Embed Size (px)
Citation preview

Vol. 53, No. 2INFECTION AND IMMUNITY, Aug. 1986, p. 372-3770019-9567/86/080372-06$02.00/0Copyright © 1986, American Society for Microbiology
Pathogenesis of Shigella Diarrhea: Rabbit Intestinal Cell MicrovillusMembrane Binding Site for Shigella Toxin
GEORGE FUCHS,"2 MUNIR MOBASSALEH,"2 ARTHUR DONOHUE-ROLFE,2'3 ROBERT K. MONTGOMERY,"2
RICHARD J. GRAND,"12 AND GERALD T. KEUSCH23*Division of Gastroenterology and Nutrition, Department of Pediatrics,1 Division of Geographic Medicine, Department ofMedicine,3 and the Center for Gastroenterology Research on Absorptive and Secretory Processes,2 New England Medical
Center and Tufts University School of Medicine, Boston, Massachusetts 02111
Received 3 February 1986/Accepted 28 April 1986
This study examined the binding of purified 1251-labeled shigella toxin to rabbit jejunal microvillusmembranes (MYMs). Toxin binding was concentration dependent, saturable, reversible, and specificallyinhibited by unlabeled toxin. The calculated number of toxin molecules bound at 4°C was 7.9 x 1010 (3 x 1010to 2 x 101I)/Ijg ofMVM protein or 1.2 x 106 per enterocyte. Scatchard analysis showed the binding site to beof a single class with an equilibrium association constant, K, of 4.7 x 109 M-' at 4°C. Binding was inverselyrelated to the temperature of incubation. A total of 80% of the labeled toxin binding at 4°C dissociated fromMVM when the temperature was raised to 37°C, but reassociated when the temperature was again brought to4°C. There was no structural or functional change of MVM due to toxin as monitored by electron microscopyor assay ofMVM sucrase activity. These studies demonstrate a specific binding site for shigella toxin on rabbitMVMs. The physiological relevance of this receptor remains to be determined.
The role of the cell-free toxin produced by Shigellaspecies in the pathogenesis of the intestinal manifestations ofclinical shigellosis has been controversial (19). This toxin hasbeen purified and shown to be a multimeric protein of Mr64,000, consisting of one biologically active A subunit (Mr,32,000) which inhibits cytoplasmic protein synthesis at theribosomal level, and five B subunits (Mr, 6,500) whichmediate the binding of toxin to cell receptors (6). A role fortoxin in pathogenesis is suggested by several observations:all Shigella species produce the toxin (20, 30; F. Auclair, A.Donohue-Rolfe, and G. T. Keusch, unpublished data); thepurified protein causes fluid secretion by rabbit small intes-tine as well as the previously described neurotoxic andcytotoxic manifestations associated with the toxin (6); andpatients make a serum toxin-neutralizing antibody duringinfection, indirectly demonstrating the production of toxinantigen in vivo (20, 23). In addition, by using a new enzyme-linked immunofluorescence assay method, we recentlyfound toxin antigen in stools of patients with acute Shigelladysenteriae 1 infection (A. Donohue-Rolfe, M. Kelley, M.Bennish, and G. T. Keusch, submitted for publication).Controversy has persisted, however, because of the
known pathogenic importance of the invasive capacity of theorganism and the failure of noninvasive but toxinogenicShigella variants to cause disease in either experimentalanimal models or human volunteers (11, 24). The latterargument is mitigated, however, because noninvasive strainsfail to colonize the host. Even more importantly, it hasrecently been learned that noninvasive but colonizing entericpathogens such as enteropathogenic serotypes of Esche-richia coli (which do not produce either heat-labile orheat-stable E. coli toxins) and strains of Vibrio choleraedeleted of the genes for both the A and B subunits of choleratoxin, do cause diarrhea and produce a toxin (Shiga-liketoxin) apparently identical or closely related to shigella toxin(25, 29).
* Corresponding author.
Because the rabbit has been used as an experimentalmodel to detect the enterotoxic activity of shigella toxin (19),we examined rabbit enterocyte microvillus membranes(MVMs) for the presence of a toxin receptor. In this paperwe report the characteristics of this receptor.
(Presented in abstract form at the Annual Meeting of theAmerican Gastroenterological Association, May 1984 [Gas-troenterology 86:1083].)
MATERIALS AND METHODSPurification and assay of Shigella toxin. Toxin was pro-
duced from S. dysenteriae 1, strain 60R, and purified by ourpublished methods (6). Bacterial cell lysates of broth-grownorganisms were chromatographed on Blue Sepharose(Pharmacia, Inc., Piscataway, N.J.), eluted in 0.5 M NaCl,and subjected to chromatofocusing (LKB, Uppsala, Swe-den). The toxin was recovered in the pH 7.0 to 7.1 fractionsand passed over a Bio-Gel P-60 column (Bio-Rad Laborato-ries, Richmond, Calif.) to remove ampholytes. Purified toxinwas lyophilized in NH4HCO3 buffer and stored at -70°Cuntil reconstituted for use.Toxin was assayed by cytotoxicity in HeLa cell monolay-
ers and by enterotoxicity in ligated rabbit ileal loops aspreviously described (22, 23). Toxin protein was measuredwith the Bio-Rad Assay Kit II (Bio-Rad), with bovine serumalbumin as standard.
Radiolabeling of toxin. Purified shigella toxin was iodin-ated by a modification of the chloramine T method (15). A1-mCi amount of carrier-free 125I-labeled sodium iodide wasadded to 10 to 20 ,ug of toxin in 150 RI of sodium phosphate(0.1 M, pH 7.4). Next, 20 [lI of chloramine T (2.5 mg/ml) wasadded, followed in 30 s by 20 RI of sodium metabisulfite (5mg/ml). Finally, 100 ,ug of rabbit hemoglobin was added as acarrier protein, and the unbound iodine was separated on a10-ml Sephadex G-25 column. Multiple iodinated toxin prep-arations were used for the different experiments; specificactivity of these preparations ranged from 16,000 to 30,000cpm per ng of toxin protein. There were no differencesdetected between native toxin and iodinated toxin when
372

INTESTINAL BINDING SITE FOR SHIGELLA TOXIN 373
examined by sodium dodecyl sulfate-polyacrylamide gelelectrophoresis as previously described (6). The labeledtoxin was fully active in HeLa cell cytotoxicity and in rabbitjejunal loop enterotoxicity assays. In some experimentstoxin was also labeled by the Bolton-Hunter method (1).
Preparation of intestinal MVMs. Adult New Zealand Whiterabbits were sacrificed by using an overdose of nembutal,and the entire small bowel was removed, washed thoroughlywith iced normal saline, and divided into five equal segmentsfrom the ligament of Treitz to the ileocecal valve. Proximalsegment 2 was used to prepare MVMs by a modification ofthe method described by Kessler et al. (17). The mucosa washarvested by scraping with a glass microscope slide on achilled 4°C glass plate. All subsequent steps were carried outat 4°C. Mucosal scrapings were suspended in 50 mM man-nitol-2 mM Tris hydrochloride buffer (pH 7.1) and homoge-nized in a Waring blender at high speed for 2 min. CaCl2 wasadded to homogenates to give a final concentration of 10mM. After incubation for 15 min, the homogenate wascentrifuged at 3,000 x g for 15 min. The pellet containingcellular debris and organelles was discarded, and the super-natant was recentrifuged at 27,000 x g for 30 min. Theresulting pellet, which contained the crude MVMs, wassuspended by homogenizing with a Potter-Elvejhem homog-enizer in 50 mM mannitol-10 mM Tris hydrochloride buffer(pH 7.1) and then was centrifuged at 27,000 x g for 30 min.This pellet, containing the final MVM preparation, wassuspended in 0.01 M K2PO4 (pH 6.8) and stored in aliquots at-70°C for subsequent use. Unless otherwise indicated, alldata were obtained by using this method of MVM prepara-tion.
In one experiment, MVMs were also prepared by the sametechnique from a known number of isolated intestinal epi-thelial cells recovered by a modification of the method ofHarrison and Webster (14). Everted jejunal segments werevibrated on a metal rod in 0.1 M phosphate buffered saline(PBS), (pH 7.4) at 4°C. Mucus and debris were first removedby vibrating at high frequency and low amplitude for 1 min.After this, villus epithelial cells were released by vibratingfor 5 min. Cells were recovered by centrifugation, suspendedin PBS (pH 7.4), and counted in a Neubauer hemocytometerchamber. When examined by phase contrast microscopy,isolated villus epithelial cells represented more than 90% ofthe population, as determined by their columnar morphologyand the presence of a distinct apical brush border.
Protein content was measured by the method of Lowry etal. (26). The purification of the MVMs was assessed bydetermining the specific activity of sucrase, lactase, andmaltase as described by Grand and colleagues (12, 13, 16).The purified MVMs were enriched by 15- to 20-fold forsucrase activity. Where required, percent recovery of MVMprotein was corrected on the basis of sucrase-specific activ-ity.
Binding of 1251-labeled shigella toxin to MVMs. A modifi-cation of the membrane filter technique of Cuatrecasas wasused for all binding studies (5). Triplicate reaction mixtureswere prepared which contained various amounts of MVMprotein (0.1 to 100,ug) in PBS (pH 7.4) containing 0.1%bovine serum albumin (PBS-BSA) and 125I-labeled shigellatoxin, in a final volume of 100,ul. All experiments includedduplicate controls not containing MVM protein. Althoughless than 1% of the added radioactive toxin bound to filters inthe absence of MVM, all data were corrected for backgroundradioactivity determined from these controls. The mix wasincubated at4°C for 20 min, unless otherwise noted. Labeledshigella toxin bound to MVMs was then separated from free
toxin by rapid filtration under vacuum through a 22-p.mpolyvinylidine fluoride Durapore filter (Millipore Corp.,Bedford, Mass.). Filters were then washed with 10 ml ofPBS-BSA, air dried, and counted in a gamma scintillationspectrometer (Beckman Instruments, Inc., Fullerton,Calif.). Specific binding of shigella toxin to MVMs wasdetermined by the simultaneous addition of labeled toxinplus a 1,000-fold excess of unlabeled toxin to duplicate setsof reaction mixtures. Residual counts under these conditonswere subtracted from total counts in the absence of addedcold toxin. Nonspecific binding was never more than 2% oftotal binding. Values for the equilibrium association constant(K) and the number of receptors per microgram of MVMprotein (n) were obtained by a least-squares fit of the data toa one-ligand, one-receptor Scatchard model (8), using theplot of the bound counts versus toxin concentration. Thedata were corrected for the binding efficiency of toxinexperimentally measured in this study; that is, calculationsassumed that the maximum binding of toxin was 50% of theinput. Data were weighted in inverse proportion to thenumber of bound counts, as described by Rodbard (31).
Dissociation of 125I-labeled shigella toxin from MVMs wasdetermined as follows. Labeled toxin was incubated withMVM at 4°C for 20 min, when a 1,000-fold excess of coldtoxin was added. Portions were removed before and atvarious time intervals after the addition of the cold toxin,filtered, and washed as above, and the radioactivity bound tothe filters was determined as described.To determine temperature dependence of binding, a 1-ml
sample of reaction mixture containing 75 ,ug of MVMs and 50ng of 125I-labeled shigella toxin was brought to equilibrium at4°C, then warmed to 37°C, and finally chilled to 4°C again.Samples were removed after 20 min at each temperatureduring the experiment and were rapidly filtered and countedas above.
Electron microscopy. MVM (0.5 p.g of protein) was incu-bated for 20 min in the presence or absence of 20 ng ofshigella toxin in 100 ,ul of PBS-BSA. MVMs were fixed bythe addition of an equal volume of ice-cold 8% gluter-aldehyde in 0.1 M sodium phosphate buffer (pH 7.4). After 2h, the fixed vesicles were collected for processing by cen-trifugation, then dehydrated through graded ethanols, em-bedded in Epon, and sectioned with a diamond knife. Thinsections were examined with a JEOL electron microscope at60 kV.
Statistical evaluation. All experiments were performed intriplicate and were repeated at least three times with dif-ferent MVM preparations. Data are reported as the mean ±one standard deviation or standard error.
RESULTS
Binding of purified shigella toxin to MVMs. When 125i-labeled shigella toxin (3 to 4 ng in different experiments) wasadded to increasing amounts of MVM protein, toxin bindingwas directly related to the concentration of MVM presentand was linear up to a concentration of 5,ug of MVM per 100,ul when the amount of labeled toxin became limiting (Fig. 1).The binding efficiency of iodinated toxin in the presence ofexcess MVM was 45 + 5% (mean + standard deviation).Labeling with the Bolton-Hunter reagent did not improvebinding efficiency. Toxin that failed to bind to an excess ofMVM was recovered in the filtrate. This material was devoidof cytotoxic activity in the HeLa cell system and did not bindto fresh MVM (data not shown). Iodinated toxin stock (i.e.,
VOL. 53, 1986

374 FUCHS ET AL.
not reacted with MVM) and toxin bound to MVM orrecovered in the filtrate were solubilized in sodium dodecylsulfate and subjected to polyacrylamide gel electrophoresis.Autoradiography of these three labeled toxin preparationsrevealed neither differences in relative density of the radio-active bands representing the two toxin subunits nor thepresence of contaminants (data not shown). We excluded thepossibility that toxin bound to MVM vesicles might escapecapture by the membrane filters by measuring protein con-tent and sucrase activity in filtrates of control reactionmixtures containing only MVM and buffer. No protein andonly trace sucrase activity (<1% of the total applied to thefilter) was found.
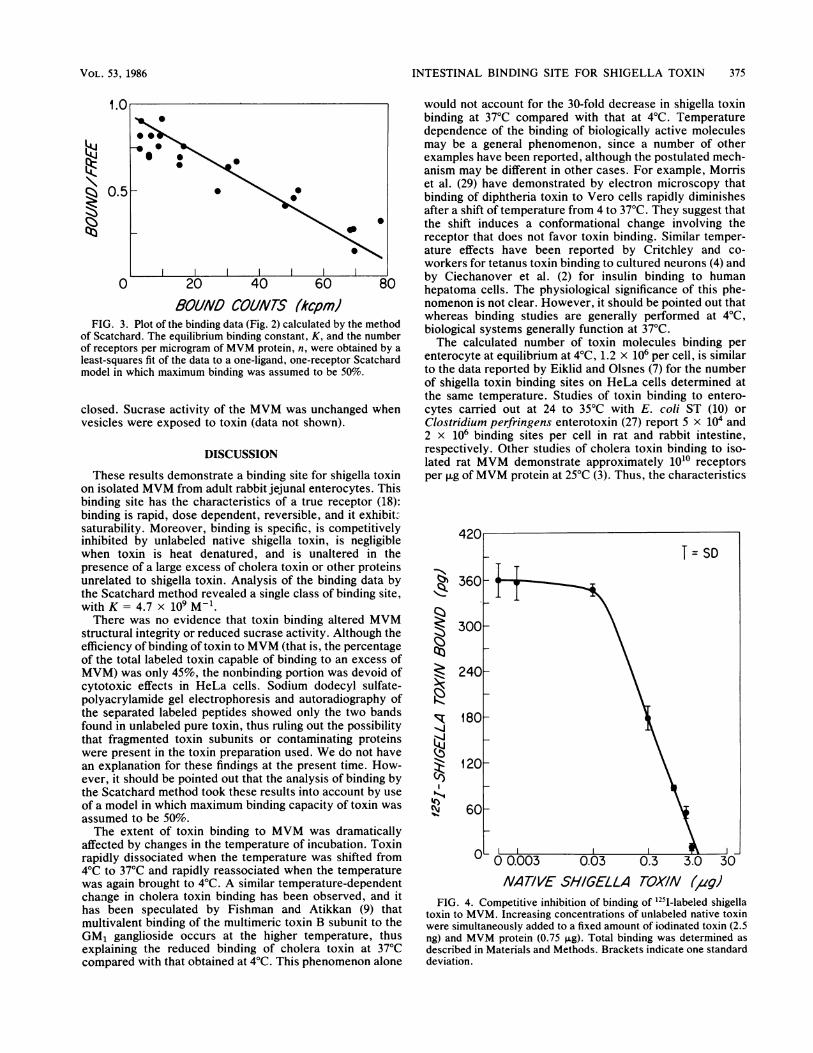
Binding was saturable when the concentration of MVMwas fixed and increasing amounts of 1251-labeled toxin wereadded (Fig. 2). Until saturation was achieved, binding wasdirectly proportional to the amount of labeled toxin that wasadded. Based on a molecular weight of 64,000 for theholotoxin (6), we calculated that 7.9 x 1010 (3 x 1010 to 2 x1011) molecules of toxin were bound per ,ug ofMVM proteinin different MVM preparations from individual rabbits. Anal-ysis of equilibrium binding data according to the Scatchardmodel (as described in Materials and Methods) revealed asingle class of binding sites with a K of 4.7 x 109 M-1,determined in six separate MVM preparations (Fig. 3).MVMs were also simultaneously obtained from isolated
enterocytes and from mucosal scrapings in the same animalin order to determine the number of binding sites perenterocyte. In this experiment, 1 p.g ofMVM was calculatedto represent 6.8 x 104 cells after correction for the actualrecovery of MVMs based on sucrase-specific activity. Byusing this value to convert our data for number of bindingsites per microgram of MVM protein to number of bindingsites per cell, we calculate a value of 1.2 x 106 binding sitesper enterocyte.
Specificity of binding. Heat-denatured toxin (90°C, 30 min)
1500- T= SEM
1250
750_
5000
250
0 5 10 i5 20 25 XJ80MVM (lag
FIG. 1. Increasing concentrations of MVM were mixed with afixed amount of 251I-labeled shigella toxin (100,000 cpm; specificactivity, 2.5 x 104 to 3 x 104 cpm per ng of toxin protein) at 4°C, andtoxin binding was measured as described in Materials and Methods.Brackets indicate one standard error.
2.4-
k 2.0-
:;~ 1.6-
00.8
0.4
0 8 16 24 32 40 48 56'251-SHIGELLA TOJXIN ADDED (ng)
FIG. 2. Increasing amounts of 125I-labeled shigella toxin wereadded to 0.75 F±g ofMVM at 4°C, and toxin binding was measured asdescribed in Materials and Methods. Brackets indicate one standarderror.
failed to bind to MVM (data not shown). In contrast, whenvarious amounts of undenatured unlabeled shigella toxinwere added simultaneously with a fixed amount of l25j1labeled native toxin to the reaction mix, competitive inhibi-tion of binding occurred in a dose-related fashion (Fig. 4).Unrelated proteins, including 10 ,ug of purified cholera toxinper ml (a gift of D. Michael Gill), Concanavalin A, and wheatgerm agglutinin, or 0.1% BSA or 0.01% hemoglobin, allfailed to inhibit binding of 125I-labeled shigella toxin toMVM.
Dissociation of 12SI-labeled shigella toxin. When an excessof cold toxin was added to MVM after the binding of labeledtoxin at 4°C, dissociation of toxin was rapid, and 52 to 63%of specific counts were displaced within 5 min of the additionof the cold toxin, increasing to 75% by 1 h. However,dissociation was never complete, and even after 48 h, 8 to9% of the label remained associated with MVM.
Effects of temperature on toxin binding. Toxin binding toMVMs was directly related to the incubation temperature(Fig. SA). Binding at 4°C or 20°C reached equilibrium by 20min; binding to MVM incubated at 37°C was detectableimmediately upon the addition of toxin and did not increasewith time. The level of binding at 37°C was approximately1/30 of that observed at 4°C. Binding at all temperatures wascompetitively inhibited by excess unlabeled toxin. Whenbinding was allowed to proceed to equilibrium at 4°C (20min) and the temperture was then shifted to 37°C for another20 min, approximately 80% of the bound toxin dissociated(Fig. SB). However, shifting the temperature back to 4°C for20 min resulted in reassociation of the toxin with the MVMto the same extent as originally observed at 4°C.
Effects of shigella toxin on integrity of MVM. Examinationof electron photomicrographs of MVM incubated in thepresence or absence of toxin revealed homogenous mem-brane vesicles of uniform size. There was no evidence ofstructural damage and no increase in the number of visiblemembrane fragments, and the vesicles remained intact and
INFECT. IMMUN.
INTESTINAL BINDING SITE FOR SHIGELLA TOXIN 375
1.
0.
0
FIG. 3. Plot of theof Scatchard. The equof receptors per micrcleast-squares fit of themodel in which maxir
closed. Sucrase actvesicles were expo,:
These results der
would not account for the 30-fold decrease in shigella toxinbinding at 37°C compared with that at 4°C. Temperaturedependence of the binding of biologically active moleculesmay be a general phenomenon, since a number of otherexamples have been reported, although the postulated mech-anism may be different in other cases. For example, Morriset al. (29) have demonstrated by electron microscopy that
o binding of diphtheria toxin to Vero cells rapidly diminishesafter a shift of temperature from 4 to 37°C. They suggest thatthe shift induces a conformational change involving thereceptor that does not favor toxin binding. Similar temper-ature effects have been reported by Critchley and co-workers for tetanus toxin binding to cultured neurons (4) and
20 40 60 8 by Ciechanover et al. (2) for insulin binding to humanhepatoma cells. The physiological significance of this phe-
VOUND COUNTS (kcpm) nomenon is not clear. However, it should be pointed out thatbinding data (Fig. 2calculatedbythemetho
whereas binding studies are generally performed at 4°C,
jilibrium binding constant, K, and the number biological systems generally function at 37°C.3gram of MVM protein, n, were obtained by a The calculated number of toxin molecules binding perdata to a one-ligand, one-receptor Scatchard enterocyte at equilibrium at 4°C, 1.2 x 106 per cell, is similarmum binding was assumed to be 50%. to the data reported by Eiklid and Olsnes (7) for the number
of shigella toxin binding sites on HeLa cells determined atthe same temperature. Studies of toxin binding to entero-
tivity of the MVM was unchanged when cytes carried out at 24 to 35°C with E. coli ST (10) or
sed to toxin (data not shown). Clostridium perfringens enterotoxin (27) report 5 x 104 and2 x 106 binding sites per cell in rat and rabbit intestine,
DISCUSSION respectively. Other studies of cholera toxin binding to iso-lated rat MVM demonstrate approximately 1010 receptors
nonstrate a binding site for shigella toxin per ,ug of MVM protein at 25°C (3). Thus, the characteristicson isolated MVM from adult rabbit jejunal enterocytes. Thisbinding site has the characteristics of a true receptor (18):binding is rapid, dose dependent, reversible, and it exhibit-saturability. Moreover, binding is specific, is competitivelyinhibited by unlabeled native shigella toxin, is negligiblewhen toxin is heat denatured, and is unaltered in thepresence of a large excess of cholera toxin or other proteinsunrelated to shigella toxin. Analysis of the binding data bythe Scatchard method revealed a single class of binding site,with K = 4.7 x 09 M-1.There was no evidence that toxin binding altered MVM
structural integrity or reduced sucrase activity. Although theefficiency of binding of toxin to MVM (that is, the percentageof the total labeled toxin capable of binding to an excess ofMVM) was only 45%, the nonbinding portion was devoid ofcytotoxic effects in HeLa cells. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis and autoradiography ofthe separated labeled peptides showed only the two bandsfound in unlabeled pure toxin, thus ruling out the possibilitythat fragmented toxin subunits or contaminating proteinswere present in the toxin preparation used. We do not havean explanation for these findings at the present time. How-ever, it should be pointed out that the analysis of binding bythe Scatchard method took these results into account by use
of a model in which maximum binding capacity of toxin wasassumed to be 50%.The extent of toxin binding to MVM was dramatically
affected by changes in the temperature of incubation. Toxinrapidly dissociated when the temperature was shifted from4°C to 37°C and rapidly reassociated when the temperaturewas again brought to 4°C. A similar temperature-dependentchange in cholera toxin binding has been observed, and ithas been speculated by Fishman and Atikkan (9) thatmultivalent binding of the multimeric toxin B subunit to theGM1 ganglioside occurs at the higher temperature, thusexplaining the reduced binding of cholera toxin at 37°Ccompared with that obtained at 4°C. This phenomenon alone
"I"%
(f)
NATIVE SHIGELLA TOXIN (,agJFIG. 4. Competitive inhibition of binding of 125"-labeled shigella
toxin to MVM. Increasing concentrations of unlabeled native toxinwere simultaneously added to a fixed amount of iodinated toxin (2.5ng) and MVM protein (0.75 ,ug). Total binding was determined asdescribed in Materials and Methods. Brackets indicate one standarddeviation.
NS 0
0
.5 _
_
VOL. 53, 1986

376 FUCHS ET AL.
-'414-i
C%4
1l800
1500Q:
1200
R 900.-4
g 600
@ 300
O
B T= SD
T
TIME (minutes) TEMPERATURE (°C)
FIG. 5. Effect of temperature on binding of 'l25-labeled shigella toxin to MVM. (A) Time course of binding of toxin at 4, 20, and 37°C. (B)Reaction mixture containing a fixed amount ofMVM (75 ,g) and '25I-labeled toxin (50 ng) in 1 ml of PBS-BSA was incubated for 20 min at4°C, when a sample was removed and total binding was determined (left bar). The reaction mixture was warmed to 37°C and brought toequilibrium. Samples were removed, and binding was measured (center bar). The reaction mix was then chilled to 4°C, brought to equilibrium,and binding was again determined (right bar). Brackets indicate one standard deviation.
of the rabbit MVM shigella toxin binding site are at leastappropriate for it to be a functional toxin receptor.However, the physiological relevance of a receptor cannot
be ascertained from binding parameters alone. There issubstantial evidence that the receptor on HeLa cells medi-ating the cytotoxic effect of shigella toxin is a tunicamycininhibitable N-linked glycoprotein. For example, treatment ofHeLa cells with the antibiotic tunicamycin, a specific inhib-itor of N-linked glycoprotein synthesis, renders the cellsresistant to toxin effects (21). More recently, we foundevidence of a different nonglycoprotein receptor on bothHeLa cells and rabbit intestinal MVM (1Sa). This secondreceptor is a glycolipid extractable in chloroform-methanol,is tunicamycin resistant, and does not mediate toxincytotoxicity to HeLa cells (1Sa). Because there is no evi-dence of cytotoxicity in rabbit (33) or monkey (32) jejunumexposed to shigella toxin or viable organisms, respectively,and because our data demonstrate a single class of toxin-binding site on jejunal MVMs, we propose that this bindingsite is different from the glycoprotein receptor on HeLa cellsand that it may mediate the in vivo secretory response of thejejunum to the toxin. Although further investigation will berequired to determine whether or not this MVM shigellatoxin binding site is, in fact, a functional receptor, thepresent study clearly identifies for the first time the presenceof a specific shigella toxin binding site on rabbit jejunalenterocyte MVMs.
ACKNOWLEDGMENTS
We gratefully acknowledge the excellent technical assistance ofMarianna Sybicki and Judith Reichler. We thank Henry A. Feldmanfor performing the Scatchard analysis of the binding data and MaryJacewicz for cytotoxicity assays.These studies were supported in part by a Digestive Disease
Center Grant (AM 39428) and a Pediatric Gastroenterology Re-search Training Grant (AM 07471), by Public Health Service grantsAM 32658, HD 18189, Al 16242, and Al 20325, from the National
Institutes of Health, and by a grant in Geographic Medicine from theRockefeller Foundation.
LITERATURE CITED1. Bolton, A. E., and W. M. Hunter. 1973. The labeling of proteins
to high specific radioactivities by conjugation to a 1251Icontaining acylating agent. Biochem. J. 133:529-539.
2. Ciechanover, A., A. L. Schwartz, and H. F. Lodish. 1983. Theasialoglycoprotein receptor internalizes and recycles indepen-dently of the transferrin and insulin receptors. Cell 32:267-275.
3. Critchley, D. R., J. L. Magnani, and P. H. Fishman. 1981.Interaction of cholera toxin with rat intestinal brush bordermembranes. J. Biol. Chem. 256:8724-8731.
4. Critchley, D. R., P. G. Nelson, W. H. Habig, and P. H. Fishman.1985. Fate of tetanus toxin bound to the surface of primaryneurons in culture: evidence for rapid internalization. J. CellBiol. 100:1499-1507.
5. Cuatrecasas, P. 1973. Interaction of Vibrio cholerae enterotoxinwith cell membranes. Biochemistry 12:3547-3558.
6. Dononue-Rolfe, A., G. T. Keusch, C. Edson, D. Thorley-Lawson,and M. Jacewicz. 1984. Pathogenesis of shigella diarrhea. IX.Simplified high yield purification of shigella toxin and charac-terization of subunit composition and function by the use ofsubunit-specific monoclonal and polyclonal antibodies. J. Exp.Med. 160:1767-1781.
7. Eiklid, K., and S. Olsnes. 1980. Interaction of Shigella shigaecytotoxin with receptors on sensitive and insensitive cells. J.Receptor Res. 1:199-213.
8. Feldman, H. A. 1983. Statistical limits in Scatchard analysis. J.Biol. Chem. 258:12865-12867.
9. Fishman, P. H., and E. E. Atikkan. 1980. Mechanism of actionof cholera toxin: effect of receptor density and multivalentbinding on activation of adenylate cyclase. Membrane Biol.54:51-60.
10. Frantz, J. C., L. Jaso-Friedman, and D. C. Robertson. 1984.Binding of Escherichia coli heat-stable enterotoxin to rat intes-tinal cells and brush border membranes. Infect. Immun.43:622-630.
11. Gemski, P., Jr., A. Takeuchi, 0. Washington, and S. B. Formal.1972. Shigellosis due to Shigella dysenteriae 1: relative impor-tance of mucosal invasion versus toxin production in pathogen-
INFECT. IMMUN.

INTESTINAL BINDING SITE FOR SHIGELLA TOXIN 377
esis. J. Infect. Dis. 126:523-530.12. Grand, R. J., D. A. Chong, and K. J. Isselbacher. 1972.
Intracellular processing of disaccharidases: the effect of actino-mycin D. Biochim. Biophys. Acta. 261:341-352.
13. Grand, R. J., R. K. Montgomery, and A. Perez. 1985. Synthesisand intracellular processing of sucrase-isomaltase in rat jeju-num. Gastroenterology 88:531-538.
14. Harrison, D. D., and H. L. Webster. 1969. The preparation ofisolated intestinal crypt cells. Exp. Cell Res. 55:257-260.
15. Hunter, W. M., and F. C. Greenwood. 1962. Preparation ofiodine-131-labeled human growth hormone of high specific activ-ity. Nature (London) 194:495-496.
15a.Jacewicz, M., H. Clausen, E. Nudelman, A. Donohue-Rolfe, andG. T. Keusch. 1986. Pathogenesis of shigella diarrhea. XII.Isolation of a shigella toxin-binding glycolipid from rabbit jeju-num and HeLa cells and its identification as globotriaosyl-ceramide. J. Exp. Med. 163:1391-1404.
16. Jonas, M. M., R. K. Montgomery, and R. J. Grand. 1985.Intestinal lactase synthesis during postnatal development in therat. Pediatr. Res. 19:956-962.
17. Kessler, M., 0. Acuto, C. Storelli, H. Murer, M. Muller, and G.Semenza. 1978. A modified procedure for the rapid preparationof efficiently transporting vesicles from small intestinal brushborder membranes. Biochim. Biophys. Acta 506:136-154.
18. Keusch, G. T. 1979. Specific membrane receptors: pathogeneticand therapeutic implications in infectious diseases. Rev. Infect.Dis. 1:517-529.
19. Keusch, G. T., A. Donohue-Rolfe, and M. Jacewicz. 1985.Shigella toxin and the pathogenesis of shigellosis, p. 193-205. InD. C. Evered and J. Whelan (ed.), Microbial toxins and diar-rhoeal disease. Ciba Foundation Symposium 112. The CibaFoundation, London.
20. Keusch, G. T., and M. Jacewicz. 1976. The pathogenesis ofshigella diarrhea. VI. Toxin and antitoxin in S. flexneri and S.sonnei infections in humans. J. Infect. Dis. 135:552-556.
21. Keusch, G. T., M. Jacewicz, and A. Donohue-Rolfe. 1986.Pathogenesis of shigella diarrhea. X. Evidence for an N-linkedglycoprotein receptor and receptor modulation by P-galac-tosidase. J. Infect. Dis. 153:238-248.
22. Keusch, G. T., M. Jacewicz, and S. Z. Hirschman. 1972.Quantitative microassay in cell culture for enterotoxin of Shi-
gella dysenteriae 1. J. Infect. Dis. 125:539-541.23. Keusch, G. T., M. Jacewicz, M. M. Levine, R. B. Hornick, and
S. Kochwa. 1976. Pathogenesis of shigella diarrhea: serumanticytotoxin response produced by toxigenic and nontoxigenicShigella dysenteriae 1. J. Clin. Invest. 57:194-202.
24. Levine, M. M., H. L. DuPont, S. B. Formal, R. B. Hornick, A.Takeuchi, E. J. Gangarosa, M. J. Snyder, and J. P. Libonati.1973. Pathogenesis of Shigella dysenteriae 1 (Shiga) dysentery.J. Infect. Dis. 127:261-270.
25. Levine, M. M., J. B. Kaper, R. E. Black, and M. L. Clements.1983. New knowledge on pathogenesis of bacterial entericinfections as applied to vaccine development. Microbiol. Rev.47:510-550.
26. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193:265-275.
27. McDonel, J. L. 1980. Binding of Clostridium perfringens (1251)enterotoxin to rabbit intestinal cells. Biochemistry 21:4801-4807.
28. Morris, R. E., A. S. Gerstein, P. F. Bonventre, and C. B.Saelinger. 1985. Receptor mediated entry of diphtheria toxininto monkey kidney (Vero) cells: electron microscopic evalua-tion. Infect. Immun. 50:721-727.
29. O'Brien, A. D., and G. D. LaVeck. 1983. Purification andcharacterization of a Shigella dysenteriae 1-like toxin producedby Escherichia coli. Infect. Immun. 40:675-683.
30. O'Brien, A. D., M. R. Thompson, P. Gemski, B. P. Doctor, andS. B. Formal. 1977. Biological properties of Shigella flexneri 2atoxin and its serological relationship to Shigella dysenteriae 1toxin. Infect. Immun. 15:796-798.
31. Rodbard, D., R. H. Lennox, H. L. Wray, and D. Ramseth. 1976.Statistical characterization of the random errors in the radioim-munoassay dose-response variable. Clin. Chem. 22:350-358.
32. Rout, W. R., S. B. Formal, R. A. Gianella, and G. J. Dammin.1975. Pathophysiology of shigella diarrhea in the Rhesus mon-key: intestinal transport, morphological, and bacteriologicalstudies. Gastroenterology 68:270-278.
33. Steinberg, S. E., J. G. Banwell, J. H. Yardley, G. T. Keusch, andT. R. Hendrix. 1975. Comparison of secretory and histologicaleffects of Shigella and cholera enterotoxins in rabbit jejunum.Gastroenterology 68:309-317.
VOL. 53, 1986





























