Embed Size (px)
Citation preview

Vol. 4, 611-618, Marc/i 1998 Clinical Cancer Research 611
Phase lb Trial of Bryostatin 1 in Patients with Refractory
Malignancies1
Steven Grant,2 John Roberts, Elizabeth Poplin,
Mary Beth Tombes, Brenda Kyle, Dawn Welch,
Marc Carr, and Harry D. Bear
Departments of Medicine [S. G.. J. R.. E. P.. M. B. T.. B. K.. M. C.],Surgery [H. D. B.l. Biostatistics [D. W.], Microbiology andImmunology [S. 0.. H. D. B.], and Pharmacology [S. G.1, VirginiaCommonwealth University. Richmond, Virginia 23298
ABSTRACT
A Phase lb trial of bryostatin 1, a macrocyclic lactoneand protein kinase C (PKC) activator, was conducted inpatients with refractory nonhematological malignancieswith the primary goal of determining whether down-regu-
lation of peripheral blood mononuclear cell (PBMNC) PKCactivity could be achieved in vivo in humans. Patients (four
patients/cohort) received bryostatin 1 (25 jig/m2) as a 1-h
infusion weekly three times every 4 weeks, but to study the
schedule dependence of pharmacokinetics and pharmacody-
namics, the first dose was administered according to one of
three schedules: (a) a 1-h infusion; (b) a 24-h infusion; or (c)
a split course (12.5 �iWm2 as a 30-mm infusion) on days 1and 4. Conventional toxicities (grades I-Ill) included myal-gias, fever, anemia, fatigue, phlebitis, and headache; in ad-
dition, two patients in cohort 3 experienced transient eleva-
tions in liver function tests, although these patients had
preexisting liver metastases. No objective clinical responses
were encountered. Effects on PBMNC PKC activity wereheterogeneous. Several patients in cohorts 1 and 2 expen-
enced significant declines in activity (-50%) that were sus-tamed in some cases for periods of �72 h. Comparison of
72-h with baseline values for all three patient cohorts corn-
bined revealed a trend toward PKC down-regulation (P =
0.06; signed rank test). For each schedule, plasma bryostatin
1 levels were below the level of detection of a platelet aggre-gation-based bioassay (3-4 nM). Bryostatin 1 administration
failed to produce consistent alterations in lymphocyte im-munophenotypic profiles, interleukin 2-induced prolifera-
Received 5/30/97: revised I 1/25/97: accepted 12/19/97.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadt’ertise,nent in accordance with I 8 U.S.C. Section 1734 solely toindicate this fact.
I Supported by NIH Grants R03 CA66990 and RO1 CA63753 and
Award P30 CA16059 to the Massey Cancer Center and in part by NIHGeneral Clinical Research Center Grant MO1 RR00065. Portions of thiswork were presented in preliminary form at the AACR Annual Meeting,Washington, D.C., April 20-24, 1996.
2 To whom requests for reprints should be addressed, at Division ofHematology/Oncology, Massey Cancer Center, Medical College of Vir-ginia. Virginia Commonwealth University. P. 0. Box 980230, Rich-
mond, VA 23298-0230. Phone: (804) 828-5211: Fax: (804) 828-8079.
tion, or cytotoxicity, although two of three samples from
patients in cohort 3 did show significant posttreatrnent in-
creases in proliferation. Moreover, in some patients, bryo-
statin 1 treatment increased lymphokine-activated killer cell
activity. These findings indicate that bryostatin 1 doses of 25
�ig/rn2 can induce in vivo PBMNC PKC down-regulation inat least a subset of patients and raise the possibility that
higher bryostatin 1 doses may be more effective in achievingthis effect.
INTRODUCTION
Bryostatin 1 is a macrocyclic bactone derived from the
marine bryozoan, Bugula neritina ( 1 ). In addition to immuno-
modulatory actions (2), bryostatin 1 has been shown to exert in
vitro and in vivo activity against a variety of experimental tumor
types, including carcinomas of the breast (3) and lung (4),
melanoma (5), bymphoma (6), and leukemia (7), among others.
In contrast to its inhibitory effects toward neoplastic cells.
bryostatin 1 stimulates the in vitro growth of normal hemato-
poietic progenitors (8), possibly through an indirect mechanism
(9). Based on these and other properties, bryostatin I has been
targeted for clinical development by the National Cancer Insti-
tute, and several Phase I trials have now been completed. When
administered as a weekly 1-h bolus or 24-h infusion, the max-
imum tolerated dose has been reported as 25-50 �ig/m2 (10-
13). Myalgias have been the major dose-limiting toxicity; other
side effects have included fevers, anemia, leukopenia, fatigue,
phlebitis, headaches, dyspnea. hypotension, and bradycardia
(10-13). Limited responses have been observed in patients with
ovarian carcinoma and lymphoma ( 12). Bryostatin I also exerts
in vivo immunomodulatory actions, enhancing expression of
lymphocyte IL-23 receptors and selectively stimulating CD8
cells (13).
Although the mechanism by which bryostatin 1 exerts its
antineoplastic actions remains unknown, considerable evidence
suggests the involvement of the Ca2�- and lipid-dependent
serine-threonine kinase PKC. In in vitro studies using nanomo-
lar concentrations, bryostatin 1 has been shown to bind to and
activate PKC (14). This results in translocation ofthe enzyme to
the membrane, accompanied by depletion of cytosolic activity
(15). In this regard, bryostatin 1 mimics, to a certain extent, the
actions of tumor-promoting phorboids such as phorbol 12-my-
ristate 13-acetate, although its spectrum of activity is distinctly
different from such compounds. For example, bryostatin 1
blocks those phorboid-associated actions that it does not pos-
sess, including tumor promotion in mouse skin ( 1 6) and induc-
tion of differentiation in some leukemic cell sublines ( 15, 17).
3 The abbreviations used are: IL, interleukin: PKC, protein kinase C:PBMNC, peripheral blood mononuclear cell; ara-C, l-�3-o-arabino-furanosylcytosine: LAK, lymphokine-activated killer cell: Ab, antibody.
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

612 Phase lb Trial of Bryostatin 1
4 5. Grant and H. D. Bear, unpublished observations.
The basis for the unique actions of bryostatin 1 is obscure but
may stem from specific patterns of PKC isoform activation (18)
or nuclear transbocation ( 19). In addition, prolonged exposure of
cells to bryostatin 1 (e.g., for 24 h) beads to profound reduction
in total cellular PKC activity (20), possibly as a consequence of
proteosomal degradation after ubiquitination (21). This phe-
nomenon, referred to as down-regulation, is distinct from the
early loss of cytosolic activity accompanying PKC activation,
which results from enzyme transbocation to the cell membrane
(22).
Bryostatin 1 has been shown to modulate the apoptotic
response of human leukemia cells to the antimetabolite ara-C
(23). For example, pretreatment of human promyebocytic beu-
kemia cells (HL-60) with bryostatin 1 substantially increases
their susceptibility to ara-C-induced apoptosis, an event tempo-
rally associated with PKC down-regulation (24). This finding is
consistent with reports that PKC activation opposes apoptosis
(25, 26), whereas PKC inhibition promotes cell death (27, 28).
In light of evidence that various chemotherapeutic drugs, in-
cluding ara-C, induce leukemic cell apoptosis in vitro and in
vivo (29, 30), it seems plausible that prior administration of
bryostatin 1 might increase the efficacy of certain antileukemic
agents.
To be effective, this strategy would require that bryostatin
I exert its in vitro actions, i.e. , PKC down-regulation, in vivo.
Currently, little is known about the in vivo effects of bryostatin
I on PKC activity. Recently, we reported that CS7BL/6 mice
receiving a single iv. bolus injection of bryostatin 1 (1 p.g)
experienced significant and sustained PKC down-regulation in
normal splenocytes, an effect accompanied by splenic enlarge-
ment (3 1 ). We have observed that in vitro exposure of normal
PBMNCs to bryostatin 1 results in PKC down-regulation virtu-
ally identical to that displayed by HL-60 cells.4 Thus, PBMNCs
may serve as a surrogate hematopoietic target tissue capable of
mimicking the response of leukemic blasts to this agent. To
assess the potential clinical relevance of these preclinical find-
ings, a Phase lb trial of bryostatin 1 has been conducted in
patients with advanced malignancies. The primary goal of this
trial was to determine whether and to what extent in vivo
administration of bryostatin 1 modulates PKC activity in human
hematopoietic cells.
MATERIALS AND METHODS
Study Design. A fixed dose of bryostatin 1 (25 p.g/m2)
was studied on the basis of experience from previous Phase Ia
trials ( 10-13). Patients received bryostatin 1 (25 pg/m2) as a 1-h
infusion weekly X 3 every 4 weeks; however, the first dose was
administered by random assignment to one of three schedules:
(a) 1-h iv. infusion; (b) 24-h iv. infusion; or (c) 12.5 p.g/m2 as
a 0.5-h iv. infusion on days 1 and 4.
Bryostatin 1 Source and Administration. Bryostatin 1
(NSC 339555) was supplied by the Cancer Treatment and
Evaluation Program, Division of Cancer Treatment, National
Cancer Institute. It was stored at 4#{176}Cin flint vials containing 0.1
mg of bryostatin 1 and S mg of povidone USP lyophilized from
40% t-butanol. Material was reconstituted in 1 ml of sterile PET
(60% polyethylene glycob 400, 30% ethanol, and 10% Tween
80) diluent. The resulting solution was diluted further with 9 ml
of 0.9% sodium chloride. Tubing for the drug infusion was
primed with bryostatin 1 in the PET formulation diluted with
sodium chloride to minimize adsorption of drug. For infusions
of � 1 h, bryostatin 1 was administered by syringe pump via a
peripheral venous line. For 24-h infusions, bryostatin 1 was
administered by syringe pump via either a peripheral venous or
central line with coinfusion of 2 biters of normal saline.
Patient Eligibility. All patients were � 1 8 years of age
and had histologically confirmed lymphoma or solid tumor
malignancy refractory to standard treatment. Other eligibility
criteria included a life expectancy � 12 weeks, Zubrod perform-
ance status � 2, the presence of measurable disease, the absence
of primary or metastatic malignant central nervous system dis-
ease, and no chemotherapy or radiotherapy within 4 weeks of
planned therapy. Laboratory criteria included a WBC in the
range 3.7-15.0 x l034t1, platelets in the range 130-500 X
103/lib, hemoglobin � 10 g/dl, normal protime, bilirubin � 1.5,
normal aspartate aminotransferase and alanine aminotransferase
(�2.5 x the upper limit of normal for metastatic disease), and
creatinine � 1 .5 or creatinine clearance > 70. Pregnant or
nursing patients were excluded from the study.
Toxicity Grading. All patients received an initial evalu-
ation that included a complete physical examination, serum
chemistries, complete blood count, protime, and urinalysis. lox-
icities were graded on a scale of 0-4 according to Cancer and
Acute Leukemia Group B common toxicity criteria, with the
exception of myalgias, which were evaluated based on criteria
previously described by Philip et a!. (10).
Response Criteria. The following criteria were used to
assess the responses of patients to bryostatin 1 : (a) complete
response, absence of all measurable disease, signs, symptoms,
and biochemical changes related to the tumor, including recal-
cification of all lytic bone lesions, for >4 weeks and no new
lesions; (b) partial response, �50% reduction in the sum of the
products of the perpendicular diameters of all measurable le-
sions compared to baseline measurements persisting for �4
weeks, no enlargement of an existing lesion, and no new lesions;
(c) stable disease, neither response nor progression for >8
weeks; and (d) progression of disease, an increase in the product
of the perpendicular diameters of any measured lesion by �25%
compared to baseline measurement or any new lesion.
PBMNC PKC Activity. At designated intervals (prein-
fusion and 1, 8, 24, 48, 72, 96, and 120 h postinfusion), 20 ml
of blood were collected in a syringe containing EDIA (Sigma
Chemical Co., St. Louis, MO) and stored at 4#{176}Cfor not more
than 20 mm before processing. Samples were then diluted 1:2
with sterile RPMI 1640 (Life Technologies, Inc., Grand Island,
NY) and layered over a lO-ml cushion of Ficoll-Hypaque (spe-
cific gravity, 1.077-1.081) in 50-mI sterile polypropylene con-
ical centrifuge tubes. The tubes were centrifuged at 400 X g at
room temperature according to the method of Boyum (32). At
the end of this period, the interface layer containing mononu-
clear cells was extracted with a sterile pipette, transferred to
15-mb plastic centrifuge tubes, and washed twice with fresh
medium.
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Clinical Cancer Research 613
Total PKC activity in PBMNC was determined as de-
scribed previously (33) using materials supplied by Life Tech-
nologies, Inc. Briefly, pebleted cells were homogenized in 2 X
10_2 M Iris, S X iO-� M EDIA, and 5 x l0� M EGTA (pH
7.5) containing 25 pg/ml protease inhibitors (aprotinin and
leupeptin), with thirty-five b-s strokes of a motorized mi-
cropestle homogenizer (Glas-Col) at setting 35. The homoge-
nate was incubated on ice for 30 ruin, after which normalized
quantities of protein ( 15 pg) were added to an assay mixture
containing mixed micelbes of phosphatidylserine and phorbol
12-myristate 13-acetate in suspension. The reaction was initi-
ated by addition of 2.5 X b0�� Ci/ml [-y-32P]ATP, 2 X l0� M
nonisotopic AlP, and 5 X b0� M synthetic peptide substrate
(acetylated myelin basic protein amino-terminal peptide
AcMB4_14) in a buffer containing 1 mrvi CaCI, as per the
manufacturer’s instructions. After a 5-ruin incubation at 30#{176}C,
aliquots of the reaction mixture were transferred to phosphocel-
bubose filters, and the reaction was terminated by immersion of
discs in cold 1% (v/v) phosphoric acid. Preliminary studies
established that this reaction was linear over at least a 30-mm
interval. The discs were washed thoroughly, and radioactivity
was quantified by conventional liquid scintilbography. With this
method, replicate determinations generally did not vary by more
than 15%. Baseline pretreatment PKC values for cells obtained
from the 12 patients studied ranged from 1705-4136 prnol
myelin basic protein phosphorybated/mg protein/S mm.
Bryostatin 1 Plasma Assay. Samples were collected in
tubes containing EDTA and centrifuged at 400 X g for 10 ruin
at 4#{176}C,generally within S mm of collection, after which the
plasma was extracted with a sterile Pasteur pipette and trans-
ferred to sterile polypropylene centrifuge tubes. All assays were
performed within 30 mm of blood collection. Bryostatin 1
plasma levels were measured by addition of aliquots of patient
plasma samples to donor whole blood and monitoring AlP
release as described previously (67). Plasma samples (450 p.1)
were mixed with 450 p.1 of whole blood, and after S ruin at room
temperature, 450 p.1 were added to 450 p.1 of 0. 15 M NaCI in an
aggregometer cuvette. Platelet AlP release was measured using
Chrono-Lume reagent and a Chrono-bog whole blood lumi-
aggregometer. Standard curves using known quantities of au-
thentic bryostatin 1 were assayed in parallel and used to deter-
mine concentrations of bryostatin 1 present in plasma samples.
Alternatively, parallel studies were performed assaying the lag
phase of platelet aggregation as described previously (34).
These methods permit detection of plasma bryostatin I levels at
concentrations as bow as S n�i (34).
IL-2 Responsiveness and Lymphocyte Markers. Sam-ples of whole blood were obtained from each patient before
treatment with bryostatin I and at intervals of 24, 72, and 144 h
after initiation of treatment. In cohort 3 (days 1 and 4), day 4
samples were obtained after the second infusion of bryostatin 1.
After density gradient centrifugation as described above, mono-
nuclear cells were harvested, washed, and frozen in FCS + I 0%
DMSO (Sigma). Subsequently, samples were thawed, washed,
and resuspended in complete medium consisting of RPMI 1640,
10% FCS (Hycbone, Logan, UT), 100 units/ml penicillin, 100
p.g/ml streptomycin, 2 msi L-glutamine, 1 m�i sodium pyruvate,
0. 1 msi nonessential amino acids, 10 mtvi HEPES buffer, and
5 x i�-� M �3-mercaptoethanol (Sigma). For proliferation as-
says, cells were plated in triplicate at l0� cells/well in a total
volume of 0.2 ml in 96-well plates (Costar, Cambridge, MA)
with or without 40 lU/mI recombinant IL-2 (Chiron, Emeryville,
CA). After 3 days of culture at 37#{176}Cin a 5% CO,. fully
humidified atmosphere, 0.05 mCi of [3H]thyrnidine (Amer-
sham, Arlington Heights, IL) was added to each well. After an
8-h incubation, cells were harvested onto glass fiber filters using
a PHD Harvester (Cambridge Biotechnology, Inc., Cambridge,
MA), and radioactivity was quantified by liquid scintillography.
Cell samples were also plated at 106 cells/well in 24-
well plates (Costar) under the following conditions: (a) 40
lU/mI IL-2; (b) wells precoated with anti-CD3 monoclonal
Ab (OKT3; Ortho, Raritan, NJ) at 1 p.g/ml, after which the
cells were washed and cultured in the presence of 40 IU/ml
IL-2 as described above; and (c) 500 IU/ml IL-2 alone to
stimulate LAKs. After a 3-day incubation at 37#{176}C.the cells
were harvested, washed, and assayed for cytotoxicity against
target cells using a 4-h 5tCr release assay as described
previously (35). Target cells tested included the P815 murine
mastocytoma, the murine lymphocytic lymphoma EL-4, and
human Daudi cells. Assays involving P815 cells, which have
cell surface Fc receptors, were performed with and without
addition of anti-CD3 monocbonab Ab (1 p.g/ml); in the pres-
ence of anti-CD3, CILs mediate Ab-redirected cytotoxicity
against Fc receptor + targets (36).
For analysis of cell surface markers, PBMNCs were stained
with fluorescein-labeled antibodies directed against CD3, CD4,
CD8, CD2S, CDS6, and DR (MHC class II) and analyzed using
a FACScan analyzer (Becton Dickinson, Sunnyvale, CA) as
described previously (37).
Informed Consent. The study was conducted in accord-
ance with local and federal regulations for the protection of
human research subjects, and all patients offered an informed
consent before enrollment.
Statistical Analysis. PBMNC PKC data for cohorts as a
whole were analyzed by the signed rank test. The significance of
differences between PKC values at individual time points (rel-
ative to baseline control bevels) was determined using Student’s
t test for unpaired observations.
RESULTS
Patient Characteristics. The characteristics of the pa-
tients participating in this trial are summarized in Table 1 . The
I 2 patients included 7 males and 5 females. The median age was
55 years, with a range of 39-76 years. Eastern Cooperative
Oncology Group performance status was 0 in 2 patients and 1 in
10 patients. Disease types and previous therapy were as shown
in Table 1.
Ioxicities. All patients entered on study were available
for evaluation of toxicity. Table 2 lists the mean number of
cycles administered and the associated side effects. In no patient
was treatment discontinued due to bryostatin 1-rebated toxicity.
Patients treated initially according to schedule I (1-h bolus
infusion) experienced myalgias (three of four patients), anemia
(two of four patients). fever (two of four patients), and pruritus
(one of four patients). In all cases, toxicity was grade II or lower.
Myalgias occurred during both early and late treatment cycles
and were responsive to standard analgesic therapy.
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

614 Phase lb Trial of Bryostatin 1
Table 1 Patient characteristics
Patient Age (yr) Gender Tumor type Prior treatment
I 49 M Pancreatic cancer Surgery, chemotherapy
2 62 M Pancreatic cancer No prior treatment
3 69 M Melanoma Surgery, chemotherapy4 59 F Non-small cell lung cancer Chemotherapy, radiotherapy
5 62 F Non-small cell lung cancer Chemotherapy, radiotherapy
6 39 F Breast cancer Surgery, chemotherapy, radiotherapy7 54 M Pancreatic cancer Surgery, chemotherapy, hormonal therapy
8 56 F Breast cancer Surgery, chemotherapy, radiotherapy, hormonal therapy, immunotherapy9 66 M Kidney cancer Surgery, chemotherapy
10 44 F Colon cancer Chemotherapy1 1 49 M Melanoma Surgery, chemotherapy, immunotherapy12 43 M Soft tissue sarcoma No prior treatment
Table 2 Toxicities observed
Type of No. ofCohort toxicity Side effect Grade subjects
Cohort 1 HematologicGeneral
AnemiaFeverMyalgia
Pruritus
�2s2�2
I
223
1Cohort 2
Cohort 3
Hematologic
General
GastrointestinalHeartNeurobogic
MetabolicHematologicGeneral
Liver
AnemiaLeukopeniaThrombocytopeniaFeverMyalgiasPhlebitisFullness (head)Performance statusAnorexiaCardiac ectopyMotorHeadacheHypocalcemiaAnemiaFeverMyalgias
Bilirubin
TransaminaseAlkaline phosphatase
�21I
�222I
�3�2
I21122
�2
3
�2�3
41I3111221111113
1
22
Patients initially receiving bryostatin 1 as a 24-h continu-
ous infusion also experienced grade 2 or lower myalgias (one of
four patients), fever (three of four patients), and anemia (four of
four patients). The anemia did not require transfusions and
resolved after discontinuation of treatment. In addition, two of
four patients experienced significant fatigue, leading to a decline
in performance status (to 3) in one patient (grade 3 toxicity).
This symptom resolved after the bryostatin 1 was discontinued
as a consequence of disease progression. Other toxicities en-
countered in this cohort were of grade 2 or lower and included
leukopenia, anorexia, ectopy, thrombocytopenia, phlebitis,
headache, and neuromotor dysfunction.
Patients receiving bryostatin 1 as a 0.5-h infusion (12.5
p.g/m2) on days 1 and 4 (schedule 3) also experienced grade 2
or lower myalgias (three of four patients), fever (one of four
patients), and anemia (one of four patients). In addition, two
patients in this cohort developed grade 3 liver function abnor-
malities, including hyperbilirubinemia (one of four patients) and
elevated alkaline phosphatase bevels (one of four patients).
Grade � II liver function abnormalities included elevated as-
partate aminotransferase (two of four patients), alanine amin-
otransferase (two of four patients), and alkaline phosphatase
(one of four patients). The latter toxicities were self-limited and
resolved following drug discontinuation after disease progres-
sion. It should be noted that each of the patients exhibiting liver
function abnormalities had preexisting liver metastases.
Responses. There were no objective complete or partial
responses in this group of patients. Three patients (with carci-
noma of the pancreas, breast, and soft tissue sarcoma) exhibited
stable disease during treatment that persisted for at beast 8 weeks
before disease progression was noted. Eight patients developed
disease progression during or after the first cycle of therapy, and
one patient elected to discontinue treatment before being eva]-
uated.
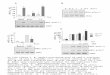
PBMNC PKC Activity. The effects of bryostatin 1 on
PBMNC PKC activity are shown in Fig. 1. Patients receiving
bryostatin 1 as a b-h bobus infusion (schedule 1) displayed an
initial increase in PBMNC PKC activity immediately after drug
administration. By 24 h, a decline in PBMNC PKC activity of
50% was observed in two of four patients and -30% in a third
(P � 0.05 in each case). Ihese declines were sustained for
72-120 h, a pattern similar to that previously reported in an
animal model (31). In one patient, PBMNC PKC activity re-
mained elevated at 24 h, fell to pretreatment control levels at
72 h, and subsequently showed a delayed increase in activity
before declining to baseline at 144 h.
Patients receiving bryostatin 1 as a 24-h continuous infu-
sion (schedule 2) showed a heterogenous pattern of PBMNC
PKC activity. Responses at 24 h ranged from a significant
decline in activity (-65%) in one patient, a more modest decline
(-30%) in a second (P � 0.05 in each case), no change in a
third, and a 100% increase in the fourth. The batter patient
subsequently displayed a late decline in activity that approached
50% at 96 h (P � 0.05). Activity returned to control levels at
72-120 h in all but the first patient, whose cells displayed a
more prolonged recovery phase.
Patients receiving bryostatin 1 as a split dose (12.5 p.g/m2
on days 1 and 4; schedule 3) also exhibited a heterogeneous
response. In one patient, a modest decline in PBMNC PKC
activity was observed postinfusion on day 1 (-40%; P � 0.05),
whereas an increase was noted in one of four patients. At 24 h,
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Schedule #1(25 p.gIm2; 1-hr infusion)
-.-- Cohort #1
-a-- Cohort #2
-.-- Cohort #3
V.C.9
x
S
C.,
Hours
Schedule #2(25 p.g/m2; 24-hr infusion)
C.)
0.
� t
C.)
0.
�
0.
C.)
0.
0.
Hours post-treatment
-0- Patient #1
-6- Patient #2
-9-- Patient #3
-.-- Patient #4
Patent #1-o’- Pa�ent #2
Patient #3
Patient #4
-a- Patient #1-a-- Patient #2
-9-- Patient #3
-.-- Patient #4
Hours
Schedule #3(12.5p.9/rn2; dl,d4)
0 1 2 3 424 48 72 96 120 144
Hours
activity declined modestly (-30%) in one patient (P � 0.05),
remained unchanged in two patients, and increased (-60%) in
a fourth. After the second dose of bryostatin 1, PBMNC PKC
activity did not change appreciably in three of four patients and
increased in one patient. In this patient, PBMNC PKC activity
remained elevated at the 144-h interval.
Clinical Cancer Research 615
Fig. 1 Three cohorts of patients (four patients/cohort) were treatedwith bryostatin 1 (25 �ig/m2) according to three schedules as describedin the text. At the indicated intervals, blood samples were obtained, andmononuclear cell populations were isolated by density centrifugation.After cell disruption, PKC activity was determined by monitoring phos-phorylation of myelin basic protein using a commercially available kit.Values correspond to a percentage of pretreatment controls and areexpressed as the means ± SD for triplicate determinations. Baseline(pretreatment) PKC values for samples obtained from the 12 patientsvaried from 1706 ± 1 15 to 4136 ± 216 pmol phosphorybated myelinbasic protein/mg protein/S mm. *. significantly different from T0 values(P � 0.05).
Fig. 2 Tritiated thymidine uptake by IL-2-stimulated peripheral bloodlymphocytes was monitored in 10 of I 2 patients at the designatedintervals after bryostatin 1 administration as described in the text.Symbols denote the individual patient cohorts. Data are expressed asmean cpm for triplicate wells for each sample after incubation inmedium containing 40 lU/mi IL-2. SDs (data not shown) were < 10% ofthe means in all cases.
For the three patient cohorts as a whole, comparison of 72 h
with baseline PBMNC PKC activities showed a trend toward
down-regulation (P = 0.06; signed rank test). However, no
significant differences among the three cohorts could be iden-
rifled.
Pharmacokinetics Studies. Bryostatin 1 plasma bevels
were undetectable in samples obtained during or after infusion
of bryostatin 1 for all of the schedules used. Based on the level
of sensitivity of the platelet aggregation-based assay (34), this
finding suggests that at the 25 p.g/m2 bryostatin 1 dose level,
plasma bryostatin 1 levels are � S flM.
IL-2 Response and Lymphocyte Markers. Analysis of
phenotypic markers failed to demonstrate consistent changes in
the surface markers of PBMNC (data not shown). For example,
1-25% of PBMNC were CD25+ before treatment, and this
value decreased in 4 of 1 1 patients tested, increased in 2 of 1 1
patients, and remained the same in S of 1 1 patients. In addition,
DR expression, observed in 7-25% of PBMNC before treat-
ment, did not change significantly after administration of bryo-
statin 1 , nor did the expression of other phenotypic markers.
Comparison of phenotypes among different patient cohorts also
did not reveal consistent patterns (data not shown).
When IL-2-stimulated proliferation was assessed, no sig-
nificant change was detected in 7 of the 10 samples assessed at
any time after administration of bryostatin 1 (Fig. 2). However,
in three of the specimens assayed, increases in [3H]thymidine
incorporation ranging from 100-1000% were noted (Fig. 2).
Interestingly, two of the three patients demonstrating an increase
in IL-2-stimulated proliferation were in cohort 3 (split dose),
whereas the third patient was treated with a 1-h infusion of
bryostatin 1.
Lastly, cytotoxicity studies also failed to detect significant
bryostatin 1 effects in the majority of patients (data not shown).
In one patient treated according to schedule 2, Ab-redirected
I-cell cytotoxicity versus P8 1 5 target cells rose from 42%
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

616 Phase lb Trial of Bryostatin 1
before treatment to 77% on day 2 (data not shown); values
subsequently declined to basal bevels by day 8. In 3 of 1 1 patient
samples assayed, significant increases in LAK-mediated killing
was observed, most notably on days 4 or 5. An example of such
a response in one of these patients is shown in Fig. 3.
DISCUSSION
Bryostatin 1 has shown a variety of interesting biological
properties both in vitro and in vito. At very low (i.e. , nanomo-
lar) concentrations, it stimulates the in vitro growth of normal
human hematopoietic progenitors (8), possibly through an ac-
cessory cell mechanism (9). Comparable concentrations inhibit
the growth of leukemic progenitors (7, 38), raising the possibil-
ity of a selective antileukemic effect. In animals, bryostatin 1
exhibits activity against a variety of tumor types, including
carcinoma of the breast (3), lung (4). melanoma (5). lymphoma
(6), and in a xenobiotic model, Waldenstrom’s macrogbobuline-
mia (39). It has been assumed that the antineoplastic activity of
bryostatin 1 stems from its ability to activate PKC, although
recent studies bring this concept into question (40). In human
trials to date, bryostatin 1 has been administered by several
schedules, and limited responses have been observed in patients
with malignant melanoma, renal cell carcinoma, and lymphoma
(10-13). The dose-limiting toxicity in each of these studies has
been myalgias, a phenomenon that seems to be cumulative
(10-13). This has led to a recommended Phase II dose of 25-35
p.g/m2 every 1-2 weeks for a 1-h bolus or 24-h continuous
infusion schedule, although recent studies suggest that when
administered as a 72-h infusion, considerably higher bryostatin
1 doses may be achieved (41).
In addition to a potential activity as a single agent, bryo-
statin I may also have a clinical role as a facilitator of neoplastic
cell apoptosis (23, 24). One of the known actions of PKC is to
oppose programmed cell death induced by growth factor depri-
vation (25) or cytotoxic agents (26, 27). Whereas the mecha-
nism by which PKC opposes apoptosis is uncertain, recent
attention has focused on the opposing influences of the PKC and
sphingomyebinase/ceramide pathways on cell viability (42). It
has recently been proposed that factors that modulate the PKC/
ceramide “rheostat” may shift the balance between cell survival
and death (43, 44). The apparent paradox that bryostatin 1 , a
known PKC activator, promotes apoptosis, may be explained by
its delayed ability to induce profound PKC down-regulation
(20) through a mechanism involving ubiquitination and proteo-
somal degradation of the enzyme (2 1 ). Consistent with this
concept, the effect of acute exposure of human leukemia cells to
bryostatin 1 , which leads to increased PKC activity, inhibits
ara-C-induced apoptosis, whereas a more prolonged incubation
(i.e. , 24 h) results in a decrease in enzyme activity and poten-
tiation of apoptosis (24).
In animal studies, in vivo administration of bryostatin 1 has
been shown to induce rapid reductions in cytosolic PKC activity
in peritoneal neutrophibs (45). However, this action, which
stems from transbocation of the enzyme to the cell membrane, is
distinct from the decline in total cellular activity that results
from enzyme down-modulation (22). In a recent study involving
CS7BL/6 mice receiving a single i.v. injection of bryostatin 1 (1
p.g), significant down-regulation of total splenocyte PKC activ-
70
C’,
� 60C.)
I’) 50
�400
�30
�.1 20
10
040:1 20:1 10:1 5:1
Err Ratio
Fig. 3 LAK cytotoxicity against P815 target cells from a single patienttreated according to schedule 2 as described in the text. Data shownrepresent the percentage of specific release from 51Cr-labeled P815target cells by PBMNC effectors after incubation with 500 IU/ml IL-2.Specific release is plotted against E:T ratios. Curves correspond to thenumber of days after bryostatin 1 treatment (shown in the inset). Similar
results were obtained in two additional patients.
ity was observed, which persisted for periods as long as 96 h
(31). This finding indicates that the ability of bryostatin 1 to
induce PKC down-regulation is not restricted to the in vitro
setting. The central goal of this study was to determine whether
in vivo administration of bryostatin 1 in humans by one or more
schedules could reduce PKC activity in a hematopoietic target
tissue, analogous to results obtained in a murine model (3 1).
Once identified, such a bryostatin 1 schedule could then be
combined with a cytotoxic agent (e.g.. ara-C) to mimic in vitro
regimens displaying enhanced antileukemic activity (23, 24).
Although responses in the current trial were quite heterogene-
ous, marked reductions in PBMNC PKC activity were observed
in several patients, and there was a trend (P = 0.06, signed rank
test) toward reduction at 72 h for the three cohorts as a whole.
Both the magnitude and duration of these reductions were
comparable to those encountered in an antecedent murine study
(31). It is possible that down-regulation of PBMNC PKC activ-
ity was limited by the bryostatin 1 dose used in this trial. In this
regard, Jayson et a!. (12) measured total PKC activity in
PBMNC from three patients at various intervals during the
course of a 24-h continuous infusion of 25 p.g/m2 of bryostatin
1. Although changes in PKC activity were noted in these pa-
tients, consistent depletion of activity at the end of the 24-h
infusion could not be documented. Effects on PBMNC PKC
activity at later time points were not examined in that study. It
should be noted that the dose of bryostatin 1 used in this and in
previous trials (25 p.g/m2; Refs. 10-13) has been limited by
cumulative toxicity (chiefly myalgias) that accompanies chronic
administration. It is conceivable that less frequent administra-
tion of bryostatin 1 (e.g., every 4-6 weeks) might permit higher
individual doses to be given, with greater effects on PKC
activity. Studies designed to test this hypothesis are currently
under development.
Plasma bevels of bryostatin 1 , regardless of schedule of
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Clinical Cancer Research 617
administration, were below the level of detection of the platelet
aggregation-based bioassay, which is sensitive to bryostatin 1
concentrations as low as 2-5 nsi (34). These results, as well as
those of several earlier studies in animals, suggest that the
failure to achieve high plasma bryostatin 1 concentrations stems
from rapid plasma clearance of this compound. For example,
Berkow et a!. (45), using a neutrophib activation assay, reported
that in mice receiving a single i.v. injection of bryostatin 1 (1
�i.g), plasma concentrations declined to undetectable levels (e.g.,
� 60 nM) within minutes of drug administration. More recently,
Zhang et al. (46) described the pharmacokinetics of [C26-
3H]bryostatin 1 (40 p.g/kg) in CD1IF2 mice after iv. or i.p.
administration. When given by the iv. route, btyostatin 1 was
rapidly cleared from the plasma, with concentrations falling to
- 15 nM after 4 h, to < 10 nM after 12 h, and to subnanomobar
concentrations thereafter. Both urinary and gastrointestinal ex-
cretion were detected, and bryostatin 1 was widely distributed in
tissues including lung, liver, heart, lymph nodes, and fat. If the
disposition of bryostatin 1 in humans is similar, rapid plasma
clearance secondary to excretion in conjunction with wide-
spread tissue distribution could account for our failure to detect
plasma bryostatin 1 levels of �5 ntvt in any of the patient
cohorts.
No consistent pattern of alterations in lymphocyte pheno-
type, IL-2-induced proliferation, or cytotoxicity against a van-
ety of targets was observed in the majority of subjects after
treatment with bryostatin 1 . In three patients, however, a striking
increase in IL-2-induced proliferation was noted. Our results
differ somewhat from those of previous trials in which increases
in IL-2 responsiveness and LAK activity were noted (12, 13).
However, in one of these studies, which involved a 24-h infu-
sion schedule, peak increases in IL-2-induced proliferation were
observed 2 h after initiation of drug infusion ( 13). If such
maximal responses typically occur within this time frame, they
would not have been detected at the later intervals examined in
the present study. It may be significant that two of three patients
displaying a significant increase in IL-2-induced lymphocyte
proliferation received bryostatin 1 as a split dose on days 1 and
4. Taken in conjunction with evidence that administration of
bryostatin 1 at or near the maximum tolerated dose in mice
inhibits their resistance to bacterial infection (47), these findings
raise the possibility that lower bryostatin 1 doses may be more
appropriate when this agent is intended as an immunomodulator.
In any case, based on the small number of patients in each
cohort, definitive conclusions regarding the schedule-dependent
immunomodulatory effects of bryostatin 1 cannot be drawn.
In summary, the results of the present study suggest that
administration of bryostatin 1 at a fixed dose of 25 p.g/m2
according to several schedules may be capable of down-regu-
bating PBMNC PKC activity in a subset of patients, and that in
these individuals, down-modulation is qualitatively and quanti-
tatively similar to that observed in a precbinical animal model. In
this study of limited power, response patterns seemed to favor
the 1-h bolus and 24-h infusion over the split-course schedule if
down-regulation of PKC activity is a targeted goal. However,
based on the inconstancy of the PKC modulatory effects, it is
unclear which of the tested schedules will prove optimal for
projected Phase II successor trials. It also remains possible that
higher bryostatin 1 doses could be more effective than the
25-p.g/m2 dose used in the present study. In view of evidence
that bryostatin 1 doses as high as I 20 p.g/m2 are tolerable when
administered as a 72-h continuous infusion (41 ), it is conceiv-
able that such schedules might result in more profound and/or
sustained PKC down-regulation than that observed in the pres-
ent study. Other issues remaining to be addressed include: (a)
whether bryostatin I is capable of down-regulating PKC in
neoplastic target tissues such as leukemic blasts; (b) if so.
whether this action increases the in vivo susceptibility of such
cells to cytotoxic drugs (i.e. , ara-C); and (c) whether the com-
bination of bryostatin 1 and a cytotoxic agent offers the prospect
of improved therapeutic selectivity. To answer these and related
questions, a Phase I trial of escalating doses of bryostatin 1
administered in conjunction with high-dose ara-C in patients
with refractory hematologicab malignancies is currently under
development.
REFERENCES
I. Pettit, G. R., Herald, C. L., Doubek, D. L., Herald. D. L.. Arnold, E..
and Clardy. J. Isolation and structure of bryostatin I . J. Am. Chem. Soc..104: 6846-6948, 1982.
2. Hess, A. D., Silankis, M. K., Esa, A. H., Pettit, G. R., and May, W. S.
Activation of human T lymphocytes by bryostatin 1. J. Immunol.. 141:
3263-3269. 1988.
3. Kennedy. M. J.. Prestigiacomo. L. J.. Tyler, G.. May. W. S.. andDavidson, N. E. Differential effects of bryostatin I and phorbol ester on
human breast cancer cell lines. Cancer Res., 52: 1278-1283, 1992.
4. Dale, I. L.. and Gescher, A. Effects of activators of protein kinase C.
including bryostatins 1 and 2. on the growth of A549 human lung
carcinoma cells. Int. J. Cancer, 43: 158-163, 1989.
5. Schuchter, L. M., Esa, A. H., May. S.. Laulis. M. K.. Pettit. G. R..
and Hess, A. D. Successful treatment of murine melanoma with bryo-
statin I . Cancer Res., 51: 682-687, 1991.
6. Hornung, R. L., Pearson, J. W., Beckwith, M., and Longo, D. L.Preclinical evaluation of bryostatin as an anticancer agent against sev-
eral murine tumor cell lines: in vitro versus in vito activity. Cancer Res..52: 101-107, 1992.
7. Jones. R. J., Sharkis, S. J., Miller, C. B., Rowinsky, E. K.. Burke.
P. J., and May, W. S. Bryostatin I, a unique biologic response modifier:
antileukemic activity in vitro. Blood, 75: 1319-1323, 1990.
8. May. W. S.. Sharkis, S. S., Esa, A. H., Gebbia. V., Kraft, A. S., Pettit.
G. R.. and Sensenbrenner, L. L. Antineoplastic bryostatins are multipo-tent stimulators of human hematopoietic progenitor cells. Proc. NatI.
Acad. Sci. USA, 84: 8483-8487, 1987.
9. Sharkis, S. J., Jones, R. J., Bellis, M. L., Demetri, G. D., Griffin.J. D., Civin, C., and May, W. S. The action of bryostatin I on normalhuman hematopoietic progenitors is mediated by accessory cell releaseof growth factors. Blood, 76: 716-721, 1990.
10. Philip. P. A., Rea, D., Thavasu, P., Carmichael, J., Stuart, N. S.,
Rockett, H., Talbot, D. C., Ganesan, T., Pettit, G. R., Balkwill, F., and
Harris, A. L. Phase I study of bryostatin 1 : assessment of interleukin-6
and tumor necrosis factor a in vito. J. Natl. Cancer Inst., 85: 1812-
1818, 1993.
I 1. Prendiville, J., Crowther, D., Thatcher, N., Woll, P. J., Fox, B. W.,
McGown, A., Testa, N., Stem, P., McDermott, R., Potter, M., and Pettit,
G. R. A Phase I study of intravenous bryostatin I in patients with
advanced cancer. Br. J. Cancer, 68: 418-424, 1993.
12. Jayson, G. C.. Crowther, D., Prendiville, J., McGown, A. T.,
Scheid. C., Stem, P., Young, R., Brenchley, P.. Chang. J., Owens, S.,
and Pettit, G. R. A Phase I trial of bryostatin 1 in patients with advanced
malignancy using a 24-hour intravenous infusion. Br. J. Cancer, 72:
461-468, 1995.
13. Scheid, C., Prendiville, J., Jayson, G., Crowther, D., Fox, B., Pettit,
G. R.. and Stem, P. L. Immunomodulation in patients receiving bryo-statin 1 in a Phase I clinical study: comparison with effects of bryostatin
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

618 Phase lb Trial of Bryostatin 1
I on lymphocyte function in vitro. Cancer Immunol. Immunother., 39:
223-230, 1994.
14. Berkow, R. L., and Kraft, A. S. Bryostatin, an activator of the
calcium phospholipid-dependent protein kinase. blocks phorbol ester-
induced differentiation of human promyelocytic leukemia cells HL-60.
Biochem. Biophys. Res. Commun., 13!: 1109-1116, 1985.
15. Kraft, A. S., Smith, J. B., and Berkow, R. L. Bryostatin, an activator
of the calcium phospholipid-dependent protein kinase. blocks phorbol
ester-induced differentiation of human promyelocytic leukemia cells
HL-60. Proc. NatI. Acad. Sci. USA, 83: 1334-1338, 1986.
16. Hennings. H., Blumberg. P. M., Pettit, G. R., Herald, C. L., Shores,R. A.. and Yuspa, S. H. Bryostatin 1, an activator of protein kinase C,
inhibits tumor promotion by phorbol esters in SENCAR mouse skin.
Carcinogenesis (Lond.), 8: 1343-1346, 1987.
I 7. Ng. S. B.. and Guy, G. R. Two protein kinase activators, bryostatin
1 and phorbol-1 2-myristate- 13-acetate, have different effects on haemo-poietic cell proliferation and differentiation. Cell Signalling, 4: 405-
416, 1992.
18. Szallasi, Z., Smith, C. B., Pettit, G. R., and Blumberg, P. M.Differential regulation of protein kinase C isozymes by bryostatin I and
phorbol 12-myristate 13-acetate in NIH 3T3 fibroblasts. J. Biol. Chem.,
269: 2118-2124. 1994.
19. Hocevar, B. A., and Fields, A. P. Selective translocation of �protein kinase C to the nucleus of human promyelocytic (HL-60) leu-
kemia cells. J. Biol. Chem., 266: 28-33, 1991.
20. lsakov, N., Galron, D., Mustelin, T., Pettit, G. R., and Altman, A.Inhibition of phorbol ester-induced T cell proliferation by bryostatin I is
associated with rapid degradation of protein kinase C. J. Immunol., 150:
1195-1204, 1993.
21. Lee, H-W., Smith, L., Pettit, G. R., Vinitsky, A., and Smith, J. B.Ubiquitination of protein kinase C-a and degradation by the proteo-
some. J. Biol. Chem.. 271: 20973-20976, 1996.
22. Rodriguez-Pena, R., and Rozengurt, E. Disappearance of Ca2�-sensitive, phospholipid-dependent kinase activity in phorbol ester-
treated 3T3 cells. Biochem. Biophys. Res. Commun., 120: 1053-1059,
1984.
23. Grant, S., Jarvis, D., Swerdlow, P., Turner, A., Traylor, R., Wallace,H., Lin, P-S., Pettit, G. R., and Gewirtz, D. A. Potentiation of theactivity of 1-�3-o-arabinofuranosylcytosine by the macrocyclic lactonePKC activator bryostatin I in HL-60 cells: association with enhanced
fragmentation of mature DNA. Cancer Res., 52: 6270-6278, 1992.
24. Jarvis, W. D., Gewirtz, D. A., Povirk, L., Turner, A., Traylor, R.,Pettit, G. R., and Grant, S. Effects of bryostatin I and other pharmaco-
logical activators of protein kinase C on l-�3-D-arabinofuranosylcy-
tosine-induced apoptosis in HL-60 human promyelocytic leukemia
cells. Biochem. Pharmacol., 47: 839-852, 1994.
25. Lotem, J., Cragoe, E. J., and Sachs, L. Rescue from programmedcell death in leukemia and normal myeloid cells. Blood, 78: 953-960,1991.
26. McConkey, D. J.. Hartzell, P., Jondal, M., and Orrenius, S. Inhibi-tion of DNA fragmentation in thymocytes and isolated thymocyte nuclei
by agents that stimulate protein kinase C. J. Biol. Chem., 264: 13399-13402, 1989.
27. Bertrand, R., Solary, E.. O’Connor, P.. Kohn. K. W.. and Pommier,Y. Induction of a common pathway of apoptosis by staurosporine. Exp.
Cell Res., 211: 314-321, 1994.
28. Grant, S., Turner, A., Bartimole, T. M., Nelms, P. A., Joe, V. C.,
and Jarvis, W. D. Modulation of 143-D-arabinofuranosylcytosine-in-
duced apoptosis in human myeloid leukemia cells by staurosporine andother pharmacological inhibitors of protein kinase C. Oncol. Res.. 6:87-99, 1994.
29. Kauffman, S. H. Induction of endonucleolytic DNA cleavage by in
human acute myelogenous leukemia cells by etoposide, camptothecin.and other cytotoxic anticancer agents: a cautionary note. Cancer Res..49: 5870-5878, 1989.
30. Gorczyca, W., Bigman. K., Mittelman, A., Ahmed, T., Gong, J.,
Melamed, M. R., and Darzynkiewicz, Z. Induction of DNA strand
breaks associated with apoptosis during treatment of leukemias. Leuke-
mia (Baltimore), 7: 659-670, 1993.
31. Bear, H. D., McFadden, A., Turner, A. J., and Grant, S. Bryostatin
1 induces long-term depletion of protein kinase C in t’ivo following a
single intravenous injection. Anticancer Res., 6: 384-391, 1995.
32. Boyum, A. Isolation of mononuclear cells and granulocytes fromhuman blood. Scand. J. Clin. Lab. Invest., 21: 77-84, 1968.
33. Grant, S., Turner, A. J., Freemerman, A. J., Wang, Z.. Kramer, L.,
and Jarvis, W. D. Modulation of protein kinase C activity and calcium-
sensitive isoform expression in human myeboid leukemia cells by bryo-statin I : relationship to differentiation and ara-C-induced apoptosis.
Exp. Cell Res., 228: 65-75, 1996.
34. Carr, M. E., Jr., Can, S. L., and Grant, S. A sensitive platelet
activation-based functional assay for the antileukemic agent bryostatin
1. Anticancer Drugs, 6: 384-391, 1995.
35. Bear, H. D., Susskind, B. M., Close, K. A., and Barrett, S. K.Phenotype of tumor-specific cytotoxic T-lymphocytes and requirementsfor their in vitro generation from tumor-bearing host and immune
spleens. Cancer Res., 48: 1422-1427, 1988.
36. Hengel, H., Wagner, H., and Heeg, K. Triggering of CD8� cyto-toxic T lymphocytes via CD3-#{128}differs from triggering via a/�3 T cellreceptor: CD3-#{128}-induced cytotoxicity occurs in the absence of proteinkinase C and does not result in exocytosis of serine esterases. J. Immu-
nol.. 147: 1115-1120, 1991.
37. Conrad, D. H. Fcc RII/CD23. The low affinity receptor for for IgE.Annu. Rev. Immunol., 8: 623-645, 1990.
38. Grant, S., Traybor, R., Bhalla, K., McCrady, C., and Pettit, G. R.
Effect of a combined exposure to l-�3-D-arabinofuranosylcytosine, bryo-statin 1 , and rGM-CSF on the in vitro clonogenic growth of normal and
leukemic human hematopoietic progenitor cells. Leukemia (Baltimore),5: 432-439, 1993.
39. Mohammad, R. M., Al-Katib, A., Pettit, G. R., and Sensenbrenner,
L. L. Successful treatment of human Waldenstrom’s macroglobubinemia
with combination biological and chemotherapy agents. Cancer Res., 54:
165-168, 1994.
40. Szallasi, Z., Du, L., Levine, R., Lewin, N. E., Nguyen, P. N..
Williams, M. D.. Pettit, G. R., and Blumberg, P. M. The bryostatins
inhibit growth of B16IF1O melanoma cells in vitro though a proteinkinase C-independent mechanism: dissociation of activities using 26-
epi-bryostatin 1. Cancer Res., 56: 2105-21 11, 1996.
41. Varterasian, M., Eilender, D., Mohammad, R., Chen, B., Hulburd,
K., Rodriguez, D., Pluda, J., Valdivieso, M., and Al-Katib, A. Phase Itrial of bryostatin 1 in relapsed lymphoma and CLL. Proc. Am. Soc.
Clin. Oncol.. 15: 481, 1996.
42. Hannun, Y. A. Functions of ceramide in coordinating cellularresponses to stress. Science (Washington DC), 274: 1855-1859, 1996.
43. Cuvillier, 0., Pirianov, G.. Kleuser, B., Vanek, P. 0., Coso, 0. A.,
Gutkind, J. S., and Spiegel, S. Suppression of ceramide-mediated pro-grammed cell death by sphingosine-l-phosphate. Nature (Lond.), 381:
800-803, 1996.
44. Grant. S., and Jarvis, W. D. Modulation of drug-induced apoptosisby interruption of the protein kinase C signal transduction pathway: anew therapeutic strategy. Clin. Cancer Res., 2: 1915-1920, 1996.
45. Berkow, R. L., Schlabach. L., Dodson, R., Benjamin, W. H., Jr.,
Pettit, G. R., Rustagi, P., and Kraft, A. S. In vivo administration of
the anticancer agent bryostatin 1 activates platelets and neutrophilsand modulates protein kinase C activity. Cancer Res., 53: 2810-2815,
1993.
46. Zhang, X., Zhang, R., Zhao, H., Cai, H., Gush, K. A., Kerr, R. G.,Pettit, G. R., and Kraft, A. S. Preclinical pharmacology of the naturalproduct anticancer agent bryostatin I , an activator of protein kinase C.Cancer Res., 56: 802-808, 1996.
47. Kraft, A. S., Adler, V., Hall, P., Pettit, G. R., Benjamin, W. H., andBriles, D. E. In vivo administration of bryostatin 1, a protein kinase C
activator, decreases munine resistance to Salmonella tvphimurium. Can-cer Res., 52: 2143-2147, 1992.
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1998;4:611-618. Clin Cancer Res S Grant, J Roberts, E Poplin, et al. malignancies.Phase Ib trial of bryostatin 1 in patients with refractory
Updated version
http://clincancerres.aacrjournals.org/content/4/3/611
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/4/3/611To request permission to re-use all or part of this article, use this link
Research. on December 10, 2020. © 1998 American Association for Cancerclincancerres.aacrjournals.org Downloaded from