Embed Size (px)
Citation preview

RESEARCH ARTICLE Open Access
Peptide Deformylase (def) is essential inMycobacterium smegmatis, but theessentiality is compensated by inactivationof methionine formylationNoga Naor†, Omer Gadot†, Michal Meir2 and Daniel Barkan1*
Abstract
Background: Co-translational processes in bacteria are attractive drug targets, but while some processes areessential, others are not. The essentiality of Peptide Deformylase (PDF, def) for vitality of mycobacteria wasspeculated, but never unequivocally proven.
Results: Here we show by targeted deletion experiments that def can only be deleted from M. smegmatis when anadditional copy is present; that prior deletion of tRNAfMet-Formyl Transferase (FMT, encoded by fmt) renders defcompletely dispensable; and that re-introduction of fmt into a Δdef mutant is not possible – constituting adefinitive proof for the essentiality of def in mycobacteria.
Conclusions: Peptide deformylase is essential in M. smegmatis, but the fact that inactivation of fmt renders the genecompletely dispensable, and thus any inhibitor of def useless, casts doubt on the usefulness of PDF as a drug-target inmycobacteria.
Keywords: Mycobacteria, Protein synthesis, Co-translation, Peptide Deformylase, Formyl Transferase
BackgroundMycobacteria continue to be a major health threatthroughout the world. M. tuberculosis causes over 1.6million deaths worldwide [1], but the burden of M.leprae and M. ulcerans (mostly in the developing world),and that of pathogenic rapidly-growing mycobacteriasuch as M. abscessus and M. kansasii is rising as well[2–4], mostly in the developed world. Many mycobac-teria exhibit substantial drug resistance and tolerance,making the need for novel therapeutics more urgent [5].Understanding of physiologic processes unique to bac-teria is important for identifying essential pathways thatcould be targeted. One such process is the prokaryote-specific formylation and de-formylation of the methio-nine that starts the synthesis of every protein [6, 7]. In
mycobacteria, methionine is first formylated by tRNAf-
Met-Formyl Transferase (FMT, encoded by fmt), and onlya formylated methionine can be used as the first aminoacid in a new protein [8, 9]. Shortly after it emerges fromthe ribosome, the leading methionine is de-formylatedby Peptide Deformylase (PDF, encoded by def). Follow-ing deformylation, methionine is removed from the ma-jority of mycobacterial proteins by either MethionineAmino-Peptidase A (MetAPa, encoded by mapA) orMetAPc (encoded by mapB) [8, 10]. FMT was suggestedto be essential by a transposone-mutant library analysisin M. tuberculosis [11–13], but directed deletion experi-ments showed it to be completely dispensable for M.smegmatis, and to cause only mild growth retardation inM. bovis when deleted [9]. In MetAP’s, chemical inhib-ition studies first suggested mapA was essential whereasmapB was not [8], but directed gene deletion experi-ments on M. bovis proved it to be the other way around– mapB, and not mapA, was the essential one, withsome redundancy [10].
© The Author(s). 2019 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, andreproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link tothe Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver(http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
* Correspondence: [email protected]†Noga Naor and Omer Gadot contributed equally to this work.1Koret School of Veterinary Medicine, The Robert H. Smith Faculty ofAgriculture, Food and Environment, The Hebrew University of JerusalemRehovot Campus, Rehovot, IsraelFull list of author information is available at the end of the article
Naor et al. BMC Microbiology (2019) 19:232 https://doi.org/10.1186/s12866-019-1611-7

PDF (and def) is postulated to be vital in all bacteria –probably as MetAP’s cannot function on proteins wherethe formyl group was not removed [14]. def was sug-gested to be essential in M. bovis-BCG, and a chemicalinhibitor of it had antibacterial activity [15]. However,given the fact chemical inhibition experiments, as well asthe genetic method used to prove essentiality in thatstudy, may mistake a slow-growth phenotype for a lethalone, thus leading to incorrect essentiality attributions (aswas the case for fmt and mapA/mapB). We opted to de-finitively examine PDF’s role in mycobacterial viabilityby deletion experiments, as well as deletion on an fmt-null background.
Resultsdef cannot be deleted from the chromosome of M.smegmatisFor the deletion of def, we used the two step allelic ex-change system, for which we constructed the plasmidpDB354. The plasmid contains both zeocinR and strepto-mycinR resistance genes (for positive selection), sacB (fornegative selection), and the two flanking regions of def(~ 600 bp each, placed adjacent to each other). The plas-mid was electroporated into wt M. smegmatis, colonieswere selected on zeocin and streptomycin, and examinedfor successful recombination at one of the def flankingregions by PCR. One correct colony was found, andnamed M.smegint (for “intermediate”). These bacteriawere grown without antibiotic selection for 15 genera-tions to allow for a second recombination event, andplated on sucrose 10% for negative selection. One hun-dred colonies that grew on sucrose were patched on ei-ther no selection or zeocin/streptomycin to differentiatetrue second recombination events from sacB inactivatingmutations. Sixty colonies were found to be true second
recombination events (sucroseR/ZeocinS/streptomycinS).All were examined by PCR that produces an 1800 bpfragment in the case of reversal to wt, and 1200 bp incase of a def deletion genotype. All 60 colonies werefound to revert to wt, with none having the deletiongenotype (Fig. 1). We therefore conclude that deletion ofdef is, at the very least, highly deleterious in wt M.smegmatis.
In the presence of an additional copy, def can be deletedWe complemented M.smegint with an additional copy ofdef at the attB site, on a hygromycin-selected plasmid. Wethen proceeded to the second recombination as describedin the previous section. This time, out of the 12 coloniesthat underwent a second recombination event, four weredef deletions, whereas the other eight reverted to wt geno-type (Fig. 2), indicating the in the presence of an add-itional copy of def, the native def is completely dispensable(and that the deletion strategy used was valid).
The lethality of def deletion is compensated by tRNAfMet-Formyl Transferase (FMT, fmt) inactivationThe physiologic function of the def product (PeptideDeformylase, PDF) is to remove the formyl group fromthe leading methionine. Since this formyl group is ori-ginally attached by another enzyme, the tRNAfMet-For-myl Transferase (FMT, encoded by fmt), we speculatedthat on an fmt-deletion background, def may becomedispensable. We previously described a complete dele-tion of fmt in M. smegmatis (strain mDB22) and in M.bovis [9]. Contrary to previous reports, the deletion offmt was not lethal, causing only a trivial growth defect inM. smegmatis, and a 2–3 fold slower growth in M. bovis[9]. We decided to examine if fmt deletion renders defcompletely dispensable. We created an mDB22int strain
Fig. 1 def cannot be deleted from wt M. smegmatis. On the right, colonies after a confirmed second recombination event were checked by PCRto differentiate def-deletion (resulting in 1.2 kb product) from reversal to wt (1.8 kb fragment). 13 colonies are shown, out of 60 examined. On theleft – wt control, and a true Δdef mutant (obtained as described in section 3.2)
Naor et al. BMC Microbiology (2019) 19:232 Page 2 of 6
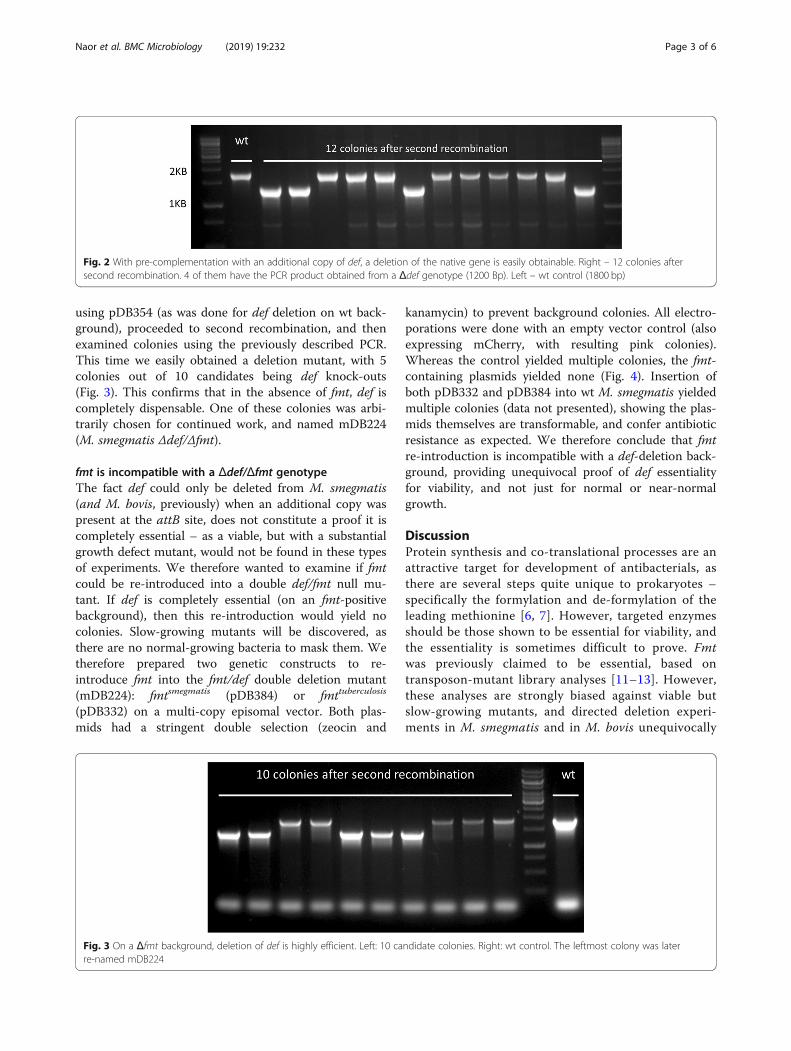
using pDB354 (as was done for def deletion on wt back-ground), proceeded to second recombination, and thenexamined colonies using the previously described PCR.This time we easily obtained a deletion mutant, with 5colonies out of 10 candidates being def knock-outs(Fig. 3). This confirms that in the absence of fmt, def iscompletely dispensable. One of these colonies was arbi-trarily chosen for continued work, and named mDB224(M. smegmatis Δdef/Δfmt).
fmt is incompatible with a Δdef/Δfmt genotypeThe fact def could only be deleted from M. smegmatis(and M. bovis, previously) when an additional copy waspresent at the attB site, does not constitute a proof it iscompletely essential – as a viable, but with a substantialgrowth defect mutant, would not be found in these typesof experiments. We therefore wanted to examine if fmtcould be re-introduced into a double def/fmt null mu-tant. If def is completely essential (on an fmt-positivebackground), then this re-introduction would yield nocolonies. Slow-growing mutants will be discovered, asthere are no normal-growing bacteria to mask them. Wetherefore prepared two genetic constructs to re-introduce fmt into the fmt/def double deletion mutant(mDB224): fmtsmegmatis (pDB384) or fmttuberculosis
(pDB332) on a multi-copy episomal vector. Both plas-mids had a stringent double selection (zeocin and
kanamycin) to prevent background colonies. All electro-porations were done with an empty vector control (alsoexpressing mCherry, with resulting pink colonies).Whereas the control yielded multiple colonies, the fmt-containing plasmids yielded none (Fig. 4). Insertion ofboth pDB332 and pDB384 into wt M. smegmatis yieldedmultiple colonies (data not presented), showing the plas-mids themselves are transformable, and confer antibioticresistance as expected. We therefore conclude that fmtre-introduction is incompatible with a def-deletion back-ground, providing unequivocal proof of def essentialityfor viability, and not just for normal or near-normalgrowth.
DiscussionProtein synthesis and co-translational processes are anattractive target for development of antibacterials, asthere are several steps quite unique to prokaryotes –specifically the formylation and de-formylation of theleading methionine [6, 7]. However, targeted enzymesshould be those shown to be essential for viability, andthe essentiality is sometimes difficult to prove. Fmtwas previously claimed to be essential, based ontransposon-mutant library analyses [11–13]. However,these analyses are strongly biased against viable butslow-growing mutants, and directed deletion experi-ments in M. smegmatis and in M. bovis unequivocally
Fig. 2 With pre-complementation with an additional copy of def, a deletion of the native gene is easily obtainable. Right – 12 colonies aftersecond recombination. 4 of them have the PCR product obtained from a Δdef genotype (1200 Bp). Left – wt control (1800 bp)
Fig. 3 On a Δfmt background, deletion of def is highly efficient. Left: 10 candidate colonies. Right: wt control. The leftmost colony was laterre-named mDB224
Naor et al. BMC Microbiology (2019) 19:232 Page 3 of 6

showed the gene was dispensable [9]. Similarly, achemical inhibition assay suggested MetAP1a was es-sential in M. tuberculosis, and that MetAP1c was not[8]. Again, directed deletion experiments showed theopposite [10]. As opposed to the well-established,complete essentiality of MetAp’s in all life forms [17,18], the role of DEF in different organisms is deemedcontroversial [19]. A targeted deletion in M. bovis-BCG only yielded colonies when an additional copy ofthe gene was present (in a two-step allelic exchangetechnique), leading to the conclusion def was essential(as also expected from what is known in otherbacteria) [15]. However, these analyses are based ontesting of several dozen colonies appearing on anegative-selection plate. Even if the gene in question iscompletely dispensable, at least 50% of colonies willrevert to wt, and if the deletion of the gene is viablebut causes growth retardation, the proportion of thedeletion mutants will become much smaller, as the wtcolonies will appear well before the deletion ones.Therefore, this kind of experiment can suggest an im-portant role in growth, but not absolute essentiality.Here, we first repeated the previous path, where weshowed the gene could be deleted when an additionalcopy was present, and no deletion mutants could beobtained when no pre-complementation was done. Wethen showed that on a background of fmt deletion, therole of def becomes completely insignificant, as evi-dent by a 50% deletion rate versus reversal to wt.However, we then showed that re-introduction of fmt(both smegmatis and tuberculosis origin) is impossibleinto a mutant that has no def – meaning a functionalfmt is incompatible with a Δdef genotype – and thisconstitutes a definitive proof of def essentiality.Why deformylation is essential is not completely clear.
One can postulate the formyl group blocks the activity ofthe methionine aminopeptidase enzymes (MetAP), who’sactivity is known to be essential in all life forms [17], in-cluding prokaryotes. One should remember the leadingmethionine is not removed from all synthesized proteins,
and it is possible that in proteins retaining this methio-nine, the formyl group may be retained as well, with nocompromise of viability. This may explain how mitochon-drial proteins in eukaryotes constantly retain the formylgroup [18, 20, 21]. Nonetheless, genes homologous to defwere shown to exist in the nuclear genomes of higher eu-karyotes, together with studies demonstrating mitochon-drial DEF expression with highly selective functions inhuman cells [19, 22, 23]. This suggests DEF does play arole in specific deformylations in eukaryotic organisms aswell. Moreover, it was shown that PDF of human origincould compensate for def deficiency in an E. coli mutantwith conditional expression of def [24], pointing the con-jecture that functionally, human PDF might be similar tothe bacterial one.The essentiality of def raises the question of whether it
can serve as an effective drug target. A previous studyshowed chemical inhibition of def does have antimyco-bacterial properties [25]. However, it appears from ourstudy that in the case of fmt inactivation, the inhibitionof def becomes irrelevant to growth. The barrier forspontaneous resistance mutations is therefore relativelylow, as inactivating mutations (unlike mutations that re-tain activity, but do not bind an inhibitor) are fairlycommon. It remains to be seen if the co-translationpathway is indeed a viable drug target.
ConclusionsPDF (def) was unequivocally shown to be essential in M.smegmatis, and probably in all mycobacteria. However, asgenetic inactivation of FMT (fmt) rendered the bacteriacompletely neutral as for the existence or absence of PDF,the role of PDF as a viable drug target remains question-able. It could, however, be part of a combination regimen,with other drugs that act on non-related targets.
MethodsStrains and growth conditionsMycobacterium smegmatis (mc2–155) were grown in Mid-dlebrook 7H9 media, supplemented with 0.5% glycerol,
Fig. 4 A functional fmt is incompatible with a Δfmt/Δdef genotype. mDB224 (Δfmt/Δdef) was electroporated with a control vector (left),control+fmttuberculosis (middle), and control+fmtsmegmatis (right). The picture was taken after 4 days incubation, but plates were kept for 10 daysbefore discarding, not to miss slow-growing mutant
Naor et al. BMC Microbiology (2019) 19:232 Page 4 of 6

0.5% dextrose, and 0.02% Tween80. For solid agar plates,we used 7H10 Middlebrook with glycerol and dextrose,but no tween. Antibiotic concentrations were: kanamy-cin 40 μg/mL, zeocin 25 μg/mL, streptomycin 20 μg/mL,and hygromycin 50 μg/mL (150 μg/mL for E. coli). ΔfmtM. smegmatis was previously described [9].
Deletion and complementation of defDeletion was done in the two-step allelic exchange, aspreviously widely described [16]. The plasmid used forthe first step recombination was pDB354. The upstreamand downstream flanking regions of def were PCR amp-lified from M. smegmatis genome, fused into a singlePCR product using primers defsmko1–5, and clonedinto a plasmid containing zeocinR, streptomycinR andsacB. After the second recombination, to differentiatebetween a deletion mutant and reversal to wt, we usedprimers defKOchqD and defKOchqU that produce an1800 bp fragment in wt, and 1200 bp fragment in a dele-tion mutant. To verify the 1200 bp product is indeed theresult of a successful deletion, the PCR product was sentfor Sanger sequencing, and was shown to be identical tothe wt-genotype, 1800 bp product at both ends, but tolack the bulk of the def gene, as constructed in the plas-mid used for the deletion (pDB354), resulting in short-ening by ~ 600 bp. To complement with an additionalcopy of defsmegmatis, def was amplified using primersdefSmU and defSmD, and cloned into the integratingplasmid pYUB412. All the primers sequences are shownin Additional file 1: Table S1.
Complementation with fmtA multicopy episomal plasmid, selected by zeocin andkanamycin, and also expressing mCherry, with a copy offmttuberculosis was previously described by us (pDB332, 9].A similar plasmid (with no mCherry) for fmtsmegmatis wasconstructed using primers fmt smg_R_xbaI, and fmtsmg_BamHI, and cloned into pDB151, producingpDB384. Electroporations were done as previouslywidely described.
Supplementary informationSupplementary information accompanies this paper at https://doi.org/10.1186/s12866-019-1611-7.
Additional file 1: Table S1. Primers (oligonucleotides) used in thisstudy.
AbbreviationsFMT: Formyl methionine transferase; MetAP: Methionine aminopeptidase;PDF: Peptide deformylase; WT: Wild type
AcknowledgementsNot applicable.
Author’s contributionsNN and OG performed most of the experiments. NN and MM participated inthe draft and final writing. MM and DB conceptualized the study. DBperformed experiments and wrote the final version.
FundingMM is supported by an internal grant from Rambam Medical Canter,“Ofakim”, and by the Israeli Science Fund (ISF). The granting body did notparticipate in the design of the study, in collection, analysis, andinterpretation of data, nor in writing the manuscript.
Availability of data and materialsNo large databases were used for this study. The datasets used and/oranalyzed during the current study are available from the correspondingauthor on reasonable request.
Ethics approval and consent to participateNot applicable.
Consent for publicationNot applicable.
Competing interestsThe authors declare that they have no competing interests.
Author details1Koret School of Veterinary Medicine, The Robert H. Smith Faculty ofAgriculture, Food and Environment, The Hebrew University of JerusalemRehovot Campus, Rehovot, Israel. 2The Ruth Rappaport Children’s Hospital,Rambam Health Care Campus, Haifa, Israel.
Received: 20 August 2019 Accepted: 6 October 2019
References1. WHO. Global Tuberculosis Report 2017. Geneva, Switzerland; 2018.2. De Groote MA, Huitt G. Infections due to rapidly growing mycobacteria.
Clin Infect Dis. 2006;42:1756–63.3. Snapper SB, Melton RE, Mustafa S, Kieser T, Jr WRJ. Isolation and
characterization of efficient plasmid transformation mutants ofMycobacterium smegmatis. Mol Microbiol. 1990;4:1911–9.
4. Duker AA, Portaels F, Hale M. Pathways of Mycobacterium ulceransinfection: a review. Environ Int. 2006;32:567–73.
5. Zhang Y. The magic bullets and tuberculosis drug targets. Annu RevPharmacol Toxicol. 2005;45:529–64.
6. Anderson WF. Initiation of protein synthesis in prokaryotic and eukaryoticsystems summary of EMBO workshop held in Noordwijkerhout, theNetherlands on 17–19 April 1974. FEBS Lett. 1974;48:1–6.
7. Giglione C, Pierre M, Meinnel T. Peptide deformylase as a target for newgeneration, broad spectrum antimicrobial agents: MicroReview. MolMicrobiol. 2000;36:1197–205.
8. Olaleye O, Raghunand TR, Bhat S, He J, Tyagi S, Lamichhane G, Gu P, ZhouJ, Zhang Y, Grosset J. Methionine aminopeptidases from Mycobacteriumtuberculosis as novel antimycobacterial targets. Chem Biol. 2010;17:86–97.
9. Vanunu M, Lang Z, Barkan D. The gene fmt, encoding tRNA fMet-formyltransferase, is essential for normal growth of M. bovis, but not for viability.Sci. Rep. 2017;7:15161.
10. Vanunu M, Schall P, Reingewertz T-H, Chakraborti PK, Grimm B, Barkan D.MapB protein is the essential methionine Aminopeptidase inMycobacterium tuberculosis. Cells. 2019;8:393.
11. Griffin JE, Gawronski JD, DeJesus MA, Ioerger TR, Akerley BJ, Sassetti CM.High-resolution phenotypic profiling defines genes essential formycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011;7:e1002251.
12. Lamichhane G, Zignol M, Blades NJ, Geiman DE, Dougherty A, Grosset J,Broman KW, Bishai WR. A postgenomic method for predicting essentialgenes at subsaturation levels of mutagenesis: application to Mycobacteriumtuberculosis. Proc Natl Acad Sci. 2003;100:7213–8.
13. Sassetti CM, Boyd DH, Rubin EJ. Genes required for mycobacterial growthdefined by high density mutagenesis. Mol Microbiol. 2003;48:77–84.
Naor et al. BMC Microbiology (2019) 19:232 Page 5 of 6

14. Yuan Z, Trias J, White RJ. Deformylase as a novel antibacterial target. DrugDiscov Today. 2001;6:954–61.
15. Teo JWP, Thayalan P, Beer D, Yap ASL, Nanjundappa M, Ngew X,Duraiswamy J, Liung S, Dartois V, Schreiber M. Peptide deformylaseinhibitors as potent antimycobacterial agents. Antimicrob AgentsChemother. 2006;50:3665–73.
16. Pavelka MS, Jacobs WR. Comparison of the construction of unmarkeddeletion mutations in Mycobacterium smegmatis, Mycobacterium bovisbacillus Calmette-Guerin, and Mycobacterium tuberculosis H37Rv by allelicexchange. J Bacteriol. 1999;181:4780–9.
17. Bradshaw RA, Brickey WW, Walker KW. N-terminal processing: themethionine aminopeptidase and Nα-acetyl transferase families. TrendsBiochem Sci. 1998;23:263–7.
18. Meinnel T. Peptide deformylase of eukaryotic protists: a target for newantiparasitic agents? Parasitol Today. 2000;16:165–8.
19. Lee MD, She Y, Soskis MJ, Borella CP, Gardner JR, Hayes PA, Dy BM, HeaneyML, Philips MR, Bornmann WG. Human mitochondrial peptide deformylase,a new anticancer target of actinonin-based antibiotics. J Clin Invest. 2004;114:1107–16.
20. Walker JE, Lutter R, Dupuis A, Runswick MJ. Identification of the subunits ofF1F0-ATPase from bovine heart mitochondria. Biochemistry. 1991;30:5369–78.
21. Yagi T, Hatefi Y. Identification of the dicyclohexylcarbodiimide-bindingsubunit of NADH-ubiquinone oxidoreductase (complex I). J Biol Chem.1988;263:16150–5.
22. Nguyen KT, Hu X, Colton C, Chakrabarti R, Zhu MX, Pei D. Characterizationof a human peptide deformylase: implications for antibacterial drug design.Biochemistry. 2003;42:9952–8.
23. Serero A, Giglione C, Sardini A, Martinez-Sanz J, Meinnel T. An unusualpeptide deformylase features in the human mitochondrial N-terminalmethionine excision pathway. J Biol Chem. 2003;278:52953–63.
24. Leeds JA, Dean CR. Peptide deformylase as an antibacterial target: a criticalassessment. Curr Opin Pharmacol. 2006;6:445–52.
25. Apfel CM, Locher H, Evers S, Takács B, Hubschwerlen C, Pirson W, PageMGP, Keck W. Peptide deformylase as an antibacterial drug target: targetvalidation and resistance development. Antimicrob Agents Chemother.2001;45:1058–64.
Publisher’s NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations.
Naor et al. BMC Microbiology (2019) 19:232 Page 6 of 6