Embed Size (px)
Citation preview

i
Pattern of Host Seeking and Oviposition Preferences of Malaria
Vectors in Kilifi County, Coastal Kenya
Abdullah Said Hemed
ADEHS (Vector Control)
A thesis submitted to the School of Biological Sciences of the University of Nairobi in partial
fulfillment of the requirements for the award of the degree of Master of Science in Applied
Parasitology
June 2013

ii
DECLARATION
I, Abdullah Said Hemed, declare that this thesis is my original work and has not been
presented for examination in any other University
Candidate
Signed …………………………………….. Date ……………..
Abdullah Said Hemed
Supervisors
This thesis has been submitted with our approval as supervisors
Signed........................................................... Date .......................
Wolfgang Richard Mukabana
(Associate Professor, School of Biological Sciences, University of Nairobi, Nairobi Kenya)
Signed............................................................. Date .......................
Prof. Charles M. Mbogo
(Chief Research Scientist, Vector Biology Department, KEMRI Centre for Geographic
Medicine Research - Coast, Kenya)

iii
ACKNOWLEDGEMENTS
This work was only feasible with support and contributions of various people and institutions
in different ways. I would like to express my appreciation to the management and staff of the
University of Nairobi for giving me admission and academic support. Further, I am deeply
grateful to my supervisors, Prof. Wolfgang Mukabana of the University of Nairobi, I am also
thankful to Prof. Charles Mbogo, Dr. Simon Muriu and Dr. Joseph Mwangangi all of the
Kenya Medical Research Institute (KEMRI), for their tireless guidance in shaping this work.
I am greatly indebted to Mr. Joseph Nzovu and Mr. Festus Yaah for their support and
instructions in laboratory work. I can’t forget the residents of Jaribuni village for their
priceless contribution in making this work possible in those homes, houses and farms. I am
indebted to field workers Mr. Arnold Mramba and Japhet Mwafondo who worked hard with
me in the field to fulfill my goals. I would like to express my thanks to Mrs. Rosemary
Wamae for helping in data compilation, Miss. Benyl Ondeto for technical advice on
computer applications and Mr. Christopher Nyundo who helped me to develop a map of
habitats in the study area. Special thanks to Ifakara Health Institute (IHI) for fully funding my
MSc studies. In particular I thank Dr. Gerry Francis Killeen for his efforts to nominate my
name for the scholarship. My family deserves the very special appreciations for their patience
during the whole period of my stay away from them during this work. I acknowledge the
Academy of Sciences for the Developing World, for supply of CDC backpack motorized
aspirator, used successfully in this study to collect indoor resting mosquitoes. Sincerely, I
express my heartfelt gratitude to the KEMRI/WellcomeTrust, Research Programme, Kilifi
and staff for hosting me and funding this study.

iv
TABLE OF CONTENTS
ITEM PAGE
TITLE ..................................................................................................................................... i
DECLARATION ................................................................................................................... ii
ACKNOWLEDGEMENTS .................................................................................................. iii
TABLE OF CONTENTS ...................................................................................................... iv
LIST OF FIGURES ............................................................................................................. vii
LIST OF TABLES .............................................................................................................. viii
ACCRONYMS AND ABREVIATIONS .............................................................................. ix
ABSTRACT ......................................................................................................................... xi
1 CHAPTER ONE: INTRODUCTION AND LITERATURE REVIEW ............................ 1
1.1 Introduction ............................................................................................................ 1
1.2 Literature Review ................................................................................................... 3
1.2.1 Biology and Epidemiology of Malaria ............................................................... 3
1.2.2 Malaria vectors (Afro-tropical species) ............................................................. 5
1.2.3 Life cycle of mosquitoes ................................................................................... 6
1.2.4 Medical importance of mosquito ....................................................................... 7
1.2.5 Oviposition and spatial distribution of habitats .................................................. 7
1.2.6 Mosquito habitat productivity ........................................................................... 7

v
1.2.7 Mosquito host seeking ...................................................................................... 8
1.2.8 Anopheline mosquito foraging behavior ............................................................ 9
1.2.9 Malaria vector control ..................................................................................... 11
1.3 Problem statement ................................................................................................. 12
1.4 Justification and significance of the research ......................................................... 13
1.5 Hypothesis ............................................................................................................ 14
1.6 Objectives ............................................................................................................. 14
1.6.1 General objective ............................................................................................ 14
1.6.2 Specific objectives .......................................................................................... 14
2 CHAPTER TWO: MATERIAL AND METHODS ....................................................... 15
2.1 Study area ............................................................................................................. 15
2.2 Experimental design .............................................................................................. 17
2.2.1 Anopheles habitat productivity and diversity ................................................... 17
2.3 Ecological factors influencing Anopheles productivity, and emerging mosquito
fitness. ..................................................................................................................... 18
2.3.1 Artificial habitats ............................................................................................ 18
2.3.2 Emergence cages ............................................................................................ 19
2.3.3 Spatial connectivity of Anopheles mosquitoes to pre-adult aquatic habitats ..... 20
2.4 Data analysis ......................................................................................................... 25

vi
2.5 Ethical consideration ............................................................................................. 25
3 CHAPTER THREE: RESULTS .................................................................................... 26
3.1 Habitat productivity and diversity ......................................................................... 26
3.2 Larval productivity................................................................................................ 26
3.3 Seasonal variation in larval habitat productivity .................................................... 28
3.4 Adult productivity ................................................................................................. 30
3.5 Influence of physicochemical factors on habitat productivity ................................ 32
3.5.1 Conductivity ................................................................................................... 32
3.5.2 Dissolved Oxygen ........................................................................................... 32
3.5.3 Salinity ........................................................................................................... 32
3.6 Body size variation in wild Anopheles mosquito population .................................. 34
3.7 Correlation of habitat productivity and distance from human habitation ................ 36
3.8 Effects of mosquito larval habitat distribution on indoor resting densities ............. 38
3.9 ELISA for sporozoite rate and blood meal analysis ............................................... 42
4 CHAPTER FOUR: DISCUSSION, CONCLUSION AND RECOMMENDATIONS .... 45
4.1 Discussion ............................................................................................................ 45
4.2 Conclusion ............................................................................................................ 52
4.3 Recommendations ................................................................................................. 52
REFERENCES .................................................................................................................... 53

vii
LIST OF FIGURES
ITEM………………… …………………………………………………………….….. PAGE
Figure 1: Malaria control cycle between the human host, parasite and vector mosquito ......... 4
Figure 2: Distribution of Mosquito breeding habitat and houses in Jaribuni village Kilifi .... 16
Figure 3: Artificial habitat for monitoring mosquito oviposition preferences ....................... 19
Figure 4: Emergent cage placed over natural habitat to monitor productivity ....................... 20
Figure 5: A characteristic house in Jaribuni village .............................................................. 24

viii
LIST OF TABLES
ITEM…….………………………………………………………………… ………….. PAGE
Table 1: Mosquito larval densities per dip per habitat sampled ............................................ 27
Table 2: Temporal variation of Anopheles larval density per dip per habitat ........................ 29
Table 3: Temporal variation of habitat productivity per M2 of adult mosquito counts .......... 31
Table 4: Regression of adult mosquito productivity against physicochemical variables ....... 33
Table 5: The mean wing length variations amongst the wild Anopheles mosquitoes ............ 35
Table 6 Temporal mosquito larval density recorded from artificial breeding habitats ........... 37
Table 7: Temporal variation of indoor resting mosquitoes collected .................................... 40
Table 8: Correlation of indoor resting Anopheles mosquito densities ................................... 41
Table 9: Plasmodium falciparum infection rate for indoor collected female Anopheles ........ 43
Table 10: Blood meal sources for Anopheles species collected indoors ................................ 44

ix
ACCRONYMS AND ABREVIATIONS
ASDW Academy of sciences for the developing world
ACT Artemisinin-based combination therapy
ANOVA Analysis of variancea
CDC Centres for disease control and Prevention
Co2 Carbon dioxide
CS Circumsporozoites
DDT Dichlorodiphenyltrichloroethene
DEET Diethyl ether
DO Dissolved oxygen
GMP Gates Malaria Partnership
GPS Geographical Positioning System
GBI Goat Blood Index
HBI ` Human Blood Index
ID Identification number
IGR Insect Growth Regulator
IHI Ifakara Health Institute
IPTp Intermitent Preventive Treatment for pregnants
IRS Indoor residual spray

x
ITN Insecticide-treated mosquito net
IVM Intergrated vector management
KEMRI Kenya Medical Research Institute
LLIN Long Lasting Insecticide Net
M2 Meter square
MAbs Monoclonal antibodies
MoPHS Ministry of Public Health and Sanitation
PBS Phosphate buffered saline
pH Hydrogen-ion exponent
RBM Roll Back Malaria partnership
s.l. sensu lato
s.s. sensu stricto
UoN University of Nairobi
USAID United States Agency for International development
WHO World Health Organisation

xi
ABSTRACT
Most malaria vectors alternate between their vertebrate hosts and stagnant water bodies for
blood meal and oviposition sites respectively. These two resources are obligatory
requirements for completion of the mosquito gonotrophic cycle. Knowledge of spatial
distribution and factors influencing habitat productivity of Anopheles mosquitoes is
mandatory in planning their control. The current study was carried out with the aim of
estimating the host seeking and oviposition patterns of malaria vector mosquitoes.
Mechanized aspiration for indoor resting mosquito collection and larval habitat
characterization was done to evaluate the productivity and fitness of emerging adult
mosquitoes at Jaribuni village in Kilifi at the coast of Kenya, from December 2010 to May
2011. Emergence cages measuring 0.5x0.5x0.5 meters were placed on natural habitats to
monitor emerging adults. Similarly, 5 replicates of artificial aquatic habitats were placed
between 0-100m at interval of 25m from selected houses to monitor Anopheles oviposition
preferences in distance. All larval habitats were monitored longitudinally for mosquito
immature stages and physicochemical characteristics. ELISA technique was used to analyze
parasite rate and blood meal source from malaria vectors. A total of 454 sampling visits were
made in five types of aquatic habitats, in which 71.79% the habitats found with Anopheles
larvae. The highest larval productivity was recorded in pools, river and ditches although the
river was the most stable habitat throughout the year. During the 6-month (December 2010-
May 2011) sampling period, the highest proportion of mosquitoes collected indoors were
anophelines 57% (296/519). An. funestus was the predominant species comprising of 97.1%
of all Anopheles collected indoors. Compared to the other Anopheles species, An. funestus
had higher indoor densities, a higher human blood index (0.90) and a higher sporozoite rate
(5.03%). In conclusion, Anopheline breeding sites were diverse ranging from the river
system, stream pools, ditches, trenches and abandoned water storage tanks. Nevertheless, the

xii
river was the most preferred breeding site with abandoned water storage tanks being the least
preferred. Generally, salinity, dissolved oxygen, conductivity and temperature were found to
have significant influence on habitat productivity. Furthermore, adult anophelines preferred
to feed close to their breeding sites.

1
1 CHAPTER ONE: INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction
Malaria is increasingly becoming a complex public health problem, especially in developing
countries. Several efforts have been made by the World Health Organization (WHO) and its
partners to combat the disease in particular through the Roll Back Malaria (RBM) initiative
(WHO, 2008). One of the main issues of concern with malaria is lack of clear understanding
of patterns of malaria vectors in host seeking and oviposition which is the basis for vector
management.
Malaria is estimated to cause 350-500 million clinical cases annually with 90% of all cases
reported from Africa (RBM, 2003). The risk of death from malaria is considerably higher in
Africa than other parts of the world. It is estimated around one million (over 90%) deaths due
to malaria occur in Africa South of the Sahara. Majority of deaths occur among children aged
five years and below (WHO, 2006) while each year approximately 25% of pregnant women
get infected with malaria during pregnancy (WHO, 2004). Malaria constitutes 30-50% of
outpatient consultations and 20-30% inpatients in Africa (Uneke, 2009). It has been estimated
to cause economic burden of 12 billion US dollars per year and slows economic growth by
approximately 1.3% (Rugemalila, et al 2006).
In Kenya, it is estimated that about 70% of the population lives in areas where malaria is the
leading cause of morbidity and mortality, (USAID, 2010). In Kenya, recent evidence indicates
that malaria accounts for 30% of all outpatients and 19% of inpatients, with 20% deaths of
children aged five years and below (Kenya Ministry of Public Health & Sanitation, 2007).
Every year, about six thousand pregnant women suffer from malaria-associated anemia and as
a result four thousand babies are born with low birth weight (MoPHS, 2007). The highest
malaria transmission intensity in the country occurs in the western and coastal regions.

2
Mbogo et al., (2003) reported malaria prevalence of about 60% among the school children
along the Kenya coast suggesting high transmission intensity in late 1990’s.
Malaria is an acute infectious disease caused by parasitic protozoa of the genus Plasmodium
and is spread by the arthropod vector the female Anopheles mosquito. This association
between malaria and mosquitoes was suspected several years ago, during early civilization. It
is said in history that the Queen of Egypt, Cleopatra, slept under a mosquito net and in 11th
century the drainage of swamps was practiced by ancient Romans as among efforts to prevent
malaria. In 1897 Ronald Ross proved the role of mosquitoes in malaria transmission when he
demonstrated oocysts in the mosquito gut (Kakkilaya, 2008).
Takken and Knols, (2007) observed that malaria vectors preference of humans as major
source of blood meal, increases the chances of malaria transmission. Development of the
human malaria parasite Plasmodium spp form a cycle occurring both in the female Anopheles
mosquito (sporogony) and human host (schizogony) (Paaijmans, 2008). It is during these
recurrent cycles involving vector blood feeding, digestion and oviposition that malaria
transmission occurs. Therefore, the risk of malaria infection can be explained by studying
adult mosquito foraging behavior and ecology (Le Menach et al., 2005, Chaves et al., 2010).
Malaria control strategies require the consideration of both disease management and
transmission reduction through vector control. Moreover, like many other developing
countries, Kenya has for long been depending on insecticide treated bed nets (ITN) and indoor
residual spraying (IRS) to protect people from mosquito bites. The Kenya National Malaria
Strategy 2009-2017 expanded its policy on ITNs to include universal coverage, defined as
“one net per two people” in prioritized regions of the country by 2010, from the previous one
that focused on vulnerable populations (Kenya Ministry of Public Health & Sanitation, 2009).
The work in this thesis was undertaken to improve the understanding of patterns of host

3
seeking and oviposition of malaria vectors in an effort towards successful integrated vector
management (IVM) in malaria control campaigns.
1.2 Literature Review
1.2.1 Biology and Epidemiology of Malaria
Malaria transmission involves the interaction between susceptible human host, protozoan
parasite, arthropod vector, physical environments and socio-economic situations of the
society. As regards physical environments, malaria prevalence and entomological factors are
interdependent. Sub-Sahara Africa is the most malaria affected area in the world (Killeen, et
al., 2001), due to several factors that favor disease transmission. These include; (a)
predominance of Plasmodium falciparum Welch which causes severe forms of malaria
disease (Shillcutt, 2008), (b) the efficient malaria vectors, Anopheles gambiae Giles complex
(Anopheles gambiae sensu stricto and Anopheles arabiensis) and Anopheles funestus Giles,
(Beier, 1999), (c) suitable hot humid tropical climate and local weather conditions including
high temperatures, rainfall and humidity optimal for both parasite and vector mosquitoes
(Kiszewski & Teklehaimanot, 2004), and (d) the instability of socio-economic conditions
worsening situations for societies living in high malaria risk localities coupled with poor
health infrastructures and lack of suitable interventions and protective measures (Carter, et al.,
2000).
Plasmodium parasites known to cause malaria to humans belong to four species, P.
falciparum, P. malariae, P. vivax and P. ovale, with Plasmodium falciparum, being the most
virulent and common cause of febrile illness and associated mortalities. There is however an
increasingly infectious P. knowlesi simian species making up to the fifth human malaria
parasite (Chandler and Read, 1961). The disease is transmitted from person to person through
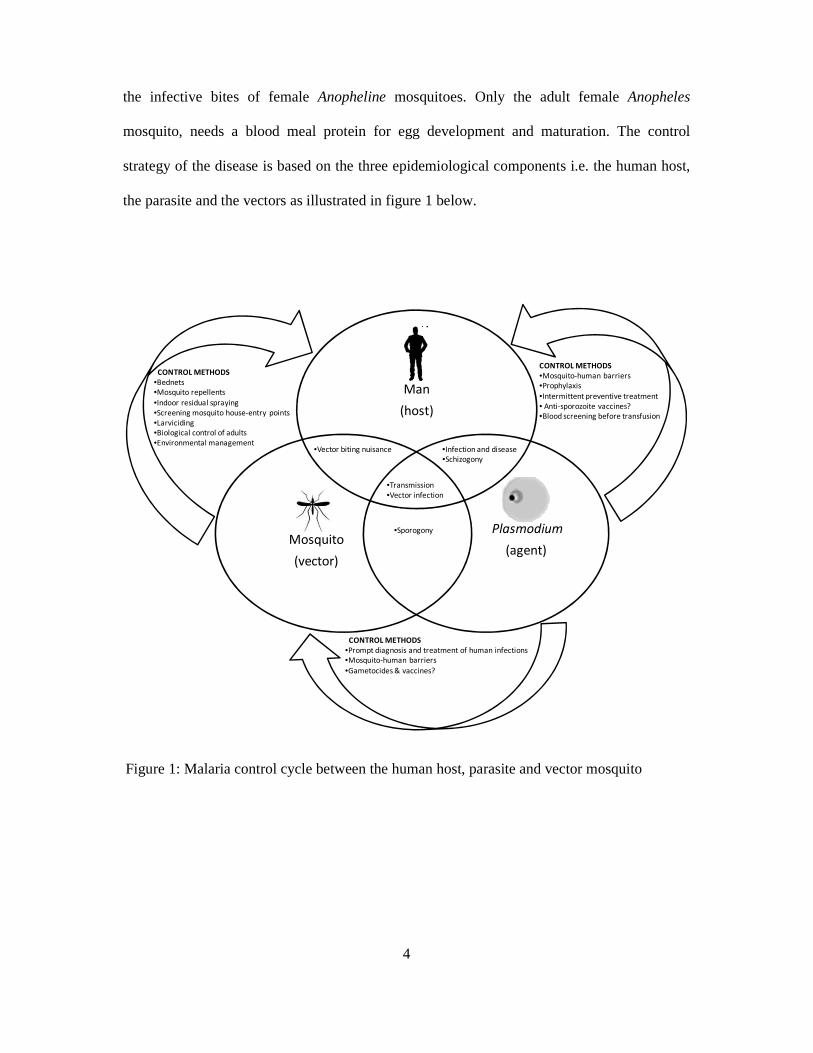
4
the infective bites of female Anopheline mosquitoes. Only the adult female Anopheles
mosquito, needs a blood meal protein for egg development and maturation. The control
strategy of the disease is based on the three epidemiological components i.e. the human host,
the parasite and the vectors as illustrated in figure 1 below.
Man
(host)
Plasmodium
(agent)Mosquito
(vector)
•Transmission
•Vector infection
•Vector biting nuisance
•Sporogony
•Infection and disease
•Schizogony
CONTROL METHODS
•Bednets
•Mosquito repellents
•Indoor residual spraying
•Screening mosquito house-entry points
•Larviciding
•Biological control of adults
•Environmental management
CONTROL METHODS
•Prompt diagnosis and treatment of human infections
•Mosquito-human barriers
•Gametocides & vaccines?
CONTROL METHODS
•Mosquito-human barriers
•Prophylaxis
•Intermittent preventive treatment
• Anti-sporozoite vaccines?
•Blood screening before transfusion
Figure 1: Malaria control cycle between the human host, parasite and vector mosquito

5
1.2.2 Malaria vectors (Afro-tropical species)
Anopheles mosquitoes can be distinguished from other mosquitoes by the palps, which are as
long as proboscis, and by presence of distinct marks on the wings. Adult Anopheles can also
be identified by their typical resting position, adults rest with their bodies in an acute or near
right angle to the surface upon which they rest (Gillies and DeMeilon, 1968).
Anopheles mosquitoes responsible for malaria transmission are widely distributed around the
world (except in Antarctica) from endemic areas to where the disease has been eliminated
(CDC, 2010). It is estimated that there are over 400 Anopheles species worldwide but only 30-
40 species are involved in malaria transmission, and about 1-2 species are major drivers of
disease transmission in a given area (White, 1974). Anopheles gambiae Giles complex is the
most efficient vector species of the fatal malaria parasite Plasmodium falciparum Welch
(Coetzee et al., 2000).
Anopheles gambiae complex comprises of seven sibling species; An. gambiae sensu stricto
Giles, An. arabiensis Patton, An. merus Donitz, An. melas Theobald, An. quadriannulatus
Theobald, An. bwambae White and An. quadriannulatus B, (Coetzee, 2004; Brooke et al.,
2002). In Sub-Sahara Africa, the major malaria vectors are Anopheles gambiae sensu stricto,
Anopheles arabiensis and Anopheles funestus Giles (Devine and Killeen, 2010) which are
widely distributed in Africa. In addition to the above 3 major malaria vectors, (Mbogo et al,
2003), reported An.merus as a secondary vector along the coast of Kenya. Other malaria
vectors along the coast of Kenya include Anopheles coustani, Anopheles squamosus,
Anopheles pharoensis and Anopheles nili (Mbogo et al., 1995). Anopheles gambiae complex
and Anopheles funestus tend to show seasonality trends in abundance and breeding. Anopheles
funestus breeds in swamps and the population density peaks at the end of rainy seasons and
onset of dry season (Gillies and DeMeilon, 1968).

6
1.2.3 Life cycle of mosquitoes
Mosquitoes have a holometabolous lifecycle (complete metamorphosis). They undergo four
different stages in their lifecycle; egg, larva, pupa and adult, the first three are aquatic and the
adult is terrestrial.
Some species such as Anopheles and Aedes mosquitoes lay their eggs singly on water surface,
but others such as Culex the eggs are glued together and laid in rafts. The Anopheles egg is
boat shaped, with bilateral floats, and cannot resist desiccation. A female adult mosquito can
lay between 50-200 eggs per single oviposition. In tropics the eggs hatch within 2-3 days due
to high temperatures, while in cold climates they last up to 3weeks.
Mosquito larva has got a well-developed head with a mouth brush for feeding, a thorax and
nine segments abdomen and has no legs. The Anopheles larvae unlike other mosquito, lack
respiratory siphon for respiration, instead they use spiracle placed on the eighth segment of
the abdomen and often lie parallel below the water surface. The Anopheles larvae feed on
bacteria, yeast, viruses, algae and other organic matter. Since they are surface feeders, they
spend most of the time on the water surface unless disturbed. The mosquito larval
development involves four larval instars, 1st, 2nd, 3rd, and 4th instars. In each instar molting
takes place after casting the exoskeleton for growth. Larval development to pupa takes about
4-7days under optimal conditions.
The pupa is comma shaped with fused head and thorax forming a cephalothorax, it doesn’t
feed and it is very active when disturbed. It frequently comes to water surface to breath by
using a pair of respiratory trumpets. The pupa molts to an adult mosquito after 1-3days
depending on ambient temperature.

7
The complete process from egg to adult varies from one species to another, under favorable
temperatures takes a minimum of 7 days, but usually takes 10-14 days in tropical countries.
1.2.4 Medical importance of mosquito
Mosquitoes apart from their biting annoyance are important vectors of diseases such as
malaria, lymphatic filariasis, dengue, yellow fever and many other arboviral diseases.
1.2.5 Oviposition and spatial distribution of habitats
Malaria vectors lay eggs in relatively clean slow moving and standing water such as irrigation
water, rice fields, shallow ditches, flooded depressions, river banks, marshes and burrow pits
(Oyewole et al., 2009). In addition, An. gambiae mosquitoes prefer small sunlit pools and
man-made habitats (Service, 1977, Shililu et al, 2003) in the vicinity of human habitations
(Carter, et al., 2000). In all mosquito species, location and selection of an oviposition site is
essential in life history because potential breeding sites vary with respect to both biotic and
abiotic characteristics (Sumba et al., 2004). The gravid Anopheles mosquito selects a suitable
breeding site to lay its eggs, (Tsila, et al, 2010) hence it determines the larval distribution in a
locality (Pates and Curtis, 2005). This selection influences reproductive potential and
determines survival rate of larvae (Bond et al, 2005). In selecting a suitable oviposition site,
gravid Anopheles mosquito use biological and chemical cues as attractants (Mokay & Shine,
2003). One female mosquito reproduces up to several hundred eggs over several broods of life
time. Soon after emerging, an adult female mosquito is normally ready to mate (Takken and
Lindsay, 2006) after which she begins to seek a blood meal.
1.2.6 Mosquito habitat productivity
Habitat productivity refers to the rate of adult mosquito emergence from individual aquatic
habitats. Proper knowledge of gravid mosquito foraging behaviour for oviposition is key in

8
understanding the habitat productivity (Gu et al, 2008). For instance, most gravid An.
gambiae mosquitoes tend to oviposit in habitats closer to the human habitations; hence these
habitats are relatively more productive than those located far away. The productivity of
available aquatic habitats within the vicinity of human habitations, determines adult vectors
population and the risk of malaria transmission in a locality (Munga et al, 2009). (Gu and
Novak, 2009) observed that aquatic habitats differ in mosquito productivity in heterogeneous
environments and this may facilitate targeted intervention. Empirical studies have suggested
that the major determinants of oviposition and larval abundances in a habitat are type and
location of aquatic habitats (Diabate et al., 2005; Munga et al., 2006; Mwangangi et al.,
2007). For instance, one of such studies reported that brick pits alone accounted for 60-80
percent of the total pupal samples in western Kenya (Mutuku et al., 2006).
1.2.7 Mosquito host seeking
Le Menach et al., (2005) reported that a set of cues such as host movement, body temperature,
odours, and carbon dioxide (Co2) are used to locate sources of blood-meal by newly emerged
mosquitoes from aquatic habitats. The availability of preferred hosts for blood meal within the
flight-range of malaria vectors influence the emergency rate, feeding cycle length, malaria
transmission dynamics and even survival of the vectors (Burkot, 1988, Killeen et al., 2001).
Success of host finding and oviposition varies from species to species and is dependent on the
flight ability of the mosquito, (Gu et al., 2006). Anopheles mosquitoes particularly differ in
host seeking behavior. Some species prefer to feed on humans, these are categorized and
referred to as anthropophagic, and others prefer to feed on animals and are referred to as
zoophagic. Mosquitoes with preference to feed indoors are referred to as endophagic whereas
outdoor feeding mosquitoes are referred to as exophagic; however these names are mere
references of the particular behavioral categories and not distinct nomenclature units.

9
Mosquito feeding behaviour as respects timing is a primary requisite for the understanding of
malaria transmission (Chandler and Read, 1961).
1.2.8 Anopheline mosquito foraging behavior
Foraging is a term used to refer to a definite set of behaviours exhibited when a female
mosquito searches for a host as a source of blood meal and oviposition site. These two
requirements consist central obligations and requirements for completion of the mosquito
gonotrophic cycle (Gu and Novak, 2009). Generally, endophily in Anopheles mosquitoes is
characterized by resting on the walls, ceiling, dark objects and under furniture. This often
occur prior to or after a blood meal; in the last case of which is crucial for digestion and egg
development. Engorged females fly to the nearby suitable water bodies for egg laying
(Mboera, 2005; Mboera et al 2006; Takken and Lindsay, 2006). A sexually mature female
mosquito usually needs two to three blood meals for initial oviposition. However, subsequent
feeding and oviposition patterns may vary slightly, in addition, the duration of the
gonotrophic cycle varies with ambient temperature ( Lyimo and Takken, 1993).
Generally, the distribution of humans and suitable habitats for mosquito larval development
vary across the landscape and the density of disease vectors fluctuates seasonally due to
dynamics of the habitat availability. These fluctuations are driven by environmental factors
mainly rainfall, temperature and topography which dictate the level of human-vector contact
and consequently disease transmission. Malaria vectors have a typical flight range of 1-2km
(Carter, et al., 2000) an essential component influencing distribution. The productivity of
breeding habitats and the effective dispersal range of the malaria vectors determine the
dimensions of malaria transmission in a region. A limited energy reserve restricts An.
gambiae from long-range flights to lay eggs in aquatic habitats far from human habitations.

10
(Le Menach et al., 2005) observed that vector dispersal can be influenced by the patterns of
host locality as well as the ecological situations of oviposition sites.
Survival and development of mosquitoes depends largely on appropriate physicochemical
parameters in the habitat, such as pH, temperature, salinity, oxygen content and nutrient
composition (Oyewole et al., 2009). ( and Takken 1993) observed that malaria parasite
transmission is dependent on the number of blood meals required to complete a gonotrophic
cycle which is influenced by body size of the female Anopheles.
Studies to assess the association of physicochemical factors and larval development have been
conducted before in Dakar, Senegal, (Robert et al., 1998) and western Kenya, (Gimnig et al.,
2001). In both studies, it has been shown that presence or development of Anopheles larvae in
pools correlate with water temperature, salinity, concentration of carbonates and nitrates. In a
separate study, (Mwangangi et al., 2007), along the coast of Kenya, using emergent cages
placed over natural habitats, found no association between physicochemical parameters with
body size of emerging mosquitoes, except for chlorophyll a. However, in a similar study of
habitat productivity, (Midega et al., 2007) concluded that there was variation in habitat
productivity along the coast of Kenya. The current study used both natural and artificial larval
habitats to evaluate this association.
(Kigadye et al., 2010) reported that due to variation in host preferences and abundance of
malaria vectors, there is need to identify and map species distribution in heterogeneous
environments. (Chaves et al, 2010) observed that Anopheles gambiae s.s prefer to feed mostly
on humans even when introduced to other hosts under controlled field trials while (Mahande
et al, 2007) reported that Anopheles arabiensis prefer feeding on animals. The availability of
humans as the sole source of blood meal alongside vector mosquito blood meal preference is
the primary reasons for anthropophagic behaviour (Burkot, 1988).

11
1.2.9 Malaria vector control
Vector control is an important aspect in the management of malaria transmission, being one of
the key components of the Roll Back Malaria (RBM), and can be directed either against the
immature stages or adult mosquitoes. The main goal of malaria vector control is to minimize
the vector population so as to reduce considerably the incidence and prevalence of both
parasite infection and clinical malaria. Several regions of the world use vector control as a
tool for malaria eradication and it has considerably reduced the incidence in some countries
(Killeen, et al 2004). From the late nineteenth century to early twentieth century, before the
invention of Dichlorodiphenyltrichloroethane (DDT), environmental management was the
main approach for malaria vector control, targeting aquatic stages; this included
environmental modification with permanent outcome and environmental manipulation with
temporary outcome (Castro, et al., 2010). The discovery of DDT which was extremely
effective made the control and eradication of malaria vectors feasible in some regions of the
world, and was used as an IRS in malaria control campaigns (Gu et al., 2008).
Among the current available chemical methods of mosquito control is the use of pesticides
such as Pyrethrin from plant flower extracts and synthetic pyrethroids which are used in ITNs
and IRS targeting adult mosquitoes as contact poisons. Mosquito population reduction can
also be achieved by hindering the dispersal through interrupting the life cycle using chemical
larvicides like organophosphates e.g. temephos, and insect growth regulators (IGRs) e.g.
methoprene (Hardin and Jackson, 2009). Others include use of biological methods like
bacteria. The bacterial larvicide applications consist of two bacterial strains of Bacillus
thuringiensis israeliensis (Bti) and Bacillus sphaericus (Bs). (Becker, 1998) reported the
introduction of large-scale routine operations of Bti for the first time in Europe in the first five
years of its invention. Tropical countries consider integrating biolarvicides into their control

12
programmes (Fillinger, et al., 2003). Tanzania has been successfully used Bti for killing
mosquito larval stages in a malaria control programme in Dar es salaam, (Fillinger et al.,
2008).
WHO defined Integrated vector management (IVM) as a “rational decision making process
for the optimal use of resources for vector control”. Several strategies are employed in control
of vector borne diseases, yet they remain a chief global public health challenge. Recent
evidence indicates the inclusion of IVM in national malaria control programmes in tropical
African countries has high potential for success of control, (Beier et al., 2008, Geissbuhler et
al., 2007). WHO has approved as first line control, IVM programme combining ITN, IRS and
source reduction in the fight against malaria vectors, (Muturi et al., 2008).
1.3 Problem statement
Tropical African countries rely on case management and insecticide-based control i.e.
insecticide treated nets (ITN) and indoor residual spraying (IRS) of the major vector species
in their malaria campaigns, despite resistance challenge. In these campaigns, insecticide
treated nets/long lasting insecticide nets (ITN/LLIN) have mostly been ranked as the top most
reliable tool for malaria vector control (GMP Report, 2001-2006). However, environmental
management for larval control has been neglected in Africa (Gu et al., 2006; Fillinger et al.,
2004). Increased coverage of ITNs and IRS in many parts of Africa has led to achievement of
vast benefits but the level of control is limited. Both ITNs and IRS target indoor mosquitoes
but the focus towards elimination in most settings should also target outdoor exposure to
mosquitoes (Ferguson, et al., 2010). Anthropophagic Anopheles mosquitoes are regarded to
be key vectors of malaria and lymphatic filariasis, (Snow, 1983), especially along the coast of
East Africa where the diseases are widely spread. Considering the importance of
environmental management, control of multiple vector borne diseases can be achieved when

13
integrated with other available tools. The success story of malaria eradication in Europe,
United States and the Middle East was based on integrating rigorous larval control measures
and other tools (Fillinger et al., 2003).
This study intended to develop a new perspective of environmental management by source
reduction, targeting mosquito foraging behavior. A study conducted by (Mbogo et al., 2003)
in the coastal area of Kenya demonstrated a spatial heterogeneity in the species composition
of Anopheles mosquitoes, however, factors associated with the spatial occurrence of larval
habitats were still not clear. In a different study of the same area (Mwangangi et al., 2007)
observed that targeting aquatic habitats surrounding house compounds could drastically
reduce mosquito abundance and malaria incidence, suggesting interference with foraging
movements as a promising target. The current study aimed at estimation of indices of spatial
connectivity to measure the availability of human hosts and aquatic habitats to host seeking
and gravid mosquitoes respectively by mapping and analyzing distribution patterns of host
seeking mosquitoes and larval populations.
1.4 Justification and significance of the research
The majority of female Anopheles mosquitoes alternate between human host for blood meal
and suitable aquatic habitats for oviposition (Sumba et al., 2004). Aquatic habitats vary in
physicochemical variables which affect oviposition preference, productivity and fitness of
larval populations. (Gu et al., 2008), noted that oviposition foraging is normally overlooked as
a factor governing distributional patterns of mosquito productivity. In planning malaria vector
control programs targeting mosquito aquatic stages, prior understanding and knowledge of the
local ecological situation and spatial distribution of breeding habitats are very important
aspects for success. However, the oviposition pattern and habitat distribution of most
ecosystems are not clearly understood thus the need to evaluate and understand them. In the

14
current study, spatio-temporal analysis of habitat productivity was conducted in Jaribuni
village along the Kenyan coast with an aim of elucidating crucial information on Anopheles
mosquito habitat suitability and productivity. Findings of this study will be useful to regional
authorities in planning strategies for mosquito larval control using environmental
management.
1.5 Hypothesis
Human host seeking patterns of Anopheles mosquitoes are influenced by oviposition patterns
1.6 Objectives
1.6.1 General objective
To estimate the host seeking and oviposition patterns of Anopheline mosquitoes
1.6.2 Specific objectives
1. To determine Anopheline habitat productivity and diversity.
2. To evaluate ecological factors which influence Anopheline productivity and emerging
mosquito fitness.
3. To assess spatial connectivity of Anopheline mosquitoes to their pre-adult aquatic
habitats.

15
2 CHAPTER TWO: MATERIAL AND METHODS
2.1 Study area
This study was carried out in Jaribuni village (03o37.3’S; 039o44.6’E) which lies
approximately 30km west of Kilifi town in the Kilifi County, along the Kenyan coast (Fig
2.1). Jaribuni lies at an altitude range of 40-400 meters above sea level and experiences two
wet seasons annually, with the long rains in April/June and short rainy period occurring in
October/December with mean annual precipitation range of 400mm-1,200mm. The mean
annual temperature ranges between 22oC and 30oC and the average relative humidity is
approximately 70% (Mtwapa Meteorological Station, 2010). Jaribuni River cuts across the
study site with abundant small pools of water and vegetation along both sides. The vegetation
cover in the area consists of bushes and shrubs. Jaribuni area is characterized by subsistence
farming such as cassava and maize, with some plantations of coconuts, and cashew nuts, and
scattered village houses. Small scale rearing of domestic animals such as cattle, goats and
poultry are also carried out in the area. The rural population mainly lives in mud-walled
houses with coconut-leaf-thatch (Makuti) roofs. Mosquitoes have free access to these houses
through eaves because they have no ceilings, moreover, unscreened windows/ventilation
openings and others are structurally defective. Homesteads are scattered and separated from
one another by agricultural land. The Kenya Medical Research Institute (KEMRI) has
established a semi-field station set up for mosquito experiments in Jaribuni village. The
present study area covered an area of (2x2) km2 grid in the village.
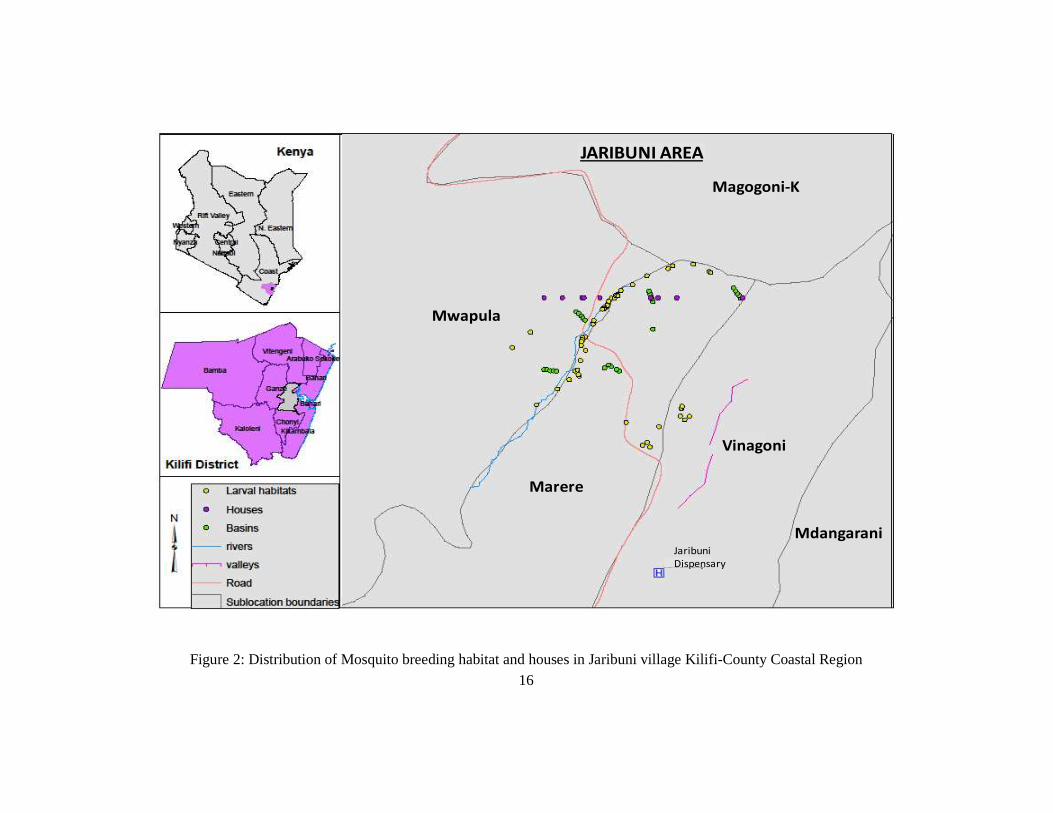
16
Mwapula
Marere
Jaribuni
Dispensary
Vinagoni
Magogoni-K
Mdangarani
JARIBUNI AREA
Figure 2: Distribution of Mosquito breeding habitat and houses in Jaribuni village Kilifi-County Coastal Region

17
2.2 Experimental design
2.2.1 Anopheles habitat productivity and diversity
2.2.1.1 Breeding site identification and mapping
A longitudinal survey of potential mosquito breeding habitats was conducted within 4km2
grid of the Jaribuni village on fortnight basis for a period of five months between December
2010 and April 2011. The grid was selected to cover part of the river Jaribuni which was the
main breeding site during the dry season and adjacent human habitations. The grid was set by
measuring 1km each direction towards North, South, West and East, using hand held
geographical positioning system (GPS) machine (eTrex© Vista HCx, Garmin), the centre
point was a bridge crossing the river Jaribuni. All temporary habitats such as ditches, trenches
and abandoned water storage tanks and permanent open water bodies such as river and
adjacent pools were considered as potential mosquito breeding habitats and recorded.
Immature aquatic mosquito stages were sampled by a standard dipper (350mls) depending on
the size of the habitat. During sampling, 10-20 dips were made in each habitat depending on
its size, pooled together and sieved into enamel trays to constitute a single sample for the
particular habitat. Field collected samples were placed into whirl packs and then transferred to
field laboratory for further processing. Larval habitats and human habitations were allocated
different codes and geo-referenced by taking coordinates using (GPS).
2.2.1.2 Mosquito larval identification
All mosquito larvae and pupae were placed in enamel trays in the laboratory and sorted out
into anopheline and culicines from other accidentally collected aquatic organisms and debris.
Anopheles and culicine larvae were further categorized as early (1st and 2nd) or late (3rd & 4th)
instars larvae. Pupae were generalized for both anophelines and culicines and placed in

18
emergent cages for adults to emerge. The emerged adults were then morphologically
identified into species using the keys of Gillies and Coetzee, 1987.
2.3 Ecological factors influencing Anopheles productivity, and emerging mosquito fitness.
2.3.1 Artificial habitats
Plastic basins (Figure 2) were placed at selected sites along transects identified in the study
village (Jaribuni) with the reference point being five inhabited houses that were also sampled
for adult mosquitoes on fortnight basis (Section 3.7). Each transect was 100m long and five
artificial habitats were placed at intervals of 25 meters along its length (0m, 25m, 50m, 75m
and 100m). Each habitat was filled with approximately 1.5kg of fresh mud and three liters of
sieved river water. The habitats were coded for easy reference and left open to allow
colonization by wild mosquitoes. The habitats were then monitored on daily basis for
oviposition by wild mosquitoes. Mosquito larvae in the artificial habitats as well as
physicochemical characteristics such as temperature, conductivity, Dissolved oxygen (DO)
and salinity were recorded daily with the aid of YSI EC 300 machine, Brannum Lane, Yellow
Spring, OH, USA and DO by CORNING 312 machine, Corning Incorporated, NY USA.
Turbidity was monitored by expert visual assessment and recorded as clear, low or high.
Water was added daily into artificial habitats to replenish amount lost through evaporation.
The number of early instar larvae were daily collected, their numbers estimated, and
transferred to the laboratory for further processing. Pupae were placed in holding cages in the
laboratory and allowed to emerge as adults to facilitate morphological identification using
keys by Gillies and Coetzee (1987). For the emergent adults, only Anopheles mosquitoes were
identified to species level.

19
Figure 3: Artificial habitat for monitoring mosquito oviposition preferences and distances from houses
2.3.2 Emergence cages
In naturally occurring mosquito habitats, cages of 0.5mx0.5mx0.5m and 0.3mx0.3mx0.3m
were used to estimate habitat productivity. The cages were made of iron rod frames and
welded with 6-inches wide iron plates on the lower sides to prevent in and out movement of
water and mosquito larvae. The cages were then covered with a netting material for the
control of emerging adults (Figure 3). The cages were placed on selected representative
natural habitats identified in the study area that had aquatic stages of Anopheles mosquitoes.
The emerged adult mosquitoes from the cages were collected daily by simple mouth aspirator
and placed in paper cups. The specimens were transferred to KEMRI, Kilifi Vector biology
laboratory for further processing. In the laboratory, the identified Anopheles mosquitoes were
killed by freezing and one wing removed and mounted on the microscope slide by using DPX
mounting solution for measurement of wing length. The wing length measurement was done

20
with the help of ocular micrometer mounted on a microscope eye piece for the estimation of
average adult body size. Wing length measurements were taken from the distal end of the
alula to the tip, excluding the fringe of scales.
Figure 4: Emergent cage placed over natural habitat to monitor productivity
2.3.3 Spatial connectivity of Anopheles mosquitoes to pre-adult aquatic habitats
2.3.3.1 Sampling procedure
Probability sampling design was used to select ten houses for aspiration in the village block.
The area was divided in 4 sub-blocks or grids (A=25 houses, B=23 houses, C=18 houses and
D=16 houses) for even distribution of the sampled houses. Number of houses in each of the 4
sub-blocks of 1km2 was pre-determined. The stratified sampling method was applied to get
the number of houses for indoor sampling in each grid, then, simple random method was

21
employed in selecting the required number of houses. The following formula was used:
Sample in a sub-block = number of houses in a sub-block x sample size n (10)
Total number of houses in a block
A= (25x10)/82=3houses, B= (23x10)/82=3houses, C= (18x10)/82=2houses and
D= (16x10)/82=2houses
2.3.3.2 Indoor resting mosquito collection method
Sampling of human host seeking mosquitoes (indoor resting) was done using aspiration
collection method as described by Service (1993) in ten selected houses between 06.00am and
11.00am on a fortnight basis for six months (December 2010-May 2011). The method used a
motorized improved “CDC backpack Aspirator” Model 1412 (John W. Hock Company,
Florida, USA), originally developed by U.S. Public Health Services to collect the indoor-
resting adult mosquitoes such as Aedes aegypti. The collection cup was inserted into the wand
with slight twist until it fitted. The blower was then turned on with the switch located in the
wand. The aspirator was powered by 12v rechargeable battery. Collections were then made by
systematically moving the flexible wand on wall surfaces, roofs, hanging clothes and under
furniture to collect resting mosquitoes. After each collection, the collection cup was closed
before turning off the blower to prevent escape of caught mosquitoes. The adult mosquitoes in
collection cups were then stored in cold boxes to prevent drying up during transfer to the
laboratory where they were then killed in a freezer under -20oC temperatures. Caught
mosquitoes were identified by the use of morphological appearance with the aid of
entomological Microscope and identification key by (Gillies and Coetzee, 1987); (Gillies and

22
DeMeillon, 1968). Anophelines were identified to species level whereas the rest were
generally regarded as culicines. The physiological status of mosquitoes was determined
through abdominal examination of freshly collected samples as unfed, fed, half gravid or
gravid.
2.3.3.3 Sporozoite rate
Enzyme linked immunosorbent assay (ELISA) technique was performed to establish infection
of female Anopheles mosquitoes collected indoors. This was based upon presence of specific
anti-Plasmodium falciparum sporozoite monoclonal antibodies (MAbs) from mosquito
salivary glands. Female Anopheles mosquitoes were cut between thorax and abdomen; heads
and thoraces were left to soak in the Nonidet P-40 for 20 minutes before being used to test for
the presence of Plasmodium falciparum circumsporozoite proteins. The “sandwich” ELISA
began by adsorption of the capture MAbs to the wells of the microtitre plate. After 30 minutes
incubation at room temperature, the well contents were aspirated and the remaining active
binding sites on the plate were blocked with blocking buffer. Mosquito samples were ground
individually in a 1.5 ml vial by plastic rod in blocking buffer containing Nonidet P-40, diluted
with blocking buffer. Resultant preparations were then aliquoted into microtitre plate wells.
Positive and negative controls were also added to specific wells. After 2 hour incubation, the
mosquito triturate was aspirated and the wells were washed two times with PBS-Tween 20
and banged to dry. The respective peroxidase-linked MAbs were then added to the wells,
completing the formation of the “sandwich”. After one hour incubation at room temperature
the well contents were aspirated, the plate was washed three times with PBS-Tween 20
solution and banged to dry and the clear peroxidase substrate was added. A dark green
product was formed as the peroxidase enzyme reacted with the substrate, the intensity of the

23
color being relative to the amount of circumsporozoite (CS) antigen present in the test sample.
The results were read visually after 30 minutes of incubation.
The sporozoite rate was estimated as a proportion (n/N), calculated as the number of infected
mosquitoes (n) divided by the total number tested (N).
2.3.3.4 Blood meal ELISA analysis
Blood fed and half gravid abdomens of individual mosquitoes were triturated manually with
plastic pestle in1.5ml microfuge tube containing 100µl phosphate buffer saline (PBS). Nine
hundred microlitres of the blocking buffer was then added to each sample to make a final
volume of 1ml. Samples were then transferred to polyvinyl microtitre plate wells in aliquots
of 50µl and incubated overnight at room temperature. The plate was then washed twice with
PBS-Tween 20 followed by addition of 50µl host specific conjugate in 5ml boiled casein-
Tween 20 enzyme diluents then incubated for one hour at room temperature. The wells
washed 3 times with PBS-Tween 20 then 100µl of ABTS peroxidase substrate was added to
each well. After 30 minutes of incubation, the dark green positive reactions for peroxidase or
dark yellow reactions for phosphotases were visually assessed. A second host source was
determined in the same microtitre plate where mosquitoes were screened for human blood.
The phosphotase-labeled anti-bovine IgG was added to the peroxidase-labeled antihuman IgG
solution. Blood meal were screened first for human IgG by addition of peroxidase substrate,
reading made after 30 minutes and wells washed 3 times with PBS-Tween 20 then to each
well 100µl of phosphatase substrate was added. Plate was read after 1 hour to determine
positive bovine reactions. Non-reacting samples were then tested for goat blood. Test serum
from each host was added except that for which enzyme conjugate was added. Microtitre plate
contained control serum from each host and blank controls containing 50µl of PBS alone.

24
Figure 5: A characteristic house within Jaribuni village where indoor resting mosquitoes were sampled

25
2.4 Data analysis
Field data collection was done and entered in pre-designed field data forms then transferred
into Excel spreadsheets. The data was first cleaned for errors and analysis done using R-
statistical package. One way analysis of variance (ANOVA) was used to compare the
productivity of different larval habitats while the student t-test was used to compare early and
late instars abundance in habitats, the same test was used to compare the mean larval density
of each habitat against the overall mean. Regression analysis was used to test for correlation
between the various physicochemical factors and habitat productivity of mosquitoes and
distance from aquatic habitats to indoor resting Anopheles population. To generate a map
showing selected houses and locations of breeding sites, a computerized program, ESRI
ArcMap version 10 was employed (Figure 1). The productivity in artificial habitats was
determined by the number of larvae per habitat per day. The productivity of malaria vectors in
natural habitats was determined by the number of emerging adult mosquitoes per m2 per day.
The sporozoite rate was calculated as a proportion of number infected over total number
tested. Human blood index (HBI) and Goat blood index (GBI) was calculated as proportion of
number positive for a particular host over number of tested mosquito samples.
2.5 Ethical consideration
Permission to conduct this study was granted by KEMRI National ethical review committee
(permit number KEMRI SSC 1719). The informed verbal consent was sought from heads of
households for the study to be carried out in their farms and household surroundings for larval
habitat searching and artificial habitat and aspiration to be performed in their houses after

26
3 CHAPTER THREE: RESULTS
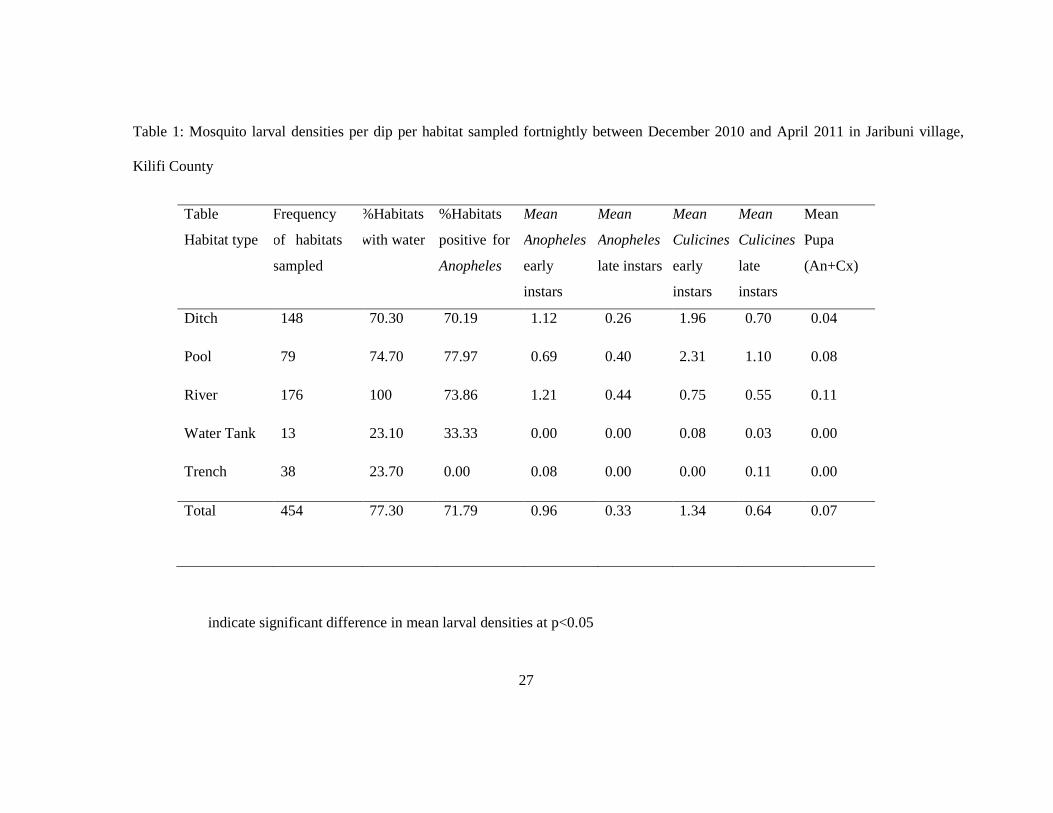
3.1 Habitat productivity and diversity
A total of five different habitat types were identified and sampled for mosquito productivity in
Jaribuni village during the study. The five habitat types included temporary pools, abandoned
water tanks, ditches, river and trenches. A total of 454 sampling visits were made to the
habitats with a proportion of 77.3% of the habitats having water (Table 1). Of this, the river
was the most stable habitat in terms of its capacity to retain water most of the time (100%)
followed by temporary pools (74.7%) and ditches (70.3%). Abandoned water tanks and
trenches were less stable habitats, they were only found with water when it rained.
Abandoned water tanks in 76.9% of the sampling visits were found dry and trenches were dry
in 76.7% of the sampling visits. The highest proportion of presence of anopheline mosquitoes
was recorded in pools with the least recorded in water tanks.
3.2 Larval productivity
The mean early and late stage larva, and pupal densities per dip per habitat for anophelines
and culicines were monitored during the entire six month study period (Table 1). The data
showed gradual decrease in larval densities per dip per habitat from early to late instars larvae
and from late instars to pupae in all five types of aquatic habitats. Early instar densities were
significantly higher than late instar densities for both anophelines and culicines at p<0.05
(Table 1). One way ANOVA test showed a statistically significant difference in Anopheles
larval productivity among the five habitat types. When comparing the means of Anopheles
larval density of each habitat type (ditches, pools, river and abandoned water tanks), against
overall mean larval density, the river had significantly higher mean density.
27
Table 1: Mosquito larval densities per dip per habitat sampled fortnightly between December 2010 and April 2011 in Jaribuni village,
Kilifi County
Table
Habitat type
Frequency
of habitats
sampled
%Habitats
with water
%Habitats
positive for
Anopheles
Mean
Anopheles
early
instars
Mean
Anopheles
late instars
Mean
Culicines
early
instars
Mean
Culicines
late
instars
Mean
Pupa
(An+Cx)
Ditch 148 70.30 70.19 1.12 0.26 1.96 0.70 0.04
Pool 79 74.70 77.97 0.69 0.40 2.31 1.10 0.08
River 176 100 73.86 1.21� � 0.44� � 0.75 0.55 0.11
Water Tank 13 23.10 33.33 0.00 0.00 0.08 0.03 0.00
Trench 38 23.70 0.00 0.08 0.00 0.00 0.11 0.00
Total 454 77.30 71.79 0.96� � 0.33 1.34�
�
0.64 0.07
� � indicate significant difference in mean larval densities at p<0.05

28

28
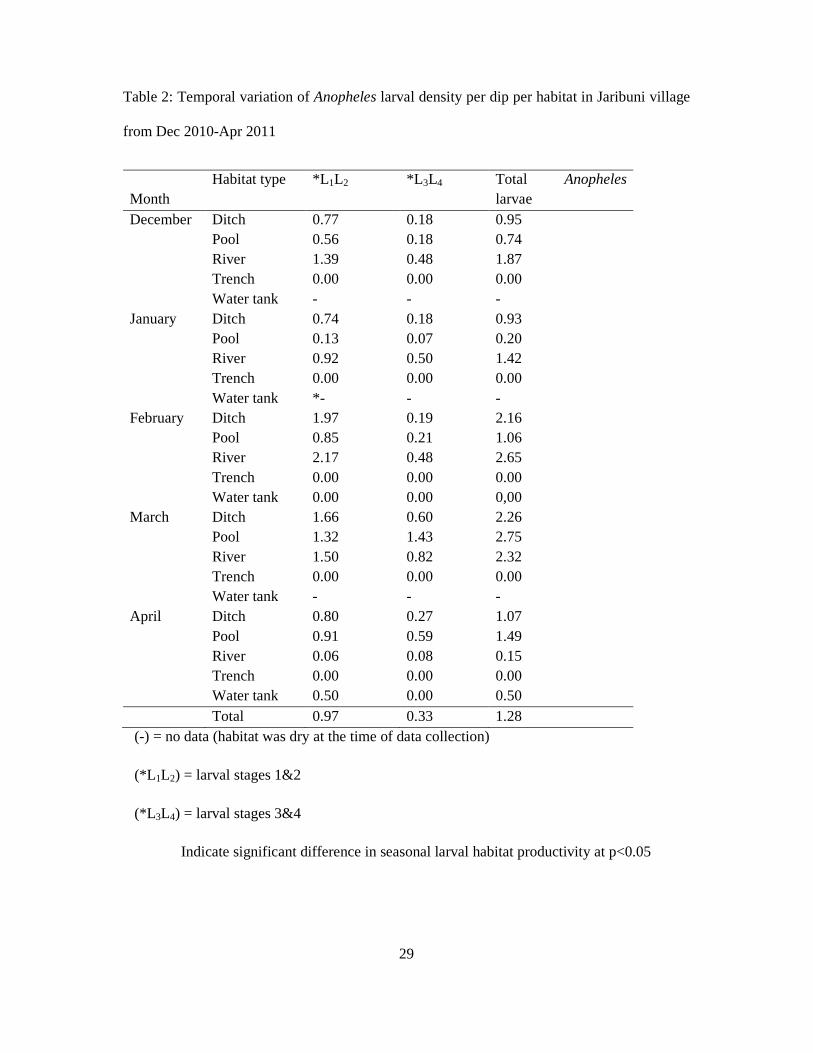
3.3 Seasonal variation in larval habitat productivity
On temporal basis, all the habitats showed differential productivity with the month of March
recording significantly higher overall productivity of anopheline larvae (p<0.05). Temporal
productivity varied significantly among different habitats (Table 2). Normally, the river
became more productive during the dry season when water volume and velocity was low. The
month of April was the wettest month compared to the rest of the four months. The number of
small pools along the river and ditches increased and the ability to retain water longer for
mosquito larval development and consequently the productivity also increased. Overall larval
density was 1.28 larvae per dip, with observed early instar density recorded as 0.97 and late
instar density as 0.33 larvae per dip per habitat (Table 2). Higher larval densities were
recorded in the river habitats in the first three months of larval sampling, i.e., December to
February (range 1.49-2.65) while the highest larval density of 2.75 was recorded in March
compared to April (1.49 larvae per dip) in pools (Table 2).
29
Table 2: Temporal variation of Anopheles larval density per dip per habitat in Jaribuni village
from Dec 2010-Apr 2011
Month
Habitat type *L1L2 *L 3L4 Total Anopheles larvae
December Ditch 0.77 0.18 0.95 Pool 0.56 0.18 0.74 River 1.39 0.48 1.87 Trench 0.00 0.00 0.00 Water tank - - - January Ditch 0.74 0.18 0.93 Pool 0.13 0.07 0.20 River 0.92 0.50 1.42 Trench 0.00 0.00 0.00 Water tank *- - - February Ditch 1.97 0.19 2.16 Pool 0.85 0.21 1.06 River 2.17 0.48 2.65 Trench 0.00 0.00 0.00 Water tank 0.00 0.00 0,00 March Ditch 1.66 0.60 2.26� � � Pool 1.32 1.43 2.75� � � River 1.50 0.82 2.32� � � Trench 0.00 0.00 0.00 Water tank - - - April Ditch 0.80 0.27 1.07 Pool 0.91 0.59 1.49 River 0.06 0.08 0.15 Trench 0.00 0.00 0.00 Water tank 0.50 0.00 0.50 Total 0.97 0.33 1.28
(-) = no data (habitat was dry at the time of data collection)
(*L1L2) = larval stages 1&2
(*L3L4) = larval stages 3&4
� �� � Indicate significant difference in seasonal larval habitat productivity at p<0.05

30
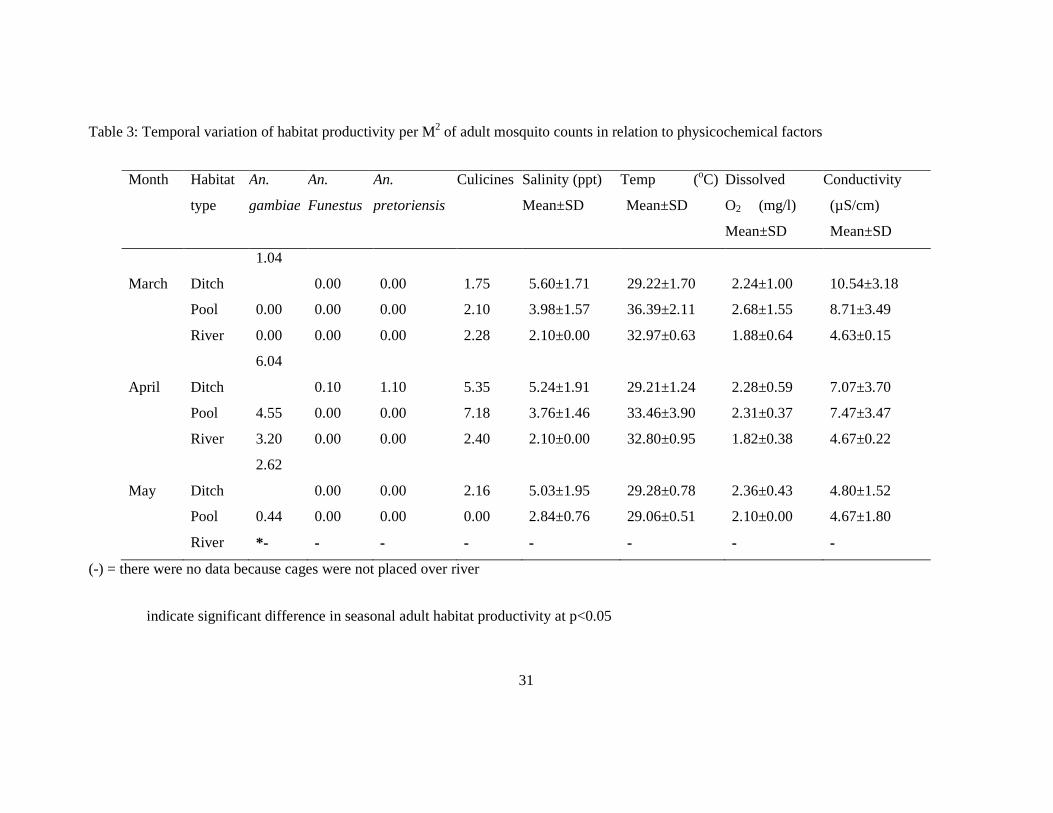
3.4 Adult productivity
Three wild type Anopheles species and culicines of various species were collected from
naturally occurring habitats using emergent cages placed over the habitats and monitored for
adult productivity (Table 3). A total 247 adult Anopheles mosquitoes were collected in three
months from March-May 2011 and were dominated by An. gambiae, 225(91.1%) followed by
An. pretoriensis 21(8.5%) and An. funestus 1(0.4%). The proportion of An. gambiae
mosquitoes was significantly higher (91.1%) with ditches being the most productive habitats
throughout the experiment. An ANOVA and Turkey’s HDS test of significance for An.
gambiae adult productivity showed that the productivity within the cages differed
significantly over time (F=53.13, df=2, 8, p<0.05) with the month of April being more
productive compared to other months. Similarly, ditches were found to be the most productive
habitats for An. gambiae adults per m2, (F=12.75, df=2, 6, p=0.02). Only one An. funestus
emerged and a low density of An. pretoriensis (1.1mosquito per m2) was recorded in April
from ditches (Table 3). In May the river was not monitored for adult productivity, because
water level was high and the cages could not be placed. High water volume during the month
of May lowered the adult density per square metre from pools.
31
Table 3: Temporal variation of habitat productivity per M2 of adult mosquito counts in relation to physicochemical factors
Month Habitat
type
An.
gambiae
An.
Funestus
An.
pretoriensis
Culicines Salinity (ppt)
Mean±SD
Temp (oC)
Mean±SD
Dissolved
O2 (mg/l)
Mean±SD
Conductivity
(µS/cm)
Mean±SD
March Ditch
1.04�
� � 0.00 0.00 1.75 5.60±1.71 29.22±1.70 2.24±1.00 10.54±3.18
Pool 0.00 0.00 0.00 2.10 3.98±1.57 36.39±2.11 2.68±1.55 8.71±3.49
River 0.00 0.00 0.00 2.28 2.10±0.00 32.97±0.63 1.88±0.64 4.63±0.15
April Ditch
6.04�
� � 0.10 1.10 5.35 5.24±1.91 29.21±1.24 2.28±0.59 7.07±3.70
Pool 4.55 0.00 0.00 7.18 3.76±1.46 33.46±3.90 2.31±0.37 7.47±3.47
River 3.20 0.00 0.00 2.40 2.10±0.00 32.80±0.95 1.82±0.38 4.67±0.22
May Ditch
2.62�
� � 0.00 0.00 2.16 5.03±1.95 29.28±0.78 2.36±0.43 4.80±1.52
Pool 0.44 0.00 0.00 0.00 2.84±0.76 29.06±0.51 2.10±0.00 4.67±1.80
River *- - - - - - - -
(-) = there were no data because cages were not placed over river
�� � � indicate significant difference in seasonal adult habitat productivity at p<0.05

32
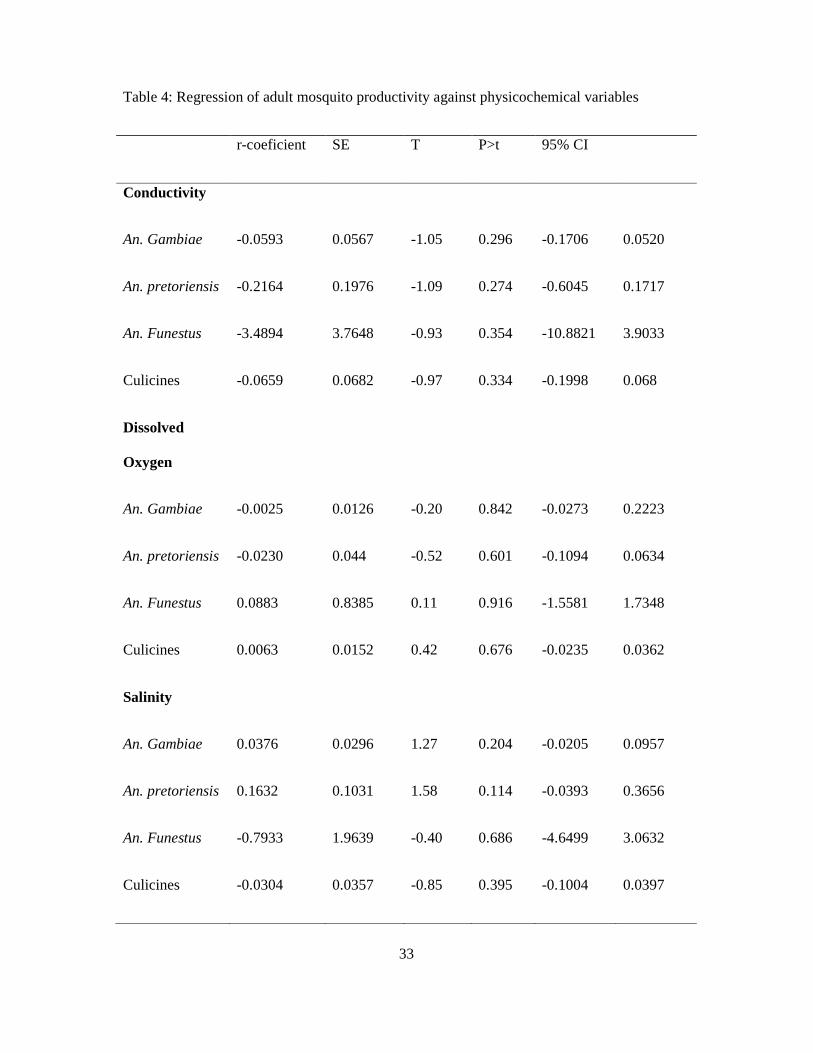
3.5 Influence of physicochemical factors on habitat productivity
A summary of the mean range of physicochemical factors and corresponding adult
productivity in different habitat types is shown in table 5. Pearson’s χ2 gave slight association
between habitat type and temperature (p<0.001) with mosquito productivity. Table 4 shows
regression analysis to test association of adult Anopheles species productivity against
conductivity, dissolved oxygen (DO) and salinity.
3.5.1 Conductivity
For each unit increase in An. gambiae, there was a decrease in conductivity by 0.059 units.
This indicates that there was negative correlation between conductivity and An. gambiae
productivity (r<0).
3.5.2 Dissolved Oxygen
An. gambiae and An. pretoriensis showed negative correlation to DO (r<0). Increasing
dissolved oxygen was associated with a decrease in the two species. Anopheles funestus and
culicines showed positive correlation; an increase in dissolved oxygen resulted in the increase
of the two (r>0).
3.5.3 Salinity
An. gambiae and An. pretoriensis showed positive correlation to salinity (r>0), a slight
increase of salinity resulted to increase in the two species (Table 4). Anopheles funestus and
culicines showed a negative correlation to salinity (r<0), increasing salinity led to a reduction
in the density of the two.
33
Table 4: Regression of adult mosquito productivity against physicochemical variables
r-coeficient SE T P>t 95% CI
Conductivity
An. Gambiae -0.0593 0.0567 -1.05 0.296 -0.1706 0.0520
An. pretoriensis -0.2164 0.1976 -1.09 0.274 -0.6045 0.1717
An. Funestus -3.4894 3.7648 -0.93 0.354 -10.8821 3.9033
Culicines -0.0659 0.0682 -0.97 0.334 -0.1998 0.068
Dissolved
Oxygen
An. Gambiae -0.0025 0.0126 -0.20 0.842 -0.0273 0.2223
An. pretoriensis -0.0230 0.044 -0.52 0.601 -0.1094 0.0634
An. Funestus 0.0883 0.8385 0.11 0.916 -1.5581 1.7348
Culicines 0.0063 0.0152 0.42 0.676 -0.0235 0.0362
Salinity
An. Gambiae 0.0376 0.0296 1.27 0.204 -0.0205 0.0957
An. pretoriensis 0.1632 0.1031 1.58 0.114 -0.0393 0.3656
An. Funestus -0.7933 1.9639 -0.40 0.686 -4.6499 3.0632
Culicines -0.0304 0.0357 -0.85 0.395 -0.1004 0.0397

34
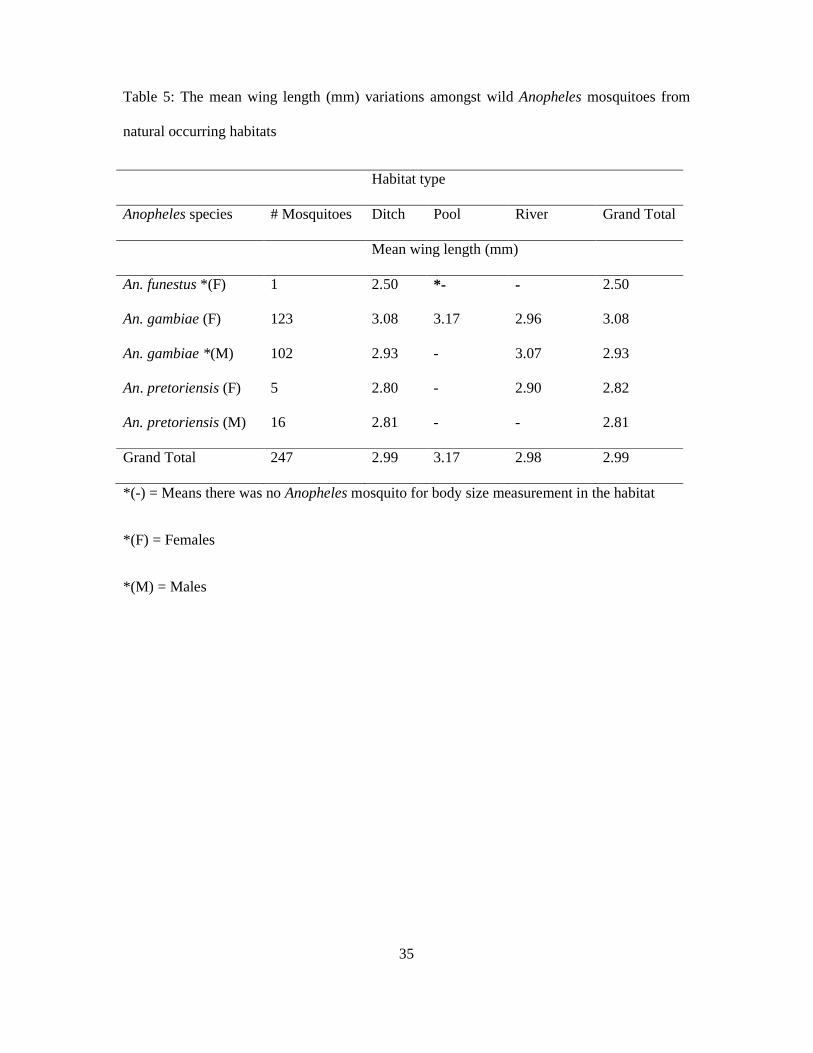
3.6 Body size variation in wild Anopheles mosquito population
Table 5 shows the mean body size (mm) variations amongst wild mosquitoes from natural
occurring habitats. A total of 247 samples of wild Anopheles mosquitoes collected from
emergent cages were analyzed for body size variation through wing length measurements
amongst different caged habitats in the study area. Of this, 225 (123females & 102males)
were An. gambiae, 21 (5females & 16males) were An. pretoriensis and only 1(female) was
An. funestus (Table 5). There were slight differences with respect to mosquito body sizes by
habitats. Anopheles gambiae females which emerged from ditches had mean body size of
3.08mm (range 2.5mm-3.8mm), those from pools had mean body size of 3.17mm (range
3mm-3.4mm) and those from the river had 2.96mm (range 2.6mm-3.5mm) mean body size.
Anopheles pretoriensis females emerged from ditches had mean body size of 2.8mm (range
2.5mm-3mm) while those from the river had 2.9mm mean size. One An. funestus female
emerged from ditch and had a body size of 2.5mm.
35
Table 5: The mean wing length (mm) variations amongst wild Anopheles mosquitoes from
natural occurring habitats
Habitat type
Anopheles species # Mosquitoes Ditch Pool River Grand Total
Mean wing length (mm)
An. funestus *(F) 1 2.50 *- - 2.50
An. gambiae (F) 123 3.08 3.17 2.96 3.08
An. gambiae *(M) 102 2.93 - 3.07 2.93
An. pretoriensis (F) 5 2.80 - 2.90 2.82
An. pretoriensis (M) 16 2.81 - - 2.81
Grand Total 247 2.99 3.17 2.98 2.99
*(-) = Means there was no Anopheles mosquito for body size measurement in the habitat
*(F) = Females
*(M) = Males

36
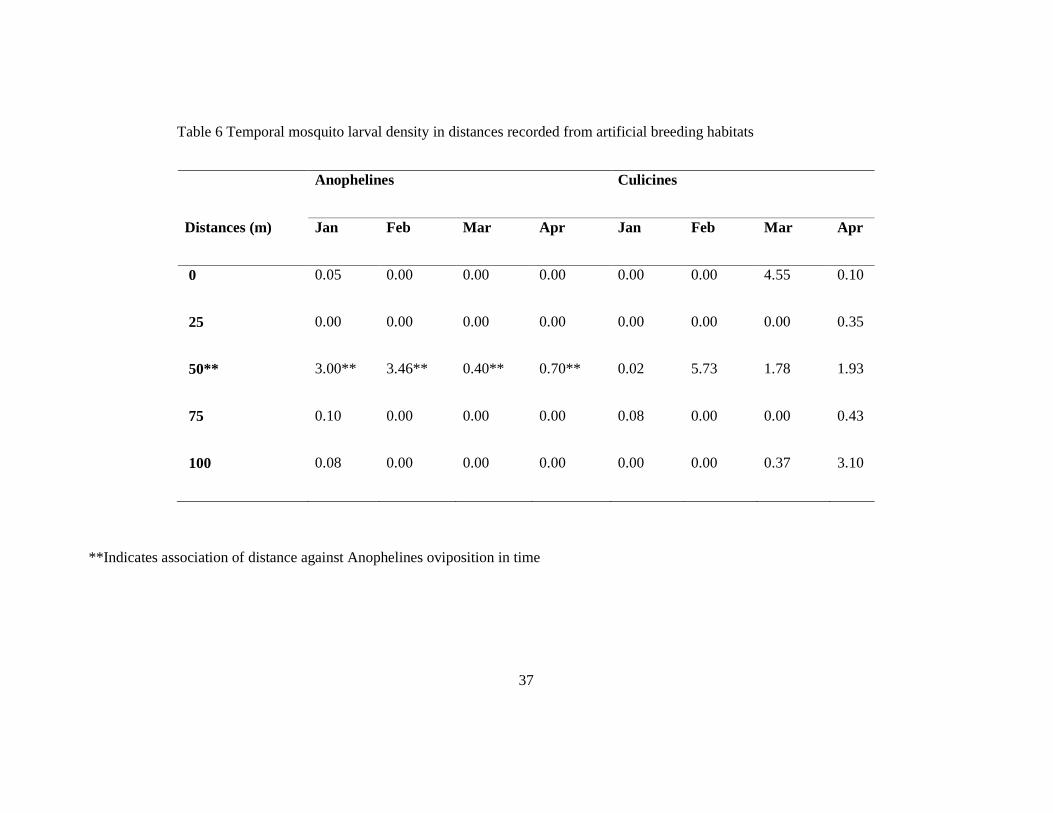
3.7 Correlation of habitat productivity and distance from human habitation
The experiment done using artificial breeding habitat to monitor Anopheline breeding
preference in distance from human habitation, Anopheles gambiae was the only malaria
vector species that was identified as having oviposited in the artificial habitats placed at 0, 25,
50, 75 and 100 meter distances (Table 6). The highest density of 3.46 Anopheles larvae per
habitat per month was recorded in February from habitats placed at 50 meter positions and the
lowest density of 0.05 larvae per habitat per month at location 0 meters in January. The
highest culicine larval density of 5.73 larvae per habitat was recorded in February in habitats
located 50 meters from human house. Both anopheline and culicine larvae were recorded in
habitats located at position 50 meters in all four months of the experiment (Table 6). Culicines
were not specific in distance and time preferences. There was no seasonal variation in
productivity of mosquitoes with distance from houses.
37
Table 6 Temporal mosquito larval density in distances recorded from artificial breeding habitats
Anophelines Culicines
Distances (m) Jan Feb Mar Apr Jan Feb Mar Apr
0 0.05 0.00 0.00 0.00 0.00 0.00 4.55 0.10
25 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.35
50** 3.00** 3.46** 0.40** 0.70** 0.02 5.73 1.78 1.93
75 0.10 0.00 0.00 0.00 0.08 0.00 0.00 0.43
100 0.08 0.00 0.00 0.00 0.00 0.00 0.37 3.10
**Indicates association of distance against Anophelines oviposition in time

38
3.8 Effects of mosquito larval habitat distribution on indoor resting densities
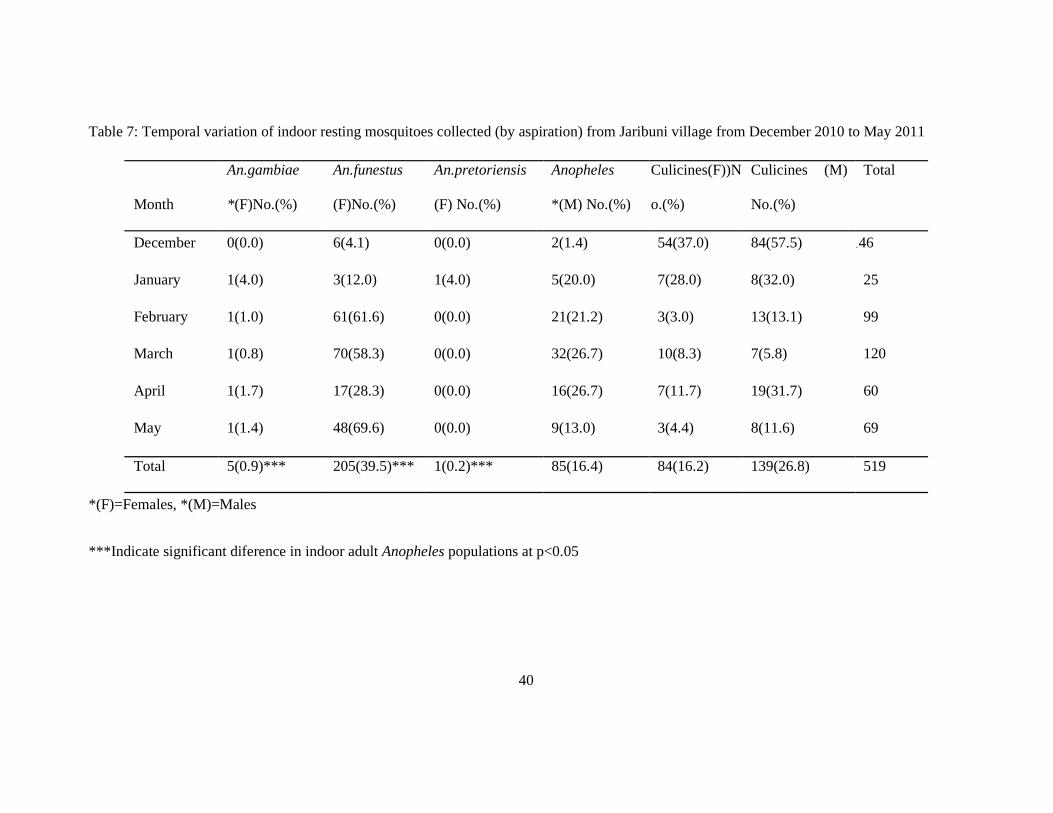
A total of 519 adult mosquitoes were collected from ten randomly selected houses in Jaribuni
village. A summary of the different species and their distribution is shown on table 7 where
anopheline mosquitoes constituted 57% (296) of the total collection while various species of
culicines constituted 43% (223). Among the anophelines, 85(28.7%) males were collected
resting indoors. Female anophelines comprised of three species namely, An. funestus
205(97.1%), An. gambiae 5(2.4%), and An. pretoriensis 1(0.5%). Generally, the female An.
funestus were the most predominant vector species collected indoors. The temporal variation
of indoor female Anopheles populations, (Table 7) shows that December and January had
very low proportions of indoor resting Anopheles populations 8(5.5%) and 10(40%)
respectively. There was a significant association between months and number of each
Anopheles species (Pearson χ2=287.57, df=25, P=0.000)
Table 8 shows correlation of indoor resting Anopheles mosquito densities to distance from
nearest breeding site in Jaribuni village. Houses located between 60-250meters from the
nearest breeding habitats had higher mosquito densities compared to those houses that were
less than 50 and more than 250 meters from the breeding site. Generally, more than 98% of all
adult indoor collected malaria vectors were collected within this range but the peak densities
were in houses located at 60meters and 220meter distances. The nearest breeding habitats in
these two distances were the river and pools. However, lower An. gambiae and An. funestus
densities, (0.18 and 0.27 respectively) per house per collection were realized in houses near
pools. The correlation analysis of indoor resting Anopheles densities against habitat type
indicated the number of An. gambiae and An. pretoriensis reduced from ditch to river by
8.25% and 30.15% respectively, while An. funestus increased by 38.76%, the reason is the
species habitat preferences (Table 8) and was highest in the river. There was negative
correlation between the mean densities of all three Anopheles species and distances, the

39
correlation analysis showed an increase in one meter led to 40.51% An. gambiae, 32.89% An.
pretoriensis and 16.23% An. funestus reduction.
40
Table 7: Temporal variation of indoor resting mosquitoes collected (by aspiration) from Jaribuni village from December 2010 to May 2011
Month
An.gambiae
*(F)No.(%)
An.funestus
(F)No.(%)
An.pretoriensis
(F) No.(%)
Anopheles
*(M) No.(%)
Culicines(F))N
o.(%)
Culicines (M)
No.(%)
Total
December 0(0.0) 6(4.1) 0(0.0) 2(1.4) 54(37.0) 84(57.5) 146
January 1(4.0) 3(12.0) 1(4.0) 5(20.0) 7(28.0) 8(32.0) 25
February 1(1.0) 61(61.6) 0(0.0) 21(21.2) 3(3.0) 13(13.1) 99
March 1(0.8) 70(58.3) 0(0.0) 32(26.7) 10(8.3) 7(5.8) 120
April 1(1.7) 17(28.3) 0(0.0) 16(26.7) 7(11.7) 19(31.7) 60
May 1(1.4) 48(69.6) 0(0.0) 9(13.0) 3(4.4) 8(11.6) 69
Total 5(0.9)*** 205(39.5)*** 1(0.2)*** 85(16.4) 84(16.2) 139(26.8) 519
*(F)=Females, *(M)=Males
***Indicate significant diference in indoor adult Anopheles populations at p<0.05
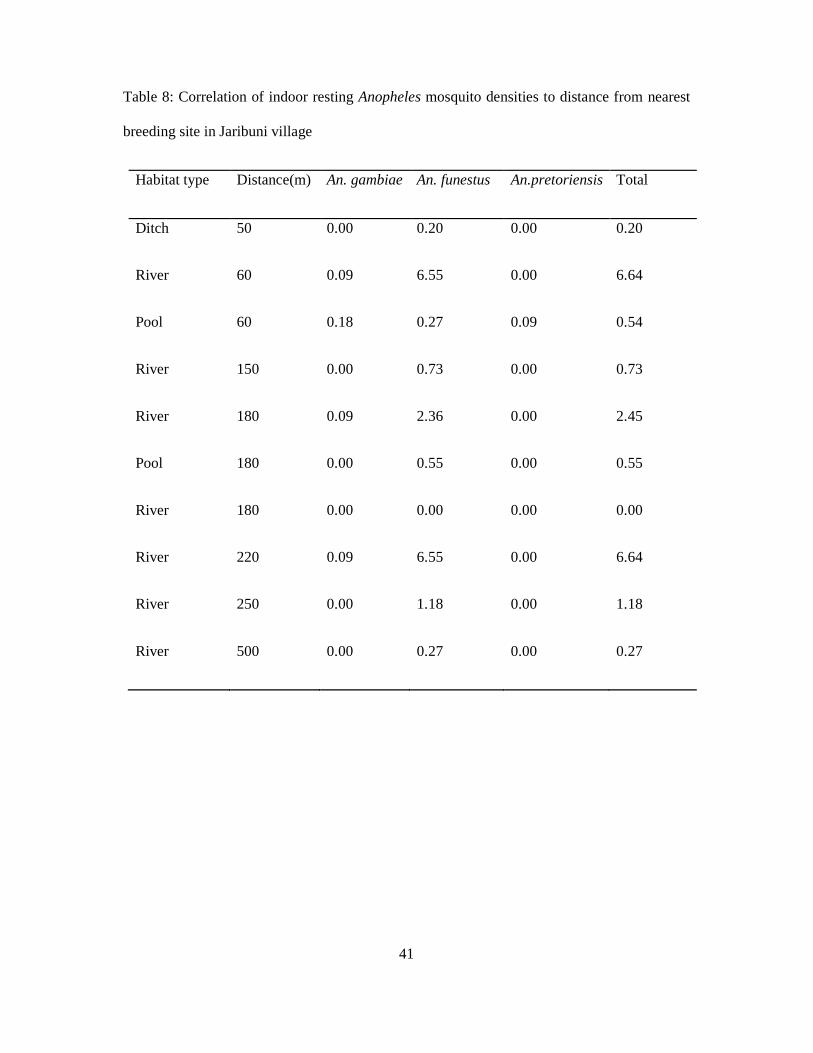
41
Table 8: Correlation of indoor resting Anopheles mosquito densities to distance from nearest
breeding site in Jaribuni village
Habitat type Distance(m) An. gambiae An. funestus An.pretoriensis Total
Ditch 50 0.00 0.20 0.00 0.20
River 60 0.09 6.55 0.00 6.64
Pool 60 0.18 0.27 0.09 0.54
River 150 0.00 0.73 0.00 0.73
River 180 0.09 2.36 0.00 2.45
Pool 180 0.00 0.55 0.00 0.55
River 180 0.00 0.00 0.00 0.00
River 220 0.09 6.55 0.00 6.64
River 250 0.00 1.18 0.00 1.18
River 500 0.00 0.27 0.00 0.27

42
3.9 ELISA for sporozoite rate and blood meal analysis
A total of 203 samples of indoor collected female Anopheles mosquitoes from Jaribuni village
were analyzed for sporozoites by ELISA technique (Table 9). The samples comprized of An.
funestus 199(98%), An. gambiae 3(1.5%) and An. pretoriensis 1(0.5%). The
circumsporozoites (CS) protein of Plasmodium falciparum was only detected in 10 specimens
of An. funestus (5.03%) mosquitoes, while an overall sporozoite rate was 4.93%.
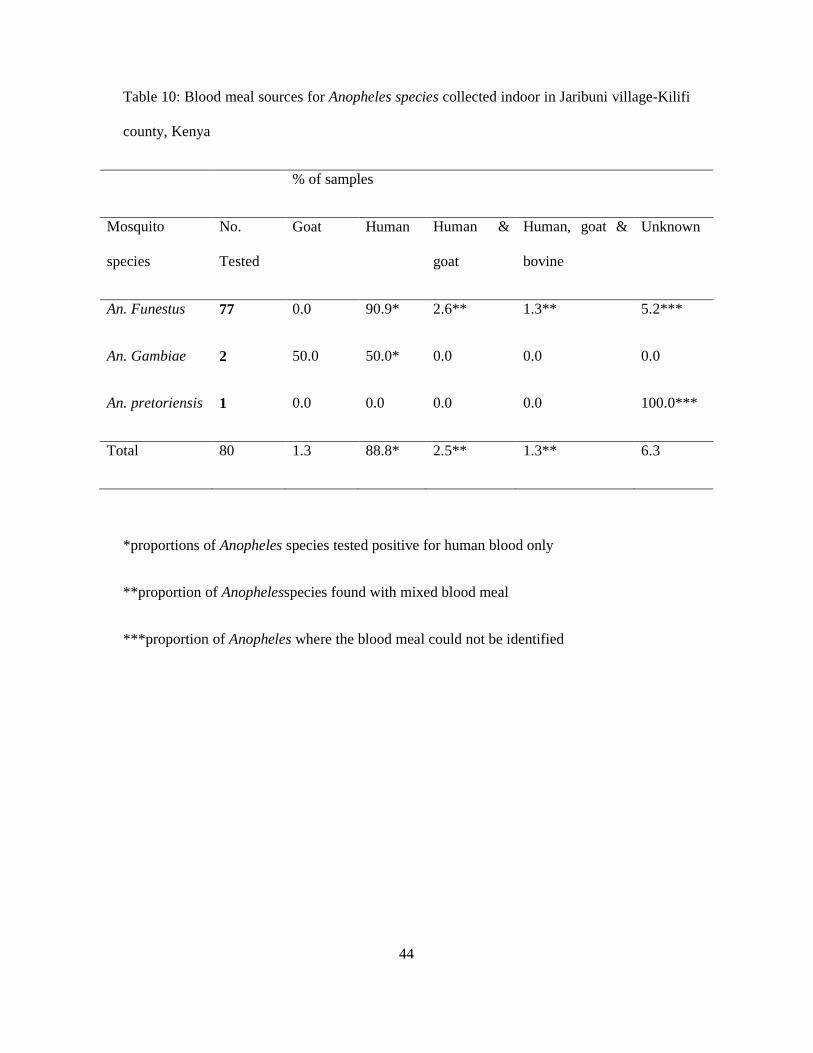
A total of 80 specimens were analyzed for blood meal sources among indoor collected
anopheline mosquitoes from Jaribuni village by ELISA (Table 10). The samples comprized of
77(96.2%) An. funestus, 2(2.5%) An. gambiae and only 1(1.3%) An. pretoriensis. The results
showed that, the overall preference of blood meal source for malaria vectors in the area was
humans (HBI) 0.89, followed by mixed blood sources human and goat (HGBI) 0.03, human,
goat and bovine (HGBBI) 0.01 then goat (GBI) 0.01 and 0.06 unknown. Table 10 shows the
human blood index (HBI) for An. funestus was high 0.91 (n=77) of the tested samples, while
mixed blood sources were 0.03 human and goat, and 0.01human, goat and bovine blood. The
human blood index (HBI) for An.gambiae was 0.5 (n=2) and Goat blood index (GBI) was 0.5.
A single sample of An. pretoriensis tested, the source of blood could not be detected.

43
Table 9: Plasmodium falciparum infection rate for indoor collected female Anopheles
mosquitoes from Jaribuni village Kilifi county (December 2010 to May 2011)
Species No. Tested No. Positive Sporozoite rate(%)
An. Funestus 199 10 5.03**
An. Gambiae 3 0 0.00
An. Pretoriensis 1 0 0.00
Total 203 10 4.93
** Proportion of infected Anopheles funestus species by sporozoites
44
Table 10: Blood meal sources for Anopheles species collected indoor in Jaribuni village-Kilifi
county, Kenya
% of samples
Mosquito
species
No.
Tested
Goat Human Human &
goat
Human, goat &
bovine
Unknown
An. Funestus 77 0.0 90.9* 2.6** 1.3** 5.2***
An. Gambiae 2 50.0 50.0* 0.0 0.0 0.0
An. pretoriensis 1 0.0 0.0 0.0 0.0 100.0***
Total 80 1.3 88.8* 2.5** 1.3** 6.3
*proportions of Anopheles species tested positive for human blood only
**proportion of Anophelesspecies found with mixed blood meal
***proportion of Anopheles where the blood meal could not be identified

45
4 CHAPTER FOUR: DISCUSSION, CONCLUSION AND
RECOMMENDATIONS
4.1 Discussion
Five major potential mosquito breeding habitats were identified in Jaribuni during the dry
period including river (stream), small stream pools along the river margins, manmade ditches
resulting from sand digging close to the river, abandoned water storage tanks and small
temporary trenches. Most of these man-made ditches and stream pools dried out during the
dry season while abandoned water tanks and trenches contained water in few occasions when
it rained. The river was the main source of water for stream pools and adjacent man-made
ditches. There was significant difference in Anopheles larval productivity between these
habitats, on temporal bases with the month of March showing higher larval productivity due
to rain showers during the month. It was evident that the highest Anopheles larval density was
taken from the river an observation attributed to its stability. The availability of optimal
mosquito vector species breeding habitat primarily influenced the species fecundity in a given
area (Imbahale et al., 2011).
This study found that anopheline and culicine larvae in naturally occurring habitats to co-
exist, similar to results of Mwangangi et al., (2007) indicating they are still important
breeding sites. Early instar densities for both anophelines and culicines were significantly
high in most of the sampling occasions in all types of breeding habitats compared to late
instars and pupae. This implied that, the gravid mosquitoes were attracted to oviposit on these
habitats but there was high larval mortality as suggested by the decline in populations of late
instars and pupae. The ability of the habitat to support larval survival and development to
complete life cycle was low in ditches and water tanks compared to the river and stream
pools. A combination of factors contribute to low survival rate of mosquitoes in a particular

46
habitat (Gimnig et al., 2002), which include high temperatures and low humidity. These
climatic factors may contribute to the drying up of small habitats, and thus making them less
suitable for mosquito breeding. Other factors that may limit the increase in larval populations
include overcrowding of larvae in a habitat leading to resources competition, physicochemical
factors imbalances and intra/inter-specific predation among mosquitoes (Carlson et al, 2004).
However, Minakawa et al (2005) observed that larval habitat stability was the most important
factor affecting survivorship of mosquito larvae. The hot dry season had resulted into several
small temporary ditches and pools drying out before the larva reached the adult stage.
Nevertheless, dry weather accelerated larval development in aquatic habitats; reduced river
water volume and velocity made the margins and stream pools with submerged vegetation
favorite breeding habitats for An. funestus in the area resulting into increased adult population.
Emerging adult mosquitoes monitored by emergent traps over natural occurring habitats
showed the river, stream pools and ditches were the most productive. An. gambiae s.l was the
predominant collected adult mosquito species among malaria vectors in the area but these
results were not comparable to those of indoor resting mosquito collection where An. funestus
was the predominant species. This difference might be caused by more than a few reasons,
including breeding preferences of these two anopheles species, number of cages placed over
the permanent breeding habitats against small temporary ones, and their feeding and resting
behaviours. Findings similar to this have been reported from the Gambia, West Africa by
Bogh et al., (2003), between two Anopheles species, An. melas and An. gambiae ss, and also
in Western Kenya between An. gambiae ss and An. arabiensis (Minakawa et al., 1999).
If this difference is attributable to the feeding and resting behaviour of the Anopheles species,
then most probably, majority of the collected An. gambiae s.l were An. arabiensis. In the
current study, further analysis of anophelines to sibling species could not be done but one

47
sample of An. gambie s.l. collected indoor was analyzed for blood meal by ELIZA and was
found with goat’s blood. This result indicated the existence of zoophagic An. gambiae
complex species in the area. An. arabiensis are zoophagic and exophilic mosquito (Mahande
et al., 2007) and they are normally predominant in dry, hot season (Githeko et al., 1996).
Moreover, it’s probable that cages were useful in reducing the sunlight intensity to the habitat.
This could have reduced water evaporation rate and prolonged wet time for An. gambiae s.l to
complete their development to adults in temporary pools and ditches compared to bare
habitats. There was continuous emergence of adult mosquitoes from caged habitats although
in low numbers throughout the experiment. This implied that there were mixed aquatic
mosquito stages confined in habitats by cages showing the difference in oviposition time
among the species.
Prior sampling was done for the presence and absence of aquatic stages before placing an
emergence cage for adult mosquito emergence. In three months of this experiment, ditches
were found to be the most productive habitats for adults over time. Due to the fact that they
were higher in number and more cages were placed on them. For instance, in March only
ditches were productive for anophelines and the higher numbers were recorded in the
following months of April and May. This experiment revealed that An. gambiae s.l.
mosquitoes were the most emerged malaria vector species in natural occurring habitats. The
emergent cages placed on river margins were less productive for adult mosquitoes. Probably,
this was due to predation but also in several occasions during this experiment, the river
flooded when it rained. When the river was flooded, the cages were pushed away and pre-
mature mosquitoes were drowned. Setting up emergent cages in flooded river was not
feasible. Consequently, less number of cages placed over the river resulted into less emerged
adult mosquitoes collected. Some few An. pretoriensis and one An. funestus were among

48
collected malaria vectors from ditches in the month of April. In comparison, the month of
April was the most productive for emerging adult mosquitoes from natural habitats to other
two months of experiment i.e. March and May. The few rains in April averaged 2.13mm and
atmospheric temperature 32.5oC improved the habitat stability and suitability for the aquatic
stage mosquito developments to adults. Other physicochemical factors such as salinity,
dissolved oxygen and conductivity were in the range favoring Anopheles production in the
habitats.
Body size variation of the Anopheles results showed that, An. gambiae s.l that emerged from
ditches and pools had mean body sizes of 3.08mm and 3.17mm respectively. This implied that
they were efficient malaria vectors since body size of 3mm or above has been shown to
enhance the vectorial capacity of mosquitoes (Lyimo and Takken, 1993). Anopheles gambie
from the river were smaller with mean body size of 2.96mm these have less chances of
survival and are thus less efficient vectors. Body size is an important feature in mosquito
fitness because it is associated with longevity, survival, blood meal capacity (Kelly and
Edman, 1992), and number of eggs per single oviposition (Woke, et al.,, 1956), all of which
influence the vectorial capacity. Well grown mosquitoes of 3mm or above have higher chance
of living longer than smaller mosquitoes. Vector longevity provides enough time for a
parasite they harbour to develop up to its infective stage (Howley, 1985). A fully grown
female Anopheles mosquito needs a single blood meal for each gonotrophic cycle (Lyimo and
Takken, 1993). This has an advantage of reducing the risk of being killed by the vertebrate
host out of repeated feeding visits. Schwartz and Koella, (2001) noted that “biting is risky, the
mosquito mortality increases with its biting rate”. A mosquito is capable of producing many
eggs in a single batch, this reproductive success enables a single mosquito to easily maintain
its colony in the locality and sustain disease transmission. The same fitness applies to males in

49
terms of mating success and mating frequencies (Takken et al., 2006). The vectorial capacity
of the mosquito is greatly influenced by combination of these factors. In all the samples,
sexual dimorphism was evident as exemplified by the differences in size for both female and
male in all the species
Artificial habitats placed at 0, 25, 50, 75 and 100m distances from human habitations to
monitor anopheline breeding preferences, showed that, An. gambiae was the only malaria
vector that oviposited although the area has higher population of An. funestus. This was due to
the fact that An. gambiae prefer breeding in small temporary water bodies subjected to direct
sunlight (Gillies and Coetzee, 1987), as was the artificial habitats, although they tend to adapt
in different new environments (Filliger et al., 2004). An. funestus did not oviposit in artificial
habitats because their preference is large permanent shaded water bodies with vegetation
(Gillies and DeMeilon, 1968) like the Jaribuni River. Higher densities of Anopheles larvae
were observed in habitats located at 50m distances from human houses in different occasions,
compared to only once in 0, 75, and 100m distances. Culicine larvae were widely distributed
in all habitat locations for the whole period of experiment because they are non-selective
breeders. There were no seasonal variations in productivity observed during four months of
experiment, although mosquito populations were low because the area was hot and dry.
The closeness of An. gambiae to human habitations had also been observed in indoor resting
mosquito collections, relatively high proportions were collected in houses located between
60m and 120m from the nearest breeding habitats. This implied that spatial distribution of An.
gambiae breeding habitats is associated with distribution of human habitations. Habitat
specificity was also observed in indoor resting collections, peak densities of An. funestus were
collected from houses located near to the river while An. gambiae from houses near to the
ditches and pools. Generally, the results showed that there was a decrease in indoor Anopheles

50
mosquito population with an increase in distance from the nearest breeding habitat, suggesting
that malaria vectors in the study area bred proximal to human habitations. The mark-release-
capture studies conducted in the same area by Midega et al., (2007), showed that house
location and distance did not correlate with aquatic habitats of Anopheles mosquitoes. The
results in the current study show that during the extensive dry season in Jaribuni village, the
river and adjacent stream pools and ditches were the main reliable breeding habitats
throughout the year and An. funestus was the predominant species. These results comply with
the study by Mbogo et al., (2003); Braginets et al., (2003) along the coast of Kenya. It is said
that An. funestus is the “bridging species” between the dry and wet season (Mbogo et al.,
2003), especially in the settings like Jaribuni village having a permanent river which is a
stable habitat all year round. Such vector population dynamical outcomes sustain malaria
transmission.
The ELISA analysis for malaria vectors gave a 4.93% infection rate of mosquitoes infected
with circumsporozoites (CS) protein of P. falciparum, which probably correlate with high
infectiousness of human population in the area. From all malaria vector species collected
resting indoors; only An. funestus mosquitoes were found infected with parasite by 5.03%.
This has higher implication in malaria transmission dynamics in the locality, since the main
reliable breeding source in the dry season is the river crossing the village and adjacent pools
and ditches which have shown to be very productive for An. funestus. Although this study
found high population of An. gambiae s.l and few An. pretoriensis from the aquatic habitats,
low populations of these two species were found indoors and none of them were found
infected with circumsporozoite protein of P. falciparum. This however, does not propose that
An. gambiae and An. pretoriensis are not playing significant role in malaria transmission in
the study area. The social and cultural behaviour of the hosts and host preferences of the

51
mosquito species can drive into host selection and eventually influence the prevalence of the
disease in a locality (Burkot, 1988). The study conducted in southern Ethiopia on An.
arabiensis feeding behaviour (Tirados, et al., 2006) found that, in the areas where humans
were more readily available, the mosquitoes fed on humans, in the situation where animals
were accessed easier, they fed upon them.
The results of the blood meal analyses confirmed high anthropophagic, endophilic behaviour
of An. funestus. Their specific human blood index (HBI) was high 0.90, concomitant with
their established disposition as main malaria vectors in tropical Africa. Results similar to
these on sporozoite rate and blood meal tests for An. funestus in the area have been reported
by Mbogo et al (2003). On the other hand, these results suggest that residents in the village
are facing a high malaria risk although most of the houses (80%) sampled indoor resting
mosquitoes had bed nets. These results suggest more research on biting behaviour of
mosquitoes as well as sleeping behavior of the people in the area. The mixed blood meals
identified in 3 samples of An. funestus, possibly was due to the failure of the mosquitoes to
complete their gonotrophic circle by only one blood meal. The observed mixed blood meals
from two and three hosts can also be considered to these mosquito species, as adaptable to
feeding from any available host. This multiple hosts feeding behavior of mosquitoes may
facilitate the transmission of zoonotic diseases, such as West Nile fever and Rift Valley fever
as was alerted by Muriu et al., (2008) in a study conducted in Mwea rice scheme, where the
multiple hosts feeding was very high among collected mosquito species. A sample of An.
gambiae s.l collected resting indoor and tested positive for goat’s blood only, evidenced that
there were some mosquitoes in the area feeding outside but enter the house to rest or they
were still seeking more blood meal from human for eggs development.

52
4.2 Conclusion
This study revealed that mosquito breeding habitats are diverse in Jaribuni village but river
and its adjoining pools provided a stable source of mosquitoes. Mosquitoes from more
permanent habitats were generally small in size. Temperature, salinity, conductivity,
dissolved oxygen and distance from human habitations were found to correlate with malaria
vector production in the habitats. The spatial distribution of aquatic habitats correlates with
human host seeking mosquito indoor densities. Malaria vectors colonized habitats proximal to
the residential houses (250m). An. funestus is the predominant malaria vector during hot dry
season in Jaribuni village given its high density indoors and in permanent breeding habitats,
higher proportion of anthropophagic behaviour and high proportion of sporozoite rate.
4.3 Recommendations
Vector populations emerging from our experimental habitats did not match the vector
populations from house catches. Therefore, further research needs to be done with the
objective of ascertaining outdoor-resting malaria vector populations and their feeding
behaviours as regards the potential of malaria transmission. Further work should be done to
establish An. gambiae s.l sibling species dispersion patterns from source, host seeking and
resting behaviours. Environmental management for malaria vector control should be adopted
as a key interventional method alongside existing ones for reduction of local vector
populations in the area of Jaribuni, in Kilifi County at the coast of Kenya.

53
REFERENCES
Becker, N., 1998, The use of Bacillus thuringiensis israeliensis against mosquitoes, with
special emphasis on the ecological impact. Israel Journal of Entomology, 32:63-69.
Beier J., Keating, J., Githure, J., Macdonald, M., Empoinvil, D. and Novak, R., 2008,
Integrated vector management for malaria control. Malaria Journal, 7(suppl.1):S4:1475-
2875-7.
Beier, J., Killeen, G. and Githure, J., 1999, Short report: Entomologic Inoculation Rates and
Plasmodium falciparum Malaria Prevalence in Africa. American Journal of Tropical
Medicine and Hygiene, 61(1): 109-113.
Bogh, C., Clarke, S., Jawara, M., Thomas C. and Lindsay, S., 2003, Localized breeding of the
Anopheles gambiae complex (Diptera: Culicidae). Bulletin of Entomological Research,
93:279-287.
Bond, J., Arredondo-Jimenez, J., Rodriguez, M., Martinez, H. and Williams, T., 2005,
Oviposition habitat selection for predator refuge and food source in a mosquito.
Ecological entomology, 30: 255-263.
Braginets, O., Minakawa, N., Mbogo, C., and Yan, G., 2003, Population genetic structure of
the Africa malaria mosquito Anopheles funestus in Kenya. American Journal of Tropical
Medicine and Hygiene, 69(3), 303-308.
Brooke, B., Hunt, R., Chandre, F., Carnevale, P. and Coetzee, M., 2002, Stable chromosomal
inversion polymorphisms and insecticide resistance in the malaria vector mosquito
Anopheles gambiae (Diptera: Culicidae). Journal of Medical Entomology, 39(4):568-573.

54
Burkot, T., 1988, Non-random host selection by Anopheles mosquitoes; Tropical health
program Queensland institute of medical research, Brastone Terrace, Herstone Brisbane-
Queensland 4006 Australia. Parasitology Today, 4(6):156-62.
Carlson, J., Byrd, B. and Omlio, F., 2004, Field assessment in Western Kenya link malaria
vectors to environmentally disturbed habitats during the dry season. BMC public Health,
4:33.
Carter, R., Mendis, K. and Roberts, D., 2000, Spatial targeting of intervention against malaria.
Bulletin of the World Health Organization, 78(12).
Castro, C., Kanamori, S., Kannady, K., Mkude S., Killeen, G. and Fillinger, U. 2010, The
importance of drains for larval development of Lymphatic filariasis and malaria vectors
in Dar es Salaam, United Republic of Tanzania. PloS Neglected Tropical Diseases, 4(5):
e693.
CDC, 2010, Malaria; Anopheles mosquitoes; Global Health-Division of Parasitic diseases.
Chandler, A., and Read, C., 1961, Introduction to Parasitology, New York: London, John
Wiley and Sons, Incl.
Chaves, L., Harrington, L., Keogh, C., Nguyen, A., and Kitron, U., 2010, Blood feeding
patterns of mosquitoes; random or structured? Frontiers in Zoology, 7:3.
Coetzee, M., 2004, Distribution of the African malaria vectors of the Anopheles gambiae
complex. American Journal of Tropical Medicine and Hygiene, 70(2):103-104.
Coetzee, M., Craig, M., and Le Sueur, D., 2000, Distribution of African malaria mosquitoes
belonging to the gambiae complex, Parasitology Today, 16(2): 74-77.

55
Devine, G., and Killeen, G., 2010, The potential of a new larviciding method for the control
of malaria vectors. Malaria Journal, 9:142.
Diabate, A., Dabire, K., Kim E., Dalton, R., Milongo, D., Baldet, T., Simard F., Gimnig, J.,
Hawley, W. and Lehman, T., 2005, Larval development of the molecular forms of
Anopheles gambiae (Diptera: Culicidae) in different habitats a transplantation
experiment. Journal of Medical Entomology, 42:548-553.
Ferguson, H., Dornhaus, A., Beeche, A., Borgemeister, C., Gottlieb, M., Mulla, M., Gimnig
J., Fish, D. and Killeen G., 2010, Ecology: A prerequisite for malaria elimination and
eradication. PloS Medicine, 7(8): e1000303.
Fillinger, U., Bart, G., Knols, and Becker, N., 2003, Efficacy and efficiency of new Bacillus
thuringiensis var. israeliensis and Bacillus sphericus formulations against Afrotropical
Anophelines in Western Kenya; Tropical Medicine and International Health, 8(1): 37-47.
Fillinger, U., Kannady, Kh., William, G., Venak, M., Dongus, S., Nyika, D., Geissbuhler, Y.,
Chaki, P., Govella, N., Mathenge, E., Singer, B., Mshinda, H., Lindsay, S., Tanner, M.,
Mtasiwa, D., Castro, M. and Killeen, G., 2008, A tool box for operational mosquito larval
control: preliminary results and early lessons from Urban Malaria Control Programme in
Dar es Salaam Tanzania. Malaria Journal, 7: 20.
Fillinger, U., Sonye, G., Killeen, G., Knols, B., and Becker, N., 2004, The practical
importance of permanent and semi-permanent habitats for controlling aquatic stages of
Anopheles gambiae sensu lato mosquitoes: operational observation from a rural town in
Western Kenya. Tropical Medicine and International Health, 9(12): 1274-1289.
Gates Malaria Partnership Report, 2001-2006, London School of Hygiene and Tropical
Medicine.

56
Geissbuhler, Y., Chaki, P., Emidi, B., Govella, N., Shirima, R., Mayagaya, V., Mtasiwa, D.,
Mshinda, H., Fillinger, U., Lindsay, S., Kannady, K., DeCastro, M., Tanner, M. and
Killeen, G., 2007, Interdependence of domestic malaria prevention measures and
mosquito-human interactions in Urban Dar es salaam, Tanzania. Malaria Journal 6:126.
Gillies, M. and Coetzee, M., 1987, A supplement to the Anophelinae of Africa South of the
Sahara (Afro-tropical Region), Publications of the South African Institute for Medical
Research 55: 1-143.
Gillies, M. and De Meilon, B. 1968, The Anophelinae of Africa, South of the Sahara
(Ethiopian Zoogeographical region), Publication 54, South Africa Institute for Medical
Research, Johannesburg, South Africa.
Gimnig, J., Ombok, M., Kamau, L. and Havlett, W., 2001, Characteristics of larval Anopheles
(Diptera: Culicidae) habitats in Western Kenya. Journal of Medical Entomology, 38:282-
288.
Gimnig, J., Ombok, M., Otieno, S., Kaufman, M., Vulule, J., and Walker, C., 2002, Density
dependant development of Anopheles gambiae (Diptera: Culicidae) larvae in artificial
habitats. Journal of Medical Entomology, 39:162-172.
Githeko, A., Adungo, N., Karanja, D., Hawley, H., Vuvule, J., Seroney, I., Ofulla, A., Atieli,
F., Ondejo, S., Genga, I., Odaga, K., Situbi, P., and Oloo, J., (1996), Some observations
on biting behaviour of Anopheles gambiae s.s, Anopheles arabiensis, and Anopheles
funestus and their implications for malaria control. Experimental parasitology 82(3):
306-15.

57
Gu, W. and Novak, R., 2009, Agent-based modeling of mosquito foraging behavior for
malaria control. Trans Research Society, American Journal of Tropical Medicine and
Hygiene, 103(11): 1105.
Gu, W., Regens, J., Beier , J. and Novac, R., 2006, Source reduction of mosquito larval
habitats has unexpected consequences on malaria transmission. Proc National Academy
of Sciences, USA. 14, 103(46): 17560-17663.
Gu, W., Utzinger, J. and Novak, R., 2008, Habitat-based larval interventions, A new
perspective for malaria control, American Journal of Tropical Medicine and Hygiene,
78(1): 2-6.
Hardin, J. and Jackson F., 2009, Application of natural products in the control of mosquito-
transmitted diseases, African Journal of Biotechnology, 8(25): 7373-7378.
Howley, W., 1985, The effect of larval density on adult longevity of a mosquito, Aedes
sierrensis. Epidemiological consequences. Journal of Animal Ecology, 54:955-964.
Imbahale, S., Paaijmans, K., Mukabana, W., Lammeren, R. and Githeko, A., 2011, A
longitudinal study on Anopheles mosquito larval abundance in district geographical and
environmental settings in western Kenya. Malaria Journal, 10:81doi 10.1186/1475-2875.
Kakkilaya, B., 2008, Malaria history, Malaria website. www.malariasite.com.
Kelly, R. and Edman, J., 1992, Mosquito size and multiple transmission of avian malaria in
the laboratory. Journal of American Mosquito Control Association, 8:386-388.
Kenya Ministry of Public Health and Sanitation, 2007, Malaria indicator survey, Division of
Malaria Control.

58
Kenya Ministry of Public Health and Sanitation, 2009, National malaria strategy 2009-2017
Division of Malaria Control.
Kigadye, E., Nkwengulila, G., Magesa, S. and Abdullah, S., 2010, Diversity, Spatial and
temporal abundance of Anopheles gambiae complex in the Rufiji River, Southern
Tanzania. Tanzania Journal of Health Research, 12(1):68-72.
Killeen, G., Fillinger, U. and Knols, B., 2002, Advantages of larval control for African
malaria vectors: low mobility and behavioral responsiveness of immature mosquito
stages allow high effective coverage. Malaria Journal, 1:8.
Killeen, G., McKenzie, F., Foy, B., Bogh, C. and Beier, J., 2001, The Availability of Potential
Hosts as a Determinant of Feeding Behaviors and Malaria Transmission by African
Mosquito population. Trans Research Society of Tropical Medicine and Hygiene, 95(5):
469-476.
Killeen, G., Seyoum, A. and Knols, B., 2004, Rationalizing historical success of malaria
control in Africa in terms of mosquito resource availability management. American
Journal of Tropical Medicine and Hygiene, 71(2) pp. 87-93.
Kiszewski, A. and Teklehaimanot, A., 2004, A review of the chemical and epidemiologic
burden of epidemic malaria. American Journal of Tropical Medicine and Hygiene, 71(2):
128-135.
Le Menach, A., McKenzie, F., Flahault, A. and Smith, D., 2005, The unexpected importance
of mosquito oviposition behaviour for malaria: non-productive larval habitats can be
sources for malaria transmission. Malaria Journal, 4:23.

59
Lymo, E. and Takken, W., 1993, Effect of adult body size on facundity and the pre-gravid
rate of Anopheles gambiae females in Tanzania. Medical and Veterinary Entomology, 7:
328-332.
Mahande, E., Mosha, F., Mahande, J. and Kweka, E., 2007, Feeding and resting behaviour of
malaria vector, Anopheles arabiensis with reference to zooprophylaxis. Malaria Journal,
6:100.
Mboera, L., 2005, Sampling techniques for adult malaria vectors and their reliability in the
estimation of entomological inoculation rate. Tanzania Health Research bulletin, 7(3).
Mboera, L., Magesa, S., and Molten, F., 2006, Indoor man biting mosquitoes and their
implication on malaria transmission in Mpwapwa and Iringa District, Tanzania; Tanzania
Health Research Bulletin, 8(3): 141-144.
Mbogo, C., Snow, R., Khamala, C., Kabiru, E., Ouma, J., Githure, J., Marsh, K. and Beier, J.,
1995, Relationship between Plasmodium falciparum transmission by vector populations
and the incidence of severe disease at nine sites on the Kenyan Coast. American Journal
of Tropical Medicine and Hygiene, 52(3): 201-206.
Mbogo, C., Mwangangi, J., Nzovu, J., Gu, W., Yan, G., Günter, J., Swalm, C., Keating, J.,
Regens, J., Shililu, J., Githure, J., and Beier, J., 2003, Spatial and temporal heterogeneity
of Anopheles mosquitoes and Plasmodium falciparum transmission along the Kenyan
Coast. American Journal of Tropical Medicine and Hygiene, 68 (6): 734-742.
Midega J., Mbogo, C., Mwambi, H., Wilson, M., Ojwang, G., Mwangangi, J., Nzovu, J.,
Githure, J., Yan, G., and Beier, J., 2007, Estimating dispersal and survival of Anopheles
gambiae and Anopheles funestus along the Kenyan Coast by using Mark-release-
recapture methods. Journal of Medical Entomology, 44 (6):923-929.

60
Minakawa, N., Mutero, C., Githure, J., Beier, J., and Yan, G., 1999, Spatial distribution and
habitat characterization of anopheline mosquito larvae in Western Kenya. American
Journal of Tropical Medicine and Hygiene, 61(6):1010-1016.
Minakawa, N., Sonye, G. and Yan, G., 2005, Relationships between occurrence of Anopheles
gambiae s.l (Diptera: Culicidae) and size and stability of larval habitats. Journal of
Medical Entomology, 42:295-300.
Mokay, A., and Shine, R., 2003, Oviposition site selection by mosquitoes is affected by cues
from conspecific larvae and anuran tadpoles. Australia Ecology, 28: 33-37.
Munga, S., Minakawa, N., Zhou, G., Mushinzimana, E., Barrack, O., Githeko, A., and Yan,
G., 2006, Association between land cover and habitat productivity of malaria vectors in
Western Kenya Highlands. American Journal of Tropical Medicine and Hygiene, 74(1):
69-75.
Munga, S., Yakub, L., Mushinzimana, E., Zhou, G., Ouna, T., Minakawa, N., Githeko, A. and
Yan, G., 2009, Land use and land cover changes and spatio-temporal dynamics of
Anopheline larval habitats during a four-year period in a high land community of Africa.
American Journal of Tropical Medicine and Hygiene, 81(6): 1079-1084.
Muriu, S., Muturi, E., Shililu, J., Mbogo, C., Mwangangi, J., Jacob, B., Irungu, L., Mukabana,
R., Githure, J., and Novak, R., 2008, Host choice and multiple blood feeding behavior of
malaria vectors and other Anophelines in Mwea rice scheme, Kenya. Malaria Journal,
7:43.
Mutuku, F., Alaii, A., Bayoo M., Gimnig, J., Vulule, J., Walker, E., Kabiru, E. and Hawley
W., 2006, Distribution, Description and local knowledge of larval habitats of Anopheles

61
gambiae s.l. in a village in Western Kenya. American Journal of Tropical Medicine and
Hygiene, 74(1): 44-53.
Muturi, E., Burges, P. and Novak, R., 2008, Malaria vector management: Where have we
come from and where are we headed? American Journal of Tropical Medicine and
Hygiene, 78(4): 536-537.
Mwangangi, J., Mbogo, C., Muturi, E., Nzovu, J., Githure, J., Yan, G., Minakawa, N., Novak,
R. and Beier, J., 2007, Spatial distribution and habitat characterization of Anopheles
larvae along the Kenyan Coast. Journal of Vector Borne Diseases 44: 44-45.
Oyewole, O., Momoh, O., Anyasor, G., Ogunnowo, A., Ibidapo, C., Oduola, O., Obansa, J.
and Awolola, T., 2009, Physico-chemical characteristics of Anopheles breeding sites:
Impact on fecundity and progeny development. African Journal Environmental Science
and Technology, 3 (12): 447-452.
Paaijmans K., 2008, Weather, water and malaria mosquito larvae: The impact of
meteorological factors on water temperature and larvae of the Afro-Tropical malaria
vector Anopheles gambiae Giles. PhD Thesis, Wageningen University.
Pates, H. and Curtis, C., 2005, Mosquito behavior and vector control Annual Review of
Entomology 50: 53-70.
RBM, 2003, The burden of malaria in Africa. The African malaria report, Chapter 1, World
Health Organization
Robert, V., Awono-Ambene, H. and Thioulouse, J., 1998), Ecology of larval mosquitoes with
special reference to Anopheles arabiencis (Diptera: Culicidae) in Market-Garden wells in
Urban Darkar, Senegal. Journal of Medical Entomology, 35(6):948-955.

62
Rugemalila, J., Wanga, C. and Kilama, W., 2006, Sixth Africa malaria day in 2006: how far
have we come after the Abuja Declaration? Malaria Journal, 5:102.
Schwartz, A. and Koela, J., 2001, Trade-offs, conflicts of interest and manipulation in
Plasmodium-mosquito interactions. TRENDS in Parasitology Volume 17 No.4.
Service M. 1993, Mosquito ecology. Field sampling methods, Second Edition, Chapter 3.
Service, M., 1977, Mortalities of the immature stages of species B of the Anopheles gambiae
complex in Kenya: Comparison between rice fields and temporary pools, identification of
predators, and effect of insecticidal spraying. Journal of Medical Entomology, 13: 535-
545.
Shililu, J., Ghebremeskel, T., Seulu, F., Mengistu, S., Fekadu, H., Zerom, M.,
Ghebrgziabiher, A., Sintasath, D., Bretas, G., Mbogo, C., Githure, J., Brantly, E., Novak,
R. and Beier, J., 2003, Larval habitat diversity and ecology of Anopheles larvae in
Eritrea, Journal of Medical Entomology, 40:921-929.
Shillcutt, S., Morel, C., Goodman, C., Coleman, P., Bell, D., Whitty, C. and Mills, A., 2008,
Cost effectiveness of malaria diagnostic methods in Sub-Saharan Africa in an era of
combination therapy. Bulletin of the World Health Organization, 86(2): 81-161.
Snow, W., 1983, Mosquito production and species succession from an area of irrigated rice
fields in The Gambia, West Africa. American Journal of Tropical Medicine and Hygiene,
86:237:245.
Sumba, L., Guda, T., Deng, A., Hassanali, A., Beier, J., and Knols, B., 2004, Mediation of
Oviposition site selection in African malaria mosquito Anopheles gambiae (Diptera:
Culicidae). International Journal of Tropical Insect Science, 24(3): 260-265.

63
Takken, W. and Knols, B., 2007, Novel mosquito control strategies in the fight against
malaria. Bulletin of the Netherlands Society for Tropical Medicine and International
Health, 46:4.
Takken, W. and Lindsay, S., 2006, Factors affecting the vectorial competence of Anopheles
gambiae. A question of scale. Wageningen UR Fronts series, volume 2 Ecological
Aspects for Application of Genetically Modified Mosquitoes.
Takken, W., Constantine, C., Dolo, C., Hassanali, A., Sagmon, N. and Osir, E., 2006,
Bridging laboratory & field research for genetic control of disease vectors. Chapter 17-
Mosquito mating behaviour, part 4:183-188.
Tirados, I., Costantini, C., Gibson, G. and Torr, S., 2006, Blood feeding behaviour of malaria
mosquito Anopheles arabiensis: implication for malaria control. Medical Veterinary
Entomology, 20(4): 425-37.
Tsila, H., Messi, J. and Dodji G., 2010, Adaptive responses of Anopheles gambiae in
crowding larvae conditions in laboratory. Asian Journal for Biological Sciences, 4:259-
265.
Uneke, C., 2009, Deforestation and malaria in Sub-Sahara Africa: an overview. International
Journal for Tropical Medicine, 6: (1).
USAID, 2010, Kenya malaria operational plan (MOP). President malaria initiative (PMI).
White, G., 1974, Anopheles gambiae complex and disease transmission in Africa. Trans
Research Society for Tropical Medicine and Hygiene, 68: 278-301.
WHO, 2004, A strategic framework for malaria prevention and control during pregnancy in
the African region. World Health Organization, Regional offices for Africa.

64
WHO, 2006, Malaria vector control and personal protection. Technical report series 936,
Report of World Health Organization-study group.
WHO, 2008, The world malaria report, World Health Organization.
Woke, P., Ally, M. & Rosenberger, C., 1956, The number of eggs developed related to the
quantities of human blood ingested in Aedes aegypti. Ann Entomological Society of
America 49:435-441.





























