Embed Size (px)
Citation preview

1985Development 124, 1985-1997 (1997)Printed in Great Britain © The Company of Biologists Limited 1997DEV1134
The Pax-6 gene encodes a transcription factor that isexpressed in regionally restricted patterns in the develop-ing brain and eye. Here we describe Pax-6 expression in theearly forebrain (prosencephalon) on embryonic day 9.5(E9.5) to E10.5 using both whole-mount in situ hybridiz-ation and antibody labeling. We find close correlationsbetween Pax-6+ domains and initial neural patterning, andidentify corresponding defects in embryos homozygous forthe Pax-6 allele, Small eye (Sey). Pax-6 expression definesthe prosencephalon-mesencephalon boundary, and mutantembryos lack this morphological boundary. Markers of thecaudal prosencephalon are lost (Pax-6, Lim-1, Gsh-1) anda marker for mesencephalon is expanded rostrally into theprosencephalon (Dbx). We conclude that the caudal pros-encephalon (prosomere 1) is at least partially transformedto a mesencephalic fate. This transformation results in a
specific deficit of posterior commissure axons. Sey/Seyembryos also exhibit an axon pathfinding defect specific tothe first longitudinal tract in the prosencephalon (tpoc,tract of the postoptic commissure). In wild type, tpoc axonsfan out upon coming in contact with a superficial patch ofPax-6+ neuron cell bodies. In the mutant, the tpoc axonshave normal initial projections, but make dramatic errorswhere they contact the neuron cell bodies, and fail topioneer this first tract. Thus Pax-6 is required for localnavigational information used by axons passing through itsdomain of expression.
We conclude that Pax-6 plays multiple roles in forebrainpatterning, including boundary formation, regional pat-terning, neuron specification and axon guidance.
Key words: homeobox, prosencephalon, Sey mutant, Pax-6, mouse
SUMMARY
Pax-6 functions in boundary formation and axon guidance in the embryonic
mouse forebrain
Grant S. Mastick*, Nicole M. Davis†, Gracie L. Andrews and Stephen S. Easter, Jr
Department of Biology, University of Michigan, Ann Arbor, Michigan 48109, USA†Present address: Ph.D. Program in Biological and Biomedical Sciences, Harvard Medical School, 220 Longwood Avenue, Boston, MA 02115, USA*Corresponding author (e-mail: [email protected])
INTRODUCTION
The differentiation of the first neurons in the vertebrateforebrain coincides with a rapid process of morphological andmolecular regionalization. The initial broad divisions of theneural tube form the classic three vesicles of the prosen-cephalon, mesencephalon, and rhombencephalon, embryonicstructures presaging the adult forebrain, midbrain andhindbrain. Further subdivisions form the neuromeres, a seriesof bulges separated by transverse constrictions in the prosen-cephalon (prosomeres; Puelles and Rubenstein, 1993), and therhombencephalon (rhombomeres; Lumsden and Keynes,1989). On the molecular level, a large number of regulatorygenes (many shared with Drosophila; Hirth et al., 1995) areexpressed in partially overlapping, regionally restrictedpatterns in the prosencephalon (reviewed by Puelles andRubenstein, 1993; Rubenstein and Puelles, 1994). In somecases, the borders of expression domains coincide withneuromere boundaries (Bulfone et al., 1993; Price et al., 1992),suggesting that borders of gene expression cause morphologi-cal boundaries.
This process of regionalization ultimately provides pos-itional information that is thought to regulate both the produc-tion of precise patterns of neuron cell bodies and the naviga-
tion of their axons. Supporting evidence for either part of thishypothesis has been limited to a few correlative studies. Forexample, gene expression borders coincide with the initialclusters of differentiating neuron cell bodies in the zebrafishprosencephalon (Macdonald et al., 1994). In mouse, the pros-encephalon-mesencephalon boundary coincides with thecaudal border of the neuron cell bodies that pioneer theposterior commissure (pc; Mastick and Easter, 1996). On thelevel of axons, some initial tracts in chick and fish form alonggene expression borders (Figdor and Stern, 1993; Macdonaldet al., 1994; Chedotal et al., 1995). In mouse, the leading axonsof the first tract, the tract of the postoptic commissure (tpoc),converge to cross a specific prosomere boundary, implying thatneuromeres and their boundaries may differentially influencenavigating axons (Mastick and Easter, 1996). These examplessupport the hypothesis that neuromeres and gene expressiondomains regulate neuronal patterning, but these general ideashave yet to be tested, for example, by perturbation of specificpatterning genes.
One candidate gene that could regulate prosencephalon pat-terning is Pax-6. This gene encodes a transcription factor withboth paired- and homeo-domain DNA binding motifs (Waltherand Gruss, 1991), and is expressed in the developing CNS ofa wide range of species, from vertebrates to nematodes. Most

1986 G. S. Mastick and others
studies of Pax-6 mutations have focused on eye development.Heterozygous Pax-6 mutations cause the Small eye (Sey)phenotype in mouse (Hill et al., 1991) and rat (Matsuo et al.,1993), as well as the human syndromes of aniridia (Ton et al.,1992; Glaser et al., 1992; Jordan et al., 1992) and Peter’sanomaly (Hanson et al., 1994). Pax-6 mutations in Drosophilahave been mapped to the eyeless gene (Quiring et al., 1994),and targeted overexpression of mouse Pax-6 in flies inducesectopic eyes, suggesting a highly conserved function (Halderet al., 1995).
Several lines of evidence suggest that Pax-6 also regulatesforebrain development, but the data are fragmentary. Pax-6 isexpressed very early in the rostral neural folds (Walther andGruss, 1991), with a caudal boundary thought to presage themorphological boundary between prosencephalon and mesen-cephalon. The broad prosencephalic expression becomesrestricted later to certain forebrain nuclei, suggesting functionsin specific subsets of neurons (Walther and Gruss, 1991;Stoykova and Gruss, 1994). Descriptions of mutant brains inboth late embryos and adults support this idea. Sey/Sey micehave substantial layering defects in forebrain in heterozygotesand homozygotes (Schmahl et al., 1993), and similar defectswere observed in a compound heterozygous human (Glaser etal., 1994). The late embryonic forebrain also appears to havedefects in regional patterning, as Sey/Sey embryos displaygross disturbances in regional growth in the diencephalon,accompanied by the failure of distinct nuclei to form (Stoykovaet al., 1996). Sey homozygous neonates also lack olfactorybulbs (Hogan et al., 1986). An earlier, more primary role inneuronal patterning was suggested by comparison of Pax-6expression and the initial scaffold of tracts in zebrafish(Macdonald et al., 1994). The tpoc axons project caudallyalong a Pax-6 expression boundary, and the tract of the pcforms just rostral to the caudal border of Pax-6. These corre-lations suggest that Pax-6 expression domains, or their borders,may pattern the initial neurons.
As a first step toward understanding the genetic regulationof early neuronal patterning in the prosencephalon, we haverecently described the relationship of prosomeres and initialneuronal patterns in the mouse (Easter et al., 1993; Mastickand Easter, 1996). In the context of this initial brain develop-ment, we relate Pax-6 expression patterns to prosomeres andneuronal patterns. To test the functional significance of thesecorrelations, we examine this neural patterning in homozygousSey mutant embryos, and find defects in specific prosomeres,neuronal cell bodies, and axon navigation.
MATERIALS AND METHODS
Mouse embryosPax-6 expression studies were carried out on CD-1 mouse embryos(Charles River Laboratories), obtained by timed matings, with noonof the day of the vaginal plug designated E0.5. The pregnant femaleswere killed using CO2 and the uteri were washed in ice-cold 0.1 Mphosphate. The embryos were dissected free in 4% paraformalde-hyde/0.1 M phosphate buffer, fixed in this solution for 2-4 hours atroom temperature, and stored in fixative at 4°C.
We primarily used the original Sey mutant allele (Roberts, 1967).The founder Sey/+ male was generously provided by AnthonyLaMantia (Duke University), and crossed to +/+ FVB females toproduce Sey/+ animals. Sey/+ animals were intercrossed, and approx-
imately 25% of their progeny had abnormalities of the optic vesicleand nasal pit diagnostic of Sey/Sey embryos (Hogan et al., 1986),while the remaining embryos appeared morphologically wild type.Our initial studies used these normal littermates (including both Sey/+and +/+) as controls for developmental stage in comparison to Sey/Seyembryos. We also examined three E10.5 litters from parents carryinganother Sey allele, Seyneu (Hill et al., 1991; Schmahl et al., 1993),which were generously provided by N. Brown and T. Glaser (Uni-versity of Michigan). The two alleles produced identical phenotypes(see Results).
GenotypingTo confirm the genotype of Sey/Sey embryos, and to distinguish Sey/+and +/+ embryos, we used a PCR strategy to genotype the individualembryos of one E9.5 litter (8 embryos) and two E10.5 litters (total of18 embryos). The Sey allele contains a single point mutation thatcreates a new DdeI restriction site (Hill et al., 1991), allowing PCR-based genotyping, essentially as recently described by Grindley et al.(1995). In brief, we isolated genomic DNA from the caudal half ofeach embryo, used flanking primers to amplify the Pax-6 exon 8, anddigested with DdeI, which produced a characteristic 55 bp band forthe mutant allele. The morphology and neurons of these individualgenotyped embryos were then examined, as described in the lastsection of Results.
Detection of expression of Pax-6 mRNA and proteinTo detect Pax-6 transcripts in whole mounts, a plasmid containing anEcoRI/NheI restriction fragment from the mouse Pax-6 cDNA(Walther and Gruss, 1991) was kindly provided by K. Backs and P.Gruss (Gottingen). Whole-mount in situ hybridization using a digox-igenin-labeled antisense RNA probe, detected by anti-digoxigeninantibody coupled to alkaline phosphatase, was carried out essentiallyaccording to the procedure of Wilkinson (1992), as modified by Parret al. (1993). The embryos were sagittally bisected, cleared in 80%glycerol, and photographed in a dissecting microscope.
To detect Pax-6 protein in 20 µm cryostat sections, a rabbitantiserum raised against a C-terminal peptide was generouslyprovided by J. Davis and R. Reed (Davis and Reed, 1996). Theantibody labeling protocol used dried milk as a blocker for non-specific binding (Sundin and Eichele, 1990), biotinylated goat anti-rabbit secondary antibody (Vector), followed by avidin-HRP and per-oxidase reactions using the purple VIP substrate (Vector). Thesections were dehydrated through an ethanol series into xylene, cov-erslipped with Eukitt (Calibrated Instruments, Inc.), and photographedin a compound microscope equipped with Nomarski optics.
For double labeling with Pax-6 antiserum and the neuron-specificβ-tubulin monoclonal antibody TuJ1 (Moody et al., 1987; Lee et al.,1990), the secondary antibodies were Cy3-conjugated anti-rabbit(Jackson Immunoresearch) and FITC-conjugated anti-mouse (Sigma),respectively. The sections were coverslipped in Prolong Antifademounting medium (Molecular Probes), and examined with a Bioradconfocal microscope.
Other probesThe Lim-1 (Lhx1) probe was synthesized from a 1.5 kb cDNAprovided by H. Sheng and H. Westphal (Fujii et al., 1994). The Gsh-1 probe was synthesized from a 799 bp cDNA fragment provided byT. Valerius and S. Potter (Valerius et al., 1995). The Pax-3 probe wassynthesized from a 519 bp cDNA fragment provided by K. Backs andP. Gruss (Goulding et al., 1991). The Dbx probe was synthesized froma cDNA fragment provided by S. Lu and F. Ruddle (Lu et al., 1992).
Neuromere morphologyNeuromeres were visualized in whole mounts after removal of skinand underlying mesenchyme. The embryos were then sagittallybisected, cleared in 50% glycerol/2% paraformaldehyde/0.05 Mphosphate, and photographed in a dissecting microscope using

1987Initial tract formation in Small eye
glancing illumination against a dark field. The neuromeres were alsovisualized in cryostat sections cut at 20 µm thickness, counterstainedwith toluidine blue, and mounted in Eukitt.
Neuronal labelingAxon tracing used DiI, a fluorescent lipophilic dye, applied to fixedembryos as previously described (Mastick and Easter, 1996). Fordouble labeling procedures, the fluorescence was photoconverted to astable product (McConnell et al., 1989). In brief, DiI-labeled embryoswere washed several times to remove formaldehyde for a total of 1hour in 0.05 M Tris, pH 8.3, including 5 mg/ml heparin as an RNAseinhibitor (Shimamura et al., 1995), transferred to the same solutioncontaining 0.6 mg/ml DAB, incubated on ice for 30 minutes, and cov-erslipped in a depression slide. Illumination of the embryos for 15-20minutes with a 25× objective converted the fluorescently labeledneurons to a yellow-brown color.
Neurons were also labeled in whole mounts and sections with theTuJ1 antibody, using an HRP-conjugated secondary antibody, andDAB reactions, as previously described (Easter et al., 1993; Mastickand Easter, 1996).
RESULTS
We first introduce the morphological conventions andlandmarks used in the prosencephalon, and then present theresults in two sections, the first describing the Pax-6 expressionpattern, and the second describing the morphological andneuronal defects in Sey/Sey mutant embryos.
We use a modified version of the morphological conventionsproposed by Puelles and Rubenstein (1993). The dorsal-ventraldirections are premised on the belief that the longitudinal axisof the neural tube is sharply bent ventrally at the cephalicflexure (Fig. 1A). The ventral surfaces of the prosencephalonand rhombencephalon face each other, so the dorsal-ventraldirection depends on position along the longitudinal axis. Thecaudal prosencephalon contains two bulges, prosomeres 1 and2 (p1, p2), flanked by transverse constrictions in the dorsalneural tube (Mastick and Easter, 1996; shown here in Figs 1B,4). Rostral to p2, four more subdivisions have been postulated(p3-p6; Puelles and Rubenstein, 1993), but are not evident onE9.5 or 10.5. We therefore refer to the region immediatelyrostral to p2 as p3, leaving the p3 rostral boundary undefined,as well as p4-p6. The rostral prosencephalon contains twodorsal evaginations: the cerebral vesicle and the optic vesicle.Caudal to p1 is the mesencephalon (mes), a large, apparentlyundivided neuromere, followed by the rhombomeres makingup the hindbrain (r1-r8). According to Puelles and Rubenstein(1993), the mes/p1 boundary becomes the dorsal midbrain-forebrain boundary in the adult, separating superior colliculusfrom pretectum, and the p2/p3 boundary becomes the zonalimitans interthalamica, separating dorsal and ventral thalamusin the adult (Puelles and Rubenstein, 1993; Stoykova et al.,1996).
Pax-6 expression patterns in the prosencephalonPax-6 expression and the prosomeresOverall, Pax-6 expression in the brain was extensive, region-ally specific and dynamic. On E9.5 (Fig. 1A), Pax-6 expressionwas limited to the dorsal neural tube, and extended through theentire prosencephalon, including the cerebral and opticvesicles, but excluding the roof plate (not shown). The mesen-
cephalon was unlabeled. In the rhombencephalon, r1 wasunlabeled, but r2 and more caudal rhombomeres were labeled.By E10.5 (Fig. 1D), the expression was reduced in some areas(notably p2), and increased in others (a heavily labeled clusterin caudal-ventral p1, a strip of scattered cells at the samedorsal-ventral level in mesencephalon, and a rostral expansionof the rhombencephalic strip into r1).
To determine more precisely the relationship between theborders of Pax-6 expression and neuromeric boundaries,sections perpendicular to the boundaries were labeled withPax-6 antibody. The prosomeres stood out clearly in suchsections (Fig. 1B,C); p1 and p2 are external bulges, with com-plementary internal recesses, and they are separated from oneanother by constrictions, characterized by both externalgrooves and internal ridges. The caudal limit of the p1 Pax-6expression domain closely corresponded to the mes/p1boundary on E9.5. The discontinuity was quite sharp betweenthe unlabeled mesencephalon and the labeled p1. A few labelednuclei arranged in radial columns defined the border in somesections (Fig. 1C), but in others, the discontinuity was evenmore abrupt (not shown). The vast majority of p1 nuclei werelabeled (Fig. 1C). The labeling was not uniform, as most nucleinear the ventricular surface and a few near the pial surface weremore heavily labeled, and a few unlabeled nuclei (about 1 in50) were located near the pial surface. On E9.5, expression wascontinuous across the p1/p2 (Fig. 1C) and p2/p3 boundaries(not shown). By E10.5, p1 had signs of advancing differen-tiation, including unlabeled nuclei superficially (where differ-entiating neurons appear; Figs 1E, 2D), as well as a cluster ofheavily labeled nuclei just ventral to the main domain ofexpression (Fig. 2C). As Fig. 1F shows, labeling was reducedin p2, most dramatically among the deep nuclei just caudal tothe p2/p3 boundary, while p3 remained heavily labeled, espe-cially in a superficial layer of nuclei extending across theboundary. In summary, the dynamic spatial patterns of Pax-6expression in the mouse prosecephalon are similar to thoserecently described in zebrafish (Macdonald et al., 1994). Here,we extend this analysis to show that Pax-6 expression domainsare closely bounded by at least two prosomere boundaries, andPax-6 is expressed differentially along all three directions ofthe neuromeres: rostral-caudal, dorsal-ventral and superficial-deep.
Pax-6 expression and the pcTwo heavily labeled blocks of cells were evident in caudal p1(Fig. 2A). The arrangement of these cells (a dorsal block incaudal p1 and a ventral block in caudal p1 that extends intomes) was similar to that of the neuron cell bodies that projectaxons dorsally to pioneer the pc on E10.5 (Mastick and Easter,1996). In double-labeled embryos (Fig. 2B), the pc somataoverlapped extensively with the two blocks of Pax-6 label, butalso extended outside of the Pax-6-labeled blocks, bothventrally and in the gap between them. Therefore, at least somepc neurons did not express Pax-6. Whether any of the pcneurons were Pax-6+ could not be established with in situs,because the label was too weak to distinguish lightly labeledfrom unlabeled cells, but the antibody gave strong nuclearlabeling, so we used it on sections of p1 (Fig. 2C,D). Completesets of serial sections from two embryos were analyzed. Of atotal of 279 pc cell bodies, 260 (93%) lacked nuclear labeling,one nucleus was heavily labeled, nine faintly labeled, and nine
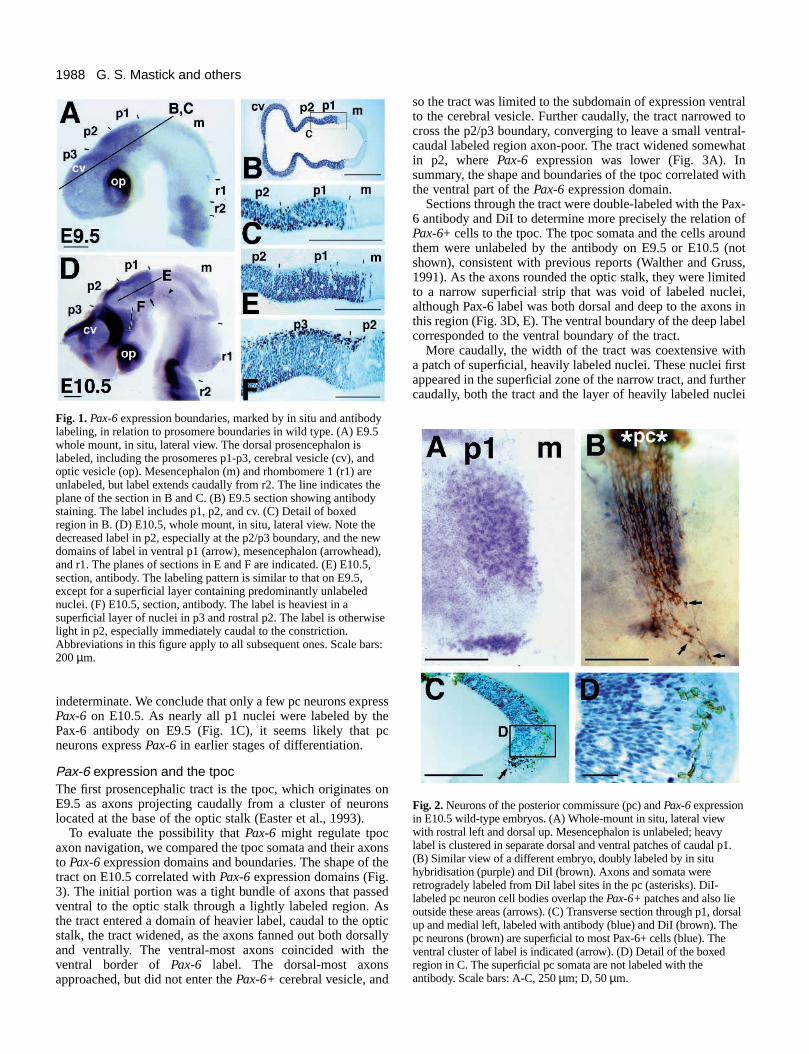
1988 G. S. Mastick and others
Fig. 1. Pax-6 expression boundaries, marked by in situ and antibodylabeling, in relation to prosomere boundaries in wild type. (A) E9.5whole mount, in situ, lateral view. The dorsal prosencephalon islabeled, including the prosomeres p1-p3, cerebral vesicle (cv), andoptic vesicle (op). Mesencephalon (m) and rhombomere 1 (r1) areunlabeled, but label extends caudally from r2. The line indicates theplane of the section in B and C. (B) E9.5 section showing antibodystaining. The label includes p1, p2, and cv. (C) Detail of boxedregion in B. (D) E10.5, whole mount, in situ, lateral view. Note thedecreased label in p2, especially at the p2/p3 boundary, and the newdomains of label in ventral p1 (arrow), mesencephalon (arrowhead),and r1. The planes of sections in E and F are indicated. (E) E10.5,section, antibody. The labeling pattern is similar to that on E9.5,except for a superficial layer containing predominantly unlabelednuclei. (F) E10.5, section, antibody. The label is heaviest in asuperficial layer of nuclei in p3 and rostral p2. The label is otherwiselight in p2, especially immediately caudal to the constriction.Abbreviations in this figure apply to all subsequent ones. Scale bars:200 µm.
Fig. 2. Neurons of the posterior commissure (pc) and Pax-6 expressionin E10.5 wild-type embryos. (A) Whole-mount in situ, lateral viewwith rostral left and dorsal up. Mesencephalon is unlabeled; heavylabel is clustered in separate dorsal and ventral patches of caudal p1.(B) Similar view of a different embryo, doubly labeled by in situhybridisation (purple) and DiI (brown). Axons and somata wereretrogradely labeled from DiI label sites in the pc (asterisks). DiI-labeled pc neuron cell bodies overlap the Pax-6+ patches and also lieoutside these areas (arrows). (C) Transverse section through p1, dorsalup and medial left, labeled with antibody (blue) and DiI (brown). Thepc neurons (brown) are superficial to most Pax-6+ cells (blue). Theventral cluster of label is indicated (arrow). (D) Detail of the boxedregion in C. The superficial pc somata are not labeled with theantibody. Scale bars: A-C, 250 µm; D, 50 µm.
indeterminate. We conclude that only a few pc neurons expressPax-6 on E10.5. As nearly all p1 nuclei were labeled by thePax-6 antibody on E9.5 (Fig. 1C), it seems likely that pcneurons express Pax-6 in earlier stages of differentiation.
Pax-6 expression and the tpocThe first prosencephalic tract is the tpoc, which originates onE9.5 as axons projecting caudally from a cluster of neuronslocated at the base of the optic stalk (Easter et al., 1993).
To evaluate the possibility that Pax-6 might regulate tpocaxon navigation, we compared the tpoc somata and their axonsto Pax-6 expression domains and boundaries. The shape of thetract on E10.5 correlated with Pax-6 expression domains (Fig.3). The initial portion was a tight bundle of axons that passedventral to the optic stalk through a lightly labeled region. Asthe tract entered a domain of heavier label, caudal to the opticstalk, the tract widened, as the axons fanned out both dorsallyand ventrally. The ventral-most axons coincided with theventral border of Pax-6 label. The dorsal-most axonsapproached, but did not enter the Pax-6+ cerebral vesicle, and
so the tract was limited to the subdomain of expression ventralto the cerebral vesicle. Further caudally, the tract narrowed tocross the p2/p3 boundary, converging to leave a small ventral-caudal labeled region axon-poor. The tract widened somewhatin p2, where Pax-6 expression was lower (Fig. 3A). Insummary, the shape and boundaries of the tpoc correlated withthe ventral part of the Pax-6 expression domain.
Sections through the tract were double-labeled with the Pax-6 antibody and DiI to determine more precisely the relation ofPax-6+ cells to the tpoc. The tpoc somata and the cells aroundthem were unlabeled by the antibody on E9.5 or E10.5 (notshown), consistent with previous reports (Walther and Gruss,1991). As the axons rounded the optic stalk, they were limitedto a narrow superficial strip that was void of labeled nuclei,although Pax-6 label was both dorsal and deep to the axons inthis region (Fig. 3D, E). The ventral boundary of the deep labelcorresponded to the ventral boundary of the tract.
More caudally, the width of the tract was coextensive witha patch of superficial, heavily labeled nuclei. These nuclei firstappeared in the superficial zone of the narrow tract, and furthercaudally, both the tract and the layer of heavily labeled nuclei

1989Initial tract formation in Small eye
widened, sharing the same dorsal and ventral boundaries (Fig.3B,C). In this wide zone, the most ventral of these nucleioverlay a ventricular zone lacking Pax-6, while the dorsal edgeof the tract was flanked by an unlabelled superficial region atthe hemispheric sulcus, and a labeled deep zone that extendedinto the cerebral vesicle. At the caudal expression border, the
Fig. 3. The tract of the postoptic commissure (tpoc) and Pax-6expression domains. (A) E10.5; whole mount, with the cv removedto reveal the tract; lateral view; doubly labeled by in situ (purple) andDiI (brown). The tpoc axons were labeled anterogradely at theasterisk. The tract is initially narrow, widens upon entering a Pax-6+domain, and narrows to cross the p2/p3 boundary. The ventral edgeof the widened tract closely corresponds to the ventral expressionborder, except in the region indicated by the arrowhead. The planesof sections shown in B-G are indicated. (B) E10.5 section; dorsal leftand medial below; labeled with antibody (blue) and DiI (brown). Theventricle (v) and hemispheric sulcus (hs) are indicated. The dorsaland ventral limits (arrowheads) of the tract coincide with the limits ofa superficial layer of nuclei heavily labeled by the antibody.(C) Enlarged view of the boxed area in B, showing the axons(yellow) intermixed with the heavily labeled superficial nuclei butseparate from the deeper, less heavily labeled ones. (D) Similar to B,but more rostral, at the level of the optic recess (or). The initial tightportion of the tract is associated with a superficial zone lackinglabeled nuclei. (E) Enlarged view of the boxed area in D. (F) Section;antibody labeling. The plane of section is similar to that of B, butlooks different because the cerebral vesicle (out of view, upper left)has not yet developed a deep hemispheric sulcus in this youngerembryo (E9.5). The superficial heavily labeled nuclei are alreadypresent, and presage the future path of the tpoc. (G) E10.5 section;similar to C, but at a higher magnification; immunolabeled with theneuronal marker, TuJ1 (green), and Pax-6 (red); confocal image. Thecells with high levels of nuclear Pax-6 label are neurons. Scale bars:A,B,D,F, 200 µm; C,E, 50 µm; G, 20 µm.
tract narrowed, and the axons became restricted to the dorsal-most heavily labeled nuclei (not shown), leaving the ventral-most nuclei in an axon-poor region, consistent with the wholemount (Fig. 3A). The most caudal of the heavily labeled nucleibridged the p2/p3 boundary (see Fig. 1F). These cells werepresent on E9.5 in a location that anticipated the tract (Fig. 3F).In summary, the wide zone of the tpoc was closely associatedwith a specific subset of Pax-6-expressing cells that precededthe tract, suggesting a role for these cells in axon guidance.
The superficial position of these heavily labeled nucleisuggested they were neurons, and this was confirmed by doublelabeling with TuJ1, a neuronal antibody (Fig. 3G). All of theheavily labeled nuclei were associated with the cytoplasmictubulin label, while few of the lightly labeled nuclei were. Weconclude that the superficial cells that express Pax-6 at highlevels are neurons.
The Small eye (Sey) prosencephalonAs a test for the function of Pax-6 in brain development, weexamined the prosencephalon in Sey embryos on E9.5 andE10.5, and found defects in general morphology, the pro-someres, the pc, and the tpoc.
We initially examined Sey/Sey embryos, which we will referto as ‘mutant’, using for comparison their morphologicallynormal littermates, pooling both +/+ and Sey/+ embryos,labeled as ‘wild type’. At the end of the Results, we describethe analysis of individual genotyped Sey/+ embryos, fromwhich we conclude that the early development of Sey/+embryos is almost entirely normal, and finish by describing theidentical mutant phenotype of another Pax-6 allele, Seyneu.
Brain morphologyHomozygous mutant embryos, readily distinguished by theirenlarged optic vesicles (E9.5 and E10.5) and the absence ofnasal pits (E10.5) (Hogan et al., 1986), appeared otherwisenearly normal. The overall size and number of somites waswithin the range of their wild-type littermates, and the mor-phology of the brain appeared relatively normal (Fig. 4A-D),both in size (measured by the length along the dorsal surface),and in shape (measured by the angle of the cephalic flexure).
The morphology of the cerebral vesicle was altered in themutant. In whole mounts, the groove marking the caudalboundary of the cerebral vesicle was less prominent on E9.5(Fig. 4A,B), as were the ventral and dorsal boundaries (notshown). By E10.5, the mutant cerebral vesicles were largerthan wild type, and their ventral boundary was indistinct (Fig.4C,D). Viewed from the dorsal side, the hemispheric sulcusformed a less acute angle between the cerebral vesicle and theadjoining prosencephalon. This morphological alteration ledto the condition shown in Fig. 4H, where the mutant cerebralvesicles were no longer separated from each other at the dorsalmidline, or from the ventral prosencephalon (compare to Fig.4G). The hemispheric sulcus was also less prominent in morerostral sections (compare Fig. 8A,C). The wall of the mutantcerebral vesicles also was thinner. Taken together, our obser-vations indicate very early defects both in the cerebral vesicleand in its junction with the ventral prosencephalon. Theseearly growth disturbances apparently continue into later devel-opment, as by E12.5 the third ventricle is abnormally large,with wide openings to the cerebral ventricles (Stoykova et al.,1996).

1990 G. S. Mastick and others
ll eye mutants have altered prosomeres. Analysis of whole mounts (A-ions (E-H) of E9.5 and 10.5 embryos, both wild type (WT) and mutantA-D) Unlabeled bisected embryos in dark-field illumination to show theinterneuromeric grooves (marked dorsally by white dashed). The mes/p1oundaries cannot be seen at either age in the mutant, while the p2/p3
s apparent, especially in D. The planes of the sections in E-H areE-H) Sections of E9.5 and E10.5 embryos, stained with toluidine blue,e lack of a mes/p1 boundary in the mutant. In G and H, similar planes ofw the altered morphology of the cerebral vesicles on E10.5. The dorsalrowhead), and the hemispheric sulcus (hs) are both less prominent in thele bars: 500 µm.
Defects in prosomere formationp1 and p2 were altered in the mutant (Fig. 4). In wild-typewhole mounts on E9.5, the groove marking the mes/p1boundary was apparent as an interface between light and darkareas when viewed with glancing illumination, but this was notseen in the mutant (Fig. 4A,B). Sections of several E9.5mutants (Fig. 4F) confirmed the absence of a neuromeric bulgewithin p1, and of an external groove and an internal ridge, thestructures normally at the boundary. In contrast, p2was prominent as a bulge, and an external grooveformed a caudal boundary with p1 in the expectedlocation. The mutant p1 neuroepithelium hadnormal thickness, but the mutant p2 was thinner.The p2/p3 boundary was apparent in wholemounts and sections (not shown).
The same defects were evident on E10.5. Themes/p1 boundary was still missing, so its absenceon E9.5 was unlikely due to a delay in the devel-opment of this region. The p1/p2 and p2/p3 bound-aries appeared normal (Fig. 4H,D). The p2/p3boundary was more visible in the mutant, probablybecause of the altered cerebral vesicle morphology(Fig. 4C,D).
The absence of the mes/p1 boundary in mutantsindicates a role for Pax-6 in the formation of thisboundary. Pax-6 is not apparently involved in theformation of the p1/p2 and p2/p3 boundaries, cor-relating with the lack, or delay, respectively, ofPax-6 expression borders at these locations in wildtype.
Alteration of p1 identityWith the absence of the mes/p1 boundary, the mor-phological distinction between mes and p1 waslost, raising the question of the fate of p1 in themutant. One possibility was that p1 regionalidentity was retained in the mutant, and that acryptic regional boundary remained in the absenceof a neuromeric boundary; alternatively, therecould be a loss of regional distinction between mesand p1.
To identify the fate of p1, we examined threehomeobox genes that were expressed in dorsal p1as markers for p1 identity: Pax-6, and two others,Lim-1 and Gsh-1, with expression that is initiatedon E10.5 (Fig. 5). The mutant Pax-6 transcript canbe used to label Pax-6 expression domains inmutant embryos (Grindley et al., 1995), since theSey allele contains a point mutation which does notdisrupt transcription (Hill et al., 1991). We foundthat the p1 domain of transcripts was missing inmutant embryos, and the most dorsal p2 label wasretained (Fig. 5A,B). This loss of Pax-6 in situlabeling was very region-specific, as other domainswere labeled as in wild type, and early, as the p1domain was lost prior to E9.5, the earliest stage weexamined. This result suggests the regulation ofPax-6 transcription is distinct in p1 and most of p2,requiring autoactivation as previously identified inlens ectoderm (Grindley et al., 1995). A secondmarker for p1 is Lim-1, which was previously
Fig. 4. SmaD) and sect(Sey/Sey). (transverse and p1/p2 bboundary iindicated. (showing thsection shomidline (armutant. Sca
described to be activated on E10.5 in a dorsal domain of rostralmesencephalon (Fujii et al., 1994). Upon reexamination of Lim-1 expression in wild type, we found that this dorsal domain wasin fact in p1, closely abutting the mes/p1 boundary on the caudalside, and extending rostrally to mid-p1, in a superficial layer ofcells (Fig. 5C, inset). Lim-1 was also expressed in several otherdomains, including a heavily labeled arc in ventral p1 and mes,a broad strip in rhombencephalon, and three patches of scattered

1991Initial tract formation in Small eye
cells in rostral prosencephalon. In the mutant, the dorsal p1domain of Lim-1 label was lost. The other domains remainedlargely unaffected, although the ventral p1 arc was less pointedat its rostral end, and the scattered cells just rostral to the p2/p3boundary were less prominent. Labeling in the spinal cord wasalso altered (not shown). A third marker for dorsal p1 is Gsh-1(Valerius et al., 1995). Sections demonstrated that Gsh-1 wasexpressed in a layer of cells that, in contrast to Lim-1, were ven-tricular, but like Lim-1, stretched from the mes/p1 boundaryrostrally to mid-p1. This dorsal p1 domain of Gsh-1 label wasmissing in the mutant, while other domains were labeled,including rhombencephalon, a strip of scattered cells in ventralp1 and mes, and another strip extending rostrally from the p2/p3boundary. This rostral strip appeared to be labeled less intenselyin the mutant. Taken together, markers for dorsal p1 (Pax-6,Lim-1, and Gsh-1) are all missing in the mutant, indicating thatthe loss of the p1/mes boundary is accompanied by the loss ofmarkers of the p1 side of the boundary.
Fig. 5. The identity of dorsal p1 is altered in Small eye mutants.Whole-mount in situ hybrization with probes against severalhomeobox genes. E10.5 wild-type and mutant littermates, withinsets in C,E,G,H showing sections through p1. (A,B) Pax-6.The wild-type embryo shown in A is the same as that in Fig.1A. In the mutant, the label in dorsal and ventral p1 is lost, andmost of dorsal p2 is unlabeled, except for a thin strip adjacentto the dorsal midline. The remaining Pax-6 domains appearnormal in the mutant, including the domain rostral to the p2/p3boundary, the cerebral and optic vesicles, a scattered strip ofcells (arrowhead) in ventral mes, and the rhombencephalon.(C) Lim-1. In wild type, Lim-1 labels dorsal-caudal p1, aventral strip extending from p1 into mes, and a strip from r1caudal. Additional labeled cells include scattered cells in p3,extending rostrally to the ventral side of the optic stalk, andpatches on the ventral, rostral, and dorsal margins of thecerebral vesicle. Inset in C: a section showing Lim-1 label inp1, in superficial cells that closely abut the mes/p1 boundary.(D) In the mutant, the dorsal domain of Lim-1 label isspecifically missing. (E,F) Gsh-1. In wild type, label is seen indorsal p1, several patches of scattered cells in the rostralprosencephalon, and in a strip from r1 caudal. Inset in E: asection through p1 showing Gsh-1 label in ventricular cells incaudal p1, closely abutting the mes/p1 boundary. In the mutant,Gsh-1 label is specifically lost from dorsal p1. (G,H) Pax-3. Inboth wild type and mutant, Pax-3 label extends rostrallythrough dorsal mes, p1, and p2. In the mutant p2, Pax-3 labeldoes not extend as far rostrally. (I,J) Dbx. In wild type, Dbxlabels dorsal mes, with a sharp border at the mes/p1 boundary(inset in I). Label in p1 and p2 is limited to a thin band ofventricular cells that curves from ventral p1 up into dorsal p2;there is also a heavily labeled domain in ventral p3, and a stripfrom r1 caudal. In the mutant, the mes domain is expandedrostrally, primarily in a superficial layer of cells (inset), and thethin band is lost from p1 and p2. Scale bars: A (applies to B-J),200 µm; C, inset (applies to insets in E, I, J), 200 µm.
The loss of p1-specific markers might not be gene-specific,but rather the result of a general reduction in transcription inthis domain. As a test for this possibility, we examined theexpression of Pax-3, another Pax family member that isexpressed in much of the dorsal CNS (Goulding et al., 1991).Pax-3 in situs labeled a dorsal domain that extended throughmes and p1, and tapered dorsally in p2 (Fig. 5G). Pax-3 thusoverlaps the mes/p1 boundary, and was coexpressed with Pax-6 in dorsal p1. In the mutant, Pax-3 labeling appeared unaf-fected in mes and p1, although labeling did not extend as farrostrally in p2 (Fig. 5H). We conclude that the dorsal p1 defectsin Pax-6, Lim-1, and Gsh-1 expression are not due to a generaldefect in gene transcription.
If p1-specific markers are lost, then what identity has beenassumed by this region? We examined the expression patternof the homeobox gene Dbx (Lu et al., 1992) as a marker forrostral mesencephalon. On E10.5 in wild type, Dbx wasexpressed in apparently all of the cells of dorsal mes, most

1992 G. S. Mastick and others
Fig. 6. Small eye mutants lack the posterior commissure (pc). Allpanels depict dorsal views of E10.5 whole mounts. (A,B) TuJ1antibody labeling, dorsal midline vertical, rostral up. The wild-typeroof plate of p1 contains pc axon bundles caudally, and neuron cellbodies rostrally (bracket); both are missing in the mutant. (C,D) DiIlabeling (asterisks), dorsal midline horizontal (dashed line), rostralleft. (C) In the wild type, both contralateral (pc) and ipsilateral fibersare labeled. On the ipsilateral side, many cell bodies are retrogradelylabeled (arrows point to examples of ventral pc cell bodies), as wellas anterogradely labeled axons, including the tract of themesencephalic nucleus of the trigeminal nerve (tmesV) andcircumferential descending axons (cda). (D) In the mutant, nocontralateral fibers are labeled, and on the ipsilateral side, onlyventral cell bodies (small arrows) in addition to tmesV and cda fibersare labeled. The arrowhead in C indicates a blood vessel that wasinadvertently labeled. Scale bar in A applies to all panels (250 µm).
heavily in superficial cells extending rostrally to the mes/p1boundary (Fig. 5 I, inset). In contrast, dorsal p1 contained rel-atively light label, and only near the ventricular surface. Otherexpression domains include a strip extending diagonally fromrostral p1 into dorsal p2, and heavily labeled domains in ventralrostral prosencephalon and rhombencephalon. In the mutant,the mes domain of Dbx label was expanded rostrally into theformer p1 territory (Fig. 5J), with light label deep and heavylabel superficial (inset), as in the wild-type mes. The loss of p1markers is thus accompanied by the gain of Dbx expression,indicating a shift in the identity of the p1 region to that of mes.
Defects in posterior commissure neuronsIf p1 is shifted to a mes identity, as suggested by the molecularmarkers, how would this transformation of identity affectneuronal development? Our previous study of axonal projec-tion patterns in wild type found two projection patterns incommon to mes and p1: tmesV (tract of the ‘mesencephalic’nucleus of the trigeminal nerve) axons appeared in both p1 andmes on E8.5, projecting ventrally then turning caudally, withcda (circumferential descending axons) axons appearing later,on E10.0, and projecting straight ventrally (Mastick and Easter,1996). Unique to dorsal p1, however, are the dorsal group ofpc neurons, which project dorsally to cross the roofplate ofcaudal p1 on E10.5, thus pioneering the commissure. The pc,and the neurons that project into it, are thus p1-specific, andare therefore markers for p1 identity on the neuronal level.
We first examined the pc in TuJ1-labeled whole mounts. AsFig. 6A,B shows, a dense array of neurons in dorsal mes, p1,and p2 projected their axons ventrally, away from the roof platein both mutant and wild-type embryos on E10.5, demonstrat-ing that neuronal development in general was not grosslydelayed or altered in the mutant. However, the pc wasabnormal; several bundles of axons crossed the dorsal midlinein caudal p1 of wild type, but no bundles were labeled inmutant embryos on E10.5. A few axons, greatly reduced innumber, were present on E11.5 (not shown). In addition, thelabeled somata in the roof plate of rostral p1 were morenumerous in wild type than in mutant.
The lack of commissural axons in p1 could be due either toa blockage of their axons by the mutant roof plate, or to mis-specification or loss of pc neuron cell bodies, resulting in afailure to project dorsally. To identify a blockage, we appliedDiI to E10.5 embryos in dorsal p1, just lateral to the roof plate,where it would be expected to label any blocked pc axons. Inwild type (Fig. 6C), a complex pattern of fibers was labeled,as predicted by the neuron map of dorsal p1. Contralateral pro-jections included fibers projecting across the commissure. Ipsi-laterally, anterogradely labeled axons projected ventrally fromdorsal p1 somata (tmesV turn caudally, and cda project towardthe ventral midline), and retrogradely labeled pc cell bodies inboth dorsal and ventral patches. Mutant embryos (Fig. 6D) hadthe ipsilateral anterogradely labeled axons, but lacked axonscrossing the dorsal midline (consistent with Fig. 6B), andlacked retrogradely labeled dorsal pc somata. The greatmajority of retrogradely labeled somata were located ventral totmesV, indicating that the loss of pc neurons was limited to thedorsal subset. We can thus account for the early loss of the pcby the selective loss of pc axons originating from dorsal p1.The axon projections of the spared ventral pc neurons probablyaccount for the sparse pc that forms on E11.5, and indicate that
dorsally directed axon projections are not generally inhibited.Therefore the effect of the mutant Pax-6 is the specific deficitof dorsal pc axons, likely from a mis-specification of dorsal pcneurons or the failure of these cells to form. This defect is con-sistent with the transformation of dorsal p1 to a mes identitysuggested by the molecular markers.
Defects in tpoc axon pathfindingThe correlation between the course of the tpoc and Pax-6labeling (Fig. 3) suggested that tpoc axon pathfinding might bealtered in the mutant, and this possibility was evaluated bylabeling the tract with DiI on E10.5 (Fig. 7A,B). The initial tightpart of the tract appeared normal in the mutant, and the tractwidened caudal to the optic stalk as expected, but the tract wasabnormal in several respects. A number of axons projected
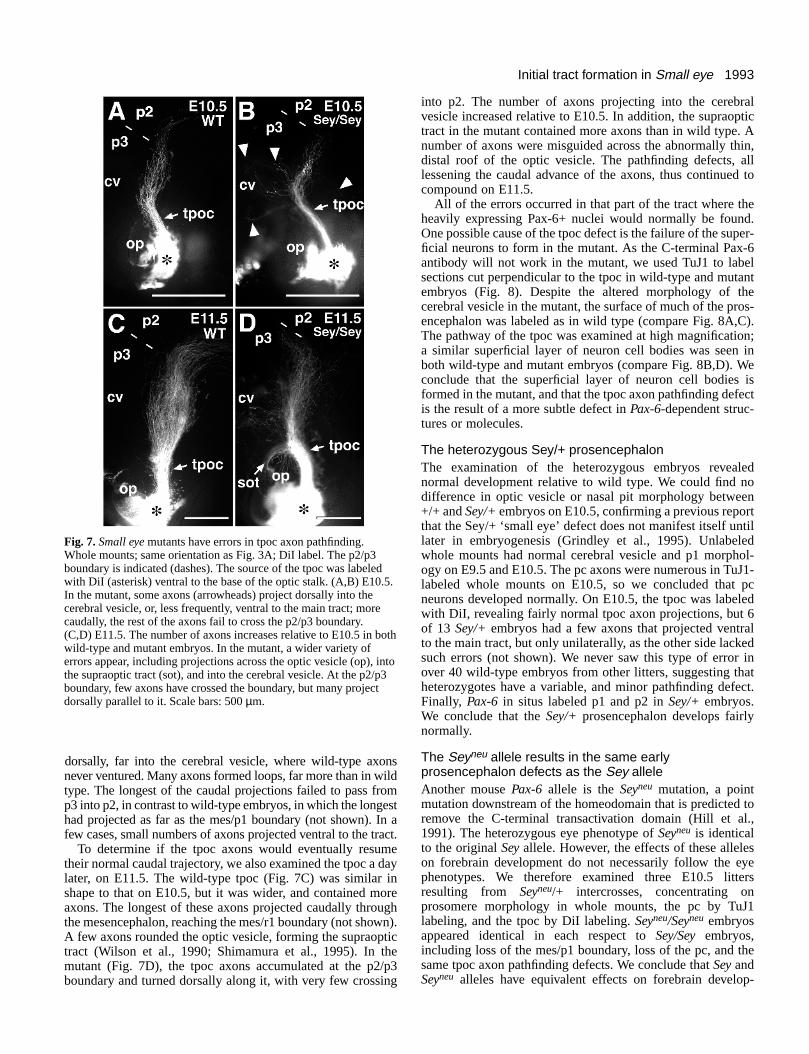
1993Initial tract formation in Small eye
Fig. 7. Small eye mutants have errors in tpoc axon pathfinding.Whole mounts; same orientation as Fig. 3A; DiI label. The p2/p3boundary is indicated (dashes). The source of the tpoc was labeledwith DiI (asterisk) ventral to the base of the optic stalk. (A,B) E10.5.In the mutant, some axons (arrowheads) project dorsally into thecerebral vesicle, or, less frequently, ventral to the main tract; morecaudally, the rest of the axons fail to cross the p2/p3 boundary.(C,D) E11.5. The number of axons increases relative to E10.5 in bothwild-type and mutant embryos. In the mutant, a wider variety oferrors appear, including projections across the optic vesicle (op), intothe supraoptic tract (sot), and into the cerebral vesicle. At the p2/p3boundary, few axons have crossed the boundary, but many projectdorsally parallel to it. Scale bars: 500 µm.
dorsally, far into the cerebral vesicle, where wild-type axonsnever ventured. Many axons formed loops, far more than in wildtype. The longest of the caudal projections failed to pass fromp3 into p2, in contrast to wild-type embryos, in which the longesthad projected as far as the mes/p1 boundary (not shown). In afew cases, small numbers of axons projected ventral to the tract.
To determine if the tpoc axons would eventually resumetheir normal caudal trajectory, we also examined the tpoc a daylater, on E11.5. The wild-type tpoc (Fig. 7C) was similar inshape to that on E10.5, but it was wider, and contained moreaxons. The longest of these axons projected caudally throughthe mesencephalon, reaching the mes/r1 boundary (not shown).A few axons rounded the optic vesicle, forming the supraoptictract (Wilson et al., 1990; Shimamura et al., 1995). In themutant (Fig. 7D), the tpoc axons accumulated at the p2/p3boundary and turned dorsally along it, with very few crossing
into p2. The number of axons projecting into the cerebralvesicle increased relative to E10.5. In addition, the supraoptictract in the mutant contained more axons than in wild type. Anumber of axons were misguided across the abnormally thin,distal roof of the optic vesicle. The pathfinding defects, alllessening the caudal advance of the axons, thus continued tocompound on E11.5.
All of the errors occurred in that part of the tract where theheavily expressing Pax-6+ nuclei would normally be found.One possible cause of the tpoc defect is the failure of the super-ficial neurons to form in the mutant. As the C-terminal Pax-6antibody will not work in the mutant, we used TuJ1 to labelsections cut perpendicular to the tpoc in wild-type and mutantembryos (Fig. 8). Despite the altered morphology of thecerebral vesicle in the mutant, the surface of much of the pros-encephalon was labeled as in wild type (compare Fig. 8A,C).The pathway of the tpoc was examined at high magnification;a similar superficial layer of neuron cell bodies was seen inboth wild-type and mutant embryos (compare Fig. 8B,D). Weconclude that the superficial layer of neuron cell bodies isformed in the mutant, and that the tpoc axon pathfinding defectis the result of a more subtle defect in Pax-6-dependent struc-tures or molecules.
The heterozygous Sey/+ prosencephalonThe examination of the heterozygous embryos revealednormal development relative to wild type. We could find nodifference in optic vesicle or nasal pit morphology between+/+ and Sey/+ embryos on E10.5, confirming a previous reportthat the Sey/+ ‘small eye’ defect does not manifest itself untillater in embryogenesis (Grindley et al., 1995). Unlabeledwhole mounts had normal cerebral vesicle and p1 morphol-ogy on E9.5 and E10.5. The pc axons were numerous in TuJ1-labeled whole mounts on E10.5, so we concluded that pcneurons developed normally. On E10.5, the tpoc was labeledwith DiI, revealing fairly normal tpoc axon projections, but 6of 13 Sey/+ embryos had a few axons that projected ventralto the main tract, but only unilaterally, as the other side lackedsuch errors (not shown). We never saw this type of error inover 40 wild-type embryos from other litters, suggesting thatheterozygotes have a variable, and minor pathfinding defect.Finally, Pax-6 in situs labeled p1 and p2 in Sey/+ embryos.We conclude that the Sey/+ prosencephalon develops fairlynormally.
The Seyneu allele results in the same earlyprosencephalon defects as the Sey alleleAnother mouse Pax-6 allele is the Seyneu mutation, a pointmutation downstream of the homeodomain that is predicted toremove the C-terminal transactivation domain (Hill et al.,1991). The heterozygous eye phenotype of Seyneu is identicalto the original Sey allele. However, the effects of these alleleson forebrain development do not necessarily follow the eyephenotypes. We therefore examined three E10.5 littersresulting from Seyneu/+ intercrosses, concentrating onprosomere morphology in whole mounts, the pc by TuJ1labeling, and the tpoc by DiI labeling. Seyneu/Seyneu embryosappeared identical in each respect to Sey/Sey embryos,including loss of the mes/p1 boundary, loss of the pc, and thesame tpoc axon pathfinding defects. We conclude that Sey andSeyneu alleles have equivalent effects on forebrain develop-

1994 G. S. Mastick and others
Fig. 8. Small eye mutants retain superficial neuron cell bodies in thetpoc pathway. Sections immunolabeled with TuJ1, detected byDAB/peroxidase reaction (brown), and counterstained with toluidineblue. (A) Section through a wild-type embryo, in a planeperpendicular to the tpoc, as in Fig. 3B. Note the layer of labelextending from ventral prosencephalon dorsally through the cerebralvesicle. (B) Detail of boxed area in A, showing a superficial layer ofneuron cell bodies and fibers (similar to the greenimmunofluorescence in Fig. 3G). (C) Section on the same planethrough a mutant embryo. Note the similar layer of neurons, despitethe altered angle of the hemispheric sulcus (hs). (D) Detail of boxedarea in C. Scale bars: A (applies to C), 400 µm; B (applies to D),100 µm.
ment, supporting the conclusion that these mutations result ina similar loss of function.
DISCUSSION
Our results indicate that Sey/Sey mutant embryos have severalearly and specific defects in prosencephalon development. Weattempt to explain the role of Pax-6 in the development of p1,the pc, and the tpoc.
Pax-6 and p1Our demonstration that the mes/p1 boundary is missing inSey/Sey embryos provides the first evidence for genetic regu-lation of prosomere formation. Pax-6 function appears to bespecific for the mes/p1 boundary, because the p1/p2 and p2/p3boundaries appear normal in the mutant. There are severalpossible functions for Pax-6 in boundary formation, dependingon which of three cellular mechanisms we can hypothesize forprosomere formation. One possibility, given that neuromeresare centers of proliferation (Bergquist and Kallen, 1954;Guthrie et al., 1991), is that Pax-6 functions to activate prolif-eration in the center of p1, thus causing the bulge. This can beexcluded because wild-type p1 is not obviously thicker thanthe corresponding region of the mutant brain (Fig. 4E,F), anda large number of neurons are still produced in mutant dorsal
p1 (Fig. 5B). A second possibility is that Pax-6 directs theformation of specialized boundary cells, distinct from eithermes or p1 cells, such as those found between rhombomeres(Heyman et al., 1995). We have yet to find any evidence forsuch cells at prosomere boundaries. We favor a third possibil-ity, that Pax-6 is necessary in p1 to establish the identity of p1cells as different from mes cells, and that this differencesomehow results in the formation of a dorsal constriction alongthe interface of the two populations. The Sey mutation doesappear to alter cell interactions; Sey/Sey and +/+ cellssegregate into separate domains in the eyes of chimeras (Quinnet al., 1996), consistent with an identity conferred by Pax-6activity. In experimental studies of rhombomeres, constrictionsform only at the interface between grafts of even and oddnumbered tissue (e.g. r2 and either r3 and r5), but not betweeneven and even (r2 and r4; Guthrie and Lumsden, 1991). Whatcould be occurring at the interface? It could be an inhibition ofproliferation, as interneuromeric cells are known to divide lessfrequently than those in the neuromeres (Martinez et al., 1992).Alternatively, it could be a limitation of cellular movementacross the interface; cells do not cross the mes/p1 boundary innormal chicks (Figdor and Stern, 1993). We predict that in themutant, such crossings would inappropriately occur.
Although the mutant embryos lacked the mes/p1 boundary,the dorsal prosencephalon had a normal rostral-caudal length,and thus did not appear to contain any obvious deletions at thisstage. In this respect, the Pax-6 mutant differs from previouslyreported mutants with neuromere phenotypes, all of whichcontain substantial early deletions: Wnt-1 (McMahon andBradley, 1990; Thomas and Capecchi, 1990; Mastick et al.,1996), En-1 (Wurst et al., 1994), Otx-2 (Acampora et al.,1995), kreisler (McKay et al., 1994), and Hoxa-1 (Mark et al.,1993). For example, Wnt-1−/− embryos have an initial loss ofregion-specific molecular markers, followed by a subsequentdisappearance of mes and r1 cells (McMahon et al., 1992), ulti-mately resulting in the fusion of p1 to r2, a shortened neuraltube, and a less acute cephalic flexure (Mastick et al., 1996).Sey/Sey mutant embryos lose a morphological boundary, butnot a neuromere, leading to the conclusion that Pax-6 is notnecessary, at least at this early stage, for survival of dorsal p1cells, but rather for their identity.
Defects in p1 identity were apparent from the expressionpatterns of several genes in the mutant. The dorsal p1 domainof three markers was lost, including markers for ventricularcells (Pax-6, Gsh-1) and superficial cells (Lim-1), while theexpression domain of Dbx, normally restricted to the mes sideof the mes-p1 boundary, was expanded rostrally into the p1domain. We conclude that the molecular identity of dorsal p1has been at least partially shifted to that of the mesencephalon.This transformation is reflected on the neuronal level by thedisappearance of the dorsal pc axons, leaving a mes-likepattern of only tmesV and cda projections.
The altered patterns of transcripts in dorsal p1 define a setof Pax-6-dependent homeobox genes involved in regulating thedevelopment of this region. The loss of Pax-6 transcripts fromdorsal p1 and most of p2 indicates regionally restrictedautoregulation, similar to that previously reported in the lensectoderm (Grindley et al., 1995). This autoregulation could bedirect, as Pax-6 can bind to and activate its own promoter intissue culture cells (Plaza et al., 1993). However, the persis-tence of Pax-6 transcription in most regions implies that

1995Initial tract formation in Small eye
autoactivation is regulated in a very domain-specific manner,and that other activator(s) are sufficient to activate the Pax-6promoter outside of p1. Our results also suggest that Gsh-1,Lim-1 and Dbx are genetically downstream of Pax-6. To date,known transcriptional targets of Pax-6 are limited to lens crys-talline genes (reviewed by Cvekl and Piatigorsky, 1996), andpossibly the cell adhesion molecule L1 (Chalepakis et al.,1994). We propose that Pax-6 also regulates a cascade of tran-scriptional regulators, each of which could direct a subset ofthe diverse developmental functions of Pax-6. The timing ofLim-1 and Gsh-1 expression in dorsal p1 are consistent withthese genes being direct regulatory targets of Pax-6, as neitheris expressed until after E9.5 (Fujii et al., 1994; Barnes et al.,1994; Valerius et al., 1995), well after Pax-6 in this region. Dbxis formally repressed by Pax-6. The promoters of these geneswill have to be characterized to test for direct Pax-6 regulation.The function of these genes in p1 development has not yet beeninvestigated, but either Lim-1 or Gsh-1 could be involved inboundary formation or pc neuron differentiation. Gsh-1mutants have postnatal pituitary and hypothalamic defects, butanalysis of early embryos has not been reported (Li et al.,1996). Lim-1 knockouts lack the entire forebrain and midbrainbecause of an early gastrulation defect (Shawlot and Behringeret al., 1995); conditional alleles will be necessary to test Lim-1 function in p1.
Pax-6 and axonsBoth the pc and the tpoc were altered in the mutant, but thechanges were profoundly different in the two structures. Thealteration in the pc resulted from a misspecification or deletionof an identified group of cells while the tpoc was changed byaberrant axonal outgrowth.
The pc defect was traced to a failure to retrogradely labelthe dorsal pc neurons, which suggests that dorsal pc neuronsfail to form or their axons are misdirected in the mutant. Theobservations that ventral pc somata are still present and forma (reduced) pc in the mutant have two important implications.First, the dorsal trajectories of ventral pc neurons indicate thatdorsal-ward guidance cues are still present, arguing against adefect in extrinsic axon pathfinding for dorsal pc neurons.Second, dorsal and ventral pc neurons have different identities,despite the fact that they project their axons together, and theidentity of the dorsal cells depends on Pax-6 expression. Wesuggest that Pax-6 functions in dorsal pc neurons by directingeither their formation or early specification. The pc precursorslikely express Pax-6, as we found that the Pax-6 antibody labelsnearly all p1 nuclei on E9.5. However, on E10.5, when the pcneurons could first be specifically labeled by DiI in theroofplate, very few of them expressed Pax-6. Either Pax-6 isexpressed early in dorsal pc precursors, acting to direct theirbirth or differentiation, then expression is rapidly down-regulated, or pc neuron differentiation is dependent on anextrinsic signal from neighboring Pax-6-expressing cells. Noearly markers for pc neurons are yet available to sort out thesepossibilities.
The tpoc is formed of axons from Pax-6− cells, and theirnumbers are not apparently altered in the mutant; instead, theaxons make errors. Their initial trajectory is normal, and whenthey enter the region where they would normally fan out, theydo so. Thus, the initial tight section of the tract and the initialfanning out are both independent of Pax-6 function, but Pax-
6-dependent navigational cues are necessary for the wide zoneof the tract. We believe that these cues probably promote axonoutgrowth rather than inhibit it, because in the mutant, eventhose tpoc axons that do project in the right direction are shortand form loops, suggesting a less favorable environment thanin wild type. Inhibitory cues cannot be excluded, however, andone possible candidate is the hemispheric sulcus, a sharplyangled fold in wild type, which could mechanically hinderattempts to project dorsally into the cerebral vesicle. In themutant, the sulcus is less prominent, but the effects of thisaltered morphology must be minimal, in that most axonsremain ventral, and do not stream en masse dorsally.
Although we do not know their identity(ies), the Pax-6-dependent guidance cues are probably produced by a layer ofsuperficial neuron cell bodies in this region. They express Pax-6 at high levels, and they are present on E9.5 (Fig. 3F), beforethe axons arrive, consistent with previous observations ofTuJ1+ cells in this region (Easter et al., 1993). Furthermore,the distribution of these cells closely matches the length andwidth of the tract (Fig. 3B). They are not the only cells tocontact the tract, since underlying neuroepithelial cells extendfrom ventricle to pia, and their pial endfeet contact growthcones and wrap axons. Differentiated neurons, in contrast, losetheir ventricular attachment, and their nuclei migrate up to thepia. Thus, the tpoc axons are in contact with the neuroepithelialendfeet and the superficial somata of cells expressing Pax-6.We infer from the spatial coincidence of the tract and the super-ficial cells that they are more important than the endfeet. Thesuperficial cells are neurons, and could express neuron-specificmolecules, e.g. cell adhesion molecules, that the neuroepithe-lial cells do not.
We were able to determine that an apparently normal layerof neuron cell bodies was present in the appropriate region ofthe mutant prosencephalon, indicating that the mutant does notsimply lack these neurons. However, the development of thisregion (p3, p4) is clearly disturbed in the mutant, as indicatedby several lines of evidence. We note here alterations in thepatterns of Lim-1, Gsh-1 and Dbx rostral to the p2/p3 boundaryon E10.5 (Fig. 5). From E12.5 onwards, Stoykova et al. (1996)observed alterations in patterns of Pax-6, Otx-2 and Dlx-1 inp3 and p4, and the failure of corresponding ventral thalamicand hypothalamic nuclei to develop. These results suggest thatdisruption of patterning on any of several molecular or cellularlevels could account for the tpoc axon pathfinding defects.
Pax-6 is only one of a large number of transcription factorsexpressed in regionally restricted patterns in the CNS. Takentogether, this mosaic of partially overlapping gene expressionpatterns potentially provides an abundance of guidance infor-mation for axons to navigate through the brain. Substantialevidence for this hypothesis has accumulated for the homeoboxgene Engrailed (reviewed by Retaux and Harris, 1996). Notonly does a gradient of En expression in the mesencephaloncorrelate with the topographic projection of optic axons, butvarious experimental manipulations of the En gradient alterthese axon projections. The guidance function of En appearsto be mediated by several candidate cell surface molecules,notably Eph ligands that are upregulated by En (Logan et al.,1996). In the prosencephalon, Pax-6 may provide local posi-tional information through similar molecules.
Wilson and others (1993) have suggested that the borders ofexpression domains are important for axon guidance. Although

1996 G. S. Mastick and others
we have emphasized guidance within the Pax-6 domain, we dofind two examples where the borders of Pax-6 expression mayinfluence the tpoc axons. First, the tpoc is closely associatedwith the ventral border of expression, as in zebrafish. (Thissimilarity is complicated by the fact that the mouse tpoc axonsproject dorsal to the border, while the fish axons remain ventralto it (Macdonald et al., 1994), but the dorsal-ventral differencesin the tpoc trajectories of mice and fish have been noted before(Mastick and Easter, 1996), and are poorly understood.) Thesecond example of guidance at a boundary is the failure of tpocaxons to cross the mutant p2/p3 boundary. This result was sur-prising, since we previously observed that axons in Wnt-1−/−mutant embryos could project without errors across a novelp1/r2 boundary (Mastick et al., 1996). One explanation for thefailure in Sey/Sey embryos is the absence of special cells at theboundary which facilitate crossing, like the guidepost cells inthe insect peripheral nervous system (Bentley and O’Connor,1991). Alternatively, the axons may normally acquire a Pax-6-dependent signal from p3, probably from the Pax-6+ neurons,that allows them to project into p2. These sorts of locallyacquired signals can influence the cell surface moleculesexpressed on axons, as shown for commissural axons encoun-tering the midline in the vertebrate spinal cord (Dodd et al.,1988), or in the Drosophila CNS (Bastiani et al., 1987). Wepropose that such locally acquired signals could allow axonsto accurately navigate long distances, reprogramming growthcones to gain responsiveness to the local cues in new regions.
We thank A. LaMantia for providing the Sey/+ founder of ourcolony, K. Backs and P. Gruss for the Pax-6 in situ probe, H. Shengand H. Westphal for the Lim-1 probe, T. Valerius and S. Potter for theGsh-1 probe, F. Ruddle for the Dbx probe, J. Davis and R. Reed forthe Pax-6 antibody, T. Glaser and J. Lauderdale for advice on PCRgenotyping, P. Eckler for expert mouse care, O. Sundin and P.Hitchcock for helpful suggestions. N. Brown and T. Glaser providedlitters of Seyneu embryos, and comments on the manuscript. This workwas supported by a postdoctoral fellowship to G. S. M.(1F32NS09701), a grant from the Office of the Vice President forResearch of the University of Michigan, and a grant from the NIH(NS33337).
REFERENCES
Acampora, D., Mazan, S., Lallemand, Y., Avantaggiato, V., Maury, M.,Simeone, A. and Brulet, P. (1995). Forebrain and midbrain regions aredeleted in Otx2−/− mutants due to a defective anterior neuroectodermspecification during gastrulation. Development 121, 3279-3290.
Barnes, J. D., Crosby, J. L., Jones, M. C., Wright, C. V. E., and Hogan, B. L.M. (1994). Embryonic expression of Lim-1, the mouse homologue ofXenopus Xlim-1, suggests a role in lateral mesoderm differentiation andneurogenesis. Dev. Biol. 161, 168-178.
Bastiani, M. J., Harrelson, A. L., Snow, P. M. and Goodman, C. S. (1987).Expression of fasciclin I and II glycoproteins on subsets of axon pathwaysduring neural development in the grasshopper. Cell 48,745-755.
Bentley, D. and O’Connor, T. P. (1991). Guidance and steering of peripheralgrowth cones in grasshopper embryos. In The Nerve Growth Cone (ed. P. C.Letourneau, S. B. Kater and E. R. Macagno), pp. 265-282. New York: RavenPress.
Bergquist, H. and Kallen, B. (1954). Notes on the early histogenesis andmorphogenesis of the central nervous system in vertebrates. J. Comp. Neurol.100, 627-659.
Bulfone, A., Puelles, L., Porteus, M. H., Frohman, M. A., Martin, G. R. andRubenstein, J. L. R. (1993). Spatially restricted expression of Dlx-1, Dlx-2(Tes-1), Gbx-2, and Wnt-3 in the embryonic day 12. 5 mouse forebrain
defines potential transverse and longitudinal segmental boundaries. J.Neurosci. 13, 3155-3172.
Chalepakis, G., Wijnholds, J., Giese, P., Schachner, M. and Gruss, P.(1994). Characterization of Pax-6 and Hox-1 binding to the promoter regionof the neural cell adhesion molecule L1. DNA Cell Biol. 13, 891-900.
Chedotal, A., Pourquie, O. and Sotelo, C. (1995). Initial tract formation in thebrain of the chick embryo: selective expression of the BEN/SC1/DM-GRASP cell adhesion molecule. Eur. J. Neurosci. 7, 198-212.
Cvekl, A. and Piatigorsky, J. (1996). Lens development and crystalline geneexpression: many roles for Pax-6. BioEssays 18, 621-630.
Davis, J. A. and Reed, R. R. (1996). Role of Olf-1 and Pax-6 transcriptionfactors in neurodevelopment. J. Neurosci. 16, 5082-5094.
Dodd, J., Morton, S. B., Karagogeos, D., Yamamoto, M. and Jessell, T. M.(1988). Spatial regulation of axonal glycoprotein expression on subsets ofembryonic spinal neurons. Neuron 1, 105-116.
Easter, S. S., Jr., Ross, L. S. and Frankfurter, A. (1993). Initial tractformation in the mouse brain. J. Neurosci. 13, 285-299.
Figdor, M. C. and Stern, C. D. (1993). Segmental organization of embryonicdiencephalon. Nature 363, 630-634.
Fujii, T., Pichel, J. G., Taira, M., Toyoma, R., Dawid, I. B. and Westphal, H.(1994). Expression patterns of the murine LIM class homeobox gene lim1 inthe developing brain and excretory system. Dev. Dyn. 199, 73-83.
Glaser, T., Walton, D. S. and Maas, R. L. (1992). Genomic structure,evolutionary conservation and aniridia mutations in the human PAX6 gene.Nat. Genet. 2, 232-239.
Glaser, T., Jepeal, L., Edwards, J. G., Young, S. R., Favor, J. and Maas, R.L. (1994). PAX6 gene dosage effect in a family with congenital cataracts,aniridia, anophthalmia and central nervous system defects. Nat. Genet. 7,463-471.
Goulding, M. D., Chalepakis, G, Deutsch, U., Erselius, J. and Gruss, P.(1991) Pax-3, a novel murine DNA binding protein expressed during earlyneurogenesis. EMBO J. 10, 1135-1147.
Grindley, J. C., Davidson, D. R. and Hill, R. E. (1995). The role of Pax-6 ineye and nasal development. Development 121, 1433-1442.
Guthrie, S. and Lumsden, A. (1991). Formation and regeneration ofrhombomere boundaries in the developing chick hindbrain. Development112, 221-229.
Guthrie, S. C., Butcher, M. and Lumsden, A. (1991). Patterns of cell divisionand interkinetic nuclear migration in the chick embryo hindbrain. J.Neurobiol 22, 742-754.
Halder, G., Callaerts, P. and Gehring, W. J. (1995). Induction of ectopic eyesby targeted expression of the eyeless gene in Drosophila. Science 267, 1788-1792.
Hanson, I. M., Fletcher, J. M., Jordan, T., Brown, A., Taylor, D., Adams, R.J., Punnett, H. H. and van Heyningen, V. (1994). Mutations at the PAX6locus are found in heterogeneous anterior segment malformations includingPeters’ anomaly. Nat. Genet. 6, 168-173.
Heyman, I., Faissner, A. and Lumsden, A. (1995). Cellular and molecularspecializations of rhombomere boundaries. Dev. Dynam. 204, 301-315.
Hill, R. E., Favor, J., Hogan, B. L., Ton, C. C., Saunders, G. F., Hanson, I.M., Prosser, J., Jordan, T., Hastie, N. D. and van Heyningen, V. (1991).Mouse Small eye results from mutations in a paired-like homeobox-containing gene. Nature 354, 522-525.
Hirth, F., Therianos, S., Loop, T., Gehring, W. J., Reichert, H. andFurukubo-Tokunaga, K. (1995). Developmental defects in brainsegmentation caused by mutations of the homeobox genes orthodenticle andempty spiracles in Drosophila. Neuron 15, 769-778.
Hogan, B. L., Horsburgh, G., Cohen, J., Hetherington, C. M., Fisher, G.and Lyon, M. F. (1986). Small eyes (Sey): a homozygous lethal mutation onchromosome 2 which affects the differentiation of both lens and nasalplacodes in the mouse. J. Embryol. Exp. Morphol. 97, 95-110.
Jordan, T., Hanson, I., Zaletayev, D., Hodgson, S., Prosser, J., Seawright,A., Hastie, N. and van Heyningen, V. (1992). The human PAX6 gene ismutated in two patients with aniridia. Nat. Genet. 1, 328-332.
Lee, M. K., Rebhun, L. I. and Frankfurter, A. (1990). Posttranslationalmodification of class III beta-tubulin. Proc. Natl. Acad. Sci. USA 87, 7195-7199.
Li, H., Zeitler, P. S., Valerius, M. T., Small, K. and Potter, S. S. (1996) Gsh-1,an orphan Hox gene, is required for normal pituitary development. EMBO J.15, 714-724.
Logan, C., Wizenmann, A., Drescher, U., Monschau, B., Bonhoeffer, F. andLumsden, A. (1996) Rostral optic tectum acquires caudal characteristicsfollowing ectopic Engrailed expression. Curr. Biol. 6, 1006-1014.
Lu, S., Bogarad, L. D., Murtha, M. T. and Ruddle, F. H. (1992) Expression

1997Initial tract formation in Small eye
pattern of a murine homeobox gene, Dbx, displays extreme spatial restrictionin embryonic forebrain and spinal cord. Proc. Natl. Acad. Sci. USA 89, 8053-8057.
Lumsden, A. and Keynes, R. (1989). Segmental patterns of neuronaldevelopment in the chick hindbrain. Nature 337, 424-428.
Macdonald, R., Xu, Q., Barth, K. A., Mikkola, I., Holder, N., Fjose, A.,Krauss, S. and Wilson, S. W. (1994). Regulatory gene expressionboundaries demarcate sites of neuronal differentiation in the embryoniczebrafish forebrain. Neuron 13, 1039-1053.
Mark, M., Lufkin, T., Vonesch, J. L., Ruberte, E., Olivo, J. C., Dolle, P.,Gorry, P., Lumsden, A. and Chambon, P. (1993). Two rhombomeres arealtered in Hoxa-1 mutant mice. Development 119, 319-338.
Martinez, S., Geijo, E., Sanchez-Vives, M. V., Puelles, L. and Gallego, R.(1992). Reduced junctional permeability at interrhombomeric boundaries.Development 116, 1069-1076.
Mastick, G. S. and Easter, S. S., Jr. (1996). Initial organization of neurons andtracts in the embryonic mouse fore- and midbrain. Dev. Biol. 173, 79-94.
Mastick, G. S., Fan, C. M., Tessier-Lavigne, M., Serbedzija, G. N.,McMahon, A. P. and Easter, S. S., Jr. (1996). Early deletion of neuromeresin Wnt-1−/− mutant mice: evaluation by morphological and molecularmarkers. J. Comp. Neurol. 374, 246-258.
Matsuo, T., Osumi-Yamashita, N., Noji, S., Ohuchi, H., Koyama, E.,Myokai, F., Matsuo, N., Taniguchi, S., Doi, H., Iseki, S. et al. (1993). Amutation in the Pax-6 gene in rat small eye is associated with impairedmigration of midbrain crest cells. Nat. Genet. 3, 299-304.
McConnell, S. K., Ghosh, A. and Shatz, C. J. (1989). Subplate neuronspioneer the first axon pathway from the cerebral cortex. Science 245, 978-982.
McKay, I. J., Muchamore, I., Krumlauf, R., Maden, M., Lumsden, A. andLewis, J. (1994). The kreisler mouse: a hindbrain segmentation mutant thatlacks two rhombomeres. Development 120, 2199-2211.
McMahon, A. P. and Bradley, A. (1990). The Wnt-1 (int-1) proto-oncogene isrequired for development of a large region of the mouse brain. Cell 62,1073-1085.
McMahon, A. P., Joyner, A. L., Bradley, A. and McMahon, J. A. (1992). Themidbrain-hindbrain phenotype of Wnt-1−/Wnt-1− mice results from stepwisedeletion of engrailed-expressing cells by 9. 5 days postcoitum. Cell 69, 581-595.
Moody, S. A., Quigg, M. S. and Frankfurter, A. (1987). Development of theperipheral trigeminal system in the chick revealed by an isotype-specific anti-beta-tubulin monoclonal antibody. J. Comp. Neurol. 279, 567-580.
Parr, B. A., Shea, M. J., Vassileva, G. and McMahon, A. P. (1993). MouseWnt genes exhibit discrete domains of expression in the early embryonicCNS and limb buds. Development 119, 247-261.
Plaza, S., Dozier, C., and Saule, S. (1993) Quail Pax-6 (Pax-QNR) encodes atranscription factor able to bind and transactivate its own promoter. CellGrowth Differ. 4, 1041-1050.
Price, M., Lazzaro, D., Pohl, T., Mattei, M. -G., Ruther, U., Olivo, J. -C.,Duboule, D. and Di Lauro, R. (1992). Regional expression of the homeoboxgene Nkx-2. 2 in the developing mouse forebrain. Neuron 8, 241-255.
Puelles, L. and Rubenstein, J. L. R. (1993). Expression patterns of homeoboxand other putative regulatory genes in the embryonic mouse forebrainsuggest a neuromeric organization. Trends. Neurosci. 16, 472-479.
Quinn, J. C., West, J. D. and Hill, R. E. (1996). Multiple functions for Pax6 inmouse eye and nasal development. Genes. Dev. 10, 435-446.
Quiring, R., Walldorf, U., Kloter, U. and Gehring, W. J. (1994). Homologyof the eyeless gene of Drosophila to the Small eye gene in mice and Aniridiain humans. Science 265, 785-789.
Retaux, S. and Harris, W. A. (1996) Engrailed and retinotectal topography.Trends Neurosci. 19, 542-546.
Roberts, R. C. (1967). Small eyes – a new dominant mutation in the mouse.Genetic Res. Camb. 9, 121-122.
Rubenstein, J. L. and Puelles, L. (1994). Homeobox gene expression duringdevelopment of the vertebrate brain. Curr. Top. Dev. Biol. 29, 1-63.
Schmahl, W., Knoedlseder, M., Favor, J. and Davidson, D. (1993). Defects ofneuronal migration and the pathogenesis of cortical malformations areassociated with Small eye (Sey) in the mouse, a point mutation at the Pax-6-locus. Acta. Neuropathol. (Berl). 86, 126-135.
Shawlot, W. and Behringer, R. R. (1995). Requirement for Lim1 in head-organizer function. Nature 374, 425-430.
Shimamura, K., Hartigan, D. J., Martinez, S., Puelles, L. and Rubenstein,J. L. R. (1995). Longitudinal organization of the anterior neural plate andneural tube. Development 121, 3923-3933.
Stoykova, A. and Gruss, P. (1994). Roles of Pax-genes in developing and adultbrain as suggested by expression patterns. J. Neurosci. 14, 1395-1412.
Stoykova, A., Fritsch, R., Walther, C. and Gruss, P. (1996) Forebrainpatterning defects in Small eye mutant mice. Development 122, 3453-3465.
Sundin, O. H. and Eichele, G. (1990). A homeodomain protein reveals themetameric nature of the developing chick hindbrain. Genes. Dev. 4, 1267-1276.
Thomas, K. R. and Capecchi, M. R. (1990). Targeted disruption of the murineint-1 proto-oncogene resulting in severe abnormalities in midbrain andcerebellar development. Nature 346, 847-850.
Ton, C. C., Miwa, H. and Saunders, G. F. (1992). Small eye (Sey): cloning andcharacterization of the murine homolog of the human aniridia gene.Genomics 13, 251-256.
Valerius, M. T., Li, H., Stock, J. L., Weinstein, M., Kaur, S., Singh, G. andPotter, S. S. (1995) Gsh-1: a novel murine homeobox gene expressed in thecentral nervous system. Dev. Dyn. 203, 337-351.
Walther, C. and Gruss, P. (1991). Pax-6, a murine paired box gene, isexpressed in the developing CNS. Development 113, 1435-1449.
Wilkinson, D. G. (1992). In Situ Hybridization: A Practical Approach. Oxford,UK: Oxford University Press.
Wilson, S. W., Ross, L. S., Parrett, T. and Easter, S. S., Jr. (1990). Thedevelopment of a simple scaffold of axon tracts in the brain of the embryoniczebrafish, Brachydanio rerio. Development 108, 121-145.
Wilson, S. W., Placzek, M. and Furley, A. (1993). Border disputes: doboundaries play a role in growth cone guidance? Trends Neurosci. 16, 316-322.
Wurst, W., Auerbach, A. B. and Joyner, A. L. (1994). Multipledevelopmental defects in Engrailed-1 mutant mice: an early mid-hindbraindeletion and pattern defects in forelimbs and sternum. Development 120,2065-2075.
(Accepted 3 March 1997)





























