Embed Size (px)
Citation preview

Physica D 211 (2005) 128–138
Phase resetting and transient desynchronization in networks ofglobally coupled phase oscillators with inertia
Kevin Dolana,b,∗, Milan Majtanika,b, Peter A. Tassa,b,c
a Institute of Medicine, Research Center Julich, 52425 Julich, Germanyb Virtual Institute of Neuromodulation, Research Center Julich, 52425 Julich, Germany
c Department of Stereotaxic and Functional Neurosurgery, University Hospital, 50924 Cologne, Germany
Received 18 August 2004; received in revised form 22 July 2005; accepted 10 August 2005Available online 12 September 2005
Communicated by Y. Kuramoto
Abstract
Recently extensive work has been done towards developing methods for effective desynchronization of globally coupled phaseoscillators (the Kuramoto model), by means of short stimulation pulses, or sequences of pulses. This is of great importance forthe treatment of neurological disorders like Parkinson’s disease and essential tremor. As a progressive step towards the goal ofbeing able to apply these desynchronization and phase-resetting techniques medically as a form of treatment, we demonstratehere how these ideas can be generalized and applied to a network of two-dimensional phase oscillators with inertia. This modelhas been previously presented as a simplification of a neuron with an axon and dendrite, and can be used to account for intrinsict l work ona n can bea which thisd additiono inciples ofp importancet©
P
K
ofof
0d
ransient behavior often seen experimentally. The stimulation techniques originally developed for the Kuramoto modenetwork of globally coupled inertial phase oscillators in a qualitatively similar way. In both cases desynchronizatio
chieved when the stimulation causes nearby trajectories to diverge from each other. However, the mechanism byivergence of trajectories is achieved, is substantially different for the network of inertial oscillators. In particular, thef inertia results in a broad range of transient dynamics not present in the Kuramoto model. Nevertheless, the basic prhase resetting and desynchronization still apply. This suggests a robustness of these techniques which is of extreme
o the medical applications.2005 Elsevier B.V. All rights reserved.
ACS: 05.45.Xt; 05.40.Ca; 87.19.La
eywords: Phase resetting; Desynchronization; Transient dynamics; Inertia
∗ Corresponding author.E-mail address: [email protected] (K. Dolan).
1. Introduction
The synchronization behavior of systemscoupled oscillators is of interest to a wide varietytopics in physics[1], chemistry [2], biology [3,4],
167-2789/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.physd.2005.08.009

K. Dolan et al. / Physica D 211 (2005) 128–138 129
neuroscience[5], and medicine[6,7]. In particular, thetopics of phase resetting[8,9,6]and desynchronization[6,10–12], are of great importance to the field of neu-roscience, where they show considerable potential forimproving our understanding of, and developing newtreatments for, neurological disorders like Parkinson’sdisease and essential tremor.[6,10–12].
An example of this type of application is in treat-ing the tremor associated with Parkinson’s disease. Inthis case, each oscillator represents a single neuron.The tremor occurs when a population of neurons firesin a synchronized way[13–15], while under healthyconditions these neurons fire in an uncorrelated man-ner [16]. One method of treating this tremor has beento use deep brain stimulation to introduce a high fre-quency, high amplitude signal to the relevant area ofthe brain[17,18]. This type of stimulation suppressesthe tremor by mimicking the effects of tissue lesion-ing, one candidate mechanism of the conventional deepbrain stimulation being the suppression of neuronal fir-ing [19,20]. There are several drawbacks to this tech-nique, including possible side effects of such a strong,persistent stimulation[21,22], as well as the fact thatthe therapeutic effect of the stimulation may vanishover time[21,22]. One goal of phase resetting researchis to develop new methods of deep brain stimulationwhich will utilize occasional, low-amplitude pulses, inorder to desynchronize the relevant neuronal popula-tion, rather than suppressing it entirely[23–26].
Extensive work has been done with the Kuramotom hatp mentot ec-e del,s gicalc s not in-t ownf pli-t ritye ou-p ddi-t tionf orec ticm ti-v new
model which introduces another level of complexitywill be used.
In this paper, we introduce the first step in this pro-cess. We will generalize the Kuramoto model to a net-work of two-dimensional phase oscillators with inertia.Inertia will be a necessary component of our final mi-croscopic model, since it is known that individual neu-rons are capable of exhibiting transient dynamics[27–30]. Specifically, recent research has shown transientdynamics which can be directly attributed to inertialeffects within the neuron[30]. Such inertial effects canpotentially have a strong influence on the appropriatestimulation timing.
Although the Kuramoto model exhibits transient dy-namics for the coupled network, an oscillator model ofat least second order is necessary to produce transientdynamics for the individual oscillators. It has recentlybeen shown that a two-dimensional phase oscillatorwith inertia can be derived as a simplified model ofa neuron with an axon and dendritic tree[31,32]. Wewill use this model to study the effects that the phaseresetting and desynchronizing stimulation techniquespreviously applied to the Kuramoto model, have on amore physically realistic system with intrinsic inertia.While this model has been derived as a simplified neu-ron model, it is also quite general, and should haverelevance to any physical system of coupled oscillatorsin which phase dynamics are of principle interest, andtransient properties of the individual oscillators mustbe taken into account.
p el ofH u-p et esyn-c thati s int n-c rred,a redf
2
hro-n heirp
odel [2], demonstrating the potential benefit thase resetting techniques could have for the treatf Parkinsonian tremor[6,10–12,23–26]. But in order
o optimize these techniques for clinical use, it is nssary to move to a full microscopic neuronal moo that they can be adapted to the given physioloonstraints. Unfortunately, such a generalization irivial task. Moving to such a complicated modelroduces many new types of behavior that are knrom studies of stationary dynamics, such as amude dynamics, multiple sources of noise, nonlineaffects (including chaos), and more complicated cling schemes. In order to resolve these various a
ional factors, we are proposing a step by step migrarom the Kuramoto model, through progressively momplicated models, finally culminating in a realisicroscopic neuron model, with physiologically mo
ated noise and coupling schemes. At each step, a
This paper is organized as follows. In Section2 weresent the two-dimensional phase oscillator modaken[32], adapting it to a network of globally coled oscillatory neurons. In Section3 we demonstrat
he mechanisms of phase resetting and transient dhronization in this model, and discuss the wayst is similar to, and different from, these processehe Kuramoto model. In Section4we discuss the resyhronization process after phase resetting has occund in particular, how to maximize the time requi
or the system to return to its synchronized state.
. Globally coupled phase oscillators
One approach towards understanding the syncization behavior of coupled neurons is to model thase dynamics with a simple phase oscillator[2,6,33–

130 K. Dolan et al. / Physica D 211 (2005) 128–138
35]. The idea is to model just the phase dynamics, eitherneglecting other dynamical factors, such as amplitudedynamics or chaotic dynamics, or approximating themas part of the internal noise.
2.1. The one-dimensional Kuramoto model
Recent work has been done demonstrating desyn-chronizing and phase resetting techniques on a sys-tem of globally coupled phase oscillators by extend-ing the Kuramoto model[2] with a stimulation term[6,10,11,23,12,24–26]:
φj = ωj + Γj +X(t)S(φj) +√
2Dξj(t). (1)
In this type of model, the phase dynamics of the neu-rons are approximated with a simple one-dimensionalphase oscillator with natural frequencyωj. The phaseφj is taken modulo 2π. Global coupling, given bythe functionΓj, is used, along with the introductionof statistically independent dynamical noise of inten-sity D, whereξj(t) is Gaussian white noise with zeromean and unit variance. The stimulation to the neuronsis modeled by a phase-dependent stimulation on thephase dynamics of each oscillator, given by the func-tion S(φj). This represents the fact that the responseof a real dynamical oscillator to a given stimulus willdepend on the current phase of the oscillator[3,4].X(t)is a step function, such thatX(t) = 0 when the stim-ulation is off, andX(t) = 1 when the stimulation iso
con-s f theo ari-a illh stan-t pendso oest n bec sea willa to ac motom bys onS
, asa the
complex order parameter.
Z(t) = R(t) exp(iφ(t)) = 1
N
N∑
j=1
exp(iφj(t)), (2)
whereN is then number of oscillators in the ensem-ble.Z(t) is a complex number, whose modulusR(t) isa measure of the degree of synchronization in the en-semble, and whose phaseφ(t) indicates the averagedphase of the synchronized ensemble[2].
It has been demonstrated that a strong pulse admin-istered at a very specific point in the cycle of the syn-chronized oscillators, will cause the entire ensemble todesynchronize[6]. The timing of the pulse is critical,as desynchronization is only achieved if the pulse isadministered during a very short portion of the wholecycle. Exactly where this critical region is located, andhow large it is, depends on the specific functional formchosen for the phase dependence of the stimulation re-sponse[6].
2.2. The two-dimensional phase oscillator modelwith inertia
It is known that individual neurons can exhibit tran-sient dynamics when exposed to changes in externalconditions, such as temperature[28,29] and electricalstimulation[27,30]. An obvious possible source of suchtransient effects is intrinsic inertia. In fact, in the recentw ectsw e-c onalp d asaI .
st ofa thed
ψ
wr ledad uroni ite,a nw ron.
n.For such a phase oscillator, the response to a
tant stimulus will not depend on the current state oscillator, because the dynamics are rotationally invnt. In a real dynamical oscillator, the limit cycle wave an instantaneous amplitude, as well as an in
aneous phase. The instantaneous amplitude den where on the limit cycle the oscillator is, as d
he instantaneous phase. Thus, the amplitude caonsidered a 2π periodic function of the phase. Themplitude dynamics influence how the stimulusffect the system, effectively making the responseonstant stimulus phase-dependent. For the Kuraodel, this dependency can be modeled explicitly
tipulating the functional form of the periodic functi(ψj) [8,6,10–12].
The degree of synchronization of the ensemblefunction of time, can then be determined from
ork of Hauptmann and Mackey, the transient effere directly attributed to such inertial effects. Rent work by Haken has shown that a two-dimensihase oscillator model with inertia can be deriven approximation of an axon with a dendritic tree[32].
t is this model which we will adapt for our analysisThe neuron model presented by Haken consisingle dendrite and an axon. The dynamics of
endrite are governed by
˙j = af (φk) − γψj +
√2Dξj(t), (3)
hereψj denotes the dendritic current of thejth neu-on. The influence of the neurons to which it is coupre given byf (φk(t)), and scaled by the factora. φkenote the phases of the axons to which the ne
s coupled.γ is the damping inherent in the dendrndD is the noise intensity, whereξj(t) is Gaussiahite noise, statistically independent for each neu

K. Dolan et al. / Physica D 211 (2005) 128–138 131
This represents the statistical fluctuations inherent inthe openings and closings of the ion channels.
The phase dynamics of the axon are governed by
φj = cψj + pext,j, (4)
where the dendritic currentsψj, are scaled by a con-stantc, andpext,j represents any external inputs to theneuron, for example, from sensory neurons.
The combination of(3) and (4)is equivalent to:
φj = Cj − γφj + Af (φk) +√
2Dξj(t), (5)
whereCj = γpext,j + pext,j, andA = ca.This type of second order phase oscillator model has
also been studied numerically for purposes of under-standing phase-frequency synchronization[36,37].
To adapt this model for the analysis we are interestedin, we make the following adjustments. First, we areinterested in neurons which fire in a self-sustained way,whereas all activity in the Haken model is the result ofsensory input to the neuron. Such autonomously firingneurons are of direct interest for the development oftreatments for Parkinson’s disease, where so-calledno-response cells can be found in the thalamus, which fireperiodically, irrespective of any kind of sensory input[13,14,38,15]. Under healthy conditions, the firing ofthese neurons is intrinsically correlated with sensoryinput.
Such self-sustained periodic behavior is usuallym rentw atest sesw es ltsi lf
hen en-d ingm
m
wi ng.Ae ate
the function describing the coupling with
Γj = K
N
N∑
k=1
sin(φk − φj), (7)
whereK is the coupling strength, andN is the numberof neurons in the network.S(φj) is a function representing the phase-
dependent response of the neuron to the stimulation,andX(t) is a step function, such thatX(t) = 0 whenthe stimulation is off, andX(t) = 1 when the stimula-tion is on.D is the intensity of the Gaussian white noiseof zero mean and unit variance, given byξj(t).
The functionS(φj) is used, as in Eq.(1), to repre-sent the phase-dependent response of the oscillator toa stimulation, which is the same for all oscillators inthe network. It is given by
S(φj) = Sdend,j + γSaxon,j + Saxon,j, (8)
whereSdend,j is the effect of the stimulation on theaxon, which would appear as an additive term on theright-hand side of Eq.(3), andSaxon,j is the effect ofthe stimulation on the axon, which would appear as anadditive term on the right hand side of Eq.(4).
In principle, this could be any periodic function ofφ,and depends entirely on the dynamics of the oscillatorbeing modeled. Such a function can be expressed as aFourier series,
S(φ) =∞∑
[Ak cos(kφ) + Bk sin(kφ)]. (9)
F wew ar-m y,a e oft on-a=
en-e is-ci iva-l x-c s ofi are,w rss andc
odelled in microscopic models as a calcium curithin the axon, and experimental evidence indic
hat this is the mechanism for the pathological cae are interested in[13,38,15]. We therefore introducuch a current to Eq.(4), as a constant term. This resun the substitutionCj = γωj, whereωj is the naturarequency of thejth neuron.
Second, we will apply a stimulation to all of teurons, which will have effects on both the drites and the axons. This results in the followodel,
φj = ωj − φj + Γj +X(t)S(φj) +√
2Dξj(t), (6)
herem = 1/γ is the inertia of the oscillator andΓjs a periodic function describing the global couplis with the modified Kuramoto model in Eq.(1), wemploy a global mean-field coupling, and approxim
k=0
or our preliminary investigation of this system,ill approximate this function using only the first honic, S(φ) = I cos(φ). Note that due to symmetrs long as only one harmonic is used, the phas
he harmonic is trivial. That is, the above is functilly equivalent toI1 cos(φ) + I2 sin(φ), whereI2
1 + I22
I2.It should be noted that this model is simply a g
ralization of the stimulated Kuramoto model dussed in Section2.1. In fact, in the limit that thenertia m goes to zero, this model becomes equent to Eq.(1). This model therefore provides an eellent opportunity to see exactly what the effectnertia on phase resetting and desynchronizationithout introducing additional confounding factouch as amplitude dynamics, limit cycle stability,haos.

132 K. Dolan et al. / Physica D 211 (2005) 128–138
3. Phase resetting and desynchronization
In the previous work done on the one-dimensionalphase oscillator model in Eq.(1), it was found that ashort pulsatile stimulus applied when the synchronizedsystem is at a critical phase, will cause the system todesynchronize[6]. The phase of the system is definedin terms of the complex order parameter, defined in Eq.(2) [2]. The phaseφ(t) indicates the average phase of thesynchronized system, and the amplitudeR(t) indicatesthe degree of synchronization.
To begin our investigation of these stimulation tech-niques on the inertial model described in Eq.(6), webegin by selecting a set of parameters which result instrong, but not complete, synchronization in the ab-sence of stimulation. We have chosenm = 2π, ωj =2π for all j (no detuning),K = 8π, andD = 0.007.This set of parameters results in an order parametermagnitude of aboutR = 0.85. We then apply stimula-tion, as described above, whereS(φ) = I cos(φ).
3.1. Dynamics of a single oscillator
In the one-dimensional model given by Eq.(1), asufficiently strong stimulation amplitude (I ≥ ω), inthe absence of coupling and noise, results in the ap-pearance of a stable and unstable fixed-point in thephase space of the oscillator, located at the two pointssatisfying cos(ψ) = ω/I. Because the model is one-dimensional, the oscillator must go to the stable fixed-pw viori tionc ptotics
ab-s xed-p ointsa t thev at iti rre-s ap-t tions ory,a
sev-e ener-a ator
in the absence of coupling or noise, with different ini-tial conditions, and plotting these trajectories on a graphof instantaneous frequency vs phase. We use the termphase velocity to refer to the instantaneous frequency,both in this figure, and throughout this paper, in order toavoid confusion with other quantities such as the natu-ral frequency of the oscillator, or the average frequencyof the network. We can see fromFig. 1a (I = ω/2) thatthe fixed-points do not yet exist. The stable periodictrajectory is slightly perturbed by the stimulation. ForI = 1.5ω (Fig. 1b) the fixed-point has appeared, andtwo separate basins of attraction are formed. Becausethe stimulation is applied to the system in its synchro-nized state, we are especially interested in initial con-ditions for which the phase velocity is equal toω = 2πrad/s, or 1 Hz. We see that forI = 1.5ω all of theseinitial conditions go to the periodic trajectory.
ForI = 3.0ω (Fig. 1c) the basin of attraction of thefixed-point extends above the 1 Hz line for some valuesof the phase. This means that, depending on the stateof the ensemble when stimulation begins, the ensemblemay go to the periodic trajectory, or to the fixed-point.If the initial phase of the ensemble lies very close tothe separation of these two basins of attraction, thenthe ensemble can even be split into two groups, oneof which goes to the fixed-point, and the other to theperiodic trajectory. This bistability plays a critical rolein the desynchronization behavior.
At some critical point, nearI = 4ω, the periodic tra-jectory collides with the boundary between the basinso gere int.N nalm r tor mf pe-r
3
onseo vea ork.F r,a thei 1)a tepso and
oint within one cycle. This is called ahard reset. Foreaker stimulation strengths, the asymptotic beha
s a periodic solution. In both cases, desynchronizaan be achieved as the system goes to its asymtate.
For the inertial phase oscillator model, in theence of noise and coupling, a stable and unstable fioint pair also appears. The phases of these fixed pre the same as for the one-dimensional model, buelocity at the fixed-points is zero. This means ths possible for an oscillator to cross the phase coponding to the stable fixed-point, without being cured. Because of this, there is a range of stimulatrengths for which both a stable periodic trajectnd a stable fixed-point, exist.
Fig. 1shows the phase space flow diagrams forral stimulation strengths. These diagrams were gted by computing the trajectories of a single oscill
f attraction. The stable periodic trajectory no lonxists, and all initial conditions go to the fixed-poote that unlike the case with the one-dimensioodel, it may take several cycles for the oscillato
each the fixed-point.Fig. 1d shows the flow diagraor I = 4.5ω, where it can be clearly seen that theiodic trajectory no longer exists.
.2. Dynamics of the globally coupled network
In the presence of noise and coupling, the respf an individual oscillator to stimulation can still gigreat deal of insight into the behavior of the netwig. 2shows the modulusR(t) of the order parametes a function of time. The vertical axis indicates
nitial phase (normalized to the interval from 0 tot which the stimulation starts, and was varied in sf 0.01. A network of 500 oscillators was used,
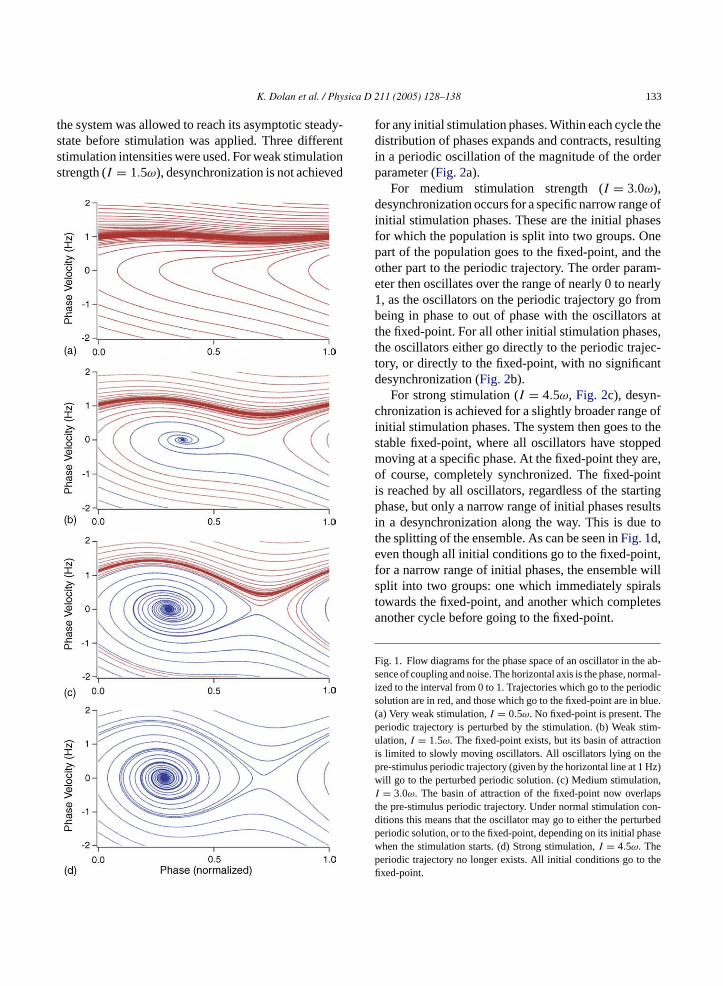
K. Dolan et al. / Physica D 211 (2005) 128–138 133
the system was allowed to reach its asymptotic steady-state before stimulation was applied. Three differentstimulation intensities were used. For weak stimulationstrength (I = 1.5ω), desynchronization is not achieved
for any initial stimulation phases. Within each cycle thedistribution of phases expands and contracts, resultingin a periodic oscillation of the magnitude of the orderparameter (Fig. 2a).
For medium stimulation strength (I = 3.0ω),desynchronization occurs for a specific narrow range ofinitial stimulation phases. These are the initial phasesfor which the population is split into two groups. Onepart of the population goes to the fixed-point, and theother part to the periodic trajectory. The order param-eter then oscillates over the range of nearly 0 to nearly1, as the oscillators on the periodic trajectory go frombeing in phase to out of phase with the oscillators atthe fixed-point. For all other initial stimulation phases,the oscillators either go directly to the periodic trajec-tory, or directly to the fixed-point, with no significantdesynchronization (Fig. 2b).
For strong stimulation (I = 4.5ω, Fig. 2c), desyn-chronization is achieved for a slightly broader range ofinitial stimulation phases. The system then goes to thestable fixed-point, where all oscillators have stoppedmoving at a specific phase. At the fixed-point they are,of course, completely synchronized. The fixed-pointis reached by all oscillators, regardless of the startingphase, but only a narrow range of initial phases resultsin a desynchronization along the way. This is due tothe splitting of the ensemble. As can be seen inFig. 1d,even though all initial conditions go to the fixed-point,for a narrow range of initial phases, the ensemble willsplit into two groups: one which immediately spiralst etesa
F e ab-s rmal-i odics blue.( ep tim-u ioni hep Hz)w ion,I apst con-d rbedp asewp thefi
owards the fixed-point, and another which complnother cycle before going to the fixed-point.
ig. 1. Flow diagrams for the phase space of an oscillator in thence of coupling and noise. The horizontal axis is the phase, no
zed to the interval from 0 to 1. Trajectories which go to the periolution are in red, and those which go to the fixed-point are ina) Very weak stimulation,I = 0.5ω. No fixed-point is present. Theriodic trajectory is perturbed by the stimulation. (b) Weak slation,I = 1.5ω. The fixed-point exists, but its basin of attract
s limited to slowly moving oscillators. All oscillators lying on tre-stimulus periodic trajectory (given by the horizontal line at 1ill go to the perturbed periodic solution. (c) Medium stimulat= 3.0ω. The basin of attraction of the fixed-point now overl
he pre-stimulus periodic trajectory. Under normal stimulationitions this means that the oscillator may go to either the pertueriodic solution, or to the fixed-point, depending on its initial phhen the stimulation starts. (d) Strong stimulation,I = 4.5ω. Theeriodic trajectory no longer exists. All initial conditions go toxed-point.
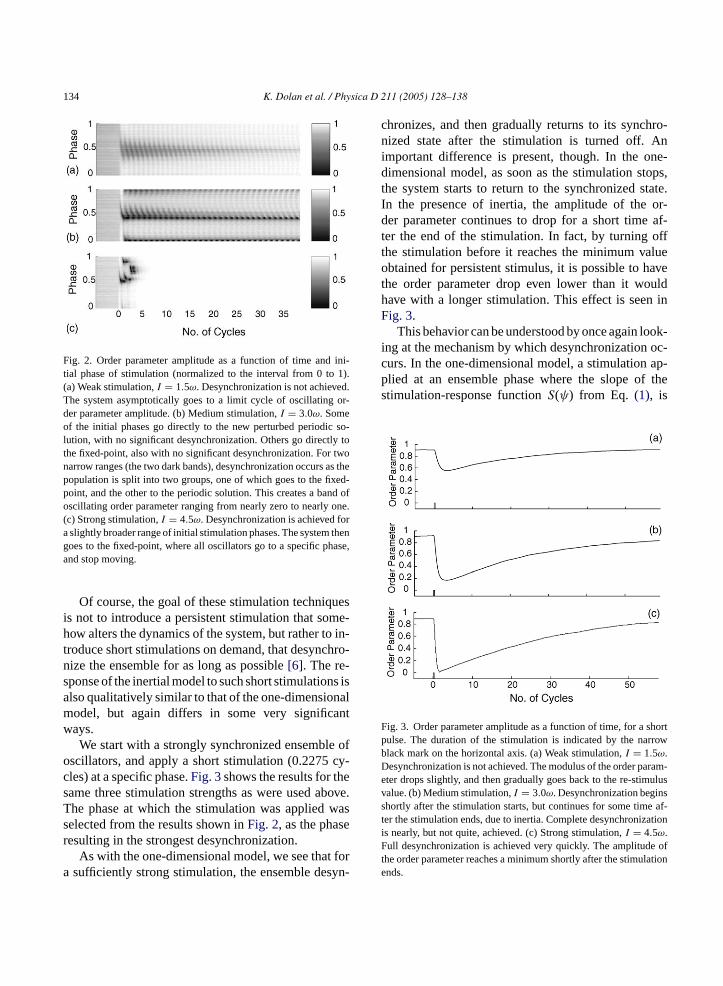
134 K. Dolan et al. / Physica D 211 (2005) 128–138
Fig. 2. Order parameter amplitude as a function of time and ini-tial phase of stimulation (normalized to the interval from 0 to 1).(a) Weak stimulation,I = 1.5ω. Desynchronization is not achieved.The system asymptotically goes to a limit cycle of oscillating or-der parameter amplitude. (b) Medium stimulation,I = 3.0ω. Someof the initial phases go directly to the new perturbed periodic so-lution, with no significant desynchronization. Others go directly tothe fixed-point, also with no significant desynchronization. For twonarrow ranges (the two dark bands), desynchronization occurs as thepopulation is split into two groups, one of which goes to the fixed-point, and the other to the periodic solution. This creates a band ofoscillating order parameter ranging from nearly zero to nearly one.(c) Strong stimulation,I = 4.5ω. Desynchronization is achieved fora slightly broader range of initial stimulation phases. The system thengoes to the fixed-point, where all oscillators go to a specific phase,and stop moving.
Of course, the goal of these stimulation techniquesis not to introduce a persistent stimulation that some-how alters the dynamics of the system, but rather to in-troduce short stimulations on demand, that desynchro-nize the ensemble for as long as possible[6]. The re-sponse of the inertial model to such short stimulations isalso qualitatively similar to that of the one-dimensionalmodel, but again differs in some very significantways.
We start with a strongly synchronized ensemble ofoscillators, and apply a short stimulation (0.2275 cy-cles) at a specific phase.Fig. 3shows the results for thesame three stimulation strengths as were used above.The phase at which the stimulation was applied wasselected from the results shown inFig. 2, as the phaseresulting in the strongest desynchronization.
As with the one-dimensional model, we see that fora sufficiently strong stimulation, the ensemble desyn-
chronizes, and then gradually returns to its synchro-nized state after the stimulation is turned off. Animportant difference is present, though. In the one-dimensional model, as soon as the stimulation stops,the system starts to return to the synchronized state.In the presence of inertia, the amplitude of the or-der parameter continues to drop for a short time af-ter the end of the stimulation. In fact, by turning offthe stimulation before it reaches the minimum valueobtained for persistent stimulus, it is possible to havethe order parameter drop even lower than it wouldhave with a longer stimulation. This effect is seen inFig. 3.
This behavior can be understood by once again look-ing at the mechanism by which desynchronization oc-curs. In the one-dimensional model, a stimulation ap-plied at an ensemble phase where the slope of thestimulation-response functionS(ψ) from Eq. (1), is
Fig. 3. Order parameter amplitude as a function of time, for a shortpulse. The duration of the stimulation is indicated by the narrowblack mark on the horizontal axis. (a) Weak stimulation,I = 1.5ω.Desynchronization is not achieved. The modulus of the order param-eter drops slightly, and then gradually goes back to the re-stimulusvalue. (b) Medium stimulation,I = 3.0ω. Desynchronization beginsshortly after the stimulation starts, but continues for some time af-ter the stimulation ends, due to inertia. Complete desynchronizationis nearly, but not quite, achieved. (c) Strong stimulation,I = 4.5ω.Full desynchronization is achieved very quickly. The amplitude ofthe order parameter reaches a minimum shortly after the stimulationends.

K. Dolan et al. / Physica D 211 (2005) 128–138 135
positive, will result in the phases of the individual os-cillators being pulled apart[6]. For the inertial model,the stimulation acts on the velocities, instead of on thephases. Stimulation when the slope ofS(ψ) from Eq.(6) is positive, results in the spreading of the distribu-tion of velocities. The spreading of the phase distri-bution then follows from the distribution of velocities.The coupling also acts on the velocities, causing oscil-lators which lie ahead of the average phase to be sloweddown, and those lying behind the average phase to besped up. Resynchronization does not begin until thiseffect has had time to cancel out the cumulative effectthat the stimulation has had on the velocity distribution.This means that there is a time lag between when thestimulation is turned off, and when the phases them-selves stop diverging away from each other, and startconverging towards each other again.
This phenomenon complicates the process of de-termining the ideal stimulation length. It is no longersufficient to simply look at how long it takes for a con-stant stimulation to achieve a minimum in the orderparameter amplitude. Instead it is necessary to look athow effective the desynchronization is as function ofboth the stimulation phase, and the stimulation length.
4. Resynchronization and stimulation timing
4.1. Calibration for effective desynchronization
tim-i ula-t pli-t s then ps.T alityo ft mbi-n sulti ap allyw ula-t ont hort,t Thisw ses,w us,n tion
Fig. 4. The average of the amplitude of the order parameter over aperiod of 40 cycles after stimulation. The horizontal axis is the stim-ulation length, and the vertical axis is the phase at which stimulationbegan. The average order parameter amplitude is given by the gray-scale. (a) Weak stimulation,I = 1.5ω. As before, desynchronizationis not achieved for any stimulation length, although the order param-eter does drop slightly for some combinations of stimulation phaseand length. (b) Medium stimulation,I = 3.0ω. Maximum desyn-chronization occurs for a narrow range of both stimulation phaseand stimulation length. (c) Strong stimulation,I = 4ω. Desynchro-nization is achieved for a broader range of stimulation phases andlengths. For long stimulation lengths, resynchronization occurs dueto the oscillators all going to the fixed-point.
length increases, the velocities become more broadlydistributed, allowing for significant desynchronizationto occur after the stimulation stops, but as the lengthcontinues to increase, the network will pass through thephase range where the slope of the stimulus responsefunctionS(ψ) is negative, which will cause the veloc-ities to be pushed back together. Persistent stimulationjust results in the distribution of velocities broadeningand contracting every period, which for the weak stimu-lation is too rapid for any significant desynchronizationto occur. The degree of desynchronization that occursafter the stimulation stops, just depends on the velocitydistribution at that time, hence the periodic dependenceon stimulation length.
For medium stimulation strength (Fig. 4b), strongdesynchronization occurs for a narrow range of ini-tial phases around 0.6, and stimulation length of about0.2 cycles. Outside of this range, initial phases rangingfrom about 0.4 to 1.0 give results which vary periodi-
To address the impact of stimulation length andng on the quality of desynchronization, we use stimions of varying length, and calculate the average amude of the order parameter over a period of 40 timeatural period of oscillation, after the stimulation stohis average provides a measure of the overall quf the desynchronization.Fig. 4 shows the results o
his analysis. We can see that there are specific coations of stimulation strength and length which re
n very strong desynchronization. InFig. 4a we seeattern of desynchronization which varies periodicith stimulation length. This is because the stim
ion operates directly on the velocities, rather thanhe phases. When the stimulation length is very she velocities are only spread by a small amount.ould result in a very gradual spreading of the phahich is quickly canceled out by the coupling. Tho desynchronization is achieved. As the stimula

136 K. Dolan et al. / Physica D 211 (2005) 128–138
cally with stimulation length, as for the weak stimula-tion case, and other initial phases go to the fixed-point,resulting in rapid resynchronization. Optimal desyn-chronization occurs for a short stimulus whose initialphase lies on the transition between these two behav-iors. Once again, this is due to the population being splitinto two groups, one of which tries to go to the fixed-point, and the other towards the periodic solution.
For strong stimulation (Fig. 4c), desynchronizationstill occurs for a narrow range of initial phases around0.6, and stimulation length of about 0.15, althoughthe range is not as narrow as for medium stimula-tion strength. A long enough stimulation always re-sults in the oscillators going to the fixed-point, though.How long this takes also depends on the initial phaseof the ensemble. As can be seen inFig. 1d, there isstill a splitting of the system into two groups, one ofwhich immediately spirals directly into the fixed-point,and the other of which goes through an additional cy-cle first. optimal desynchronization occurs for initialphases which lie near this splitting. For initial phasesin the range of 0.5–1.0, desynchronization can be at-tained for a broad range of longer stimulation lengthsas well. This can also be understood by observing fromFig. 1d that initial phases in this range must go throughseveral cycles before finally going to the fixed-point.
4.2. Resynchronization time as a function of theinduced velocity distribution
hro-n et ofa theo i-t ndt riedt eeh t oft
l syn-c ens,t s be-c withδ thep cou-p uffi-c me
Fig. 5. The amplitude of the order parameter over a period ofn cycles.Resynchronization occurs very slowly for a narrow velocity distribu-tion, and occurs more quickly as the velocity distribution broadens.As the velocity distribution continues to broaden, the resynchroniza-tion time increases again, due to the large amount of surplus energybeing put into the system.
non-zero. This fixed-point is unstable, but when thesystem is near that fixed-point, the net effect of the cou-pling is very small. As the oscillators begin to clustertogether, the effect of the coupling becomes more pro-nounced. Broadening the distribution of velocities firstcauses the clustering of phases to occur more quickly,by providing more opportunities for statistical fluctu-ations in the phase distribution to start the clusteringprocess. For a velocity distribution with standard de-viation of about 0.15 times the natural frequency, theresynchronization time is minimized. As the broadnessof the distribution continues to increase, the resynchro-nization time then grows longer. This is due to the iner-tia in the system. Oscillators which are moving muchfaster than the natural frequency will take a long time toslow down to the natural frequency, which they must doin order for synchronization to be achieved. Likewise,oscillators moving much slower than natural frequency(or even backwards), will take a long time to speed backup to the natural frequency. This frequency lockingmust be achieved before resynchronization can occur,because as long as there is any distribution of velocities,that will tend to broaden the distribution of phases.
5. Discussion
As a first step towards a physiological model of thebrain processes associated with Parkinsonian and es-
To better understand the mechanism of resyncization, a numerical simulation was done using a srtificial initial conditions, such that the phases ofscillators are initially uniformly distributed. The in
ial velocities are given by a normal distribution arouhe natural frequency of the oscillators. We then vahe width of the velocity distribution, in order to sow the distribution of velocities affects the amoun
ime required for resynchronization to occur.We can see fromFig. 5that for aδ-distribution of ve-
ocities, the system takes a relatively long time to rehronize, but as the distribution of velocities broadhe resynchronization time becomes smaller. This iause the uniformly distributed phases, combined-distributed velocities, constitute a fixed-point inhase space of the ensemble. The net effect of theling is zero, and requires the occurrence of a siently strong perturbation (due to noise), to beco

K. Dolan et al. / Physica D 211 (2005) 128–138 137
sential tremor, we have applied basic phase resettingand desynchronization techniques to a network of glob-ally coupled phase oscillators with inertia. While thismodel has been derived as a simplified neuron model,it is also quite general, and should have relevance toany physical system of coupled oscillators in whichphase dynamics are of principle interest, and transientproperties of the individual oscillators must be takeninto account. The introduction of inertia to the modelleaves many of the qualitative aspects of the desynchro-nization process found in the one-dimensional modelintact, but introduces several new phenomena not seenin the simpler model.
A bistability, where both a stable periodic trajectoryand a stable fixed-point are present, plays a critical rolein the desynchronization of the network of inertial os-cillators. This mechanism differs from the desynchro-nization mechanism of the one-dimensional model,where no such coexistence of periodic and fixed-pointbehavior is possible[6]. Even when the bistability isnot apparent because all normal pre-stimulus initialconditions lie in the basin of attraction of the periodictrajectory, the bistability still plays a role in the desyn-chronization, because the critical phases where desyn-chronization occurs are those phases where the initialconditions lie closest to the boundary between the twobasins of attraction. In contrast, in the Kuramoto sys-tem desynchronization occurs when the population ofoscillators is split by the unstable fixed point into twogroups of oscillators which then approach the stablefi
in al op-t thatt appli-c ers.I la-t them syn-c tionp re-q anda uset by as syn-c ro-n timed e os-
cillators are desynchronized, and also on the distribu-tion of phase velocities at that time. Therefore, onemust first find a combination of stimulation strengthand length that causes complete desynchronization, andthen by adjusting those parameters so as to minimizethe broadness of the velocity distribution at the timewhen desynchronization occurs, one can achieve opti-mal desynchronization.
Another distinction between the inertial phase os-cillator, and the one-dimensional model, is that for astrong enough short stimulus, desynchronization canalways be achieved, regardless of initial phase. Forweak and intermediate stimulation strengths, desyn-chronization normally occurs when the ensemble is ata phase where the stimulation response function,S(ψ)from Eq.(6), is positive. This is because the leading os-cillators in the ensemble get accelerated, and the trail-ing oscillators get slowed down, thus pulling the en-semble apart. When the slope is negative, the oppositeeffect occurs. The leading oscillators slow down, andthe trailing oscillators speed up. For weak and interme-diate stimulation strengths, this causes the ensemble tobecome more strongly synchronized. This is exactlyanalogous to the situation with the one-dimensionalmodel. The difference is that in the one-dimensionalmodel, the stimulus is directly acting on the phases,but in the inertial model, it is acting directly on the ve-locities. For a very strong stimulus, even when the slopeof S(ψ) is negative, the stimulation will still widely dis-tribute the velocities. In this case, the leading oscillatorsg ve-l uchf sys-t chro-n n bes alp iticali ccuri tioni ses,t nks,a
twop neu-r f theb theK lagb hen
xed point from opposite directions[6].The inertia inherent in the model also results
ag between when the stimulation stops, and whenimal desynchronization is achieved. This meanshe standard phase resetting curves are no longerable for calibration of optimal stimulation paramett is not sufficient to simply locate the ideal stimuion phase, and then shut off the stimulation whenaximum desynchronization occurs. Complete de
hronization can still be achieved, but the calibrarocedure for finding the stimulations parametersuires finding both an optimal stimulation phasen optimal stimulation duration. Furthermore, beca
he state of the system can not be characterizedingle parameter, simply achieving complete dehronization does not imply that optimal desynchization has been achieved. The resynchronizationepends both on how completely the phases of th
et slowed down to much slower than the averageocity, and the trailing oscillators get sped up to master than the average velocity. The inertia of theem then causes the phases to go right past the synized state, and then desynchronize. This effect caeen inFig. 4(c), where only a small range of initihases do not lead to desynchronization. That cr
nitial phase where desynchronization does not os where the slope of the stimulation response funcs close to zero. As the simulation strength increahe range of phases around this critical point shrind disappears.
The identification of these new properties servesurposes. First, since inertia is a property of realons, it can provide relevant insight into aspects oiological system that would not be realizable withuramoto model. For example, the potential for aetween when the stimulation is stopped, and w

138 K. Dolan et al. / Physica D 211 (2005) 128–138
maximal desynchronization occurs, could play a cru-cial role in situations where other types of delay arepresent as well. Second, it provides a framework fromwhich the next step can be taken. When further proper-ties like amplitude dynamics and nonlinearity are intro-duced, it will be easier to determine which new behav-iors are the result of these new properties, and which aredue to the inertial properties inherent in such systems.
Acknowledgements
The authors would like to thank Lutz Schimansky-Geier for very useful discussions. This study was sup-ported by the Volkswagen Foundation (76761), theGerman Israeli Foundation (I-667-81.1/2000), and theDAAD (D/0247269).
References
[1] H. Haken, Advanced Synergetics, Springer, Berlin, 1983.[2] Y. Kuramoto, Chemical Oscillations, Waves and Turbulence,
Springer, Berlin, 1984.[3] A.T. Winfree, The Geometry of Biological Time, Springer,
Berlin, 1980.[4] J.D. Murray, Mathematical Biology, Springer, Berlin, 1989.[5] H. Steriade, et al., Thalamic Oscillations and Signaling, John
Wiley & Sons, New York, 1990.[6] P.A. Tass, Phase Resetting in Medicine and Biology. Stochastic
Modelling and Data Analysis, Springer, Berlin, 1999..
s of
[[
[12] P.A. Tass, Europhys. Lett. 57 (2002) 164.[13] R. Llinas, H. Jahnsen, Nature 297 (1982) 406.[14] D. Pare, R. Curro’Dossi, M. Steriade, Neuroscience 35 (1990)
217.[15] F.A. Lenz, H.C. Kwan, R.L. Martin, R.R. Tasker, J.O. Dostro-
vsky, Y.E. Lenz, Brain 117 (1994) 531.[16] A. Nini, A. Feingold, H. Slovin, H. Bergman, J. Neurophysiol.
74 (4) (1995) 1800.[17] A.L. Benabid, P. Pollak, C. Gervason, D. Hoffmann, D.M. Gao,
M. Hommel, J.E. Perret, J. DeRougemont, Lancet 337 (1991)403.
[18] S. Blond, D. Caperros-Lefebvre, F. Parker, R. Assaker, H. Petit,J.-D. Guieu, J.-L. Christiaens, J. Neurosurg. 77 (1992) 62.
[19] C.C. McIntyre, M. Savasta, L. Kerkerian-Le Goff, J.L. Vitek,Clin. Neurophysiol. 155 (2004) 1239.
[20] A.L. Benabid, A. Benazzous, P. Pollak, Mov. Disord. 17 (2002)73.
[21] W.C. Koller, K.E. Lyons, S.B. Wilkenson, A.I. Troster, R.Pahwa, Mov. Disord. 16 (3) (2001) 464.
[22] M.I. Hariz, Mov. Disord. 17 (Suppl. 3) (2002) S162–S166.[23] P.A. Tass, Biol. Cybern. 85 (2001) 343.[24] P.A. Tass, Biol. Cybern. 87 (2002) 102.[25] P.A. Tass, Phys. Rev. E 66 (2002) 036226.[26] P.A. Tass, Biol. Cybern. 89 (2003) 81.[27] P. Anderson, J.C. Eccles, Y. Lowning, J. Neurophys. 27 (1964)
592.[28] H.A. Braun, K. Schafer, K. Voigt, R. Peters, F. Bretschneider,
X. Pei, L. Wilkins, F. Moss, J. Comp. Neurosci. 4 (1997) 335.[29] H.A. Braun, M. Dewald, K. Schafer, K. Voigt, X. Pei, K. Dolan,
F. Moss, J. Comp. Neurosci. 7 (1999) 17.[30] C. Hauptmann, M.C. Mackey, Biol. Cybern. 88 (2003) 459.[31] H. Haken, Eur. Phys. J. B 18 (2000) 545.[32] H. Haken, Brain Dynamics, Springer-Verlag, Berlin, 2002.[33] G.B. Ermentrout, N. Kopell, J. Math. Biol. 29 (1991) 195.[34] E.R. Grannan, D. Kleinfeld, H. Sompolinsky, Neural Comp. 4
[ 367.[[[ eu-
[7] W. Singer, C.M. Gray, Annu. Rev. Neurosci. 18 (1995) 555[8] A.T. Winfree, J. Theor. Biol. 28 (1970) 327.[9] L. Glass, M.C. Mackey, From Clocks to Chaos. The Rythm
Life, Princeton University Press, 1988.10] P.A. Tass, Europhys. Lett. 53 (2001) 15.11] P.A. Tass, Europhys. Lett. 55 (2001) 171.
(1993) 550.35] D. Hansel, G. Mato, C. Meunier, Europhys. Lett. 23 (1993)36] J.A. Acebron, R. Spigler, Phys. Rev. Lett. 81 (1998) 2229.37] L.L. Bonilla, Phys. Rev. E 62 (2000) 4862.38] H. Bergman, T. Wichmann, B. Karmon, M.R. DeLong, J. N
rophysiol. 72 (1994) 507.

![Desynchronization and sustainability of noisy ... · arXiv:q-bio/0701032v1 [q-bio.PE] 20 Jan 2007 Desynchronization and sustainability of noisy metapopulation cycles Refael Abta1,](https://img.pdfslide.us/doc/110x75/5e6b40e71bafa44ca0602752/desynchronization-and-sustainability-of-noisy-arxivq-bio0701032v1-q-biope.jpg)



























