Embed Size (px)
Citation preview

Paul A. Schmalzer, Ph.D. Ecological Program Kennedy Space Center [email protected] Northeast Florida Scrub Working Group June 2013

Leaf structures that reduce evapotraspiration
Narrow, linear leaves (e.g., Florida rosemary Ceratiola ericoides)
Waxy leaf coatings (e.g., saw palmetto, Serenoa repens)
Sclerophyllous (thick, leathery leaves) (e.g., sand live oak, Quercus geminata)
Vertical orientation (e.g., scrub oak, Quercus inopina)

Saw Palmetto
Waxy coating on leaves

Florida Rosemary
Narrow, linear leaves

Sand live oak
Sclerophyllous leaves Note: sclerophyllous leaves have also been associated with low-nutrient soils.

Root Structure
Extensive surface roots
Deep roots (some species)
Stomatal Closure
Reduces water loss but also reduces photosynthesis
Physiological adaptations Shed leaves & buds (severe drought)

Root clumps
Clumps of vegetation, roots, and soil harvested from oak scrub/scrubby flatwoods with a tree spade. Note dense root mat and large, woody rhizomes/roots.

ABS (Saha et al. 2008) O18 isotope and excavation
Q. inopina, 20 cm; Ceratiola, 50 cm; Q. chapmanii, Q. geminata 40-200 cm, Serenoa, Sabal, 250 cm
KSC (Hungate et al. 2002) Deuterium (D2O) isotope
Q. geminata 95% from water table, Q. myrtifolia 79% from water table; water table at 1.5-3.0 m

Physiology/Growth Stomatal conductance, pre-dawn water potential
reduced (Saha et al. 2008).
Net Ecosystem CO2 Exchange (NEE) & Leaf Net Photosynthetic Rate (PN) depressed (Li et al. 2007).
Biomass increment reduced. Q. myrtifolia more sensitive than Q. geminata (Dijkstra et al. 2002, Seiler et al. 2009).
Damage/Mortality Leaf shedding, bud loss; little mortality (Saha et al.
2008).

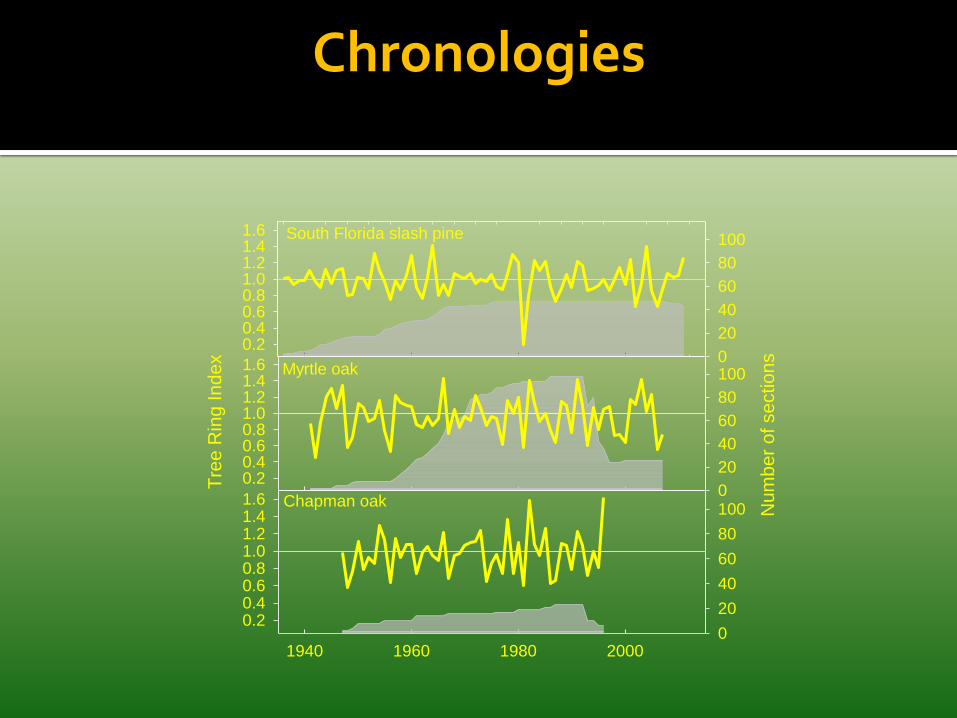
Applied to sections of myrtle oak and Chapman oak collected from long-unburned scrub and cores of South Florida slash pine.
• Created standardized chronologies for each species.
• Correlated ring-width indices with regional precipitation and temperature
• Foster et al. 2013 manuscripts

Myrtle Oak Section 1959 - 1995
1993 1987 1981 1967
Chronologies
Num
ber
of
se
ctio
ns0
20
40
60
80
100
Tre
e R
ing I
nd
ex
0.20.40.60.81.01.21.41.6
0
20
40
60
80
100
0.20.40.60.81.01.21.41.6
South Florida slash pine
Myrtle oak
1940 1960 1980 20000
20
40
60
80
100
0.20.40.60.81.01.21.41.6 Chapman oak
Corr
ela
tion c
oeffic
ient
-0.2
0.0
0.2
0.4
0.6
pJ
pA
pS
pO
pN
pD
Jan
Feb
Mar
Apr
May
Jun
Jul
Aug
Sep
Oct
Nov
Dec
pJA
SpO
ND
JF
MA
MJ
JA
SO
ND
-0.2
0.0
0.2
0.4
0.6
Myrtle oak Chapman oak
Slash pinepJ
pA
pS
pO
pN
pD
Jan
Feb
Mar
Apr
May
Jun
Jul
Aug
Sep
Oct
Nov
Dec
pJA
SpO
ND
JF
MA
MJ
JA
SO
ND
Slash pine earlywood
pJ
pA
pS
pO
pN
pD
Jan
Feb
Mar
Apr
May
Jun
Jul
Aug
Sep
Oct
Nov
Dec
pJA
SpO
ND
JF
MA
MJ
JA
SO
ND
Slash pine latewood

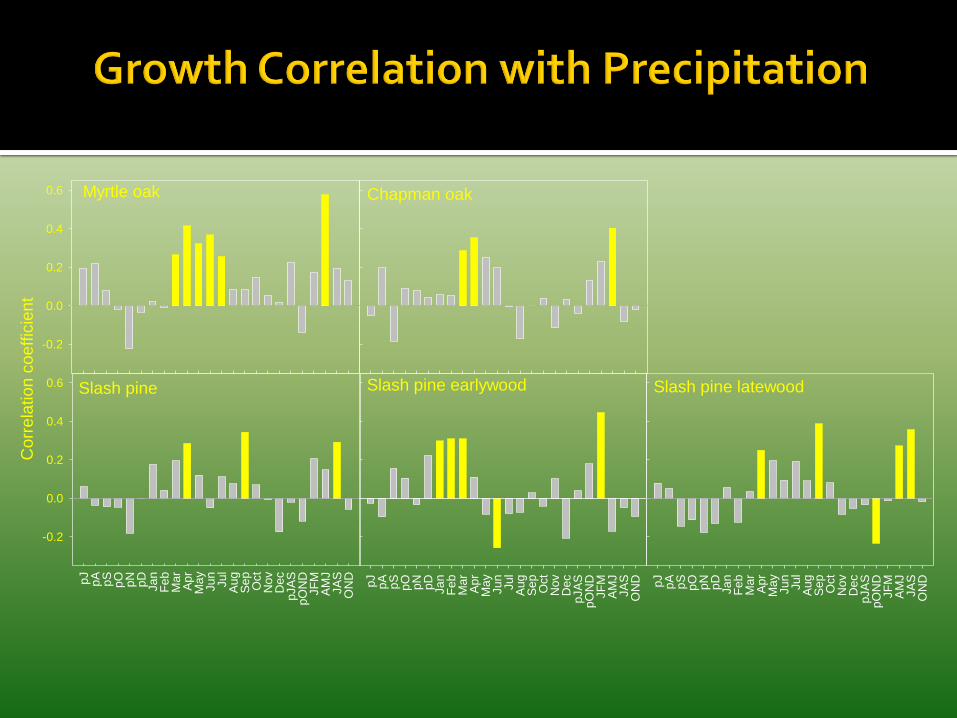
Species responded to differing climatic variables Myrtle oak and Chapman oak responded similarly
▪ Growth was primarily limited by spring precipitation
▪ Spring drought decreased growth
▪ Extreme seasonal precipitation did not influence growth
South Florida Slash pine ▪ Earlywood growth
▪ Primarily limited by winter precipitation
▪ Growth decreased the year following a spring or annual drought
▪ Latewood and total radial growth ▪ Primarily limited by April and September precipitation
▪ Growth decreased by annual and spring drought
▪ Growth increased during years with high precipitation during July – September, usually associated with tropical activity
Species Responses

Ecosystem level measures of photosynthesis, respiration. Scrub & flatwoods sites on KSC.
Scrub maintains evapotranspiration (Et) when surface soils are dry (Bracho et al. 2008).
NEE maintained under short-term dry conditions (Powell et al. 2006).
Deep rooting, access to water table

Seed only
Sand pine (Pinus clausa), killed by fire, serotinuous (+/-) cones, release seeds post-fire, establish in mineral soil (Peninsular Florida)
Florida rosemary, killed by fire, seed bank in soil, released from inhibition with death of parent plant (Peninsular Florida)
Some herbs, particularly in rosemary balds ▪ Menges & Kohfeldt 1995, Maguire & Menges 2011

Sand Pine burning
Killed by fire, seeds released from serotinous cones, may establish in mineral soil

Sprouting
Oaks, saw palmetto, scrub palmetto, scrub hickory, Lyonia spp., Vaccinium spp., many grasses, sedges, and forbs
Clonal Spread
Oaks, saw palmetto, Lyonia spp., Vaccinium spp.
Sprouting + Seed Recruitment
Wiregrass (Aristida stricta), other grasses, palmettos, prickly pear (Opuntia)

Sprouting response
Scrub transect pre- and post-burn Permanent transect, MINWR/KSC, pre-burn, post-burn, 6 months postburn, and 5 years postburn Dominant species: Myrtle oak, saw palmetto,shiny lyonia, Chapman oak (Schmalzer and Hinkle 1992, Schmalzer 2003)

Clonal spread of saw palmetto
Saw palmettos planted in former citrus grove on scrub soil on MINWR/KSC in 1993. Photos are all from 2009. Palmettos show variable growth and clonal spread Foster and Schmalzer (2012) Saw palmetto clones may reach great age (Takahashi et al. 2011)

Most (2/3 or more) of the biomass of scrub vegetation is below-ground (roots & rhizomes) and thus protected from fire.
Archbold (Johnson et al. 1986, Saha et al. 2010)
Ocala (Guerin 1993)
KSC/MINWR (Day et al. 2013)

Survive and Seed
Longleaf pine (Pinus palustris), South Florida slash pine (Pinus elliottii var. densa). Survival depends on fire intensity, season, amount of duff. Exposed mineral soil favors seedling establishment.

Seedling establishment
Difficult in intact scrub, dense root systems, drought, sandy soils that don’t retain water.
In rosemary scrub (and maybe elsewhere) biological soil crusts composed of algae, cynobacteria, bacteria, and fungi) are important (Hawkes 2003).

Rotary cutters, mulching machines, roller-choppers, shearing blades, and other such equipment have been used to aid restoration of long-unburned scrub.
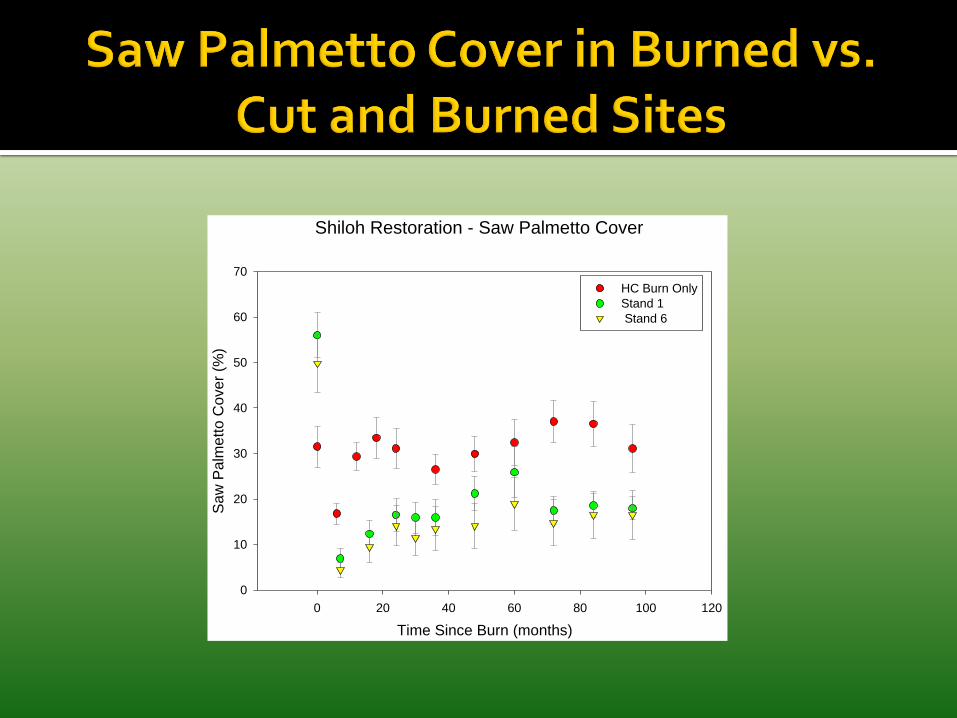
Oaks and other woody shrubs respond similarly to cutting, burning, and cutting plus burning.
Cover of saw palmetto is nearly always reduced by mechanical treatments and the reductions are long-lasting.
Why?

Mechanical treatments
Brown tree-cutter K-G Blade V-blade Roller-chopper Schmalzer & Boyle 1998 Schmalzer & Adrian 2001 Schmalzer et al. 2003
Shiloh Restoration - Saw Palmetto Cover
Time Since Burn (months)
0 20 40 60 80 100 120
Sa
w P
alm
ett
o C
ove
r (%
)
0
10
20
30
40
50
60
70
HC Burn Only
Stand 1
Stand 6
Happy Creek Restoration - Sand live and Myrtle oaks
Time Since Burn (months)
0 20 40 60 80 100 120
Cover
(%)
0
20
40
60
80
Q. geminata Stand 3
Q. geminata Stand 5
Q. myrtifolia Stand 3
Q. myrtifolia Stand 5
Happy Creek Burn - Sand live and Myrtle oaks
Time Since Burn (months)
0 20 40 60 80 100 120 140
Cover
(%)
0
10
20
30
40
50
Quercus geminata
Quercus myrtifolia

Saw palmetto
Rhizome with apical meristem usually aboveground. Protected from fire but vulnerable to mechanical damage. Sometimes sprout epicormically but not as robust.
Scrub oaks
Many dormant buds at base and on underground stems. Sprout when top-killed by fire or cutting.

Long-unburned oak scrub lacks openings important to many scrub animals & plants.
Sprout regeneration after burning or cutting & burning is generally dense.
Sprouting depends on underground rhizomes & roots.
Piling heavy stems and burning should kill belowground stems and delay regeneration.

Some large stems of oaks were piled for burning (1992).
Piles burned longer and probably hotter than fires in standing scrub or lighter fuels and produced ash beds.
Most of the roots and rhizomes of scrub shrubs killed, and the recovery pattern differed.
Cover > 0.5 m in these openings was a mix of shrubs and herbs with relatively low cover of oaks and saw palmetto. Cover < 0.5 m was predominately herbs and bare ground.
Openings gradually closed from canopy spread above, some clonal spread in, and little seed establishment.


Transect through burned pile
Shiloh transect in long-unburned (ca. 50 yr) scrub Pretreatment Cut & piled Burned 7 months postburn 15 months postburn 5 years postburn
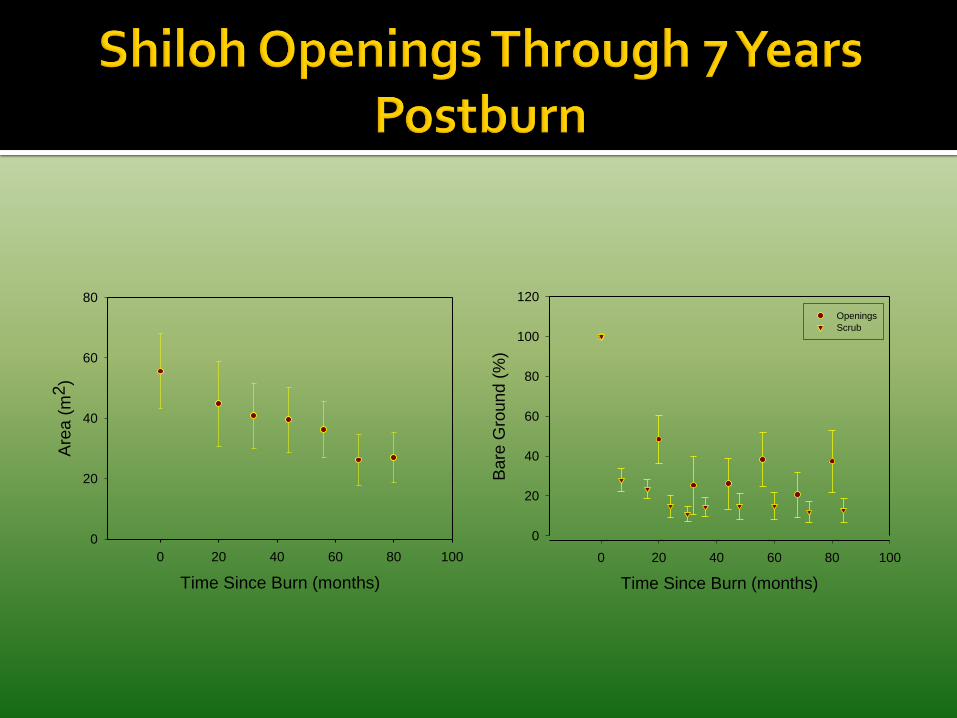
Time Since Burn (months)
0 20 40 60 80 100
Are
a (
m2)
0
20
40
60
80
Time Since Burn (months)
0 20 40 60 80 100
Bare
Gro
und (
%)
0
20
40
60
80
100
120
Openings
Scrub

10-acre planting in former citrus grove on scrub soil on KSC/MINWR beginning in 1992.
Cleared, herbicide treatement. Scrub oaks, Q. geminata, Q. myrtifolia, Q.
chapmanii (1-yr tublings) planted in 1992 with additional planting in 1993.
Saw palmetto, slash pine, Vaccinium myrsinites, Lyonia ferruginea (1-gal pots) planted in 1993.

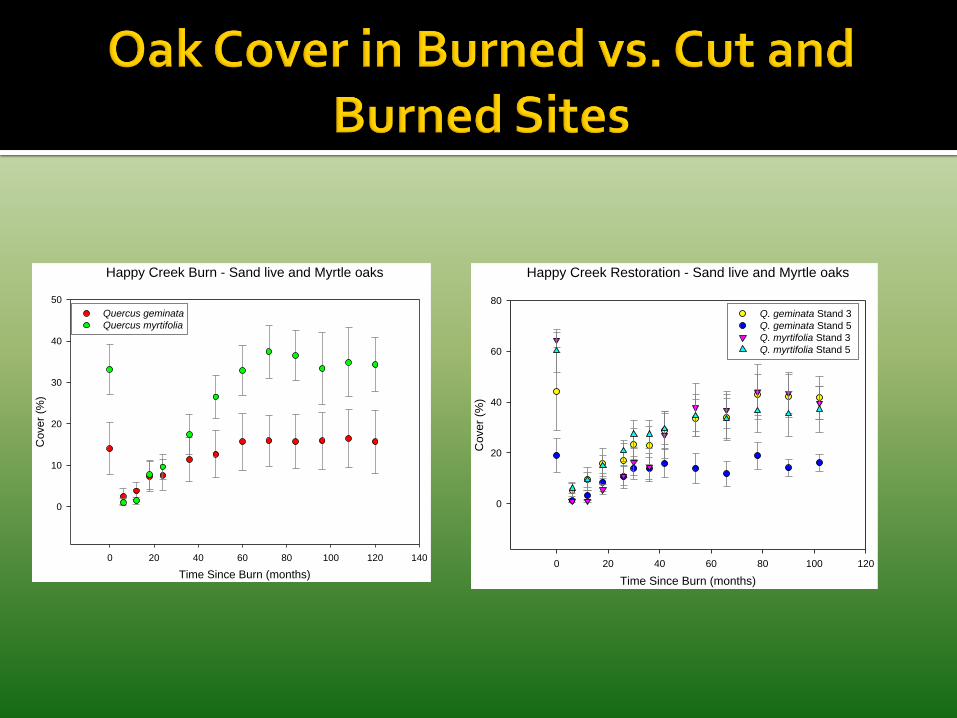
Scrub oaks had differing patterns of survival and growth. Sand live oak had higher survival and more rapid growth than myrtle or Chapman oaks. However, sand live oak grew in height rather than spread clonally.
Saw palmetto survived well initially but survival was reduced by feral pigs. Growth and clonal spread was influenced by exotic grasses.
Slash pine had high survival and rapid growth.

Sand live oak
August 1992 April 1994 April 1995 February 2001 April 2003 (5.9 m height, not shown)

Permanent transect
August 1992 September 1993 September 1997 September 2003 See Schmalzer et al. (2002)

Reestablishing scrub is difficult. Controlling exotics (esp. grasses) is critical. Planted scrub species differ in survival and
growth. Top-killing (fire/cutting) may be necessary to
encourage clonal spread in sand live oak.

First burned in 2000. Some sand live oaks were top-killed. That appeared to encourage clonal spread, but many were too large and survived fire.
Large oaks cut in Nov. 2010 and burned in 2011. Monitoring continuing.

Scrub plants have a variety of adaptations to survive periodic droughts. Mortality from drought is uncommon.
Growth is sensitive to drought, particularly spring drought.
Scrub species have several patterns of regeneration/reproduction after fire, but sprouting is particularly important.
Belowground biomass is critical in scrub recovery after disturbance.

Scrub species differ in response to mechanical disturbance related to position of meristems.
Some reduction in sprouting density through reduced belowground biomass may be necessary to reestablish scrub openings.
Scrub species can survive and grow when planted in former agricultural sites on scrub soils. Development of “normal” scrub structure may be slow, limited by exotic species, and require additional management.

Bracho, R. T.L. Powell, S. Dore, J. Li, C.R. Hinkle, and B.G. Drake. 2008. Environmental and biological controls on water and
energy exchange in Florida scrub oak and pine flatwoods ecosystems. Journal of Geophysical Research 113, G02004, doi:10.1029/2007JG000469.
Day, F.P., R.E. Schroeder, D.B. Stover, A.L.P. Brown, J.R. Butnor, J. Dilustro, B.A. Hungate, P. Dijkstra, B.D. Duval, T.J. Seiler, B.G. Drake, and C.R. Hinkle. 2013. The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem. New Phytologist doi:10.1111/nhp.12246.
Dijkstra, P., G.J. Hymus, D. Colavito, D. Vieglais, C. Cundari, D.P. Johnson, B.A. Hungate, C.R. Hinkle, and B.G. Drake. 2002. Elevated atmospheric CO2 stimulates shoot growth in a Florida scrub oak ecosystem. Global Change Biology 8:90-103.
Foster, T.E. and P.A. Schmalzer. 2012. Growth of Serenoa repens planted in a former agricultural site. Southeastern Naturalist 11(2): 331-336.
Guerin, D.N. 1993. Oak dome clonal structure and fire ecology in a Florida longleaf pine dominated community. Bulletin of the Torrey Botanical Club 120:107-114.
Hawkes, C.V. 2004. Effects of biological soil crusts on seed germination of four endangered herbs in a xeric Florida shrubland during drought. Plant Ecology 170:121-134.
Hungate, BA, M. Reichstein, P. Dijkstra, D. Johnson, G. Hymus, J.D Tenhunen, and B.G. Drake. 2002. Evapotranspiration and soil water content in a scrub-oak woodland under carbon dioxide enrichment. Global Change Biology 8:289-298.
Johnson, A.F., W.G. Abrahamson, and K.D. McCrea. 1986. Comparison of biomass recovery after fire of a seeder (Ceratiola ericoides) and a sprouter (Quercus inopina) species from south-central Florida. American Midland Naturalist 116:423-428.
Li, J.H., D.P. Johnson, P. Dijkstra, B.A. Hungate, C.R. Hinkle, and B.G. Drake. 2007. Elevated CO2 mitigates the adverse effects of drought on daytime net ecosystem CO2 exchange and photosynthesis in a Florida scrub-oak ecosystem. Photosynthetica 45: 51-58.
Maguire, A.J. and E.S. Menges. 2011. Post-fire growth strategies of resprouting Florida scrub vegetation. Fire Ecology 7(3):12-25. doi: 10.4996/fireecology.0703012.
Menges, E.S. and N. Kohfeldt. 1995. Life history strategies of Florida scrub plants in relation to fire. Bulletin of the Torrey Botanical Club 122:282-297.

Powell T. L., R. Bracho, J. Li, S. Dore, C.R. Hinkle, and B.G. Drake. 2006. Environmental controls over net ecosystem carbon exchange of scrub oak in central Florida. Agricultural and Forest Meteorology 141:19-34.
Saha, S., T.M. Strazzisar, E.S. Menges, P. Ellsworth, and L. Sternberg. 2008. Linking the patterns in soil moisture to leaf water potential, stomatal conductance, growth, and mortality of dominant shrubs in the Florida scrub ecosystem. Plant and Soil 313:113-127. doi10.1007/s11104-008-9684-3.
Saha, S. A. Catenazzi, and E.S. Menges. 2010. Does time since fire explain plant biomass allocation in the Florida, USA, scrub ecosystem. Fire Ecology 6(2): 13-25. doi: 10.4996/fireecology.0602013.
Schmalzer, P.A. 2003. Growth and recovery of oak-saw palmetto scrub through ten years after fire. Natural Areas Journal 23:5-13.
Schmalzer, P. A. and F. W. Adrian. 2001. Scrub restoration on Kennedy Space Center/Merritt Island National Wildlife Refuge, 1992-2000. Pp. 17-20 in D. Zattau. (ed.). Proceedings of the Florida Scrub Symposium 2001. U.S. Fish and Wildlife Service. Jacksonville, Florida. 63 pp.
Schmalzer, P. A., and S. R. Boyle. 1998. Restoring long-unburned oak-saw palmetto scrub requires mechanical cutting and prescribed burning (Florida). Restoration & Management Notes 16(1):96-97.
Schmalzer, P.A. and C.R. Hinkle. 1992. Recovery of oak-saw palmetto scrub after fire. Castanea 53:158-173. Schmalzer, P.A. T. E. Foster, and F.W. Adrian. 2003. Responses of long-unburned scrub on the Merritt Island/Cape
Canaveral barrier island complex to cutting and burning. In: Proceedings of the Second International Wildland Fire Ecology and Fire Management Congress, American Meteorological Society, Published on CDROM and at http://www.ametsoc.org.
Schmalzer, P.A., S.R. Turek, T.E. Foster, C.A. Dunlevy, and F.W. Adrian. 2002. Reestablishing Florida scrub in a former agricultural site: Survival and growth of planted species and changes in community composition. Castanea 67:146-160.
Seiler, T.J., D.P. Rasse, J.-H.Li, P. Dijkstra, H.P. Anderson, D.P. Johnson, T.L. Powell, B.A. Hungate, C.R. Hinkle, and B.G. Drake. 2009. Disturbance, rainfall, and contrasting species responses mediated aboveground biomass response to 11 years of CO2 enrichment in a Florida scrub-oak ecosystem. Global Change Biology 15:356-367. doi: 10.1111/j.1365-2486.2008.01740.x
Takahashi, M.K., L.M. Horner, T. Kubota, N.A. Keller, and W.G. Abrahamson. 2011. Extensive clonal spread and extreme longevity in saw palmetto, a foundational clonal plant. Molecular Ecology doi: 10.1111/j.1365-294X.2011.05212.x.












