Embed Size (px)
Citation preview

Oecologia (Berl.) 36, 69--79 (1978) Oecologia �9 by Springer-Verlag 1978
Partitioning of Respiration from Soil, Litter and Plants
in a Mixed-Grassland Ecosystem
R.E. Redmann and Z.M. Abouguendia
Department of Plant Ecology, University of Saskatchewan, Saskatoon, Sask., S7N OWO, Canada
Summary. Carbon dioxide effluxes from plants, litter and soft were measured in two mixed-grassland sites in Saskatchewan, Canada. Ecosystems at both locations were dominated by Agropyron dasystachyum (Hook.) Scribn. Respi- ration rates of intact and experimentally-modified systems were measured in field chambers using alkali-absorption. Removal of green leaves, dead leaves, and litter from a wet sward reduced respiration to as low as 58% of the rate in an intact system. In a dry sward green shoots were the only significant above-ground source of CO2.
Carbon dioxide effluxes from different parts of A. dasystachyum plants, and from soil samples were measured in laboratory vessels at 20 ~ using alkali-absorption. Respiration of green leaves (1.46 mg CO2 g-1 h - l ) was significantly higher than microbial respiration in moist, dead leaf samples (0.79 mg CO2 g- 1 h - 1) or litter (0.75 mg CO2 g- 1 h- ~). Microbial respira- tion in air-dried, dead plant material was very low. Average repiration rates of roots separated from soil cores (0.24 mg CO2 g- ~ h - 1) were lower than many values reported in the literature, probably because the root population sampled included inactive, suberized and senescent roots. Root respiration was estimated to be 17-26% of total CO2 efflux from intact cores.
Laboratory data and field measurements of environmental conditions and plant biomass were combined in order to reconstruct the CO2 efflux from the shoot-root-soil system. Reconstructed rates were 1.3 to 2.3 times as large as field measured rates, apparently because of stimulation to respira- tion caused by the experimental manipulations. The standing dead and litter fractions contributed 26% and 23% of the total CO2 efflux in a wet sward. Both field-measured and reconstructed repiration values suggest that in situ decomposition of standing dead material under moist conditions can be a significant part of carbon balance in mixed grassland.
0029-8549/78/0036/0069/$02.20

70 R.E. Redmann and Z.M. Abouguendia
Introduction
Understanding of the movement of carbon in the soil and vegetation is essential in the study of total ecosystem function. The parameter "soil respiration" has been measured in a number of grassland ecosystems (e.g., Old, 1969; Kucera and Kirkham, 1971; Redmann, in press; Wildung et al., 1975). Although rela- tively easy to measure, soil respiration is actually a complex parameter including fluxes from several ecosystem compartments (Fig. 1). All of the attempts to partition total soil and litter respiration into its components disturb the intact soil-litter-plant system, but the exact effect of this disturbance has not been determined unequivocally.
Coleman (1973) brought soil-litter cores from an Andropogon virginicus- dominated community into the laboratory and separated them into compart- ments. Roots, litter and soil, respectively, contributed 13%, 6%, and 81% of total respiration measured in late spring. Respiration from summed compart- ments ranged from 89% to more than 143% of the output from intact cores. Root respiration never exceeded 17% of the total. Kucera and Kirkham (1971) partitioned root and soil microbial activity in a tall-grass prairie ecosystem by measuring C O 2 effluxes from soils containing progressively fewer roots, and then projecting to soil with no roots. Root metabolism was estimated to contribute 40% of total soil respiration; litter respiration was not measured. The CO2 output from litter in a Festuca suIcata grassland in Czechoslovakia ranged from 5 to 50% of total soil-litter respiration, depending on the moisture content (Tesa~ov~t and Gloser, 1976).
The objective of the present study was to determine the major pathways of carbon flow through compartments of a mixed-grassland ecosystem by: (1) measuring CO2 evolution from intact and modified systems in the field and (2) measuring CO2 efflux from isolated system compartments (e.g., litter, roots) in the laboratory. The effects of precipitation (rainfall or dew) was simulated by wetting the plant material. Respiration rates in intact systems were compared with systems which were "reconstructed" using laboratory data.
The alkali-absorption technique used in this study has the advantages of simplicity and low cost which allow relatively large numbers of replicates to be sampled. The major problems with the technique result from the possible influences of the chamber on microclimate and CO2 diffusion (Sestgk et al., 1971). De Jong et al. (unpublished manuscript) found close agreement between alkali-absorption and soil-CO2 profile techniques in a grassland similar to that used in the present study.
Methods
Site Descriptions
Field studies were carried out at the Matador Field Station located 45 km north of Swift Current, Sask., Canada (50~ 107~ and at Kernan Prairie situated 6 km northeast of Saskatoon, Sask. (52~ 106~ Plant and soil materials used in the laboratory were collected from these same localities.

Partitioning of Respiration in a Mixed Grassland 71
IN 3TS
IDING
E R "
!
TS
ATMOSPHERIC CO 2
TOTAL SYSTEM RESPIRATION
SHOOT RESPIRATION
SOIL-LITTER RESPIRATION
MICROBE RESPIRATION
ROOT RESPIRATION
D ,,,
~TS _ ~ MICROBE RESPIRATION
;ANIC . . . . . . "TER
Fig. 1. Sources and fluxes of carbon dioxide in a typical mixed-grassland ecosystem. Other above- and below-ground heterotrophs are omitted
The vegetation type studied at both sites was dominated by Agropyron dasystachyum (Hook.) Scribn., a typical component of the northern part of the North American mixed grassland. Detailed descriptions of the vegetation on the sites are given by Coupland et al. (1973) and Baines (1973).
The ungrazed sward at the Matador Site was about 20 cm high in mid-summer, with a character- istic layer of standing dead leaves about 10 cm above the soil surface and a layer of litter on the surface (Fig. 1). Approximately three-fourths of the above-ground biomass consisted of dead shoot material. Root biomass at the Matador site averaged 2760 g m 2 (0-150 cm) during 1968-70 (Coupland et aI., 1974). Nearly hatf of this biomass was found in the top 20 cm of the soil. Shoot and root characteristics have not been as intensively studied at Kernan Prairie, but the general structure of the communit ies was similar.
The soils of the Matador site belong to the Sceptre Association, and consist mainly of chernoze- mic-brown soils of heavy-clay texture, developed in glacio-lacustrine deposits. Kernan Prairie soils (SutherIand Association) are clay loam-textured chernozems, also developed in lacustrine materials.
The climate of the region is classified as Type VII 5(VI) Arid, with cold season, according to the Walter and Leith (1960) system. Annual precipitation averages 388 m m (Swift Current) and 345 m m (Saskatoon), about one-third falling during Jane and July. Mean air temperatures, at Matador and Saskatoen respectively, are 19 ~ C and 19 ~ C (July) and - 1 3 ~ C and - 1 7 ~ C (January). The annual means are 3.3 ~ C and 2.1 ~ C.
Field Measurements
Carbon dioxide evolution from intact and modified swards was measured using the alkali-absorption technique (Sest~tk et al., 1971). Vessels containing up to 80 ml of 0.5 N N a O H solution (surface area 102 cm 2) were placed on the soil surface beneath plastic chambers (25 cm diameter, 9-1 volume), which were covered with a luminum foil to reduce overheating. Chambers were pressed about 2 cm into the soil. Chambers were put in place at noon and left for 24 h, after which 35 ml of 2 M BaC1 was added to the NaOH, and the solution was titrated with IN HC1 using phenolphtha- lein indicator. A particular spot was sampIed for only one 24 h period.

72 R.E. Redmann and Z.M. Abouguendia
Treatments of the grass sward included removal of: (1) green (living) leaves, (2) green and dead shoots and (3) green shoots, dead shoots and litter. The control system was left intact. Shoot material was clipped, separated into green and dead categories, dried at 80 ~ C and weighed. Root samples were obtained to a depth of 10 cm using a hand-driven soil sampler 2.5 cm in diameter.
Soil temperature was determined at intervals during the measurement period; air temperature was monitored using a maximum-minimum thermometer enclosed in the chamber. Soil moisture was determined gravimetrically.
Laboratory Studies
Samples of live and dead shoots, litter, soil and roots were removed to the laboratory where CO2 evolution was measured using the alkali-absorption technique. Samples were enclosed in sealed chambers (20 cm diameter, 4 1 volume), along with 40 ml of 0.5 N NaOH. After 24 h the solution was titrated as described previously. Temperature was maintained at 20 +_ 2 ~ C.
Green leaf samples were moistened with distilled water prior to being sealed in the chambers in order to maintain favorable water status. Dead leaves and litter were either moistened or left in an air-dry condition. Soil (including roots) was sampled in cylindrical cores, and in bulk. Roots were removed by agitation under water, or left in the soil depending upon the nature of the experiment. Roots were sterilized by immersion in 70% ethanol (Ss) followed by 10% javex (5 min) and a final rinse in sterile distilled water.
After measurement of CO2 evolution, dry weight of the plant material and soil was determined by oven-drying at 80 ~ C for 12 h. Carbon dioxide evolution was expressed on the basis of plant or soil dry weight.
Results and Discussion
Intact and Partitioned Systems in the Field
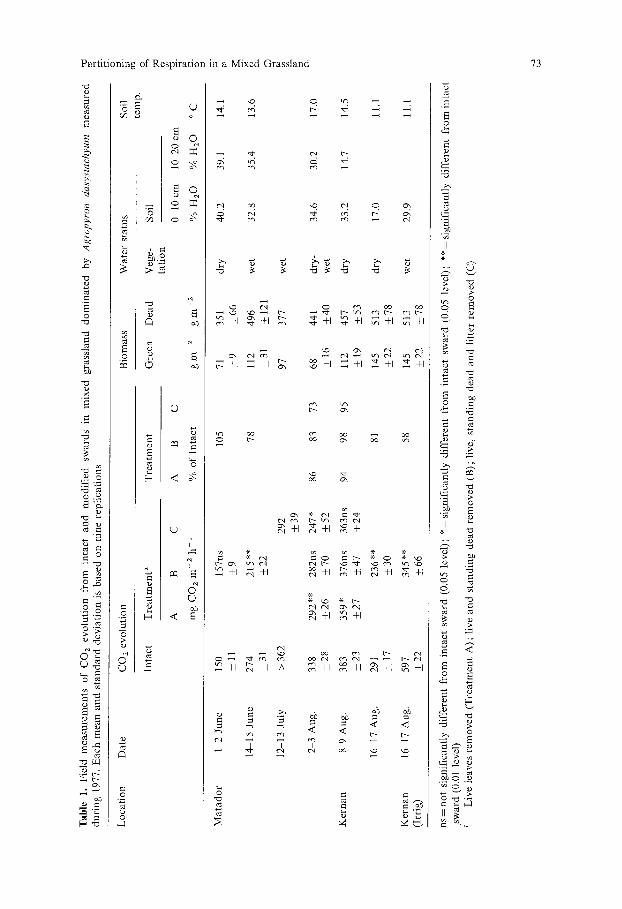
C a r b o n dioxide efflux f rom the intact grassland system dur ing 1977 ranged from 150 to 597 mg CO2 m - 2 h 1 (Table 1). The values for Ma tador are similar
in magni tude to those reported previously for the same locat ion by W a r e m b o u r g
and Paul (1977) and R e d m a n n (in press). Removal of green tissue (Trea tment A) reduced CO2 efflux to abou t 90%
of the intact system. The reduct ion was statistically significant under both wet and dry vegetat ion condi t ions. The reduct ion in total CO2 efflux was probably a direct effect of removal of respiring green tissue rather than an indirect effect
such as a reduct ion in root respira t ion owing to the clipping. The dark condi t ions in the intact chamber should have produced the same effect on root respirat ion
as the removal of green shoots, as observed by Frossard (1977). Removal of bo th green and s tanding dead tissue f rom beneath the absorp t ion
chamber (Trea tment B) caused a statistically significant reduct ion in CO 2 effiux on three of six days. The reduct ion was greatest when the vegetat ion was wet. Efflux of CO z in Trea tmen t B on 16-17 August was only 58% of the wet intact sward. Trea tment B values on other days when the sward was wet (14-15 June and 2-3 August) were 78% and 83% of the intact sward, respectively. The latter value was not statistically significant owing to the large var iabi l i ty .
T rea tmen t C (removal of green, dead and litter material) reduced CO 2 efflux by an addi t ional increment which is an estimate of litter respiration. Under condi t ions when the vegetat ion was wet, the litter layer cont r ibu ted abou t 10%
of the total CO2 efflux.
Tab
le
1.
Fie
ld
mea
sure
men
ts o
f C
Oz
evol
utio
n fr
om
inta
ct
and
mod
ifie
d sw
ards
in
m
ixed
gr
assl
and
do
min
ated
by
A
grop
yron
das
ysta
chyu
m m
easu
red
~z
du
ring
197
7.
Eac
h m
ean
an
d s
tand
ard
devi
atio
n is
bas
ed o
n ni
ne r
epli
cati
ons
Loc
atio
n D
ate
CO
2 ev
olut
ion
Bio
mas
s W
ater
sta
tus
Soi
l te
mp.
In
tact
T
reat
men
t ~
Tre
atm
ent
Gre
en
Dea
d
Veg
e-
Soil
ta
tion
A
B
C
A
B
C
0
10
cm
10 2
0cm
mg
CO
zm
-zh
i
%
of I
ntac
t g
m
z g
m~
2
%
H2
0
%
H2
0
oc
o m 9 �9
Mat
ado
r 1
2 Ju
ne
150
157n
s 10
5 71
35
1 dr
y 40
.2
ill
_+9
+9
_+
66
14
15 J
une
274
215"
* 78
11
2 49
6 w
et
32.8
+
31
+-2
2 _+
31
_+12
1
12-1
3 Ju
ly
> 36
2 29
2 97
37
7 w
et
+3
9
2 3
Aug
. 33
8 29
2**
282n
s 24
7*
86
83
73
68
441
dry-
34
.6
+-28
+
26
+
70
_+
52
+1
6
_+40
w
et
Ker
nan
8 ~
) A
ug.
383
359
* 37
6ns
363n
s 94
98
95
11
2 45
7 dr
y 33
.2
+-23
+
27
_+
47
+2
4
+-19
+
53
16
17 A
ug.
291
236*
* 81
14
5 51
3 dr
y 17
.0
• 17
_+
30
+2
2
+7
8
Ker
nan
16
-17
Aug
. 59
7 34
5**
58
145
513
wet
29
.9
(Irr
ig)
+_22
_+
66
+-2
2 _+
78
39.1
35.4
30.2
14.7
14.1
13.6
17.0
14.5
11.1
11.1
~z
ns=
no
t si
gnif
ican
tly
diff
eren
t fr
om
int
act
swar
d (0
.05
leve
l);
*=
sig
nifi
cant
ly d
iffe
rent
fro
m
inta
ct s
war
d (0
.05
leve
l);
**
=si
gn
ific
antl
y d
iffe
rent
fro
m i
ntac
t sw
ard
(0.0
1 le
vel)
"
Liv
e le
aves
rem
oved
(T
reat
men
t A
) ;
live
an
d s
tand
ing
dead
rem
oved
(B
) ;
live
, st
andi
ng d
ead
and
litt
er r
emov
ed (
C)

74 R.E. Redmann and Z.M. Abouguendia
Table 2. Carbon dioxide efflux from green leaves, dead leaves and shoot litter of Agropyron dasystachyum measured in the laboratory at 20 ~ C. Thirteen replica- tions were used
Moisture status and Green leaves coefficient of variation (CV)
Dead leaves Litter mgCO2g-lh 1
Moist samples 1.46 + 0.27 0.79 _+ 0.21 0.75 _+ 0.38 CV 18 27 51 Dry samples 0.030_+0.010 0.02_+0.01 CV 33 50
Carbon Dioxide Efflux from Leaves and Litter
Labora tory measurements showed that carbon dioxide efflux from green leaves of Agropyron dasystachyum was significantly higher than from either dead leaves or litter (Table 2). The coefficient of variation within plant material was largest in the case of litter samples, apparently because litter was a heterogeneous mixture of plant tissues of various ages and stages of decomposition.
The respiration rates of green leaves were higher than those calculated from the data for the same species presented in the previous section (0.68 mg CO2 g-1 h-l) and in Redmann (1974) (0.80 mg COa g-~ h - l ) . Mois- ture and temperature conditions were similar in all cases, suggesting that some aspect of the experimental procedure, such as the effect of tissue wounding reported in Evans (1972), might have caused the higher rates.
Microbial respiration rates in the dead leaf samples were similar to those in the litter (Table 2). Moist conditions were more favorable than dry. More detailed study of the response of mlcrobial respiration to changes in the water and nutrient status of the dead leaves and litter, and to changes in the nature of the microbial population probably would explain some of the variation in CO2 evolution rates which was observed.
The rates observed here are higher than the respiration of wet litter (0.24 mg CO2 g - 1 h - 1 at 25 ~ C) from a tall-grass prairie reported by Old (1969). Coleman (1973) measured respiration rates of about 0.65 mg CO2 g-1 h-~ for Andropogon virginicus litter (20 ~ C). Mangenot (1966; cited in Bell, 1974) deter- mined that Festuca sylvatica leaves decomposed at a rate of 1.5% per day for an initial 10-day period, after which the rate dropped to 0.38% per day (20 to 100 days). Labelled carbon in grass shoots placed on the soil surface at the Matador grassland site in 1971 lost weight at a rate of about 1.1% per day for the first 30 days (Coupland et al., 1974). The Agropyron dasystachyum leaves under opt imum moisture conditions in this study lost weight at a rate of 1.65% per day. Although such comparisons must be made with caution because of the effects of variations in environmental and microbial characteris- tics, they do show the general magnitude of decomposition rates which can be expected in grassland ecosystems.

Partitioning of Respiration in a Mixed Grassland 75
Root Respiration
The respiration rates of washed roots (0.24_+0.11 mg C O 2 g 1 h -1) did not differ statistically from those of roots which were washed and sterilized (0.23 +_0.07 mg COz g 1 h 1). Thirty-five replications were used. Populations of surface-dwelling microorganisms appeared to be absent or inactive after washing alone. No attempt was made to separate the roots into categories of age or physiological condition, therefore these respiration rates represent the average activity of a mixed root population. Environmental conditions during the experiment were probably near optimum (moist, well-aerated, 20 ~ C), thus the major questions regarding the results involve the effects on root respiration of washing and handling, and of the removal of the photosynthate supply.
The root respiration rates reported here are lower than values reported elsewhere (Table 3). In general, the highest values in the literature (21 24.5 mg CO2 g - t h 1) are for roots grown in solution culture; the lowest (1.1-1.3 mg CO2 g-1 h - l ) are for excised roots removed from natural ecosys- tems. The data also show a strong effect of substrate supply: root respiration in the dark, or in detopped plants is reduced. Roots from plants in later stages of development have lower respiration rates. The proportion of meristematic tissue, which should be positively correlated with the amount of growth-respira- tion, strongly influences root respiration. Barnard and Jorgenson (1977) found that white, succulent, unsuberized meristem tissue had a respiration rate 12 times higher than small, suberized roots.
The published data suggest several explanations for the low respiration rates measured in Agropyron dasystachyum: respiration was measured after detopping, a large proportion of the roots were suberized, and some of the roots were senescent or dormant. In addition, dry soil, or other unfarvorable environmental conditions at the time of collection might have caused reductions in activity.
Partitioning Soil Respiration
The C O 2 efflux from intact soil cores was compared with cores from which the roots had been removed (Table 4). On the first two days of measurements, the root-free cores had CO2 efflux rates equal to the intact cores, implying that root respiration was negligible. A more likely explanation is that soil respira- tion in the root-free cores was stimulated by the root-extraction procedure, as reported previously by Coleman (1973) who used a similar procedure. After two days, the disturbance-stimulus apparently was reduced or absent.
The root-free cores had significantly lower respiration rates on the third and fourth days of the experiment. The reductions (26% and 17% of the intact cores) are estimates of the root respiration in the cores. Root respiration was also calculated using the rate measured in section 3.3 of this paper, along with the root biomass in the cores. This estimate (1.7 mg CO2 g-1 h 1) is close to the rate determined by subtraction of the root-free-core rate from the intact- core rate (Table 4).

76 R.E. Redmann and Z.M. Abouguendia
Table 3. Root respiration of several species of grasses. All measurements were made at 20 ~ C, other environmental conditions are specified
Rate Experimental conditions Species Reference mg CO2 g-1 h 1
0.24 Excised roots, collected from dry soil Agropyron This study layer in the field dasystachyum
21 Shoot-root intact, high light, solution Lolium Hansen (1977) culture multiflorum
1.46 Shoot-root intact, third day of low light, Lolium Hansen (1977) 20 days after defoliation multiflorum
1.88 Severed roots, collected from old field Andropogon Coleman (1973) in May virginicus
1.33 Severed roots, collected from old field Andropogon Coleman (1973) in Dec.-Jan. virginicus
4 Excised new roots, collected from tundra Dupontia Billings et al. fisheria (1977)
1.1 Whole root system, de-topped 24 h Dupontia Billings et al. previously, collected from tundra fisheria (1977)
8.0 Shoot-root intact, collected from the Triticum Osman (1971) field 9 weeks after crop emergence, shoot aestivum and root in darkness
Shoot-root intact, shoot in light Triticum for 24 min aestivurn
Shoot-root intact, collected from the Triticum field 13 weeks after crop emergence, shoot aestivum and root in darkness
Shoot-root intact, shoot in light Triticum for 24 min aestivum
Shoot stage, solution culture, Triticum fresh-washed roots aestivum
Fruiting stage, solution culture, Triticum fresh-washed roots aestivum
9.8 Osman (1971)
2.6 Osman (1971)
3.6 Osman (1971)
24.5 Sauerbeck and Johnen (1976)
1.9 Sauerbeck and Johnen (1976)
Table 4. CO2 evolution from soil and roots of Agropyron dasystachyum separated from soil cores in the laboratory, and from intact cores (soil plus roots). Daily measurements over 4-day period, using 5 replicates
Day Intact Soil (root free) Roots Roots Roots core (intact minus soil) % of intact (soil free calculated) a
mg kg ~ (soil) h - 1 mg kg- 1 (soil) h - 1
1 7.1• 7.8• 2 5.9• 5.9• 3 6.1• 4.5• 1.6 26 1.7 4 5 .4• 4.5• 0.9 17 1.7
* = significantly different from intact core (0.05 level of probability); n s - n o t significantly different from intact sward a Calculated, using root weight from intact cores, a root respiration rate of 0.24 mg CO2 g-Z (roots) h-~, and expressing rate on basis of dry weight of soil in intact cores
Tab
le 5
. C
O2
evo
luti
on
fro
m r
eco
nst
ruct
ed s
ho
ot-
roo
t-so
il s
yst
ems
bas
ed o
n l
abo
rato
ry m
easu
rem
ents
, co
mp
ared
wit
h fi
eld
mea
sure
men
ts a
t K
ern
an P
rair
ie
To
tal
wei
ght
"Dry
" v
eget
atio
n,
mo
ist
soil
M
ois
t ve
geta
tion
, l~
0ois
t soi
l
Res
p.
rate
at
To
tal
Tre
atm
ent
b %
of
Res
p.
rate
at
To
taI
Tre
atm
ent
b %
o
f re
sp.
tota
l re
sp.
tota
l
20 ~
C
fiel
d A
B
C
rc
sp.
20 ~
C
fiel
d A
B
C
re
sp.
tem
p. '
~ te
mp
. ~
mg
CO
2g
-Ih
-I
m
gC
O2
m-2
h-i
m
gC
O2
g
i h
I m
gC
O2
m
2 h
i
7~
P~
O
Rec
on
stru
cted
res
pir
atio
n
Liv
e sh
oo
ts
112
1.46
1.
31
147
22
1.46
1.
31
147
12
Dea
d s
ho
ots
45
7 0.
03
0.02
7 12
12
2
0.79
0.
71
325
325
26
Lit
ter
428
0.02
0.
018
8 8
8 1
0.75
0.
68
289
289
289
23
Ro
ots
(0
10
cm)
1,36
8 0.
24
0.13
2 18
1 18
1 18
1 18
1 28
0.
24
0.13
2 18
1 18
1 18
1 18
1 14
S
oil
(0-t
0 c
m)
124,
000
~ 0.
0045
0.
0025
31
0 31
0 31
0 31
0 47
0.
0045
0.
0025
31
0 31
0 31
0 31
0 25
S
um
65
8 50
3 49
9 49
1 12
52
1105
78
0 49
1 %
of
tota
l re
con
stru
cted
res
pir
atio
n
100
76
76
75
100
88
62
39
Fie
ld-m
easu
red
res
pir
atio
n
383
359
376
363
597
345
% o
f to
tal
fiel
d-m
easu
red
resp
irat
ion
10
0 94
98
95
10
0 58
Rat
io,
reco
nst
ruct
ed:
fiel
d-m
easu
red
1.7
1.4
1.3
1.4
2.1
2.3
~o
C~
co
Rat
e at
fie
ld t
emp
erat
ure
cal
cula
ted
usi
ng
a Q
10 o
f 2.
0 u
Liv
e le
aves
rem
ov
ed (
Tre
atm
ent
A)
; li
ve a
nd
sta
nd
ing
dea
d r
emo
ved
(B
) :
live,
st
and
ing
dea
d an
d l
itte
r re
mo
ved
(C
) C
alcu
late
d u
sin
g a
bul
k de
nsit
y o
f 1.
24 g
cm
- 3
det
erm
ined
for
Ker
nan
Pra
irie
soi
l

78 R.E. Redmann and Z.M. Abouguendia
Comparison of CO 2 EfjTux from Ecosystem Compartments
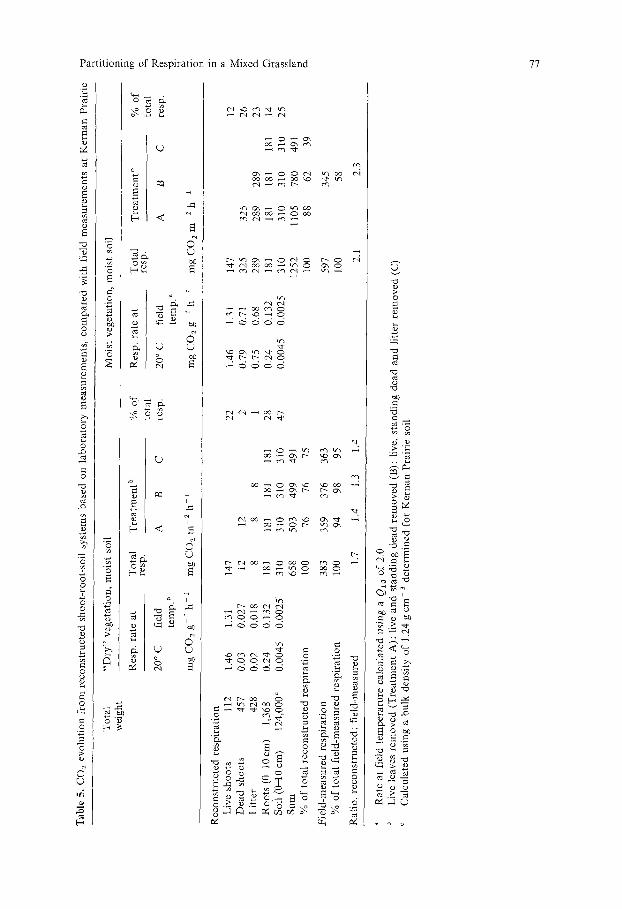
The laboratory data and field measurements of environment and plant biomass were combined in an attempt to reconstruct the CO2 efflux from the shoot-root- soil system (Table 5). The results of the field partitioning of total CO2 efflux at Kernan Prairie on 8-9 August, 1977 were compared with the "reconstructed" system.
When the standing dead and litter were dry, their contribution to total CO2 efflux was small (2% and 1% respectively). Green-shoot respiration and root respiration rates were approximately the same (22% and 28%). The balance of the CO2 (47% of the total) came from soil microbial activity. The contribution of the green-shoot compartment was larger in the reconstructed system. The total CO2 efflux from the reconstructed system was about 70% larger than the total rate measured at Kernan Prairie, even though the calculated rate included only the 0-10 cm soil layer.
Wetting of the standing dead and litter layers increased their CO2 effluxes to 26% and 23% of the total efflux in the reconstructed system. The relative contributions of the other compartments were reduced to about half those under dry conditions. The calculated total CO2 efflux was more than double that measured in the field. The contribution of the litter-root-soil fraction, relative to the total efflux, was about the same in both cases, indicating that the laboratory conditions might have stimulated all the fractions proportionately.
The results suggest that CO2 efflux from wet standing dead and litter, and, therefore, in situ decomposition of this material, can be a significant part of carbon balance in the mixed grassland. Tesa~ovfi and Gloser (1976) have already reported that CO2 released from the litter (perhaps including standing dead) can be as much as 50% of the total efflux from soil and litter. Natural grassland is considerably different in this regard from cultivated ecosystems where plant residues are artificially incorporated into the soil, and where soil microbes play a relatively greater role.
Temporal integration of total CO2 efflux arising from in situ decomposition of shoot materials would require detailed information on sward microclimate, particulary the amount of time that the vegetation is wet (from precipitation, including dew). An effort to combine environmental data with more detailed knowledge of respiration-environment responses, perhaps along the lines attemped in the deciduous forest biome (Edwards and Harris, 1977), would further advance our understanding of carbon balance in grassland ecosystems.
Conclusions
In a dry mixed-grassland sward the living shoots are the only significant above- ground source of CO2; most of the CO2 arises from the root-soil system. When the sward is wet, as much as half of the total CO2 efflux comes from in situ decomposition of standing dead and litter material. This conclusion was supported by both field-measured respiration and "reconstructed" respiration based on laboratory measurements. Air-dried dead plant material respires very slowly. Respiration of roots removed from the field tends to be low, probably because the root population includes inactive, suberized and senescent roots

Partitioning of Respiration in a Mixed Grassland 79
as well as growing :roots with active meristematic tissue. Roots are estimated to contribute about 17 26% of the CO2 arising below the soil surface in this Agropyron dasystachyum-dominated ecosystem.
Acknowledgments. This research was supported in part by a grant from the National Research Council of Canada to the senior author. The helpful review of the manuscript by M.M. Caldwell is gratefully acknowledged.
References
Baines, G.B.K. : Plant distribution on a Saskatchewan prairie. Vegetatio 28, 99 123 (1973) Barnard, E.L., Jorgensen, J.R.: Respiration of field-grown loblolly pine roots as influenced by
temperature and root type. Can. J. Bot. 55, 740-743 (1977) Bell, M.K.: Decomposition of herbaceous litter. In: Biology of plant litter decomposition, Vol.
1 (C.H. Dickinson, G.J.F. Pugh, eds.), pp. 37-67. London: Academic Press 1974 Billings, W.D., Peterson, K.M., Shaver, G.R., Trent, A.W.: Root growth, respiration and carbon
dioxide evolution in an arctic tundra soil. Arct. Alp. Res. 9, 129 137 (1977) Coleman, D.C.: Compartmental analysis of total soil respiration, an exploratory study. Oikos
24, 361 366 (1973) Coupland, R.T., Ripley, E.A., Robins, P.C.: Description of site. I. Floristic composition and
canopy architecture of the vegetative cover. Matador Project Tech. Rep. l 1, 54 pp., University of Saskatchewan, Saskatoon (1973)
Coupland, R.T., Willard, J.R., Ripley, E.A.: Summary of activities 1967-74. Matador Project Tech. Rep. 69, 244 pp., University of Saskatchewan, Saskatoon (1974)
Evans, G.C.: The quantitative analysis of plant growth. Berkeley, Los Angeles: University of California Press 1972
Edwards, N.T., Harris, W.F.: Carbon cycling in a mixed deciduous forest floor. Ecology 58, 431-437 (1977)
Frossard, J.S.: Relations entre l'eclairement des feuilles et l'absorption d'oxyg6ne par les racines chez le tournesol (Helianthus annuus L.). Ann. Agron. 27, 435 445 (1976)
Hansen, G.K.: Adaptation to photosynthesis and diurnal oscillation of root respiration rates for Lolium multiflorum. Physiologia Plantarum 39, 275-27~) (1977)
de Jong, E., Redmann, R.E., Ripley, E.A.: A comparison of methods to measure soil respiration. Unpublished manuscript
Kucera, C.L., Kirkham, D.R.: Soil respiration studies in tallgrass prairie in Missouri. Ecology 52, 912-915 (1971)
Old, S.M.: Microclimates, fire and plant production in an Illinois prairie. Ecol. Monogr. 39, 355-384 (1969)
Osman, A.M. : Root respiration of wheat plants as influenced by age, temperature and irradiance of shoots. Photosynthetica 5, 107-112 (1971)
Redmann, R.E. : Photosynthesis, respiration and water relations of Agropyron dasystachyum mea- sured in the laboratory. Matador Project Tech. Rep. 47, 22 pp., University of Saskatchewan, Saskatoon (1974)
Redmann, R.E. : Soil respiration in a mixed grassland ecosystem. Can. J. Soil Sci. (in press) Sauerbeck, D., Johnen, B.: Der Umsatz von Pflanzenwurzeln im Laufe der Vegetationsperiode
und dessen Beitrag zur Bodenatmung. Z. Pflanzenern. Bodenk. 139, 315-328 (1976) Sestflk Z., (2atsk), J., Jarvis, P.G. (eds.).: Plant photosynthetic production. Manual of methods.
The Hague: Junk 1971 Tesa~ovfl, M., Gloser, J. : Total CO2 output from alluvial soils with two types of grassland commu-
nities. Pedobiologia 16, 364 372 (1976) Walter, H., Lieth, H.: Klimadiagramm-Weltatlas. Jena: Fischer 1960 Warembourg, F.R., Paul, E.A. : Seasonal transfers of assimilated 1~C in grassland: plant production
and turnover, soil and plant respiration. Soil Biol. Biochem. 9, 295-301 (1977) Wildung, R.E., Garland, T.R., Buschbom, R.L.: The interdependent effects of soil temperature
and water content on soil respiration rate and plant root decomposition in arid grassland soils. Soil Biol. Biochem. 7, 373-378 (1975)
Received January 16, 1978





























