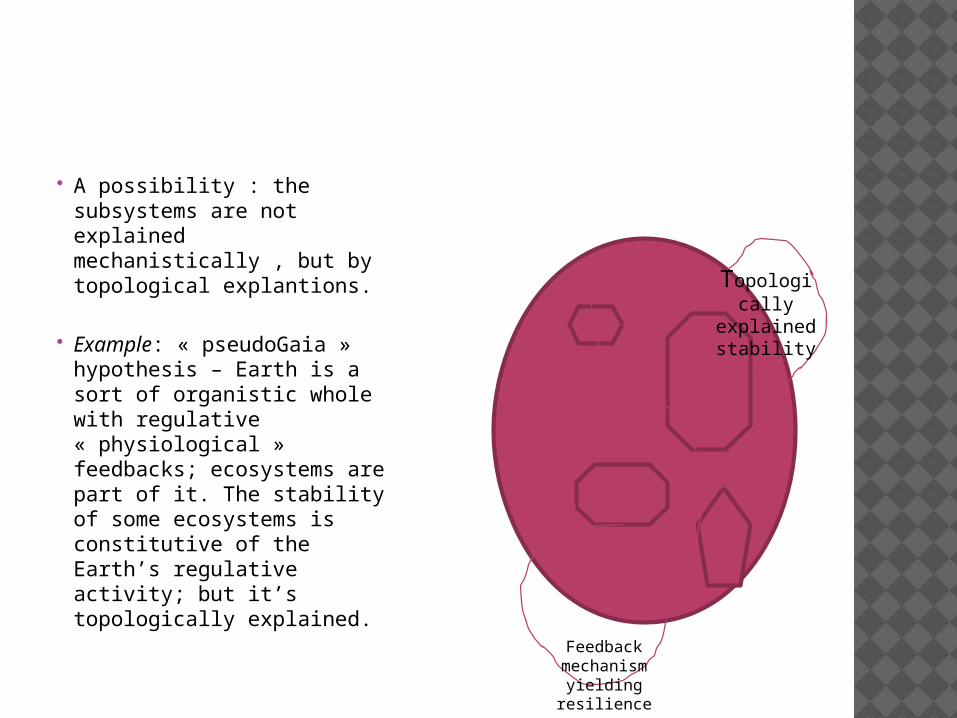
A possibility : the subsystems are not explained mechanistically , but by topological explantions.
Example: « pseudoGaia » hypothesis – Earth is a sort of organistic whole with regulative « physiological » feedbacks; ecosystems are part of it. The stability of some ecosystems is constitutive of the Earth’s regulative activity; but it’s topologically explained.
Topologically
explained stability
Feedback mechanism
yielding resilience

CAUSAL EXPLANATIONS IN BIOLOGY – DIVERSIFYING
THE PICTURE
Part III

Displaying Mechanisms : you don’t need to consider counterfactuals relations (what would hold if part A is not here etc.)
The display of the mechanism is explanatory.
Topological explanations : you abstract away from mechanisms.
The causal nature of interaction is irrelevant (predation and parasitism may fall into the same equivalence class)
Question : is it still a causal explanation ?

S, system, S’ associated abstract shape, P topological properties, X property/behaviour/outcome to be explained.
« If S had not P (in virtue of S’) the outcome/behaviour X of S would not be what it is »
-> topological explanations involve counterfactual dependencies (whose relata partly belong to the abstract space E associated to S)

In life sciences causal explanations are:
a. Displaying mechanisms
b. Topological explanations – counterfactual
c. Mixed forms of both. (Constrained topological explanations, sequence of explanations, attractor explanations, etc.)