Embed Size (px)
Citation preview

APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY
Phenotypic characterization, virulence, and immunogenicityof Edwardsiella tarda LSE40 aroA mutant
Zhao-Lan Mo & Jie Li & Gui-Yang Li & Peng Xiao
Received: 17 December 2012 /Revised: 15 February 2013 /Accepted: 24 February 2013 /Published online: 17 March 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Bacterial aro mutants are frequently used as liveattenuated vaccines for domestic animals. In this study, wecharacterized Edwardsiella tarda strain LSE40 with a deletionin the aroA gene. In addition to autotrophy, the aroA mutantappeared to have delayed cell division and reductions in itsswarming motility, biofilm formation, and production oftranslocator proteins in the type III secretory system. Themutant exhibited high virulence attenuation in turbot fish,Scophthalmus maximus (L.), where the 50 % lethal doseincreased by more than 3 log10 via intraperitoneal (i.p.) injec-tion and by >2 log10 via immersion exposure compared withthe wild-type parent strain. A tissue persistence study showedthat the mutant retained the ability to invade and spread inturbot and viable cells could be detected up to 28 days after i.p.infection and 21 days after immersion exposure. These resultssuggested a pleiotropic role for aroA in the physiologicalbehavior of E. tarda. Turbot exhibited a good humoral re-sponse and the enhanced expression of innate immune factors,interleukin 1β and lysozyme, when vaccinated with aroAmutant at 105CFU via i.p. injection and at 108CFU via im-mersion exposure. However, the aroA mutant did not provideeffective protection for turbot against edwardsiellosis follow-ing i.p. vaccination at doses of 104–106CFU or immersionvaccination at doses of 106–108CFUml−1. We hypothesized
that the aroA mutant did not trigger an appropriate T cell-immune response in turbot against infection of E. tarda.
Keywords Edwardsiella tarda . aroA . Phenotype . Proteinsecretion . Virulence . Immunogenecity
Introduction
Edwardsiella tarda is an important pathogen with a broadhost range, including, fish, amphibian, birds, and humans.Fish infected with this bacterium develop edwardsiellosis,which is an enterohemorrhagic septicemic disease that canlead to high stock mortality in fresh and marine fish (Mohantyand Sahoo 2007). The pathogenesis of E. tarda is multifacto-rial, and many virulence determinants associated with infec-tion of this bacterium have been described, such as thecapacity to adhere to cell surfaces (Rao et al. 2003; Sakai etal. 2003, 2009) and to invade host cells (Wang et al. 2010),hemolysins (Hirono et al. 1997), chondroitinase (Cook andTappe 1985), serum resistance (Mathew et al. 2001), biofilmformation (Zhang et al. 2008), and effector proteins deliveredvia the type III or VI secretory systems (T3SS or T6SS; Xie etal. 2010; Jobichen et al. 2010).E. tardamutants with deletionsin important virulence factors are highly attenuated, and theyhave been evaluated as potential live vaccines againstedwardsiellosis (Mo et al. 2007; Xiao et al. 2011; Choi andKim 2011).
Bacteria use a common pathway (the shikimate pathway) tobiosynthesize aromatic amino acids as well as folate, ubiqui-none, and/or menaquinone and catecholate siderophores(Pittard 1996). Mutants with deletion in aro genes that makeup this pathway are auxotrophic for aromatic amino acids, andthis is usually considered to be the main reason for virulenceattenuation because of their inability to synthesize aromaticamino acids that are required for growth and replication in
Electronic supplementary material The online version of this article(doi:10.1007/s00253-013-4813-1) contains supplementary material,which is available to authorized users.
Z.-L. Mo (*) : J. Li :G.-Y. LiYellow Sea Fisheries Research Institute, Chinese Academyof Fishery Sciences, 106 Nanjing Road,Qingdao 266071, People’s Republic of Chinae-mail: [email protected]
P. XiaoKey Lab of Experimental Marine Biology, Institute of Oceanology,Chinese Academy of Sciences, 7 Nanhai Road,Qingdao 266071, People’s Republic of China
Appl Microbiol Biotechnol (2013) 97:6325–6335DOI 10.1007/s00253-013-4813-1

hosts. aro mutants are frequently used as live vaccine candi-dates for domestic animals and humans. The most commontargeted gene is aroA, encoding the first enzyme of the com-mon pathway, 5-enolpyruvylshikimate-3-phosphate synthase(Pittard 1996). aroA mutants have been proven to be effectivelive attenuated vaccine strains against fish diseases (Vaughanet al. 1993; Marsden et al. 1996; Hernanz Moral et al. 1998;Thune et al. 1999). However, aro mutants have also beendescribed with defects in their flagella (Bar-Tana et al. 1980),proton translocation proteins (Rychlik et al. 2000), cell walland outer membrane integrity (Sebkova et al. 2008), and theproduction of capsular polysaccharides and fimbriae (Malcovaet al. 2009), which might contribute to their attenuation. Inaddition, not all aro mutants are effective as live vaccinesbecause they are incapable of growing and persisting in thehost for a period or triggering an immune response in theirhosts (O’Callaghan et al. 1988; Trebichavsky et al. 2006). Forexample, Salmonella enterica serotype typhimurium witharoA deletion was attenuated, but it did not trigger an innateimmune response or cytokine production (Trebichavsky et al.2006). Similarly, a preliminary study performed in our labora-tory showed that an attenuated E. tarda aroA mutant couldinduce innate and humoral responses in turbot, Scophthalmusmaximus (L.), but did not provide protection againstedwardsiellosis. These findings suggested that in addition toautotrophy, these aromutants might have defective physiolog-ical functions, which may mean they fail to provide effectiveprotection via appropriate immune response activation. Toclarify the properties of aroA in E. tarda, we constructed anaroA mutant and characterized its physiological phenotype,virulence, and immune efficacy in turbot.
Materials and method
Bacterial strains and growth conditions
E. tarda strain LSE40 (CGMCC no. 7199) isolated fromdiseased turbot was used in this study (Mo et al. 2007).E. tarda strains were grown in tryptic soy broth (TSB, Difco),on tryptic soy agar (TSA), or in Dulbecco’s modified Eagle’smedium (DMEM; Gibco) with 5 % CO2 at 25 °C. Escherichiacoli strains were grown in Luria–Bertani (LB; MDBio, Inc.,Taiwan) broth or on LB agar plates at 37 °C. A definedminimalmedium (DMM) was developed for E. tarda, which was basedon a previously described method, with some modifications(Lisa and Ronald 1996). The DMMmedium was composed ofM9 salts (Sambrook et al. 1989), 0.2 mgml−1 CaCl2, 0.005 mgml−1 FeSO4⋅7H2O, 0.24 mgml−1 MgSO4⋅7H2O, 9.0 mgml−1
glucose, 0.11 mgml−1 DL-pyruvic acid, 0.004 mgml−1 L-pyri-doxamine, 0.084 mgml−1 L-arginine, 0.1 mgml−1 L-glutamine,and 0.146 mgml−1 L-lysine. The “aromix” consisted of 1 mgml−1 each of tyrosine, tryptophan, and phenylalanine, which
was added to DMM to test the growth of the aroA mutant.When required, appropriate antibiotics were supplemented asfollows: colistin, 12.5 μgml−1; chloramphenicol, 34 μgml−1.
Fish
Turbot (S. maximus L.) were purchased from local commer-cial fish farms in Shandong province, China. Fish weremaintained in 500-L aquaria with circulating seawater at16–18 °C. Prior to the infection and vaccination experi-ments, sera were taken from five random fish and testedby slide agglutination against formalin-inactivated E. tarda.In addition, the internal organs (liver, spleen, and kidney)were harvested from the fish samples and tested by bacteri-ological analysis. Only those fish that were negative in thesera agglutination test and bacterial recovery were used inthe later experiments.
Construction of the aroA mutant
A non-polar deletion of aroA gene was generated in E. tardaLSE40 using a published method (Mo et al. 2007). In brief,two PCR fragments were generated from LSE40 genomicDNA using the primer pairs aroA-for (5′-AAGGTACCAGGTGCTGGCGGAAAATGACTC-3′) and aroA-int-rev( 5 ′ - AACTGGCTAAAGTAGTCGGGCAACGTCATGCTAAATCACTCCTGCTG-3′), and aroA-int-for (5′-ACGTTGCCCGACTACTTTAGCCAGTTTTCCGCCCTGTGCCATCCG-3 ′ ) a nd a r oA - r ev ( 5 ′ -AAGGTACCCGTGCTGACATCCTCCCCCTCT-3′). Theresulting 593-bp fragment contained the upstream regionof aroA, while the 628-bp fragment contained the down-stream region of aroA. A 26-bp overlap in the sequences(underlined) permitted amplification of a 1,221-bp productduring a second PCR with the primers aroA-for and aroA-rev, both of which introduced a KpnI restriction site (bolditalic). The resulting PCR product, which contained a dele-tion from nucleotides 4–1,236 in aroA, was ligated into thesuicide vector pRE112 (Cmr) and transformed into E. coliSM101 pir. Single crossover mutants were obtained byconjugal transfer of the resulting plasmid into E. tardaLSE40. The second crossover mutants were screened using10 % sucrose TSA plates. The mutation of aroAwas furtherconfirmed by PCR using the primers aroA-for and aroA-revand sequence analysis of the PCR product.
In vitro characterization of the aroA mutant
The physiological phenotype of the aroA mutant was char-acterized, including its cell morphology, growth, swarmingmotility, biofilm formation, auto-aggregation, and T3SStranslocator protein production and secretion. Briefly, thecell morphology was observed by scanning electron
6326 Appl Microbiol Biotechnol (2013) 97:6325–6335

microscopy using cultures incubated in TSB for 24 h. Thebacterial growth levels during incubation with DMM,DMEM, and TSB at 25 °C with or without “aromix” addi-tion were recorded at 48 h. The swarming diameter wasmeasured after incubation of the bacterial strains on 0.5 %(w/v) soft TSA plates at 25 °C for 24 h. Biofilm formationwas assayed using the crystal violet stain method, as de-scribed previously (Fletcher 1977). Auto-aggregation wasdetermined by culturing the bacteria in DMEM with 5 %CO2 at 25 °C for 24 h (Rao et al. 2004), followed byextraction of intracellular proteins (ICPs) and extracellularproteins (ECPs) to examine the levels of EseB, EseC, andEseD by Western blotting, according to a published method(Wang et al. 2009).
Attenuation experiments
Healthy turbot (average weigh, 10 g) were infected viainjection, immersion, or oral administration. For infectionvia injection, groups of ten fish were injected intraperitone-ally (i.p.) with 0.1 ml of the wild-type LSE40 or the aroAmutant suspended in phosphate-buffered saline (PBS,pH 7.4) containing 102–108CFU; the corresponding controlgroups were injected with 0.1 ml of PBS. For infection viaimmersion, groups of ten fish were exposed to seawatercontaining 106–108CFUml−1 of the LSE40 or the aroAmutant for 30 min; the corresponding control group wasexposed to seawater without bacteria. For infection via theoral route, groups of ten fish were treated with doses of0.2 ml PBS containing ∼109CFU of the LSE40 or the aroAmutant via a 26-gauge syringe, which was passed into thestomach of each fish; the corresponding control group wasinfused with 0.2 ml of PBS in each fish. All groups wereobserved for 14 days and any dead or moribund fish wasremoved for routine examination by bacterial recovery fromthe internal organs on TSA plates. The LD50 values werecalculated using the statistical approach described by Reedand Muench (1938).
Persistence of the aroA mutant
The persistence of the aroA mutant was determined in turbotinfected by the i.p., immersion, or oral route. Groups of 30turbot (average weight, 10 g) were i.p.-infected with thearoA mutant at a dose of ∼105CFU/fish, immersed with adose of ∼108CFUml−1 for 30 min, or orally treated with adose of ∼108CFU/fish. After infection, kidneys were sam-pled from five random fish in each group at 7-day intervals,followed by weighing, homogenization in PBS, and enu-meration of the viable bacterial counts by plating the seriallydiluted homogeneous suspensions onto the TSA plates.PCR with primers aroA-for and aroA-rev was performedfor identification of the colonies as aroA mutant.
Vaccination trials
Fish were vaccinated with the aroA mutant via i.p. injectionor immersion exposure. For i.p. vaccination, two trials wereperformed with two sizes of turbot. In the first trial, threegroups of 80 turbot (average weight, 5 g) were treated with0.1 ml of the aroA mutant at serial doses of 104–106
CFU/fish. In the second trial, one group of 80 turbot (aver-age weight, 40 g) was treated with 0.1 ml of the aroAmutantat ∼106CFU/fish. The corresponding control groups weretreated with 0.1 ml PBS. Immersion exposure wasperformed in parallel using both sizes of fish. In the firsttrial, three corresponding fish groups were treated withseawater containing 106–108CFUml−1 of the aroA mutantfor 30 min. In the second trial, one corresponding fish groupwas treated with seawater containing ∼108CFUml−1. Thecorresponding control groups were treated with seawatercontaining only PBS.
After vaccination, sera were collected from five randomcontrol and vaccinated fish by puncturing the caudal bloodvessels at different time points. The agglutination titers wereassayed using indirect enzyme-linked immunosorbent assay(ELISA) as described previously (Xiao et al. 2007). At 30 dayspost-vaccination, the vaccinated and control fish groups werechallenged with wild-type LSE40 via i.p. or immersion. Forthe 5 g turbot vaccinated by i.p., 20 fish from each group werechallenged i.p. with ∼104CFU/fish of LSE40. The 40 g fishvaccinated by i.p. were divided into two subgroups and 20 fishfrom each were challenged i.p. with ∼104CFU/fish of LSE40or by immersion exposure to ∼108CFUml−1 of LSE40. Forthe 5 g turbot vaccinated by immersion exposure, 20 fish fromeach group was challenged by immersion in ∼108CFUml−1 ofLSE40. The 40 g fish vaccinated by immersion were dividedinto two subgroups and 20 fish from each were challenged i.p.with ∼104CFU/fish of LSE40 or by immersion with ∼108
CFUml−1 of LSE40. The mortalities of each group wererecorded every 24 h after infection; dead fish was used to testbacterial recovery to confirm E. tarda as the cause of death.All of the test groups were observed for 4 weeks.
Expression of immune-related genes
The 40 g turbot vaccinated by i.p. or immersion, and thecorresponding control fish, were tested to determine the ex-pression of immune-related genes, i.e., interleukin 1β (IL-1β),lysozyme, and major histocompatibility complex classes Iα(MHCIα) and IIα (MHCIIα). The spleens were sampledrandomly from five fish in each group at 24 h, 1 week, and4 weeks after vaccination. The total RNAwas extracted usingTRIzol (Invitrogen, Carlsbad, CA). One microgram of totalRNAwas used for cDNA synthesis with Superscript II reversetranscriptase (Invitrogen). Quantitative real-time reverse tran-scriptase PCR (qRT-PCR) was performed using an ABI 7300
Appl Microbiol Biotechnol (2013) 97:6325–6335 6327

Real-Time Detection System (Applied Biosystems, FosterCity, CA) with SYBR ExScript II qRT-PCR Kit (Takara,China). The primer sequences are presented in Electronicsupplementary material (ESM) Table S1. The comparativethreshold cycle method was used to analyze the mRNA level.The expression levels of genes were determined in triplicateusing the 2−ΔΔC
T method with β-actin RNA as the control(Livak and Schmittgen 2001). All data were expressed in termsof the relative mRNA by normalizing the mRNA levels of thevaccinated fish against those of the control fish.
Statistical analysis
The data obtained from the analyses of bacterial growth,swarming diameter, biofilm formation, agglutination titers,and qRT-PCR were expressed as X±SD, and we tested forstatistical differences between the wild-type strain and aroAmutant, and differences between the experimental and con-trol groups, using Student’s t test. Data were consideredstatistically significant when P<0.05.
Results
Construction of the E. tarda LSE40 aroA mutant
An in-frame deletion aroA mutant was constructed using E.tarda LSE40 by a double-selection strategy of allelic ex-change mutagenesis. The mutant was screened to determineits inability to grow on TSA plates supplemented withcolistin and chloramphenicol, which was verified by PCRusing the primers aroA-for and aroA-rev. The aroA mutantgenerated an approximately 1,221-bp fragment, whereas thewild-type LSE40 generated a 2,454-bp fragment (Fig. 1).The PCR product was sequenced and it was found to alignwith the predicted sequence.
Growth of the aroA mutant in culture medium
The growth of the aroA mutant and the wild-type LSE40was tested using different media. As expected, the aroAmutant was unable to grow in the DMM medium withoutaromatic amino acids. However, only partial growth of thearoA mutant was observed in DMM when all aromaticamino acids were provided compared with the wild-typeLSE40 (P<0.05). When cultured in DMEM, the growth ofthe aroA mutant was significantly lower than LSE40 in theabsence of aromatic amino acids (P<0.05); when aromaticamino acids were provided, the growth of the aroA mutantwas slightly, but not significantly (P>0.05), lower than thatof LSE40. When cultured in TSB, the aroA mutantexhibited slightly, but not significantly (P>0.05), lessgrowth than LSE40 (Fig. 2).
Cell morphology of the aroA mutant
The cell morphology of the aroA mutant and wild-typeLSE40 was observed using scanning electron microscopy.Compared with LSE40, the aroA mutant had lost its shortrod shape and mainly assumed a slightly elongated shape.Most of the elongated cells were formed of two cells, whichappeared to be in the process of cell division (Fig. 3).
Biofilm formation and swarming motility of the aroA mutant
Fig. 1 PCR identification of E. tarda LSE40 aroA mutant. DNA fromE. tarda LSE40 aroA mutant and wild-type strain LSE40 was used astemplate in PCR with primer pairs of aroA-for and aroA-rev. PCRproducts were separated on 1.0 % agarose gel. Marker represented theTrans2K plus II DNA marker (TransGen Biotech Co., China)
Fig. 2 Growth of E. tarda under different media. Wild-type E. tardaLSE40 and aroA mutant were cultivated in DMM, DMEM, and TSBwith or without the addition of “aromix” consisting of 1 mgml−1 eachof tyrosine, tryptophan, and phenylalanine. Bacterial counts were de-termined on TSA plates. Each experiment was repeated in triplicate.Data were expressed as X±SD. Asterisks indicated significant differ-ences between the wild-type strain and the aroA mutant. *P<0.05
6328 Appl Microbiol Biotechnol (2013) 97:6325–6335
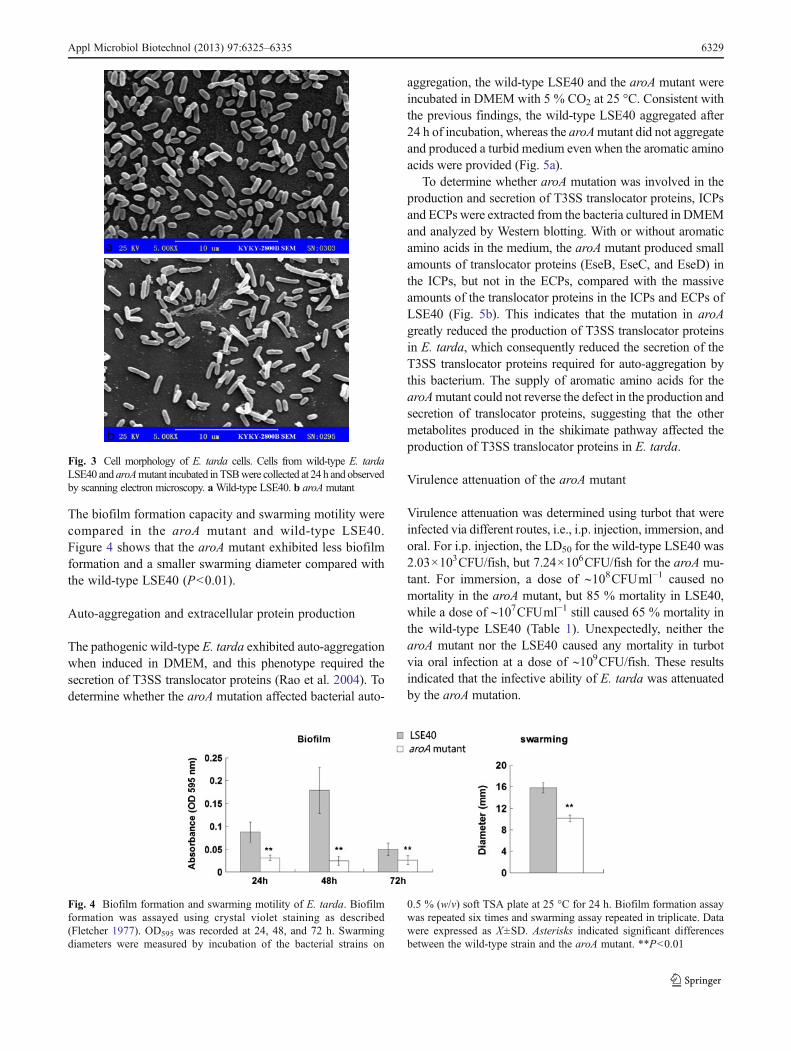
The biofilm formation capacity and swarming motility werecompared in the aroA mutant and wild-type LSE40.Figure 4 shows that the aroA mutant exhibited less biofilmformation and a smaller swarming diameter compared withthe wild-type LSE40 (P<0.01).
Auto-aggregation and extracellular protein production
The pathogenic wild-type E. tarda exhibited auto-aggregationwhen induced in DMEM, and this phenotype required thesecretion of T3SS translocator proteins (Rao et al. 2004). Todetermine whether the aroA mutation affected bacterial auto-
aggregation, the wild-type LSE40 and the aroA mutant wereincubated in DMEM with 5 % CO2 at 25 °C. Consistent withthe previous findings, the wild-type LSE40 aggregated after24 h of incubation, whereas the aroAmutant did not aggregateand produced a turbid medium even when the aromatic aminoacids were provided (Fig. 5a).
To determine whether aroA mutation was involved in theproduction and secretion of T3SS translocator proteins, ICPsand ECPs were extracted from the bacteria cultured in DMEMand analyzed by Western blotting. With or without aromaticamino acids in the medium, the aroA mutant produced smallamounts of translocator proteins (EseB, EseC, and EseD) inthe ICPs, but not in the ECPs, compared with the massiveamounts of the translocator proteins in the ICPs and ECPs ofLSE40 (Fig. 5b). This indicates that the mutation in aroAgreatly reduced the production of T3SS translocator proteinsin E. tarda, which consequently reduced the secretion of theT3SS translocator proteins required for auto-aggregation bythis bacterium. The supply of aromatic amino acids for thearoAmutant could not reverse the defect in the production andsecretion of translocator proteins, suggesting that the othermetabolites produced in the shikimate pathway affected theproduction of T3SS translocator proteins in E. tarda.
Virulence attenuation of the aroA mutant
Virulence attenuation was determined using turbot that wereinfected via different routes, i.e., i.p. injection, immersion, andoral. For i.p. injection, the LD50 for the wild-type LSE40 was2.03×103CFU/fish, but 7.24×106CFU/fish for the aroA mu-tant. For immersion, a dose of ∼108CFUml−1 caused nomortality in the aroA mutant, but 85 % mortality in LSE40,while a dose of ∼107CFUml−1 still caused 65 % mortality inthe wild-type LSE40 (Table 1). Unexpectedly, neither thearoA mutant nor the LSE40 caused any mortality in turbotvia oral infection at a dose of ∼109CFU/fish. These resultsindicated that the infective ability of E. tarda was attenuatedby the aroA mutation.
Fig. 3 Cell morphology of E. tarda cells. Cells from wild-type E. tardaLSE40 and aroAmutant incubated in TSBwere collected at 24 h and observedby scanning electron microscopy. aWild-type LSE40. b aroAmutant
Fig. 4 Biofilm formation and swarming motility of E. tarda. Biofilmformation was assayed using crystal violet staining as described(Fletcher 1977). OD595 was recorded at 24, 48, and 72 h. Swarmingdiameters were measured by incubation of the bacterial strains on
0.5 % (w/v) soft TSA plate at 25 °C for 24 h. Biofilm formation assaywas repeated six times and swarming assay repeated in triplicate. Datawere expressed as X±SD. Asterisks indicated significant differencesbetween the wild-type strain and the aroA mutant. **P<0.01
Appl Microbiol Biotechnol (2013) 97:6325–6335 6329

Persistence of the aroA mutant in turbot
Simultaneously, we determined the persistence of the aroAmutant and the wild-type LSE40 in turbot infected by i.p.injection, immersion, or oral infusion. Following i.p. injectionwith a dose of ∼105CFU/fish, the wild-type LSE40 straingrew rapidly and reached 108–109CFUg−1 of the organs byday 14, which resulted in the death of all fish (data not shown).The aroA mutant grew much less efficiently, and counts were102, 103, and 104CFUml−1 on day 3 in the liver, spleen, andkidney, respectively, followed by a continued decline. ThearoA mutant was cleared from the liver by day 7 and fromthe spleen in week 4, although the bacterium still persisted inthe kidney at 1.85×101CFUg−1 in week 4 (Fig. 6).
Following immersion infection at a dose of ∼107
CFU/fish, the wild-type LSE40 strain required longer tocause a systemic infection of the organs compared with the
injection route, and it resulted in the death of all fish byday 21 (data not shown). The aroA mutant was not isolatedfrom the liver or the spleen throughout the experimentalperiod. The counts of aroA mutant in the kidney were 103
CFUml−1 on day 3 after infection, followed by a gradual
Fig. 5 Auto-aggregation and production of T3SS translocator proteinsof E. tarda. E. tarda strains were incubated in DMEM with 5 % CO2 at25 °C and aggregation was recorded after 24 h (a). The wild-typeLSE40 and the aroA mutant were incubated in DMEM for 24 h with orwithout “aromix.” The cultures normalized to OD600=1.0 were used inextracellular protein (ECP) and intracellular protein (ICP) extractionsaccording to Wang et al. (2009). Aliquots of 10 μl of each extractionwere loaded and separated by 12 % SDS-PAGE followed by Westernblot analyses using anti-EseB, anti-EseC, and anti-EseD antiserum (b)
Table 1 Mortality of turbot (S. maximus L.) challenged with E. tardaby injection, immersion, and oral routes
Treatment Mortality (%) LD50
Injection dosea (CFU/fish)
aroA mutant 7.30×109 100 7.24×107
7.30×108 60
7.30×107 10
7.30×106 0
LSE40 2.03×105 100 2.03×103
2.03×104 90
2.03×103 50
2.03×102 10
Immersion dosea (CFUml−1)
aroA mutant 7.3×108 0 >108
7.3×107 0
LSE40 2.03×108 85 6.4×106
2.03×107 65
2.03×106 0
Oral dosea (CFU/fish)
aroA mutant 7.0×109 0 >109
LSE40 7.0×109 0
Turbot with an average weight of 10 g were i.p.-injected with 0.1 ml of102 –108 CFU of wild-type or aroA mutant, immersed in seawatercontaining 106 –108 CFUml−1 , or given oral infusion with approxi-mately 109 CFU bacterial strains. Fish were observed for 14 days.LD50 values were calculated in the injection administration using thestatistical approach described by Reed and Muench (1938)a Ten fish for each concentration were used
Fig. 6 Persistence of E. tarda in turbot (S. maximus L.). Turbot(average weight, 10 g) were i.p.-injected with 2.7×105CFU/fish ofaroA mutant or immersion-exposed in 2.7×108CFU/fish of aroA mu-tant for 30 min. Bacteria that colonized in the kidney, spleen, and liverwere enumerated on TSA agar. Each experiment was repeated intriplicate. Data were expressed as X±SD
6330 Appl Microbiol Biotechnol (2013) 97:6325–6335

decline and clearance from the kidney in week 4. After oralinfusion of ∼108CFU/fish, no bacteria were recovered fromthe liver, kidney, or spleen of the infected turbot throughoutthe experimental period (data not shown). No mortality orclinical signs of illness were observed in the fish infected viathe three routes during the experimental period. These re-sults suggest that the aroA mutant established infections infish via injection and immersion routes.
Protective effects of i.p. vaccination with aroA mutant
In the first trial, groups of turbot fish (average weight, 5 g)were vaccinated i.p. with the aroA mutant at doses of 1.88×104, 1.88×105, and 1.88×106CFU/fish and challenged i.p.with 1.47×104CFU/fish of the wild-type LSE40 (LD50=7.2)at day 28 post-vaccination. The vaccinated fish died withmortality rates of 95–100 % within 30 days post-challenge,which was comparable to the 95 % mortality observed withthe control fish.
In the second trial, groups of turbot (average weight,40 g) were vaccinated i.p. with 2.14×106CFU/fish of aroAmutant and challenged i.p. with 1.64×104CFU/fish (LD50=8.1) of LSE40 or by immersion in 1.64×108CFUml−1
(LD50=81.2) of LSE40 at day 28 post-vaccination. In eachchallenged group, only 15 % of the fish survived at 30 dayspost-challenge, which was comparable to the correspondingcontrol group, i.e., 10 % survivors in the i.p. group and 20 %in the immersion group (Table 2). All of the dead fish werepositive for wild-type E. tarda and exhibited the typicalsymptoms of edwardsiellosis. These results suggest thatthe aroA mutant did not provide turbot with protection viathe i.p. vaccination route.
Protective effects of immersion vaccination with LSE40 aroA
Groups of turbot (5 g) were vaccinated via immersion indoses of 1.32×106, 1.32×107, or 1.32×108CFUml−1 of thearoA mutant and challenged by immersion with 2.01×108
CFUml−1 of wild-type LSE40 (LD50=31.4) at 30 days post-vaccination. Only 35 % of the fish vaccinated with a higherdose survived within 30 days post-challenge, which was notsignificantly different from the 40 % survival rate in thecontrol group. No survivors were observed in fish vaccinat-ed with lower doses. A second vaccination trial wasconducted using 40 g turbot by immersion with 1.66×108
CFUml−1 of aroA mutant, and the vaccinated fish werechallenged via the immersion or i.p. route at 30 days post-vaccination. It was found that 40 % of the vaccinated fishsurvived after challenge via immersion in 1.05×108CFUml−1 of LSE40, which was not significantly different fromthe 45 % survivors in the control fish group. After i.p.challenge with 1.05×104CFU/fish of LSE40, 30 % of thevaccinated fish survived, which was not significantly
different from the 35 % survivors in the control group(Table 2). All of the dead fish had the typical symptoms ofedwardsiellosis. These results suggested that the aroA mu-tant did not provide turbot with protection via the immersionvaccination route.
Humoral antibody responses in vaccinated fish
The antibody titers against formalin-killed E. tarda LSE40were determined after vaccination by i.p. or immersion withthe aroA mutant. As shown in Fig. 7a, groups of 5 g fishvaccinated via i.p. with doses of 104–106CFU of aroAmutantshowed increasing antibody titers with time after the vaccina-tion. All of the vaccinated fish had significantly higher anti-body titers than the control fish (P<0.05) during the samplingperiod, and fish that received higher doses had higher seraantibody titers. Similarly, the 40 g fish vaccinated by i.p. with∼108CFU of the aroA mutant also had significantly higher
Table 2 Mortality of vaccinated turbot (S. maximus L.) challengedwith E. tarda LSE40 by injection and immersion
Treatment Challenge via i.p.:no. of dead fish(total no. of fish)
Challenge via immersion:no. of dead fish(total no. of fish)
i.p. vaccination dose (CFU/fish)
5 g 1.88×104 20 (20)a N
1.88×105 20 (20)a N
1.88×106 19 (20)a N
Control 19 (20)a N
40 g 2.14×106 17 (20)b 17/20d
Control 18 (20)b 16/20d
Immersion vaccination dose (CFUml−1)
5 g 1.32×106 N 20 (20)e
1.32×107 N 20 (20)e
1.32×108 N 13 (20)e
Control N 12 (20)e
40 g 1.66×108 14/20c 12 (20)f
Control 13/20c 11 (20)f
Turbot were vaccinated via i.p. injection or the immersion route. Deathwere recorded for 30 daysa At 4 weeks post-vaccination, vaccinated fish groups were i.p.-chal-lenged with 1.47×104 CFU/fish of wild-type E. tarda LSE40bAt 4 weeks post-vaccination, vaccinated fish groups were i.p.-chal-lenged with 1.64×104 CFU/fish of wild-type E. tarda LSE40c At 4 weeks post-vaccination, vaccinated fish groups were i.p.-chal-lenged with 1.05×104 CFU/fish−1 of wild-type E. tarda LSE40dAt 4 weeks post-vaccination, vaccinated fish groups were immersedin 1.64×108 CFUml−1 of wild-type LSE40e At 4 weeks post-vaccination, vaccinated fish groups were immersedin 2.01×108 CFUml−1 of wild-type LSE40f At 4 weeks post-vaccination, vaccinated fish groups were immersedin 1.05×108 CFUml−1 of wild-type LSE40
Appl Microbiol Biotechnol (2013) 97:6325–6335 6331
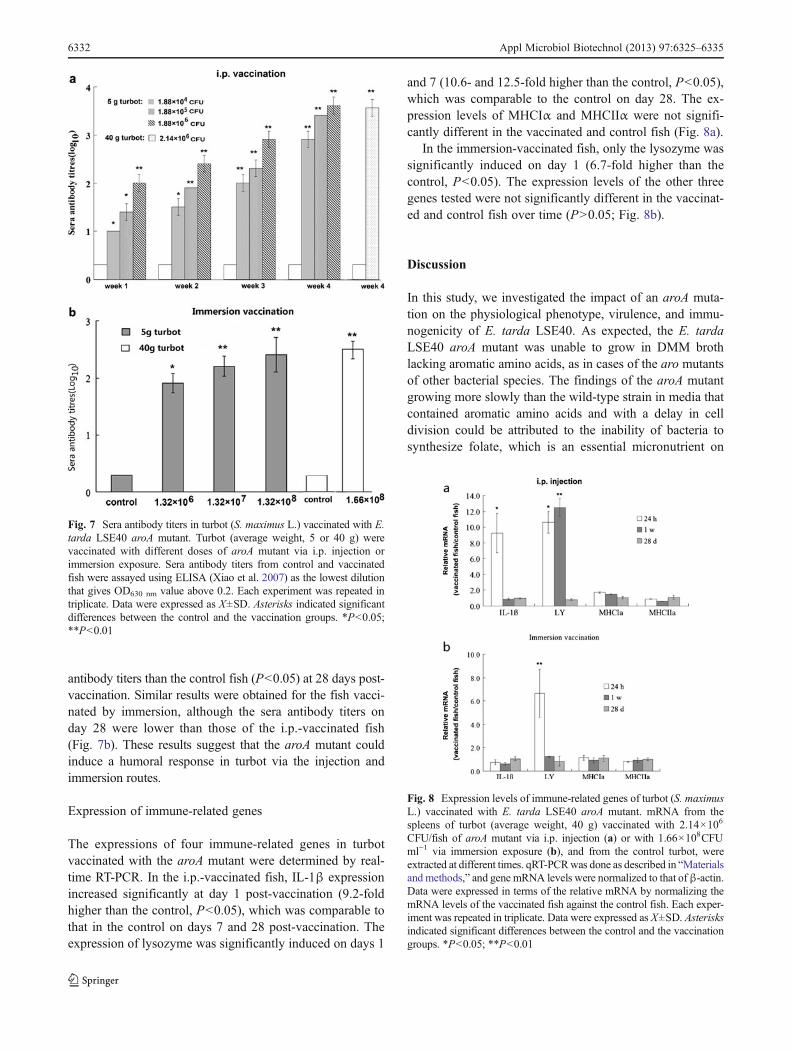
antibody titers than the control fish (P<0.05) at 28 days post-vaccination. Similar results were obtained for the fish vacci-nated by immersion, although the sera antibody titers onday 28 were lower than those of the i.p.-vaccinated fish(Fig. 7b). These results suggest that the aroA mutant couldinduce a humoral response in turbot via the injection andimmersion routes.
Expression of immune-related genes
The expressions of four immune-related genes in turbotvaccinated with the aroA mutant were determined by real-time RT-PCR. In the i.p.-vaccinated fish, IL-1β expressionincreased significantly at day 1 post-vaccination (9.2-foldhigher than the control, P<0.05), which was comparable tothat in the control on days 7 and 28 post-vaccination. Theexpression of lysozyme was significantly induced on days 1
and 7 (10.6- and 12.5-fold higher than the control, P<0.05),which was comparable to the control on day 28. The ex-pression levels of MHCIα and MHCIIα were not signifi-cantly different in the vaccinated and control fish (Fig. 8a).
In the immersion-vaccinated fish, only the lysozyme wassignificantly induced on day 1 (6.7-fold higher than thecontrol, P<0.05). The expression levels of the other threegenes tested were not significantly different in the vaccinat-ed and control fish over time (P>0.05; Fig. 8b).
Discussion
In this study, we investigated the impact of an aroA muta-tion on the physiological phenotype, virulence, and immu-nogenicity of E. tarda LSE40. As expected, the E. tardaLSE40 aroA mutant was unable to grow in DMM brothlacking aromatic amino acids, as in cases of the aro mutantsof other bacterial species. The findings of the aroA mutantgrowing more slowly than the wild-type strain in media thatcontained aromatic amino acids and with a delay in celldivision could be attributed to the inability of bacteria tosynthesize folate, which is an essential micronutrient on
Fig. 7 Sera antibody titers in turbot (S. maximus L.) vaccinated with E.tarda LSE40 aroA mutant. Turbot (average weight, 5 or 40 g) werevaccinated with different doses of aroA mutant via i.p. injection orimmersion exposure. Sera antibody titers from control and vaccinatedfish were assayed using ELISA (Xiao et al. 2007) as the lowest dilutionthat gives OD630 nm value above 0.2. Each experiment was repeated intriplicate. Data were expressed as X±SD. Asterisks indicated significantdifferences between the control and the vaccination groups. *P<0.05;**P<0.01
Fig. 8 Expression levels of immune-related genes of turbot (S. maximusL.) vaccinated with E. tarda LSE40 aroA mutant. mRNA from thespleens of turbot (average weight, 40 g) vaccinated with 2.14×106
CFU/fish of aroA mutant via i.p. injection (a) or with 1.66×108CFUml−1 via immersion exposure (b), and from the control turbot, wereextracted at different times. qRT-PCRwas done as described in “Materialsand methods,” and gene mRNA levels were normalized to that ofβ-actin.Data were expressed in terms of the relative mRNA by normalizing themRNA levels of the vaccinated fish against the control fish. Each exper-iment was repeated in triplicate. Data were expressed as X±SD. Asterisksindicated significant differences between the control and the vaccinationgroups. *P<0.05; **P<0.01
6332 Appl Microbiol Biotechnol (2013) 97:6325–6335

certain bacteria and a promoter of bacterial growth (Regeand Sreenivasan 1954; Shane and Stokstad 1975).
The aroAmutant also had reduced swarming motility andbiofilm formation. Motile bacteria use filamentouspropeller-like flagella for motility, and the flagella are asso-ciated with biofilm establishment (Verstraeten et al. 2008).Mutation in aroA gene blocks the synthesis of ubiquinonethat is required for bacterial flagellation (Hertz and Bar-Tana1977; Bar-Tana et al. 1980). It is possible that mutation inaroA impaired the flagellation in E. tarda and consequentlyaffected the flagella-mediated motility and biofilm produc-tion of this bacterium. The reduced T3SS translocator pro-tein production in the aroA mutant may also contribute tothe reduced swarming motility and biofilm formation be-cause T3SS translocation, which is mediated by a pilus-likestructure, is known to be involved in motility and biofilmformation in E. tarda (Wang et al. 2009). Furthermore, arole for aroA in capsular polysaccharide production cannotbe excluded because aro mutations in Salmonella enteritidislead to the reduced production of cellulose, N-acetyl-D-glu-cosamine, or N-acetylneuraminic acid in the capsular poly-saccharide, and fimbriae, which explains their inability toform a biofilm (Malcova et al. 2009). However, the actualreason for the association of the aroAmutation with flagellarformation, T3SS translocation protein production, and cap-sular polysaccharide production remains unclear in E. tarda.
Similar to the aro mutants of other bacterial species, thevirulence of the E. tarda LSE40 aroA mutant was stronglyattenuated by more than 2–3 log10 via the infection routes ofinjection and immersion compared with the wild-type parentstrain. A shortage of aromatic amino acids in the fish host maybe a major reason for its attenuated virulence. Other physio-logical changes might also contribute to the attenuated viru-lence of E. tarda LSE40, such as the impaired flagella motilitythat reduces access to the appropriate niche, the failure ofbiofilm attachment during colonization of the host cell sur-face, and a lack of functional T3SS with toxic effects on hostcell functions. Unlike the other aroAmutants of other bacterialspecies, which can provide efficient protection against bacte-rial infections (Vaughan et al. 1993; Marsden et al. 1996;Hernanz Moral et al. 1998; Thune et al. 1999), the LSE40aroA mutant was not successful in providing protectionagainst edwardsiellosis in turbot vaccinated by i.p. injectionor immersion exposure, where the challenge dosages usedwere LD50=7.2–8.1 for the i.p. route and LD50=31.4–81.2for the immersion route. Protection was also not successful forthe aroA mutant in the vaccinated turbot when challengedwith dosages lower than LD50=1 via i.p. or via the immersionroute (data not shown). The reason for the ineffective protec-tion provided by the aroA mutant is less clear. There havebeen similar reports for some aromutants of Salmonella spp.,and it is possible that they were unable to trigger a strongimmune response or to multiply/persist in host tissue
(O’Callaghan et al. 1988; Trebichavsky et al. 2006). In thisstudy, the inability to multiply/persist in host tissue can beruled out because the invasion and persistence of the LSE40aroA mutant was detected up to 21 days after fish wereinfected via immersion and at 28 days in i.p.-infected fish.The inability to trigger humoral adaptive immune responsealso appears unlikely because increasing antibody titers weretested in the vaccinated fish. E. tarda is able to invade intofish’s non-phagocytic cells and survive in the phagocytes(Ling et al. 2000; Rao et al. 2001), leading to the assumptionthat T cell-mediated immune response could be the mostimportant component for protection against this pathogen.Over the last decade, T cell-related genes including TCR,CD3, CD4, CD8, CD28, CTLA4, MHC I, MHC II as wellas important cytokines have been identified in some fishspecies (Castrol et al. 2011), suggesting the existence of Tcytotoxicity and T helper subtypes in fish as those in mammal.In this study, analysis of the expression levels of immune-related factors showed that only the innate immune factors,IL-1ß and lysozyme, were stimulated significantly in thevaccinated fish, whereas the Tcell-mediated adaptive immunefactors,MHCIα andMHCIIα, were not. Asmentioned above,the phenotypic changes in the aroAmutant could contribute todefects in flagellin, capsular polysaccharide, fimbriae, andT3SS translocator proteins, which are potential surface-associated antigens for immunity both to extracellular andintracellular pathogens (Barat et al. 2012). It is possible thatthe ineffective stimulation of T cell-mediated adaptive im-mune responses in turbot by the aroA mutant might make itincapable of triggering T cell subsets to kill the ingestedE. tarda cells. The role of elevated antibody induced by thearoAmutant against E. tarda infection needs further clarifica-tion since the antibody-mediated immunity is known to pro-vide protection against intracellular pathogens by variousmechanisms including direct bactericidal activity, comple-ment activation, opsonization, cellular activation, andantibody-dependent cellular cytotoxicity.
This study demonstrated that aroA has a pleiotropic effecton the physiological behavior of E. tarda, including itsgrowth, motility, biofilm formation, and T3SS translocatorproduction, which also contribute to its virulence. The de-letion of one of the E. tarda T3SS translocators could lead toabout 1 log10 virulence attenuation in blue gorami (Tan et al.2005), but it is not clear how the other features are relevantto the aroA mutant’s attenuation compared with its inabilityto synthesize aromatic amino acids. Our results also dem-onstrated that the aroA mutant was ineffective in providingimmune protection against edwardsiellosis; further work isneeded to address the mechanism of aroA on T cell activity.Unfortunately, compared to other fish species, few turbotgenomic resources are available and little is known aboutthe immune relevant genes that are essential to analyzeinfection, immunity, and host–pathogen interactions. Thus,
Appl Microbiol Biotechnol (2013) 97:6325–6335 6333

gene information of the T cell markers and cytokines ofturbot becomes much more necessary for a thorough illus-tration of the mechanism of aroA on host immune response.
Acknowledgments This project was supported by the National NaturalSciences Foundation of China (grant no. 31072245) and the planningproject of “the twelfth five-year-plan” in the National Science and Tech-nology for the Rural Development in China (2011BAD13B07) and inMarine Technology for Typical Marine Biological Development andUtilization of Important Functional Genes (2012AA092203).
Conflict of interest The authors declare that they have no conflict ofinterest.
References
Barat S, Willer Y, Rizos K, Claudi B, Mazé A, Schemmer AK, KirchhoffD, Schmidt A, Burton N, Bumann D (2012) Immunity toIntracellular Salmonella depends on surface-associated antigens.PLoS Pathog 8:e1002966. doi:10.1371/journal.ppat.1002966
Bar-Tana J, Howlett BJ, Hertz R (1980) Ubiquinone synthetic pathwayin flagellation of Salmonella typhimurium. J Bacteriol 143:637–643
Castro R, Bernard D, Lefranc MP, Sixc A, Benmansour A, Boudinot P(2011) T cell diversity and TcR repertoires in teleost fish. FishShellfish Immunol 31:644–654. doi:10.1016/j.fsi.2010.08.016
Choi SH, Kim KH (2011) Generation of two auxotrophic genes knock-out Edwardsiella tarda and assessment of its potential as a com-bined vaccine in olive flounder (Paralichthys olivaceus). FishShellfish Immunol 31:58–65. doi:10.1016/j.fsi.2011.03.006
Cook RA, Tappe JP (1985) Chronic enteritis associated withEdwardsiella tarda infection in Rockhopper penguins. J Am VetMed Assoc 187:1219–1220
Fletcher M (1977) The effects of culture concentration and age, time,and temperature on bacterial attachment to polystyrene. Can JMicrobiol 23:1–6. doi:10.1139/m77-001
Hernanz Moral C, Flańo del Castillo E, López Fierro P, Villena CortesA, Anguita Castillo J, Cascnó Soriano A, Sánchez Salazar M,Razquin Peralta B, Naharro Carrasco G (1998) Molecular char-acterization of the Aeromonas hydrophila aroA gene and potentialuse of an auxotrophic aroA mutant as a live attenuated vaccine.Infect Immun 66:1813–1821
Hertz R, Bar-Tana J (1977) Anaerobic electron transport in anaerobicflagellum formation in Escherichia coli. J Bacteriol 132:1034–1035
Hirono I, Tange N, Aoki T (1997) Iron-regulated hemolysin gene fromEdwardsiella tarda. Mol Microbiol 24:851–856. doi:10.1046/j.1365-2958.1997.3971760.x
Jobichen C, Chakraborty S, Li M, Zheng J, Joseph L, Mok YK, LeungKY, Sivaraman J (2010) Structural basis for the secretion of EvpC: akey type VI secretion system protein from Edwardsiella tarda.PLoS One 5:e12910. doi:10.1371/journal.pone.0012910
Ling SHM, Wang XH, Xie L, Lim TM, Leung KY (2000) Use of greenfluorescent protein (GFP) to study the invasion pathways ofEdwardsiella tarda in in vivo and in vitro fish models.Microbiology 146:7–19
Lisa AC, Ronald LT (1996) Development of a defined minimal medi-um for the growth of Edwardsiella ictaluri. Appl EnvironMicrobiol 62:848–852
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expressiondata using real-time quantitative PCR and the 2−ΔΔCT method.Methods 25:402–408. doi:10.1006/meth.2001.1262
Malcova M, Karasova D, Rychlik I (2009) aroA and aroD mutations ininfluence biofilm formation in Salmonella enteritidis. FEMSMicrobiol Lett 291:44–49. doi:10.1111/j.1574-6968.2008.01433.x
Marsden MJ, Vaughan LM, Foster TJ, Secombes CJ (1996) A live(delta aroA) Aeromonas salmonicida vaccine for furunculosispreferentially stimulates T-cell responses relative to B-cell re-sponses in rainbow trout (Oncorhynchus mykiss). Infect Immun64:3863–3869
Mathew JA, Tan YP, Rao PSS, Lim TM, Leung KY (2001) Edwardsiellatarda mutants defective in siderophore production, motility, serumresistance and catalase activity. Microbiology 147:449–457
Mo ZL, Xiao P, Mao YX, Zou YX, Wang B, Li J, Xu YL, Zhang PJ(2007) Construction and characterization of a live, attenuated esrBmutant of Edwardsiella tarda and its potential as a vaccine againstthe haemorrhagic septicaemia in turbot, Scophthamus maximus (L.).Fish Shellfish Immunol 23:521–530. doi:10.1016/j.fsi.2006.11.00
Mohanty BR, Sahoo PK (2007) Edwardsiellosis in fish: a brief review.J Biosci 32:1331–1344. doi:10.1007/s12038-007-0143-8
O’Callaghan D, Maskell D, Liew FY, Easmon CS, Dougan G (1988)Characterization of aromatic- and purine-dependent Salmonellatyphimurium: attenuation, persistence, and ability to induce pro-tective immunity in BALB/c mice. Infect Immun 56:419–423
Pittard AJ (1996) Biosythesis of aromatic amino acids. In: NeidhardtFC (ed) Escherichia coli and Salmonella typhimurium: cellularand molecular biology, 2nd edn. American Society forMicrobiology, Washington, DC, pp 458–484
Rao PSS, Lim TM, Leung KY (2001) Opsonized virulent Edwardsiellatarda strains are able to adhere to and survive and replicate withinfish phagocytes but fail to stimulate reactive oxygen intermediates.Infect Immun 69:5689–5697. doi:10.1128/IAI.69.9.5689-5697.2001
Rao PSS, Lim TM, Leung KY (2003) Functional genomics approachto the identification of virulence genes involved in Edwardsiellatarda pathogenesis. Infect Immun 71:1343–1351. doi:10.1128/IAI.71.3.1343-1351.2003
Rao PSS, Yamada Y, Tan YP, Leung KY (2004) Use of proteomics toidentify novel virulence determinants that are required forEdwardsiella tarda pathogenesis. Mol Microbiol 53:573–586.doi:10.1111/j.1365-2958.2004.04123.x
Reed LJ, Muench H (1938) A simple method of estimating fiftypercent end points. Am J Epidemiol 27:493–497
Rege DV, Sreenivasan A (1954) The influence of folic acid and vitaminB12 on nucleic acid metabolism in microorganisms. J Biol Chem210:373–380
Rychlik I, Cardova L, Sevcik M, Barrow PA (2000) Flow cytometrycharacterisation of Salmonella typhimurium mutants defective inproton translocating proteins and stationary-phase growth pheno-type. J Microbiol Methods 42:255–263. doi:10.1016/S0167-7012(00)00199-8
Sakai T, Kanai K, Osatomi K, Yoshikoshi K (2003) Identification of a19.3-kDa protein in MRHA-positive Edwardsiella tarda: putativefimbrial major subunit. FEMS Microbiol Lett 226:127–133.doi:10.1016/S0378-1097(03)00608-6
Sakai T, Matsuyama T, Sano M, Iida T (2009) Identification of novelputative virulence factors, adhesin AIDA and type VI secretionsystem, in atypical strains of fish pathogenic Edwardsiella tardaby genomic subtractive hybridization. Microbiol Immunol53:131–139. doi:10.1111/j.1348-0421.2009.00108.x
Sambrook JE, Fritsch EF, Maniatis T (1989) Molecular cloning: alaboratory manual, 2nd edn. Cold Spring Harbor, New York
Sebkova A, Karasova D, Crhanova M, Budinska E, Rychlik I (2008) aromutations in Salmonella enterica cause defects in cell wall and outermembrane integrity. J Bacteriol 190:3155–3160. doi:10.1128/JB.00053-08
Shane B, Stokstad EL (1975) Transport and metabolism of folates bybacteria. J Biol Chem 25:2243–2245
6334 Appl Microbiol Biotechnol (2013) 97:6325–6335

Tan YP, Zheng J, Tung SL, Rosenshine I, Leung KY (2005) Role oftype III secretion in Edwardsiella tarda virulence. Microbiology151:2301–2313. doi:10.1099/mic.0.28005-0
Thune RL, Fernandez DH, Batista J (1999) An aroA mutant ofEdwardsiella ictaluri is safe and efficacious as live, attenuatedvaccine. J Aquat Anim Health 11:358–372. doi:10.1577/1548-8667(1999)011<0358:AAMOEI>2.0.CO;2
Trebichavsky I, Splichalova A, Rychlik I, Hojna H, Muneta Y, Mori Y,Splichal I (2006) Attenuated aroA Salmonella enterica serovartyphimurium does not induce inflammatory response and earlyprotection of gnotobiotic pigs against parental virulent LT2 strain.Vaccine 24:4285–4289. doi:10.1016/j.vaccine.2006.02.054
Vaughan LM, Smith PR, Foster TJ (1993) An aromatic-dependentmutant of the fish pathogen Aeromonas salmonicida is attenuatedin fish and is effective as a live vaccine against the salmoniddisease furunculosis. Infect Immun 61:2172–2181
Verstraeten N, Braeken K, Debkumari B, Fauvart M, Fransaer J,Vermant J, Michiels J (2008) Living on a surface: swarming andbiofilm formation. Trends Microbiol 16:496–506. doi:10.1016/j.tim.2008.07.004
Wang B, Mo ZL, Mao YX, Zou YX, Xiao P, Li J, Yang JY, Ye XH,Leung KY, Zhang PJ (2009) Investigation of EscA as a chaperonefor the Edwardsiella tarda type III secretion system putative
translocon component EseC. Microbiology 155:1260–1271.doi:10.1099/mic.0.021865-0
Wang X, Wang Q, Xiao J, Liu Q, Wu H, Zhang Y (2010) HemolysinEthA in Edwardsiella tarda is essential for fish invasion in vivoand in vitro and regulated by two-component system EsrA-EsrBand nucleoid protein HhaEt. Fish Shellfish Immunol 29:1082–1091. doi:10.1016/j.fsi.2010.08.025
Xiao P, Mo ZL, Zou YX, Wang B, Xu YL, Zhang PJ (2007) Immuneresponses and efficacy of an oral-emulsified bivalent vaccineagainst Vibrio anguillarum in turbot (Scophthalmus maximus).High Technol Lett 17:979–985
Xiao J, Chen T, Wang Q, Liu Q, Wang X, Lv Y, Wu H, Zhang Y (2011)Search for live attenuated vaccine candidate against edwardsiellosisby mutating virulence-related genes of fish pathogen Edwardsiellatarda. Lett Appl Microbiol 53:430–437. doi:10.1111/j.1472-765X.2011.03126.x
Xie HX, YuHB, Zheng J, Nie P, Foster LJ,MokYK, Finlay BB, Leung KY(2010) EseG, an effector of the type III secretion system ofEdwardsiella tarda, triggers microtubule destabilization. InfectImmun 78:5011–5021. doi:10.1128/IAI.00152-10
Zhang M, Sun K, Sun L (2008) Regulation of autoinducer 2 productionand luxS expression in a pathogenic Edwardsiella tarda strain.Microbiology 157:2060–2069. doi:10.1099/mic.0.2008/017343-0
Appl Microbiol Biotechnol (2013) 97:6325–6335 6335





























