Embed Size (px)
Citation preview

VERNON FRENCH AND GRAEME DANIELS
The beginning and the end of insect limbsRecent results shed light on the mechanisms underlying
pattern formation in the development of Drosophila imaginaldiscs, which give rise to the appendages of the adult fly.
Central to the development of an organism are themechanisms of pattern formation that ensure that cellsform structures appropriate to their location. Thesemechanisms have been studied in a number of organ-isms in many different ways, and over the last decademolecular genetic analyses have provided an unparal-leled understanding of patterning in the Drosophilaearly embryo. We know (most of) the genes involved,their patterns of expression and, to some extent, weunderstand their controls and the functions of theirproducts. However, light shines brightly on only thefirst few hours of Drosophila development and similar,molecular genetic analysis of later processes - such asthe development of the major adult appendages, thelegs and wings - is only just beginning. In this article,we discuss some recent work on the development ofappendages, including studies of two genes, Distal-less(Dll) [1] and aristaless (ab [2], which are expressed atthe presumptive distal end of the appendage and whichmay function in patterning the proximal-distal axis.
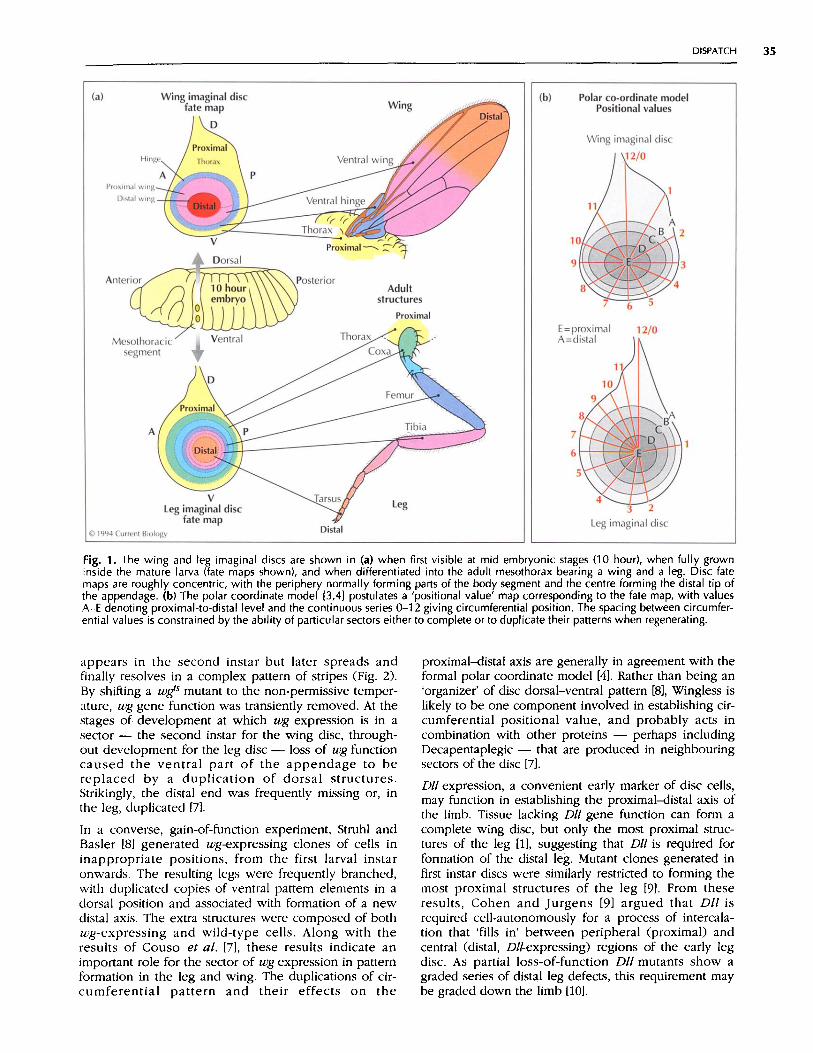
Drosophila appendages are formed by imaginal discs,specialized clusters of cells that derive from a small partof certain embryonic body segments. The discs invagi-nate and then grow inside the developing larva, finallydifferentiating into adult structures at metamorphosis.Until recently, pattern formation in discs was studiedmainly by analysing their regeneration after surgery.The results of these experiments contributed to the for-mulation of the 'polar coordinate' model [3], which pro-poses that the mature imaginal disc has a map ofcircumferential and radial sets of 'positional values',ordered around and down the future appendage,respectively (Fig. 1). After damage, short-range cell-cellinteractions result in intercalary regeneration to remove,by the shortest possible route, any discontinuity in posi-tional value between adjacent cells. In addition, themodel links the two sets of coordinates, by proposingthat distal ends are regenerated whenever all or most ofthe circumferential values are present in a group ofadjacent cells [3,4]. The model provides a formal expla-nation of regeneration - although not a perfect one -and suggests that similar cell-cell interactions patternthe disc during normal development [4]. With the toolsnow available, we hope to understand the positionalproperties of the disc cells and the mechanisms of theirinteraction at the molecular level.
The leg and wing imaginal discs each become morpho-logically distinct as separate groups of about 20 cellswithin the embryonic thoracic segments, about 10hours after egg deposition [5], but it is likely that thediscs originate at a much earlier stage. The localized
expression of two genes, wingless (wg) and decapenta-plegic (dpp) is important in patterning the anterior-pos-terior and dorsal-ventral axes of the early segment and,probably, in initiating disc formation. These two genesencode, respectively, members of the WNT (winglessand int) and transforming growth factor-13 (TGF-B)families of secreted signalling molecules.
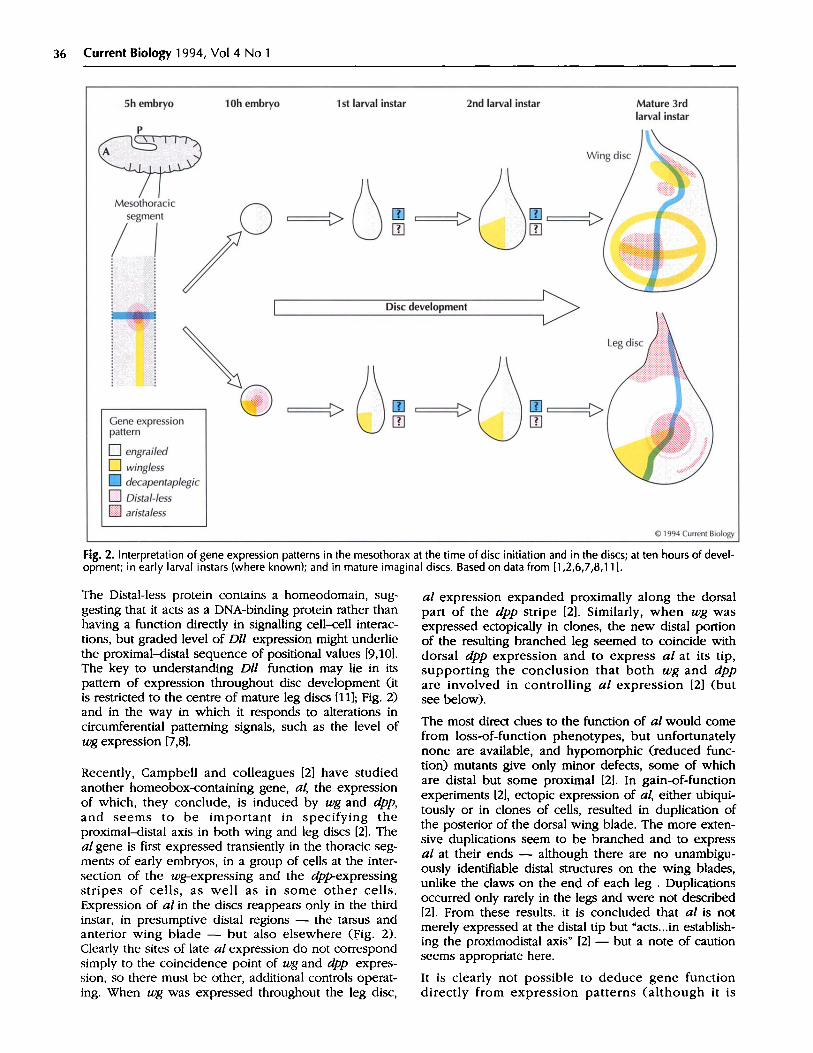
Around five hours after egg deposition, the segmentpolarity gene wg is expressed in an interrupted stripeacross each segment; the expression of dpp has by thistime resolved into longitudinal stripes down theembryo. Cohen and co-workers [1,6] have shown thatDll is expressed in each thoracic segment from aboutfive hours of development, in a group of cells aroundthe intersection of the wg and dpp expression stripes(Fig. 2). Dll gene expression marks the discprimordium, which may be specified directly by thecoincidence of Wingless and Decapentaplegic proteins,both of which are secreted and so spread a shortdistance from the cells producing them.
There is good evidence for control of disc formation bywg, as Dll expression and disc formation are bothabsent in wg-null mutant embryos, and, in embryosmutant for other segment polarity genes, both Dllexpression and disc formation are shifted to correspondto the altered sites of wg expression [6]. The relation-ship between dpp expression and disc formation is sug-gestive, but as yet there is little direct evidence.Intriguingly, the initial pattern of Dll expression seemsto characterize a common primordium of the wing andleg imaginal discs. Some of these cells then switch offDll and are displaced dorsally at about 10 hours ofdevelopment to form the wing disc, while the otherscontinue Dllexpression and form the leg disc [1].
It is becoming clear that, in addition to their roles inpatterning the primary axes of the early embryo,Wingless and Decapentaplegic proteins mediate othershort-range cell-cell interactions later in development:indeed, the genes encoding both were first known andnamed for the effects of adult-viable mutations causingdefects in the appendages, particularly the absence ofdistal parts.
In a recent study, Couso et al. [7] have traced wgexpression throughout disc development. Initially, wgexpression is found in a quadrant of the leg disc but isabsent from the wing disc. The leg disc pattern remainsapproximately constant, so wg continues to beexpressed in an anterior-ventral sector of the mature,third larval instar leg disc (Fig. 2). In the wing disc,however, an anterior-ventral sector of expression
© Current Biology 1994, Vol 4 No 134
PATTERN FORMATION
DISPATCH 35
Fig. 1. The wing and leg imaginal discs are shown in (a) when first visible at mid embryonic stages (10 hour), when fully growninside the mature larva (fate maps shown), and when differentiated into the adult mesothorax bearing a wing and a leg. Disc fatemaps are roughly concentric, with the periphery normally forming parts of the body segment and the centre forming the distal tip ofthe appendage. (b) The polar coordinate model 3,4] postulates a 'positional value' map corresponding to the fate map, with valuesA-E denoting proximal-to-distal level and the continuous series 0-12 giving circumferential position. The spacing between circumfer-ential values is constrained by the ability of particular sectors either to complete or to duplicate their patterns when regenerating.
appears in the second instar but later spreads andfinally resolves in a complex pattern of stripes (Fig. 2).By shifting a wgs mutant to the non-permissive temper-ature, wg gene function was transiently removed. At thestages of development at which wg expression is in asector - the second instar for the wing disc, through-out development for the leg disc - loss of wg functioncaused the ventral part of the appendage to bereplaced by a duplication of dorsal structures.Strikingly, the distal end was frequently missing or, inthe leg, duplicated [7].
In a converse, gain-of-function experiment, Struhl andBasler [81 generated wg-expressing clones of cells ininappropriate positions, from the first larval instaronwards. The resulting legs were frequently branched,with duplicated copies of ventral pattern elements in adorsal position and associated with formation of a newdistal axis. The extra structures were composed of bothwg-expressing and wild-type cells. Along with theresults of Couso et al. [7], these results indicate animportant role for the sector of wg expression in patternformation in the leg and wing. The duplications of cir-cumferential pattern and their effects on the
proximal-distal axis are generally in agreement with theformal polar coordinate model [4]. Rather than being an'organizer' of disc dorsal-ventral pattern [8], Wingless islikely to be one component involved in establishing cir-cumferential positional value, and probably acts incombination with other proteins - perhaps includingDecapentaplegic - that are produced in neighbouringsectors of the disc [7].
DI! expression, a convenient early marker of disc cells,may function in establishing the proximal-distal axis ofthe limb. Tissue lacking Dll gene function can form acomplete wing disc, but only the most proximal struc-tures of the leg [1], suggesting that Dll is required forformation of the distal leg. Mutant clones generated infirst instar discs were similarly restricted to forming themost proximal structures of the leg [9]. From theseresults, Cohen and Jurgens [9] argued that Dll isrequired cell-autonomously for a process of intercala-tion that 'fills in' between peripheral (proximal) andcentral (distal, Dll-expressing) regions of the early legdisc. As partial loss-of-function Dll mutants show agraded series of distal leg defects, this requirement maybe graded down the limb [10].
36 Current Biology 1994, Vol 4 No 1
Fig. 2. Interpretation of gene expression patterns in the mesothorax at the time of disc initiation and in the discs; at ten hours of devel-opment; in early larval instars (where known); and in mature imaginal discs. Based on data from [1,2,6,7,8,11].
The Distal-less protein contains a homeodomain, sug-gesting that it acts as a DNA-binding protein rather thanhaving a function directly in signalling cell-cell interac-tions, but graded level of Dll expression might underliethe proximal-distal sequence of positional values [9,10].The key to understanding Dll function may lie in itspattern of expression throughout disc development (itis restricted to the centre of mature leg discs [11]; Fig. 2)and in the way in which it responds to alterations incircumferential patterning signals, such as the level ofwg expression [7,8].
Recently, Campbell and colleagues [2] have studiedanother homeobox-containing gene, al, the expressionof which, they conclude, is induced by wg and dpp,and seems to be important in specifying theproximal-distal axis in both wing and leg discs [2]. Theal gene is first expressed transiently in the thoracic seg-ments of early embryos, in a group of cells at the inter-section of the wg-expressing and the dpp-expressingstripes of cells, as well as in some other cells.Expression of al in the discs reappears only in the thirdinstar, in presumptive distal regions - the tarsus andanterior wing blade - but also elsewhere (Fig. 2).Clearly the sites of late al expression do not correspondsimply to the coincidence point of wg and dpp expres-sion, so there must be other, additional controls operat-ing. When wg was expressed throughout the leg disc,
al expression expanded proximally along the dorsalpart of the dpp stripe [2]. Similarly, when wg wasexpressed ectopically in clones, the new distal portionof the resulting branched leg seemed to coincide withdorsal dpp expression and to express al at its tip,supporting the conclusion that both wg and dppare involved in controlling al expression [2] (butsee below).
The most direct clues to the function of al would comefrom loss-of-function phenotypes, but unfortunatelynone are available, and hypomorphic (reduced func-tion) mutants give only minor defects, some of whichare distal but some proximal [2]. In gain-of-functionexperiments [2], ectopic expression of al, either ubiqui-tously or in clones of cells, resulted in duplication ofthe posterior of the dorsal wing blade. The more exten-sive duplications seem to be branched and to expressal at their ends - although there are no unambigu-ously identifiable distal structures on the wing blades,unlike the claws on the end of each leg . Duplicationsoccurred only rarely in the legs and were not described[2]. From these results. it is concluded that al is notmerely expressed at the distal tip but "acts...in establish-ing the proximodistal axis" [2] - but a note of cautionseems appropriate here.
It is clearly not possible to deduce gene functiondirectly from expression patterns (although it is

DISPATCH 37
puzzling that al is not expressed in first and secondlarval instars [2], as the distal axis can be truncated orduplicated by the temporary absence of wg at thesestages [7]). It is difficult to deduce gene function in acomplex, interacting system, even when lack-of-func-tion mutations are available, as in the case of Dll [9,101.It is certainly unsafe to deduce normal gene functionmainly from gain-of-function experiments, in which theproduct is forced into inappropriate cells, perhaps inthe absence of proteins with which it normally interactsand in the presence of others that it does not normallyencounter. (This caveat also applies to the ectopic wgexperiments [2,8] and generally to much of the currentwork on vertebrate early development!) It seems likelythat Distal-less is an important part of the mechanismunderlying patterning of the long axis of the Drosophilaappendage, but the same cannot be said with any con-fidence for aristaless.
This is an exciting time for those interested in insectappendages. We are, however, only just beginning tounderstand their mechanisms of development. Weknow only a few of the genes involved (including somenot discussed here; see [21), generally only some fea-tures of their expression and, so far, very little abouttheir control or the functions of their products. Fromthe wide range of experimental approaches (molecular,genetic and surgical) now being applied to imaginaldiscs, we can expect new general insights into themolecular basis of cellular 'positional values' andcell-cell interactions [3,4]. In addition, an understandingof imaginal disc development may illuminate other,more zoological issues, such as the developmentalchanges underlying the evolutionary origin of thearthropod limb from the body segments of limblessancestors and the origin of the wing from - perhaps- the leg base of apterygote insects [12].
Acknowledgements: We thank Alfonso Martinez-Arias and JuanPablo Couso for animated discussion, and apologize to authorsunjustly omitted from the short reference list.
References1. COHEN B, SIMCOx AA, COHEN SM: Allocation of the thoracic
imaginal primordia in the Drosophlla embryo. Development1993, 117:597-608.
2. CAMPBELL G, WEAVER T, TOMLINSON A: Axis specification in the
developing Drosophila appendage: the role of wingless,decapentapleglc and the homeobox-containing gene arista-less. Cell 1993, 74:1113-1123.
3. FRENCH V, BRYANT PJ, BRYANT SV: Pattern regulation in epimor-
phic fields. Science 1976, 193:969-981.4. FRENCH V, BRYANT SV, BRYANT PJ: Distal regeneration and sym-
metry. Science 1981, 212:993-1002.5. BATE M, MARTINEz-ARIAS A: The embryonic origin of imaginal
discs in Drosophila. Development 1991, 112:755-761.6. COHEN SM: Specification of limb development in the
Drosophlla embryo by positional cues from segmentationgenes. Nature 1990, 343:173-177.
7. Couso J-P, BATE M, MARTINEZ-ARIAS A: A wingless-dependent
polar coordinate system in Drosophlla imaginal discs Science1993, 259:484-489.
8. STRUHL G, BASLER K: Organizing activity of wingless protein inDrosophila. Cell 1993, 72:527-540.
9. COHEN SM, JURGENS G: Proximal-distal pattern formation inDrosophila: cell-autonomous requirement for Distal-less geneactivity in limb development. EMBOJ 1989, 8:2045-2055.
10. COHEN SM, JURGENS G: Proximal-distal pattern formation inDrosophlla: graded requirement for Distal-less gene activity inlimb development. Roux's Arch Dev Biol 1989, 198:158-169.
11. COHEN SM: Imaginal disc development. In Development ofDrosophila. Edited by Martinez-Arias A, Bate M. Cold SpringHarbor: Cold Spring Harbor Press; 1993.
12. WILLIAMS JA, CARROLL SB: The origin, patterning and evolutionof insect appendages. Bioessays 1993, 15:567-577.
Vernon French and Graeme Daniels, Institute of Cell,Animal and Population Biology, University ofEdinburgh, West Mains Road, Edinburgh EH9 3JT, UK.




![THE CUTICULAR PATTERN IN AN INSECT, RHODNIUS ...[ 45 ]9 THE CUTICULAR PATTERN IN AN INSECT,RHODNIUS PROLIXUS STAL BY M. LOCKE Department of Zoology, University College of the West](https://img.pdfslide.us/doc/110x75/60d8dfdd6bafa25aa5444dad/the-cuticular-pattern-in-an-insect-rhodnius-45-9-the-cuticular-pattern-in.jpg)