Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Oct. 1983, p. 99-1050019-9567/83/100099-07$02.00/0Copyright X) 1983, American Society for Microbiology
Vol. 42, No. 1
Phagocytic Killing of Neisseria gonorrhoeae by HumanMonocytes
JOSEPH R. MEZZATESTA AND RICHARD F. RESTt*Department of Molecular and Medical Microbiology, University ofArizona Health Sciences Center, Tucson,
Arizona 85724
Received 11 March 1983/Accepted 28 June 1983
The ability of human monocytes to phagocytize and kill nonpiliated opaque (T3)and transparent (T4) gonococci was investigated in a tumbling tube suspensionassay. A serum-sensitive strain, F62, and a serum-resistant strain, FA19, werestudied. CFU remaining after incubation with monocytes were used to assess theextent of killing. The data show that 50% of T3 and T4 gonococci of both strainswere killed by monocytes over a 2-h period. Serum was necessary for the killing oftransparent gonococci of both strains as well as for FA19 T3. Concentrations ofserum ranging from 0.5 to 10% were equally effective, and heat-labile componentswere required. Killing of F62 T3, however, occurred in the absence of serum. Anincreased ratio of bacteria -to monocytes decreased the rate of killing. A 30-minpreopsonization of gonococci in 10% serum resulted in an enhanced rate of killing.Monocytes were able to kill plate-grown, but not log-phase, organisms. Disrup-tion of the monocytes by sonication to release internalized bacteria did notincrease the number of viable organisms. The addition of 10 ,ug of cytochalasin Bper ml completely inhibited the reduction in colony numbers over time. Thesedata indicate that freshly isolated human monocytes are capable of phagocytizingand killing nonpiliated gonococci.
Urogenital infections with Neisseria gonor-rhoeae result in an acute inflammatory responseproducing a purulent urethral or cervical dis-charge. These exudates contain predominantlypolymorphonuclear leukocytes (PMN) alongwith fewer numbers of epithelial cells and mono-nuclear phagocytes (MNP) (31). Consequently,the interaction of gonococci with human MNPhas not been extensively studied despite thepresence of these cells in inflammatory exudatesand the more widespread contact with bloodmonocytes and tissue macrophages in the dis-seminated form of the disease.Drutz (8) reported that human MNP kill 100%
of ingested piliated gonococci, as measured by[3H]adenine incorporation, and that monocytesare less phagocytic than macrophages. Othershave reported, however, that a small percentageof intracellular piliated organisms survive andgrow within the human MNP, possibly servingas the source of chronic infection and dissemina-tion (29, 31).Other available studies describe the interac-
tion of gonococci with mouse macrophages.Blake and Swanson (2) showed with light andelectron microscopy that nonpiliated gonococciassociate with peritoneal macrophages to agreater degree than do piliated forms. Using
t Present address: Department of Microbiology and Immu-nology, Hahnemann University, Philadelphia, PA 19102.
radiolabeling techniques, Jones and Buchanan(11) also found mouse macrophages better ableto phagocytize nonpiliated organisms. Phagocy-tosis of piliated gonococci could be enhanced byopsonization with specific antibody to pili (12).However, Ota et al. (18) reported on the phago-cytosis of nonopsonized, piliated gonococci and,with the aid of electron micrographs, speculatedthat some of these bacteria were able to surviveintracellularly. Recently, Cooper and Floyd (4)found murine macrophages to be capable ofkilling all intracellular piliated gonococci. Inaddition, the morphological appearance of inter-nalized gonococci did not reflect their viability.
In this report, we use agar plating techniquesto quantitate the percentage of viable gonococciafter incubation with human monocytes in asuspension assay. The data from these studiesdemonstrate that human peripheral blood mono-cytes are capable of killing nonpiliated gono-cocci. The transparent organisms of strainscausing local (UGI) and disseminated (DGI)forms of infection were found to be equallysusceptible to phagocytic killing in the presenceof serum, whereas only opaque UGI gonococciwere killed by monocytes in the absence ofserum.
MATERIALS AND METHODSMedia. Hank's balanced salt solution (HBSS)
(GIBCO Laboratories, Grand Island, N.Y.) containing
99
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

100 MEZZATESTA AND REST
0.35 mg of NaHCO3 per ml and 10 mM N-2-hydroxy-ethylpiperazine-N'-2-ethanesulfonic acid (pH7.1) wasused for preparation of mononuclear cells. Final sus-pensions of monocytes and gonococci were made inHBSS containing 0.1% gelatin (HBSS-gel).
Bacteria. Opaque (T3) and transparent (T4) F62(UGI) and FA19 (DGI) nonpiliated gonococci weremaintained on GC medium base (Difco Laboratories,Detroit, Mich.) agar plates with added supplements(described below) in a humidified incubator at 37°C in5% CO2 in air. Colonies were selectively passageddaily by using the criteria of Kellogg et al. (13, 14) andSwanson (26) for piliation and opacity.Broth cultures of gonococci. Log- and stationary-
phase bacteria were grown in 20 ml of GC brothenriched with 200 p.l of 4% NaHCO3, 200 p.l of asupplement containing 20 p.g of cocarboxylase per ml,400 mg of glucose per ml, and 20 mg of glutamine perml and 200 p.l of Fe(NO3)3 (0.1 mg/ml). Plate-growngonococci were suspended in 5 ml of this broth (opticaldensity at 550 nm, 0.3; Bausch & Lomb Spectronic20), transferred to a Klett flask containing 15 ml ofenriched GC broth, and placed in a 37°C shaking waterbath. For FA19 T3 and T4, the midpoint of log-phasegrowth was reached at 60 Klett units after 3 h ofgrowth. For F62 T3 and T4, the midpoint was about 90Klett units, also after 3 h of incubation. The stationaryphase was indicated by Klett values of 100 for FA19and 130 for F62 after 5 h of incubation.
Preparation of gonococci for phagocytic killing assay.Plate-grown FA19 T3 and T4 between 18 and 21 h oldwere suspended in 5 ml of GC broth to an opticaldensity of 0.35. Plate-grown F62 T3 and T4 also 18- to21-h-old, and log- and stationary-phase broth culturesof both strains and types were suspended to 0.3 in 5 mlof GC broth. These suspensions contained 1 x 108 to 2x 108 CFU/ml. Dilutions in HBSS-gel were made toprovide final suspensions of 5 x 105 to 5 x 107 CFU/mlas required when added to the phagocytic mixture.
Preparation of monocyte cell suspension. Three partsof heparinized venous blood was mixed with 1 partPlasmagel (Roger Bellon Laboratories, Neuilly,France; distributed by HTI Corp., Buffalo, N.Y.), andsedimented in a siliconized separatory funnel for 1 h atroom temperature. The erythrocytes were discarded,and the leukocyte-rich supernatant was centrifuged for10 min at 200 x g followed by two washes in HBSS.The leukocytes were suspended in HBSS, and 5 to 10ml of the suspension was layered onto 10 ml ofIsolymph (Gallard-Schlesinger Chemical Manufac-turing Corp., Carle Place, N.Y.) and centrifuged atroom temperature for 30 min at 400 x g. The interfacecontaining the mononuclear cells was recovered,washed twice, and suspended in HBSS. The cells werecounted in 0.25% trypan blue in HBSS (.98% viable),and slides were prepared in a cytocentrifuge (Cyto-spin; Shandon Southern International, Sewickley, Pa.)for Wright stain and differential counting. The inter-face contained an average of 2.0%o PMN, 57% lympho-cytes, 37% monocytes, and 4% basophils. The mono-
nuclear cells were suspended at 12.5 x 10' monocytesper ml in HBSS-gel.
Phagocytic killing assay. The tumbling tube assaywas performed in Falcon (Becton, Dickinson and Co.,Cockeysville, Md.) 12- by 75-mm sterile plastic culturetubes rotated at 4 rpm in a 37°C dry 5% C02-in-airincubator. Each tube contained 400 p.l of the mononu-
clear cell suspension (5 x 106 monocytes), 500 ,u1 ofbacteria, and 100 K. or less of pooled normal humanserum (PNHS). The final volume was always 1.0 ml.Samples of 10 ,u1 were taken at 0, 30, 60, and 120 minand diluted 1:1,000 in GC broth. Samples of 40 ,u1 werespread onto duplicate GC agar plates and incubatedovernight. Results are expressed as the percentage ofviable organisms at each time point relative to that atzero time. Where indicated, samples were diluted inHBSS by using plastic tubes and sonicated for 30 s at asetting of 4 (Heat Systems-Ultrasonics, Inc., Plain-view, N.Y.; model W-225R). In some experiments, themononuclear cells were preincubated with 10 pg ofcytochalasin B (Sigma Chemical Co., St. Louis, Mo.)per ml for 10 min at 37°C before the addition ofbacteria. The data are presented as the means ofbetween two and seven experiments as shown withinparentheses in the figure legends.
RESULTS
Effect of bacteria/monocyte ratios on phagocyt-ic killing. Human monocytes killed approximate-ly 50 to 60% of nonpiliated, plate-grown FA19 T4gonococci over 2 h in the presence of 10%PNHS. The ratio of bacteria to monocytes wasfound to influence the rate of phagocytic killing(Fig. 1). Suspensions of gonococci were adjust-ed to give ratios of 10:1, 1:1, and 0.5:1 whenmixed with 5 x 106 monocytes in HBSS-gel-10% PNHS. The rate of killing was similar forratios of 0.5:1 and 1:1; colony numbers de-creased progressively over time with maximalreduction occurring by 120 min. At a 10:1 ratio,the reduction in colony numbers occurred morerapidly with a decrease to approximately 50%within 30 min. However, no further killing oc-curred beyond this time. Rather, the number ofgonococci increased between 60 and 120 min,returning to the number present at the start ofthe assay.
Increasing the number of monocytes twofoldto 10 x 106/ml while maintaining the bacterialsuspension at 5 x 106/ml resulted in increasedkilling at 30 min and about a 50% reduction by 60min (Fig. 1). As with the 1:1 ratio, colonynumbers did not increase appreciably between60 and 120 min. In contrast, no phagocytickilling was seen when monocyte numbers werereduced to 5 x 105/ml (data not shown).Monocytes incubated for 2 h with either whole
gonococci at a 1:1 ratio or the supernatant of a 2h culture of 5 x 106 CFU/ml were >95% viable,and at least 80% of the initial number of mono-nuclear cells were recovered. Thus, the patternof phagocytic killing is not related to a concomi-tant loss of viable monocytes over time.Serum requirements for phagocytic killing. The
experiments described above were performed in10% PNHS. In other experiments, plate-grownFA19 T4 gonococci were incubated at a ratio1:1 ratio with 5 x 106 monocytes per ml in vari-
INFECT. IMMUN.
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
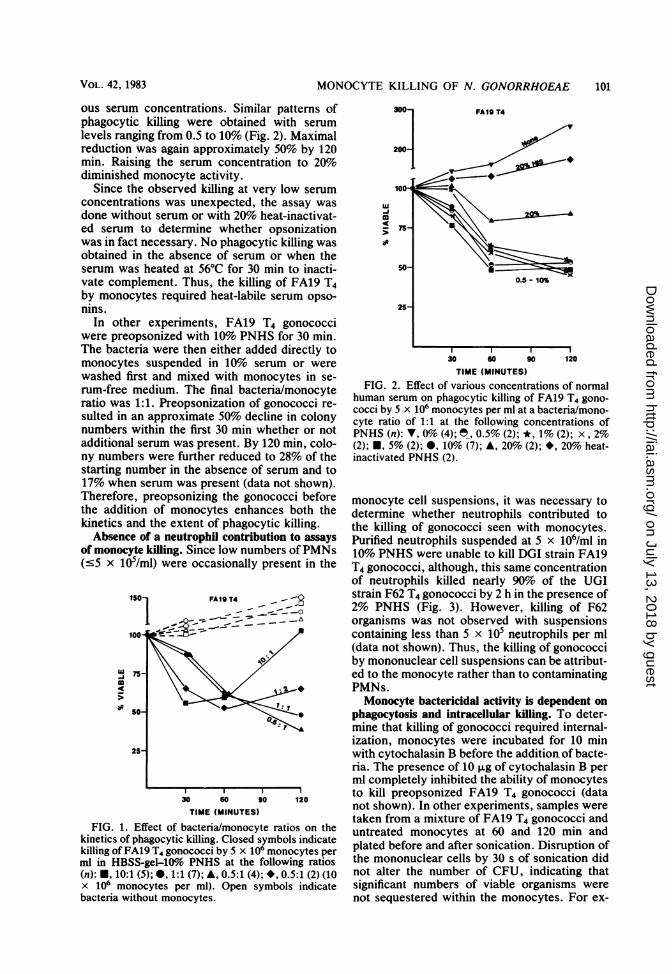
MONOCYTE KILLING OF N. GONORRHOEAE
ous serum concentrations. Similar patterns ofphagocytic killing were obtained with serumlevels ranging from 0.5 to 10% (Fig. 2). Maximalreduction was again approximately 50% by 120min. Raising the serum concentration to 20%diminished monocyte activity.
Since the observed killing at very low serumconcentrations was unexpected, the assay wasdone without serum or with 20% heat-inactivat-ed serum to determine whether opsonizationwas in fact necessary. No phagocytic killing wasobtained in the absence of serum or when theserum was heated at 56°C for 30 min to inacti-vate complement. Thus, the killing of FA19 T4by monocytes required heat-labile serum opso-nins.
In other experiments, FA19 T4 gonococciwere preopsonized with 10% PNHS for 30 min.The bacteria were then either added directly tomonocytes suspended in 10%o serum or werewashed first and mixed with monocytes in se-rum-free medium. The final bacteria/monocyteratio was 1:1. Preopsonization of gonococci re-sulted in an approximate 50% decline in colonynumbers within the first 30 min whether or notadditional serum was present. By 120 min, colo-ny numbers were further reduced to 28% of thestarting number in the absence of serum and to17% when serum was present (data not shown).Therefore, preopsonizing the gonococci beforethe addition of monocytes enhances both thekinetics and the extent of phagocytic killing.Absence of a neutrophil contribution to assays
of monocyte killing. Since low numbers ofPMNs(c5 x 105/ml) were occasionally present in the
1501SO _A -
25-
10
30 s0 90 120
TIME (MINUTES)
FIG. 1. Effect of bacteria/monocyte ratios on thekinetics of phagocytic killing. Closed symbols indicatekilling of FA19 T4 gonococci by 5 x 106 monocytes perml in HBSS-gel-109o PNHS at the following ratios(n): *, 10:1 (5); , 1:1 (7); A, 0.5:1 (4); *, 0.5:1 (2) (10x 10' monocytes per ml). Open symbols indicatebacteria without monocytes.
3001
2004
-iCa
FA19 T4
30 60 90 120TIME (MINUTES)
FIG. 2. Effect of various concentrations of normalhuman serum on phagocytic killing of FA19 T4 gono-cocci by 5 x 106 monocytes per ml at a bacteria/mono-cyte ratio of 1:1 at the following concentrations ofPNHS (n): V, 0% (4); , 0.5% (2); *, 1% (2); x, 2%(2); E, 5% (2); *, 10% (7); A, 20% (2); *, 20% heat-inactivated PNHS (2).
monocyte cell suspensions, it was necessary todetermine whether neutrophils contributed tothe killing of gonococci seen with monocytes.Purified neutrophils suspended at 5 x 106/ml in10% PNHS were unable to kill DGI strain FA19T4 gonococci, although, this same concentrationof neutrophils killed nearly 90% of the UGIstrain F62 T4 gonococci by 2 h in the presence of2% PNHS (Fig. 3). However, killing of F62organisms was not observed with suspensionscontaining less than 5 x 105 neutrophils per ml(data not shown). Thus, the killing of gonococciby mononuclear cell suspensions can be attribut-ed to the monocyte rather than to contaminatingPMNs.Monocyte bactericidal activity is dependent on
phagocytosis and intracellular killing. To deter-mine that killing of gonococci required internal-ization, monocytes were incubated for 10 minwith cytochalasin B before the addition of bacte-ria. The presence of 10 ,ug of cytochalasin B perml completely inhibited the ability of monocytesto kill preopsonized FA19 T4 gonococci (datanot shown). In other experiments, samples weretaken from a mixture of FA19 T4 gonococci anduntreated monocytes at 60 and 120 min andplated before and after sonication. Disruption ofthe mononuclear cells by 30 s of sonication didnot alter the number of CFU, indicating thatsignificant numbers of viable organisms werenot sequestered within the monocytes. For ex-
VOL. 42, 1983 101
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
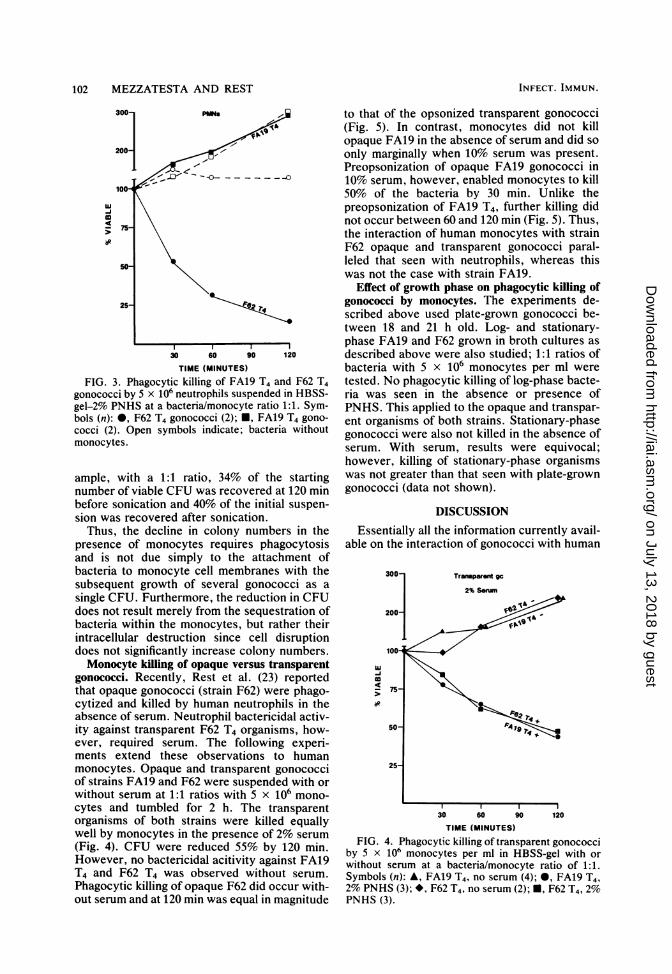
102 MEZZATESTA AND REST
-75
25
30 60 90 120
TIME (MINUTES)
FIG. 3. Phagocytic killing of FA19 T4 and F62 T4gonococci by 5 x 106 neutrophils suspended in HBSS-gel-2% PNHS at a bacteria/monocyte ratio 1:1. Sym-bols (n): 0, F62 T4 gonococci (2); *, FA19 T4 gono-cocci (2). Open symbols indicate; bacteria withoutmonocytes.
ample, with a 1:1 ratio, 34% of the startingnumber of viable CFU was recovered at 120 minbefore sonication and 40% of the initial suspen-sion was recovered after sonication.Thus, the decline in colony numbers in the
presence of monocytes requires phagocytosisand is not due simply to the attachment ofbacteria to monocyte cell membranes with thesubsequent growth of several gonococci as a
single CFU. Furthermore, the reduction in CFUdoes not result merely from the sequestration ofbacteria within the monocytes, but rather theirintracellular destruction since cell disruptiondoes not significantly increase colony numbers.Monocyte killing of opaque versus transparent
gonococci. Recently, Rest et al. (23) reportedthat opaque gonococci (strain F62) were phago-cytized and killed by human neutrophils in theabsence of serum. Neutrophil bactericidal activ-ity against transparent F62 T4 organisms, how-ever, required serum. The following experi-ments extend these observations to humanmonocytes. Opaque and transparent gonococciof strains FA19 and F62 were suspended with or
without serum at 1:1 ratios with 5 x 106 mono-
cytes and tumbled for 2 h. The transparentorganisms of both strains were killed equallywell by monocytes in the presence of 2% serum
(Fig. 4). CFU were reduced 55% by 120 min.However, no bactericidal acitivity against FA19T4 and F62 T4 was observed without serum.
Phagocytic killing of opaque F62 did occur with-out serum and at 120 min was equal in magnitude
to that of the opsonized transparent gonococci(Fig. 5). In contrast, monocytes did not killopaque FA19 in the absence of serum and did soonly marginally when 10% serum was present.Preopsonization of opaque FA19 gonococci in10% serum, however, enabled monocytes to kill50% of the bacteria by 30 min. Unlike thepreopsonization of FA19 T4, further killing didnot occur between 60 and 120 min (Fig. 5). Thus,the interaction of human monocytes with strainF62 opaque and transparent gonococci paral-leled that seen with neutrophils, whereas thiswas not the case with strain FA19.
Effect of growth phase on phagocytic killing ofgonococci by monocytes. The experiments de-scribed above used plate-grown gonococci be-tween 18 and 21 h old. Log- and stationary-phase FA19 and F62 grown in broth cultures asdescribed above were also studied; 1:1 ratios ofbacteria with 5 x 106 monocytes per ml weretested. No phagocytic killing of log-phase bacte-ria was seen in the absence or presence ofPNHS. This applied to the opaque and transpar-ent organisms of both strains. Stationary-phasegonococci were also not killed in the absence ofserum. With serum, results were equivocal;however, killing of stationary-phase organismswas not greater than that seen with plate-growngonococci (data not shown).
DISCUSSIONEssentially all the information currently avail-
able on the interaction of gonococci with human
300- Transparent gc
2%Swm
200- _
100tlu\
75-
25- l
30 60 90 120
TIME (MINUTES)FIG. 4. Phagocytic killing of transparent gonococci
by 5 x 106 monocytes per ml in HBSS-gel with orwithout serum at a bacteria/monocyte ratio of 1:1.Symbols (n): A, FA19 T4, no serum (4); *, FA19 T4,2% PNHS (3); *, F62 T4, no serum (2); U, F62 T4, 2%PNHS (3).
INFECT. IMMUN.
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
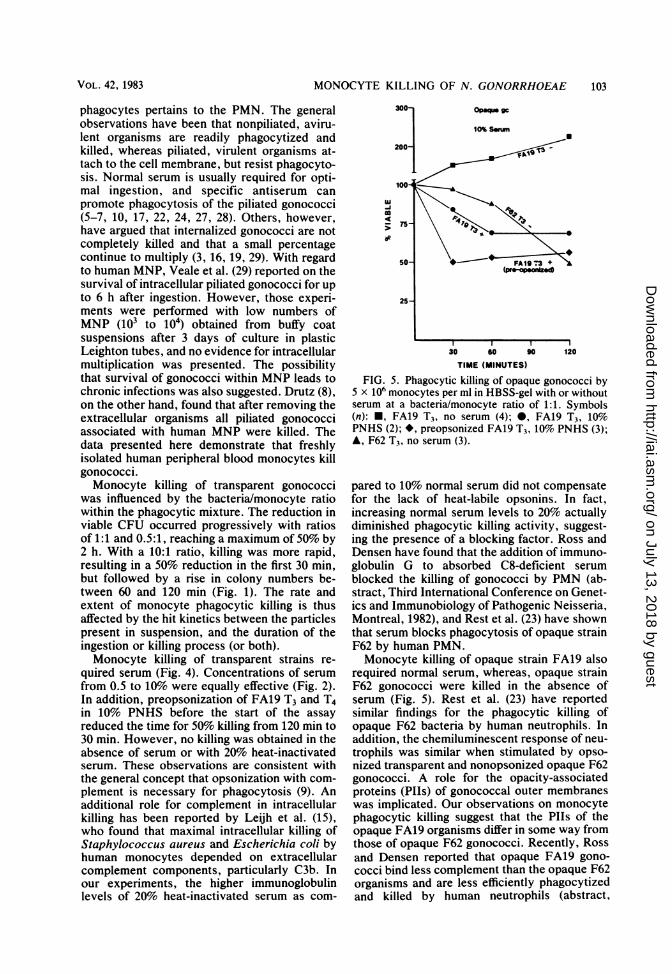
MONOCYTE KILLING OF N. GONORRHOEAE
phagocytes pertains to the PMN. The generalobservations have been that nonpiliated, aviru-lent organisms are readily phagocytized andkilled, whereas piliated, virulent organisms at-tach to the cell membrane, but resist phagocyto-sis. Normal serum is usually required for opti-mal ingestion, and specific antiserum canpromote phagocytosis of the piliated gonococci(5-7, 10, 17, 22, 24, 27, 28). Others, however,have argued that internalized gonococci are notcompletely killed and that a small percentagecontinue to multiply (3, 16, 19, 29). With regardto human MNP, Veale et al. (29) reported on thesurvival of intracellular piliated gonococci for upto 6 h after ingestion. However, those experi-ments were performed with low numbers ofMNP (103 to 104) obtained from buffy coatsuspensions after 3 days of culture in plasticLeighton tubes, and no evidence for intracellularmultiplication was presented. The possibilitythat survival of gonococci within MNP leads tochronic infections was also suggested. Drutz (8),on the other hand, found that after removing theextracellular organisms all piliated gonococciassociated with human MNP were killed. Thedata presented here demonstrate that freshlyisolated human peripheral blood monocytes killgonococci.Monocyte killing of transparent gonococci
was influenced by the bacteria/monocyte ratiowithin the phagocytic mixture. The reduction inviable CFU occurred progressively with ratiosof 1:1 and 0.5:1, reaching a maximum of 50% by2 h. With a 10:1 ratio, killing was more rapid,resulting in a 50% reduction in the first 30 min,but followed by a rise in colony numbers be-tween 60 and 120 min (Fig. 1). The rate andextent of monocyte phagocytic killing is thusaffected by the hit kinetics between the particlespresent in suspension, and the duration of theingestion or killing process (or both).Monocyte killing of transparent strains re-
quired serum (Fig. 4). Concentrations of serumfrom 0.5 to 10% were equally effective (Fig. 2).In addition, preopsonization of FA19 T3 and T4in 10% PNHS before the start of the assayreduced the time for 50% killing from 120 min to30 min. However, no killing was obtained in theabsence of serum or with 20% heat-inactivatedserum. These observations are consistent withthe general concept that opsonization with com-plement is necessary for phagocytosis (9). Anadditional role for complement in intracellularkilling has been reported by Leijh et al. (15),who found that maximal intracellular killing ofStaphylococcus aureus and Escherichia coli byhuman monocytes depended on extracellularcomplement components, particularly C3b. Inour experiments, the higher immunoglobulinlevels of 20% heat-inactivated serum as com-
-75 47
50- FA1T3 +
25-
30 G0 90 120
TIME (MINUTES)
FIG. 5. Phagocytic killing of opaque gonococci by5 x 106 monocytes per ml in HBSS-gel with or withoutserum at a bacteria/monocyte ratio of 1:1. Symbols(n): *, FA19 T3, no serum (4); 0, FA19 T3, 10%PNHS (2); *, preopsonized FA19 T3, 10% PNHS (3);A, F62 T3, no serum (3).
pared to 10% normal serum did not compensatefor the lack of heat-labile opsonins. In fact,increasing normal serum levels to 20% actuallydiminished phagocytic killing activity, suggest-ing the presence of a blocking factor. Ross andDensen have found that the addition of immuno-globulin G to absorbed C8-deficient serumblocked the killing of gonococci by PMN (ab-stract, Third International Conference on Genet-ics and Immunobiology of Pathogenic Neisseria,Montreal, 1982), and Rest et al. (23) have shownthat serum blocks phagocytosis of opaque strainF62 by human PMN.Monocyte killing of opaque strain FA19 also
required normal serum, whereas, opaque strainF62 gonococci were killed in the absence ofserum (Fig. 5). Rest et al. (23) have reportedsimilar findings for the phagocytic killing ofopaque F62 bacteria by human neutrophils. Inaddition, the chemiluminescent response of neu-trophils was similar when stimulated by opso-nized transparent and nonopsonized opaque F62gonococci. A role for the opacity-associatedproteins (Plls) of gonococcal outer membraneswas implicated. Our observations on monocytephagocytic killing suggest that the PIIs of theopaque FA19 organisms differ in some way fromthose of opaque F62 gonococci. Recently, Rossand Densen reported that opaque FA19 gono-cocci bind less complement than the opaque F62organisms and are less efficiently phagocytizedand killed by human neutrophils (abstract,
VOL. 42, 1983 103
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

TABLE 1. Summary of observations on N. gonorrhoeaeSerum bactericidal Monocyte phagocytic killing Neutrophil phagocytic
activity'aetohlpaoyiStrain Colony type Plate Log phase Plate grown Log phase killing (plate grownStan Colony type Plate Log phase with serum)
grown (both) With Without (broth) withserum serum serum
F62 Opaque (T3) S R NTb + _ NTF62 Transparent (T4) R R + - - +FA19 Opaque (T3) R R + - - NTFA10 Transparent (T4) R R + -
a The serum bactericidal assay used 5 x 106 CFU/ml in HBSS-gel at 37°C for 120 min with 2% PNHS (strainF62) or 10% PNHS (strain FA19). S, No survival; R, 100% survival.
b NT, Not tested.
American Federation of Clinical Research, p.378A, 1982).
In evaluating the killing capabilities of mono-cytes for gonococci, the phagocytic capacity ofthe monocyte should also be considered. Alesser capacity for phagocytosis by humanmonocytes as compared to neutrophils has beenshown with Listeria sp., E. coli, S. aureus, andSalmonella sp. (21, 25, 30). When both cell typeswere permitted to ingest equal numbers of bacte-ria, no difference in the bactericidal activityagainst these organisms was seen. Pesanti et al.(20) found that murine macrophages that hadphagocytized large numbers of staphylococciingested substantially fewer organisms whenchallenged 1 h later. Thus, the greater ability ofhuman neutrophils for killing gonococci com-pared with monocytes (Fig. 3) may result from afaster rate of particle ingestion by neutrophils.The decline in viable gonococci over time was
a result of monocyte phagocytic killing since thepresence of 10 ,ug of cytochalasin B per mlinhibited the reduction in colony numbers. Cyto-chalasin B is a known inhibitor of phagocytosisthrough its actions on microfilament assembly(1). Also, when ingestible particles are present,treated phagocytes display "reverse endocyto-sis," during which lysosomes fuse to the interiorside of the cell membrane and release theircontents to the extracellular medium. Therefore,the absence of killing by cytochalasin B-treatedmonocytes indicates that ingestion was requiredand that killing was not the result of externalbactericidal activity. Sonication experimentsshow that internalized gonococci were indeedkilled and not simply sequestered within themonocytes. If a significant number of ingestedbacteria were in fact viable, they would havebeen released when the monocytes were disrupt-ed, and differences in colony numbers beforeand after sonication would have been apparent.
Killing of gonococci was clearly due to mono-cytes and not low numbers of contaminatingneutrophils, since PMNs were unable to kill
strain FA19 gonococci (Fig. 3) and suspension of<5 x 105 neutrophils per ml were insufficient tokill strain F62.
Last, the use of broth-grown, log- or station-ary-phase gonococci in place of plate-grownbacteria did not improve monocyte phagocytickilling. No apparent differences were detectedbetween the opaque and transparent organismsof both strains. Whereas killing of stationary-phase bacteria occurred in some experiments,the inability of monocytes to kill log-phase gono-cocci was consistently observed. These findingsdiffer from reports in which log- and stationary-phase, egg-grown gonococci were adequatelyphagocytized and killed by human neutrophils(10, 24). The use of broth-grown versus egg-grown bacteria in our experiments may be re-sponsible. A summary chart comparing the ob-servations on colony type, serum sensitivity,and monocyte and neutrophil phagocytic killingbetween the two strains of N. gonorrhoeae ispresented in Table 1.We conclude from these data that human
monocytes possess phagocytic killing capabili-ties against N. gonorrhoeae. It appears unlikelythat significant numbers of ingested organismssurvive intracellularly. However, conclusive ev-idence will require experiments in which nonin-gested, extracellular bacteria are completely re-moved from the phagocytic mixture prior to thedisruption of the monocytes. Activated or differ-entiated human MNP may show greater microbi-cidal activity toward gonococci than that pres-ently seen with freshly isolated monocytes.Those experiments are now in progress.
ACKNOWLEDGMENTSThis work was supported in part by Public Health Service
grant A114934 from the National Institutes of Health. R.F.R.is the recipient of Public Health Service Research CareerDevelopment award A100382 from the National Institutes ofHealth. We thank Linda Eaton for many helpful discussionsand Melin Canez and Nancy Gin for excellent technicalassistance.
INFECT. IMMUN.104 MEZZATESTA AND REST
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

MONOCYTE KILLING OF N. GONORRHOEAE 105
LITERATURE CITED
1. Axline, S. G., and E. P. Reaven. 1974. Inhibition of phago-cytosis and plasma membrane motility of the cultivatedmacrophage by cytochalasin B. Role of subplasmalemmalmicrofilaments. J. Cell Biol. 62:647.
2. Blake, M., and J. Swanson. 1975. Studies on gonococcusinfection. IX. In vitro decreased association of piliatedgonococci and mouse peritoneal macrophages. Infect.Immun. 11:1402-1404.
3. Casey, S. G., D. R. Veale, and H. Smith. 1979. Demon-stration of intracellular growth of gonococci in humanphagocytes using spectinomycin to kill extracellular or-ganisms. J. Gen. Microbiol. 113:395-398.
4. Cooper, M. D., and S. A. Floyd. 1982. In vitro kinetics ofphagocytosis and intracellular killing of gonococci byperitoneal macrophages from mice deficient in comple-ment component 5. Infect. Immun. 36:363-370.
5. Densen, P., and G. L. Mandell. 1978. Gonococcal interac-tions with polymorphonuclear neutrophils. Importance ofthe phagosome for bactericidal activity. J. Clin. Invest.62:1161-1171.
6. Densen, P., M. F. Rein, J. A. Sullivan, and G. L. Mandell.1978. Morphological observations of neutrophil-gonococ-cus interaction, p. 213-219. In G. F. Brooks, E. C.Gotschlich, K. K. Holmes, W. D. Sawyer, and F. E.Young (ed.), Immunobiology of Neisseria gonorrhoeae.American Society for Microbiology, Washington, D.C.
7. Dilworth, J. A., J. W. Hendley, and G. L. Mandell. 1975.Attachment and ingestion of gonococci by human neutro-phils. Infect. Immun. 11:512-516.
8. Drutz, D. J. 1978. Intracellular fate of Neisseria gonor-rhoeae, p. 232-235. In G. F. Brooks, E. C. Gotschlich,K. K. Holmes, W. D. Sawyer, and F. E. Young (ed.),Immunobiology of Neisseria gonorrhoeae. American So-ciety for Microbiology, Washington, D.C.
9. Ehlenberger, A. G., and V. Nussenzweig. 1977. The role ofmembrane receptors for C3b and C3d in phagocytosis. J.Exp. Med. 145:357-371.
10. Gibbs, D. L., and R. B. Roberts. 1975. The interaction invitro between human polymorphonuclear leukocytes andNeisseria gonorrhoeae cultivated in the chick embryo. J.Exp. Med. 141:155-171.
11. Jones, R. B., and T. M. Buchanan. 1978. Quantitativemeasurement of phagocytosis of Neisseria gonorrhoeaeby mouse peritoneal macrophages. Infect. Immun.20:732-738.
12. Jones, R. B., J. C. Newland, D. A. Olsen, and T. M.Buchanan. 1980. Immune-enhanced phagocytosis of Neis-seria gonorrhoeae by macrophages: characterization ofthe major antigens to which opsonins are directed. J. Gen.Microbiol. 121:365-372.
13. Kellogg, D. S., Jr., I. R. Cohen, L. C. Norins, A. L.Schroeter, and G. Reising. 1968. Neisseria gonorrhoeae.II. Colonial variation and pathogenicity during 35 monthsin vitro. J. Bacteriol. 96:596-605.
14. Kellogg, D. S., Jr., W. L. Peacock, Jr., W. E. Deacon, L.Brown, and C. I. Pirkle. 1963. Neisseria gonorrhoeae. I.Virulence genetically linked to clonal variation. J. Bacter-iol. 85:1274-1279.
15. Leih, P. C. J., M. T. van den Barselaar, T. L. van Zwet,M. R. Daha, and R. van Furth. 1979. Requirement ofextracellular complement and immunoglobulin for intra-
cellular killing of microorganisms by human monocytes.J. Clin. Invest. 63:772-784.
16. Novotny, P., J. A. Short, and P. D. Walker. 1975. Anelectron microscope study of naturally occurring andcultured cells of Neisseria gonorrhoeae. J. Med. Micro-biol. 8:413-427.
17. Ofek, I., E. H. Beachey, and A. L. Bisno. 1974. Resistanceof Neisseria gonorrhoeae to phagocytosis: relationship tocolonial morphology and surface pili. J. Infect. Dis.129:310-316.
18. Ota, F., J. Morita, N. Yoshida, F. Ashton, and B. Diena.1975. Studies on gonoccal infection. I. Electron micro-scopic studies on phagocytosis of Neisseria gonorrhoeaeby macrophages. Jpn. J. Microbiol. 19:149-155.
19. Ovcinnikov, N. M., and V. V. Delektorskij. 1971. Electronmicroscope studies of gonococci in the urethral secretionsof patients with gonorrhoea. Br. J. Vener. Dis. 47:419-439.
20. Pesanti, E. L., and K. M. Nugent. 1981. Inhibition ofmacrophage phagocytosis after contact with ingestibleparticles. RES J. Reticuloendothel. Soc. 30:157-166.
21. Peterson, P. K., J. Verhoef, D. Schmeling, and P. G. Quie.1977. Kinetics of phagocytosis and bacterial killing byhuman polymorphonuclear leukocytes and monocytes. J.Infect. Dis. 136:502-509.
22. Punsalang, A. P., Jr., and W. D. Sawyer. 1973. Role of piliin the virulence of Neisseria gonorrhoeae. Infect. Immun.8:255-263.
23. Rest, R. F., S. H. Fischer, Z. Z. Ingham, and J. F. Jones.1982. Interactions of Neisseria gonorrhoeae with humanneutrophils: effects of serum and gonococcal opacity onphagocyte killing and chemiluminescence. Infect. Immun.36:737-744.
24. Schiller, N. L., G. L. Friedman, and R. B. Roberts. 1979.The role of natural IgG and complement in the phagocyto-sis of Type 4 Neisseria gonorrhoeae by human polymor-phonuclear leukocytes. J. Infect. Dis. 140:698-707.
25. Steigbigel, R. T., L. H. Lambert, Jr., and J. S. Reming-ton. 1974. Phagocytic and bactericidal properties of nor-mal human monocytes. J. Clin. Invest. 53:131-142.
26. Swanson, J. 1978. Studies on gonococcus infection. XII.Colony color and opacity variants of gonococci. Infect.Immun. 19:320-331.
27. Thomas, D. W., J. C. Hill, and F. J. Tyeryar, Jr. 1973.Interaction of gonococci with phagocytic leukocytes frommen and mice. Infect. Immun. 8:98-104.
28. Thongthai, C., and W. D. Sawyer. 1973. Studies on thevirulence of Neisseria gonorrhoeae. I. Relation of colo-nial morphology and resistance to phagocytosis by poly-morphonuclear leukocytes. Infect. Immun. 7:373-379.
29. Veale, D. R., H. Finch, and H. Smith. 1976. Penetration ofpenicillin into human phagocytes containing Neisseriagonorirhoeae: intracellular survival and growth at opti-mum concentrations of antibiotic. J. Gen. Microbiol.95:353-363.
30. Verbrugh, H. A., R. Peters, P. K. Peterson, and J. Ver-hoef. 1978. Phagocytosis and killing of staphylococci byhuman polymorphonuclear and mononuclear leukocytes.J. Clin. Pathol. 31:539-545.
31. Witt, K., D. R. Veale, and H. Smith. 1975. Resistance ofNeisseria gotnorrhoeae to ingestion and digestion byphagocytes of human buffy coat. J. Med. Microbiol. 9:1-12.
VOL. 42, 1983
on July 13, 2018 by guesthttp://iai.asm
.org/D
ownloaded from





























