Embed Size (px)
Citation preview

JOURNAL OF CELLULAR PHYSIOLOGY 153:563-574 (1992)
Partial Characterization of Skeletal Myoblast Mitogens in Mouse Crushed Muscle Extract
GANG CHEN AND LEBRlS S. QUINN* Departmcnt of t?io/ogica/ Striicturc, Univenity of Washington,
SealUe, Washington 98 195
We have utilized a model system to investigate myotrophic factors released by normal adult mouse muscles following a crush injury. We found that saline extracts from gently crushed mouse muscles (CME) contain potent mitogenic activities which act on primary newborn mouse myoblast cultures, as well as on mouse C2 cells, a mouse myoblast cell line. We compared the activity of CME on mouse myoblasts with that of basic fibroblast growth factor (bFGF) and insulin-like growth factor I (IGF-I), two growth factors known to be mitogenic for primary myoblasts (Allen, Dodson, and Lutein: Exp. Cell. Res., 152:154-I 60, 1984; DiMario and Strohman: Differentiation, 39:42-49, 1988; Allen and Boxhorn: J . Cell. Physiol., 138:311-315, 1989; Dodson, Allen, and Hossncr: Endocrinology, 11 7:2357-2363, 1985; Florini and Magri: Am. J. Physiol., 25h:C701-C711, 1989). We found that CME could act in an additive fashion to saturating doses of bFGF to increase proliferation in myoblast cultures. Additionally, CME acted additively to the combination of saturating amounts of bFGF and IGF-l on both C2 and primary myoblast cultures. We also examined additivity of CME with the combination of saturating doses of bFGF, IGF-I, transferrin (Tf), platelet-derived growth factor (PDGF), epidermal grvwth factor (EGF), adrenocorticotrophin (ACTH), and macrophage colony-stimulating factor (M-CSF). Our data indicate that CME contains Tf, a5 well as one or more uncharacterizecl mitogens for myohlasts which are distinct from Tf, the IGFs, bFtiF, EGF, PDGF, M-CSF, and ACTH. These uncharacterized mitogens may act independently of known growth factors to stimulate myoblast proliferation, or may act through modulation of known growth factor activities. D 1992 WiIey-Liss, Inc.
Adult muscle tissue is capable of regeneration from reserve myoblasts known as “satellite” cells (Mauro, 1961). In normal adult animals, satellite cells consti- tute a mitotically quiescent cell population (Schultz et al., 1978). Following muscle injury, satellite cells reen- ter the cell cycle, proliferate, and undergo terminal dif- ferentiation to form replacement muscle fibers (Carl- son and Faulkner, 1983; Campion, 1984). Little is known about the factors which control the mitotic state of satellite cells. Cell culture studies have shown that in vitro, the proliferation and differentiation of adult as well as embryonic myoblasts are under the control of growth factors (Florini, 1987; Florini and Magri, 1989). The most extensively characterized growth factors which stimulate proliferation of primary myoblasts in culture are basic fibroblast growth factor (bFGF; Allen et al., 1984; DiMario and Strohman, 1988) and the insu- lin-like growth factors (IGF-I and IGF-11; Dodson et al., 1985; Allen and Boxhorn, 1989; Greene and Allen, 1991).
Recently, Bischoff (1986b) has shown that extracts from gently crushed adult ra t muscles are mitogenic for embryonic rat myoblasts, and for adult rat satellite cells which are already in the cell cycle. Crushed mus- cle extract (CME) can also trigger quiescent, Go-phase adult rat satellite ceIIs to reenter the cell cycle (Bischoff, 198613, 1990). Extracts from uncrushed mus- t? 1992 WILEY-LISS, INC.
cles, however, do not have such activities (Bischoff, 1986b3. CME provides a model system to investigate factors released during tissue injury that stimulate qui- escent satellite cells to enter mitosis, and which allow or promote proliferation once the cells have entered the cell cycle. CME clearly contains potent factors which regulate myogenesis.
The active factors present in rat CME are unknown. Bischoff (1986a, 1989) has suggested that these factors are not bFGF or the IGFs, since the physiological activ- ities and biochemical properties of CME differ from those of purified bFGF and the IGFs. Thus, rat CME could contain some novel inyogenic regulators. Alter- natively, when combined, known growth factors could act differently than when added to cells individually (Allen and Boxhorn, 1989; Greene and Allen, 1991). In this paper, we show that extracts from crushed adult mouse muscles produce effects on mouse myoblasts similar to those reported by Bischoff for the rat model system. CME from adult mice is strongly mitogenic for primary mouse myoblasts and for C2 cells, a myogenic
Received March 16,1992; accepted July 9,1992. “‘To whom reprint requestsicorrespondence should be addressed.

564 CHEN AND QUINN
cell line from adult mouse muscle (Yaffe, and Saxel, 1977). We also examined whether the activities in mouse CME are due to bFGF, the IGFs, or other identi- fied mitogens for mammalian myoblasts. Our data in- dicate that mouse CME contains transferrin (TO, plus one or more uncharacterized mitogens which are dis- tinct from Tf, the IGFs, bFGF, epidermal growth factor (EGF), platelet-derived growth factor (PDGF), mac- rophage colony-stimulating growth factor (M-CSF), and adrenocorticotrophin (ACTH).
MATERIALS AND METHODS Sera and growth factors
Horse serum (HS) and fetal calf serum IFCS) were purchased from Hyclone. Sera were diluted in Eagles' Minimal Essential Medium (MEM; Sigma) for culture medium. Human recombinant bFGF, IGF-I, IGF-11, EGF, as well as Tf purified from human serum were from Upstate Biotechnology. Mouse Tf, purified from serum, was from Cappel. Identical results were ob- tained from murine and human Tf. Human recombi- nant M-CSF was from Genzyme. Human synthetic ACTH and the polyclonal antibody against human Tf were purchased from Sigma. Recombinant human PDGF-BB was obtained from Dr. D. Bowen-Pope (De- partment of Pathology, University of Washington). All the above growth factors made from human sequences are active on mouse cells.
Muscle extract CME from 8-10 week old Swiss-Webster mice (Tyler
Laboratories, Bellevue, WA) was made following the procedure described by Bischoff (1986b) for rat muscle extract. Mice were sacrificed by COz asphyxiation, then used immediately for tissue collection. Anterior thigh muscles and posterior leg muscles were excised by cut- ting both tendons. The muscles were then pressed gen- tly seven times with a pair of blunt forceps. The tissue was placed in cold Tris-buffered saline (TBS; 50 mM Tris, 0.9% NaC1, pH 7.6) and incubated a t 4°C with gentle shaking for 90 minutes. A ratio of 1 ml TBS for the muscles from one mouse was used for the extrac- tion. Soluble substances which were released were col- lected in the supernatant after centrifugation of mus- cles for 3 minutes in a clinical centrifuge. After sterilization by filtration using a 0.22 pm Millex-GS filter (Millipore), CME was used in cell culture studies
ACTH AFA bFGF CME EGF FCS FITC HS _. .
'H-TdR IGF-I IGF-I1 M-CSF MEM PDGF ~~
TBS TCA Tf
A hhreviations
adrenocorticotrophin alcohollformaliniacetic acid basic fibroblast growth factor crushed muscle extract epidermal growth factor fetal calf serum fluorescein isothiocyanate horse serum W t h y midine insulin-like growth factor I insulin-like growth factor I1 macrophage colony-stimulating growth factor Minimal Essential Medium platelet-derived growth factor Tris-buffered saline trichloroacetic acid transferrin
a t concentrations of 70 pg to 330 pg of total protein per milliliter. The yield of CME was approximately 3 mg of protein from one adult mouse. Protein concentration was assayed by the method of Bradford (1976).
Cell culture For primary cell cultures, 5-day-old Swiss-Webster
mice (Tyler Laboratories) were sacrificed by CO, as- phyxiation. Leg muscles were aseptically removed and cut into small pieces, then digested with 0.1% trypsin (GIBCOI and 0.05% collagenase in MEM simulta- neously for 50 minutes at 37°C with occasional shaking. The enzymes were removed from the tissue by centrifu- gation at 1,OOOg for 5 minutes. The pellet was sus- pended in 10% FCS and pipetted up and down 25 times with a Pasteur pipet. Muscle fragments were further sheared through a 21G needle five times. The suspen- sion was then filtered through two layers of lens paper. Cell yields were determined with a hemocytometer. The cells were plated onto 35 mm tissue culture dishes which had previously been coated with gelatin and treated with 25% HS for at least 5 hours a t 37°C. The plating density was 1.0-1.2 x lo5 cells per dish in 1.5 ml 10% FCS in MEM. Culture medium was changed after 18-20 hours (day l), and, if necessary, on day 3. Growth factors and CME were added a t every medium change. All cultures were maintained a t 37°C in a hu- midified incubator with 5% C02 in air. In some cases, a "preplating" procedure was used to enrich primary cul- tures for myogenic cells (Cossu et al., 1989). In this procedure, cells were preplated in untreated 100 mm tissue culture dishes a t 2.5 x 10' cells per dish, and then incubated for 90 minutes a t 37°C. The nonadher- ent fraction was recounted and plated onto 35 mm dishes a t 1.0-1.2 x lo5 cells per dish. The preplated cell cultures were maintained as described above for the nonpreplated cultures.
C2 cells were plated in 1.5 ml 10% FCS at a density of 5 x lo3 cells per dish on untreated 35 mm tissue cul- ture dishes. Medium was changed on the next day to 5% HS, 2% FCS, or to 0.5% FCS, plus factors to be investi- gated. Varying concentrations of FCS were used in in- dividual experiments in order to allow analysis of the effects of various growth factors.
Assay of cell proliferation by incorporation of "H-thymidine into DNA
Two days after the initial medium change and addi- tion of factors, cultures were given 'H-thymidine (3H- TdR; New England Nuclear, 6.7 Ci/mmol) a t a final concentration of 0.5 pCi/ml. Four hours later, the cells were rinsed with cold MEM, then 1 ml of cold 5% trichloroacetic acid (TCA) in TBS was added to each dish. DNA was allowed to precipitate for a t least 2 hours at 4°C. Cells were rinsed again with cold 5% TCA, solubilized with 0.5 N NaOH, and transferred to scintillation vials containing 10 ml Scinti-verse (Fisher Scientific). Radioactivity was measured using a Pack- ard MINAXI scintillation counter.
Assays of differentiation and proliferation by immunocytochemical staining and cell counting
To quantitate muscle differentiation and prolifera- tion, cells were stained with MF-20 (Developmental

CRUSHED MUSCLE EXTRACT 565
Studies Hybridoma Bank, University of Iowa; contrib- uted by Dr. D. Fischman), a monoclonal antibody di- rected against sarcomeric myosin (Bader e t al., 1982). Cells were also stained with ethidium bromide, a nu- clear stain. Briefly, after rinsing twice with cold MEM, cells were fixed for 30-60 seconds with cold alcohol/ formalin/acetic acid (AFA; 20 parts 70% ethanol, 2 parts formalin, and 1 part glacial acetic acid). Cells were then rinsed three times with MEM to wash off the AFA. The fixed cells were blocked with 1% normal goat serum in TBS for 2 hours to overnight. Cells were incu- bated with MF-20 (15 dilution in TBS) for 1-2 hours a t room temperature. After rinsing four times with TBS containing 0.05% Tween-20 (TBS-Tw; Sigma), the cells were incubated with secondary antibody, FITC-goat- anti-mouse IgG (Cappel) at 1:500 dilution in TBS. After 1-2 hours in the secondary antibody, the cells were stained with 0.01% ethidium bromide in TBS for 10 minutes. Cells were rinsed again three times and mounted in glycerol:TBS, 9:l. The number of nuclei in MF-20 positive cells, and the total number of nuclei, were counted using a Zeiss epifluorescence microscope. For each dish, more than 500 cells from randomly se- lected fields were counted. Results were analyzed using Student’s t-test (Brown and Hollander, 1977).
RESULTS Mouse CME is mitogenic for primary mouse
myoblasts and C2 cells To examine whether CME from adult mice contains
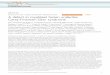
niyotrophic activity, myogenic cell cultures from 5-day- old mice were established a s described in Materials and Methods. The day after plating, experimental cultures were given 1.5 ml 10% FCS, plus 400 pg CME in a volume of 0.2 ml TBS per dish (final concentration 240 pg/ml). Control cultures were given 1.5 ml 10% FCS plus 0.2 ml TBS. The culture medium was changed again on day 3, and CME or TBS was re-added. The cells were fixed on day 5 for immunocytochemical and ethidium bromide staining. The effects of CME on total cell proliferation and on the number of differentiated muscle nuclei generated were quantitated by the counts of total nuclei per microscopic field, and by counts of nuclei in cells which stained positively for MF-20, a monoclonal antibody directed against muscle specific myosin. Cultures treated with CME showed a nearly three-fold increase in total nuclear number (Fig. lA), and a 12-fold increase in the number of nuclei in differentiated muscle cells (Fig. 1B). This could suggest a specific effect of the CME on myoblast proliferation, andlor an effect on differentiation per se. Photomicro- graphs illustrating the effects of CME on mouse pri- mary myogenic cultures are shown in Figure 2.
We also examined the mitogenic effects of increasing concentrations of CME on newborn mouse myogenic cultures by assaying incorporation of ’H-TdR. In this experiment, the primary cultures were enriched for myogenic cells by a 90 minute “preplating” procedure as described in Materials and Methods. The day after plating, the medium in the experimental cultures was changed to 1.5 ml10% FCS, plus increasing concentra- tions of CME (70-330 pgiml) in a volume of 0.2 ml TBS. Control cultures were changed to 1.5 ml 10% FCS plus 0.2 ml TBS. Two days later, cultures were given ‘H-
TdR at a final concentration of 0.5 pCiiml. After label- ing for 4 hours, the cultures were processed to deter- mine TCA-precipitable label incorporation. As shown in Figure lC, increasing concentrations of CME caused dose-dependent increases in ’H-TdR incorporation into DNA, the effect leveling off after 270 pgiml.
To examine whether CME is mitogenic for muscle tissue-derived fibroblasts, we obtained fibroblast cul- tures by repeated subculturing. Briefly, the initially adherent cells from preplating were grown to conflu- ence to allow any adherent myogenic cells to differenti- ate terminally and fuse. The cultures were then pas- saged. Infrequent myotube formation was observed after the secondary cultures reached confluence, indi- cating that the cultures were composed mostly of non- myogenic cells. These cells were passaged and plated at 1.0 x lo5 cells per dish. The effects of CME on fibro- blasts were examined by ’H-TdR incorporation assays, following the procedures used for primary m ogenic cultures. CME (240 pg/ml) did not stimulate ‘H-TdR incorporation into DNA by fibroblasts (Fig. lD), fur- ther suggesting that CME acts on myoblasts, and not on contaminating fibroblasts, to stimulate %IH-TdR incorporation in primary myogenic cultures. Fibroblast proliferation was stimulated in these conditions by the addition of bFGF (30 ngiml), but not by 100 ngiml of IGF-I (Fig. 1D).
The effects of CME on 0 2 cells, a mouse muscle satel- lite cell line (Yaffe and Saxel, 1977), were also exam- ined by ‘H-TdR incorporation assays and by cell counts. One day after plating, culture medium was changed to 2% FCS, plus CME in 0.2 ml TBS or an equal volume of TBS for control cultures. For ’H-TdR incorporation as- says, cultures were given ’H-TdR at a concentration of 0.5 $Xml on day 3 of culture. C2 cells were labeled for 4 hours, and the TCA-precipitable radioactivity incor- porated during the 4 hour period was measured. As shown in Figure 3A, CME administered at 214 pgiml increased 3H-TdR incorporation into DNA by C2 cells approximately three-fold compared to controls. For di- rect cell counts, sister cultures were fixed on day 3, stained with ethidium bromide, and the numbers of cells per microscopic field were counted. Cultures treated with CME contained twice as many cells as control cultures (Fig. 3B).
These data indicate that mouse CME caused a signif- icant increase in the total number of cells and in the number of differentiated cells in primary mouse myo- genic cultures. The effect appeared to be specific for the myogenic cells in primary cultures. CME also stimu- lated the proliferation of C2 myoblasts.
CME acts additively with saturating doses of bFGF on newborn mouse muscle cultures
Since CME stimulated myoblast proliferation, an ef- fect reported for bFGF (Linkhart et al., 1981; Spizz et al., 1986), we examined whether the activity of CME is due to the presence of bFGF, or a similar molecule which acts using the same receptors. We first deter- mined the dose-response curve of newborn mouse myo- genic cultures to bFGF, with or without the presence of CME, by means of ’H-TdR incorporation assays (Fig. 4A). Without CME (240 pgiml), the saturating amount of bFGF for primary mouse myogenic cultures was 1 ngiml, since adding more than 1 ngiml of bFGF did not

CHEN AND QUINN 566
4 150 LL
CME
T - LL w n. y 100 -J u 0 d
50 c
0
04 I
Fig. 1. The effects of mouse CME on the proliferation and differenti- ation of newborn mouse myogenic cultures. Bars indicate standard deviations. A The effect of CME on total cell proliferation, determined by nuclear counts. More than 500 nuclei from randomly selected fields were counted per sample. Experimental cultures were significantly different from control cultures (P < 0.0005; t-test). B The effect of CME on the number of differentiated nuclei generated, determined by counts of nuclei in MF-20 positive cells (MF-20 positive “cells”); more
lead to a higher response. In the presence of CME, the saturating amount of bFGF was reduced to 0.25 ngiml, suggesting that CME contains less than a saturating amount of bFGF (1 ngiml). Additionally, at all concen- trations of bFGF which were used, CME could signifi- cantly stimulate proliferation. Immunocytochemical staining for MF-20 and cell counts also confirmed that the effects of bFGF plus CME in stimulating newborn mouse myogenic culture proliferation and differentia- tion were higher than those obtained from saturating amounts of bFGF alone (Fig. 4B,C).
Similar results were also obtained when CME was tested in conjunction with 30 ngiml bFGF on C2 cells. This concentration of bFGF was determined to be well above the saturating dose for C2 cells (not shown). As assayed by 3H-TdR incorporation into DNA, the level of stimulation of C2 cell proliferation obtained from the combination of CME (240 pgiml) and bFGF (30 ng-iml) was higher than that obtained from bFGF (Fig. 4D).
These data show that CME can act in an additive fashion to saturating amounts of bFGF to stimulate mvoblast Droliferation. Therefore. the activity of CME
I I W 0 W
$ 20 ln 0 a 0 1 0 c.4 I L I 0
D n 25 (Fibroblasts)
30 bFGF h
x
2 20 V v CONTROL IGF-I d 1 5 IY 0 0
u U
I
g 1 0
7 5 n
0
than 500 nuclei from different fields were analyzed. CME treated cultures were significantly different from controls ( P < 0.0005; t-test). C: Mitogenic response of preplated newborn mouse myogenic cultures to increasing concentrations of mouse CME, assayed by 3H-TdR incor- poration into DNA. Each data point represents one culturc. D: Effects of CME on 3H-TdR incorporation of muscle-derived fibroblasts. IGF-I, bFGF, and CME were used at 100 ng/ml, 30 ng/ml, and 240 p,g/ml, respectively.
cannot be due solely to bFGF, and active factors other than bFGF must be present in CME.
The activity of CME is not due solely to the presence of IGF-I
IGF-I and IGF-I1 have also been shown t o stimulate proliferation of primary myoblasts (Dodson et al., 1985; Greene and Allen, 1991; Allen and Boxhorn, 1989). We examined whether IGF-I is present in CME, and whether the presence of IGF-I can account for the full activity of CME. First, we attempted to determine the dose-response curve of IGF-I on the proliferation of myogenic cultures from newborn mice. In 10% FCS, 2% FCS, or 5% HS, addition of IGF-I at concentrations of 0.5 ngiml to 200 ngiml did not produce any significant increases in ’H-TdR incorporation over controls. We also tested the activity of IGF-I in the presence of bFGF or dexamethasone, since bFGF and dexamethasone have been shown to potentiate the response of primary myogenic cultures (Greene and Allen, 1991; Dodson et al., 1985) to IGF-I. We did not observe any effects of IGF-I on primary mouse myogenic culture proliferation

CRUSHED MUSCLE EXTRACT 567
Fig. 2. Photomicrographs of preplated newborn mouse myogenic cul- tures grown for 5 days, with or without CME (240 pglml) treatment. Cultures were stained with ethidium bromide and MF-20, as described in Materials and Methods. A Control culture in 10%) PCS. B: Culture in 10% FCS with CME. C: Control culture in 5%, HS. D: Culture in 5% HS with CME.
in the presence of bFGF or dexamethasone (not shown). As assayed by MF-20 staining and nuclear counts, the combination of CME with 100 ng/ml IGF-I produced the same effects on primary cell proliferation and differen- tiation as CME alone (not shown).
Since a response of primary mouse myoblasts to IGF-I could not be obtained, we tested the activity of IGF-I on C2 cells. C2 cell proliferation was measured by the incorporation of 'H-TdR. Figure 5A shows the IGF-I dose-response curve of C2 cells, with or without CME. Without CME, the saturating dose of exogenously added IGF-I for C2 cell proliferation was 50 ngiml; with 240 pgiml CME, the saturating dose of exogenously added IGF-I was 3 ngiml. Additionally, the activity of CME was higher than that of IGF-I used at saturating doses. The combination of CME with saturating amounts of IGF-I (100 ngiml) produced a higher level of proliferation than CME or 100 ng/ml IGF-I alone. These findings indicate that factors other than IGF-I must be present in CME.
To examine the possible role of IGF-I1 on C2 cell proliferation, we determined dose-response curves for IGF-I1 on C2 cells (Fig. 5B), and compared the level of stimulation from saturating amounts of IGF-I1 with that obtained from saturating amounts of IGF-I (Fig. 5C). The saturating amount of IGF-I1 was 500 ngiml (Fig. 5B), 10 times as great as the saturating amounts
Of IGF-I (50 ngiml). However, the level of stimulation of 'H-TdR incorporation with saturating amounts of IGF-I1 was the same as the level with saturating amounts of IGF-I (Fig. 5C). Also, saturating amounts of IGF-I1 did not act additively to saturating amounts of IGF-I in stimulating C2 cell proliferation (Fig. 5C). These data confirm the findings of others that IGF-I1 binds to type-I IGF receptors, the receptors used by IGF-I, to mediate its effects on myoblast proliferation (Ewton et al., 1987; Duclos et al., 1991). Therefore, the possible role of IGF-I1 in CME stimulation of CZ cell proliferation was not pursued further.
The activity of CME is not due solely to the combination of bFGF and IGF-I
Allen and Boxhorn (1989) have reported that bFGF and IGF-I act additively on myoblasts to stimulate pro- liferation, We examined whether the full activity of CME could be explained by the combination of bFGF and IGF-I. Newborn mouse muscle cultures were given saturating doses of bFGF (30 ngiml) together with sat- urating doses of IGF-I (100 ngiml), with or without CME (240 pg/ml). Treatment of the myogenic cultures with bFGF plus IGF-I, or with CME alone, produced approximately 3.5-fold increases in cell number (Fig. 6A), and 12-fold increases in the number of nuclei in differentiated cells (Fig. 6B). However, treatment of cultures with CME, bFGF, and IGF-I caused approxi- mately 6.5-fold increases in total cell number (Fig. 6A), and 30-fold increases in the number of nuclei in differ- entiated cells (Fig. 6B). Therefore, the activity of CME cannot be due solely to the combination of bFGF and IGF-I. CME must contain factors other than bFGF and IGF-I which can act in a n additive fashion to saturating doses of these growth factors.
Using C2 cells, a similar observation was obtained. C2 cells were given saturating doses of bFGF (30 ngiml) combined with saturating doses of IGF-I (100 ngiml), with or without the presence of CME. The response in cell proliferation was measured by 3H-TdR incorpora- tion assays (Fig. 6C). The level of stimulation by bFGF and IGF-I combined with CME was higher than that of bFGF plus IGF-I (P < 0.025; t-test). As reported above for primary cells, these data indicate that CME can act additively on C2 cells to the combination of bFGF and IGF-I to stimulate proliferation.
CME must contain fac torb) other than PDGF, EGF, ACTH, M-CSF, and Tf
Besides FGF and the IGFs, factors which have been reported to be mitogenic for mammalian skeletal myo- blasts are PDGF (Jin et al., 1990; Yablonka-Reuveni et al., 19901, ACTH (Cossu et al., 19891, Tf (Shimo-Oka et al., 19861, and M-CSF (Jones et al., 1991). Additionally, we observed that EGF could stimulate 3H-TdR incorpo- ration by C2 cells cultured in 2% FCS (Fig. 7A). Figure 7A shows that the saturating dose of EGF on C2 cells was 1 ng/ml. Among three isoforms of PDGF, PDGF-BB is the most potent for myoblasts (Yablonka-Reuveni et al., 1990; J in et al., 1990). We examined whether EGF, PDGF-BB, ACTH, Tf, and M-CSF could account for the additivity of CME to bFGF and IGF-I. We observed that EGF, ACTH, and M-CSF, when used a t concentrations of 0.5 nglml to 200 ng/ml, did not stimulate "H-TdR

568 CIIEN AND QUINN
- 75.- w 0 - A CME
k 50 a CONTROL ._
Fig. 3. Mitogenic response of mouse C2 myoblasts to mouse CME, assayed by 3H-TdR incorporation into DNA (A) arid cell counts (B). C2 cells were cultured in 2% FCS. In iB), CME was used at 240 pgiml. Bars represent standard deviations; in several cases in (A), extent of deviation was smaller than size of data point symbols.
incorporation of C2 cells cultured in 0.5% FCS (not shown). Additionally, M-CSF (100 ngiml) did not stim- ulate C2 myoblasts cultured in 0.5% FCS to incorporate thymidine in the presence of 100 ngiml IGF-I or 240 pgiml CME (not shown). We determined dose-response curves of PDGF-BB and Tf on C2 cells. In agreement with an earlier report by Yablonka-Reuveni et al. (1990), PDGF-BB stimulated 3H-TdR incorporation by C2 myoblasts in a dose-dependent fashion (Fig. 7B). The saturating dose of PDGF-BB was 1 ngiml. Like- wise, Tfalso stimulated C2 cells to incorporate 3H-TdR, with a saturating dose of 1 pgiml (Fig. 7C).
Having determined the saturating doses of bFGF, IGF-I, PDGF-BB, and Tf, we then examined whether CME could act additively with the combination of satu- rating amounts of bFGF, IGF-I, PDGF-BB, EGF, Tf, ACTH (100 ngiml), and M-CSF (50 ngiml) to stimulate C2 cell proliferation. Figure 8A shows that the combi- nation of CME with bFGF (30 n g h l ) , IGF-I (100 ng/ ml), PDGF-BB (10 ng/ml), EGF (10 ngiml), Tf (10 pg/ ml), ACTH (100 ngiml), and M-CSF (50 ngiml) caused a higher level of stimulation than the combined growth factors without CME (P < 0.01; t-test). Similar results were obtained from studies on primary mouse myo- genic cultures (Fig. 8B). We conclude that CME must contain factors other than bFGF, IGF-I, PDGF, EGF, Tf, ACTH, and M-CSF.
Several lines of evidence indicate that Tf was present in CME at saturating or nearly saturating levels. We observed a 630% difference in stimulation of "H-TdR incorporation between the combinations of CME and six growth factors (bFGF, IGF-I, PDGF-BB, EGF, ACTH, M-CSF), and the six growth factors without CME (Fig. 9A). The difference was only 38% if Tf was included (Fig. 8A). Additionally, Tf did not act addi- tively to CME to stimulate 3H-TdR incorporation of C2 cells (Fig. 9B). Tf was detected in CME by Western blot. analysis (not shown), and was estimated to be present in CME at approximately 30 pgiml. Experiments using a neutralizing anti-Tf antibody also indicated that Tf was present in CME (Fig. 10A-C). The anti-Tf antibody completely inhibited Tf activity (Fig. 10A). The same antibody did not reduce the stimulation of 3H-TdR in-
corporation by combinations of bFGF, IGF-I, and EGF (Fig. 10B). Half of the mitogenic activity of CME could be neutralized by the antibody (Fig. lOC). However, even in the presence of the antibody, CME acted addi- tively to combinations of saturating doses of bFGF, IGF-I, and EGF on C2 cells cultured in 0.5% FCS (Fig. 1OC). Our previous studies have shown that when all were used at saturating doses, combinations of bFGF, EGF, and IGF-I caused the same level of stimulation of labeled thymidine incorporation as combinations of bFGF, EGF, IGF-I, PDGF-BB, ACTH, and M-CSF. Thus, when Tf activity is eliminated from the system, there is clear evidence for the presence of one or more mitogens which are distinct from known growth factors for myoblasts.
DISCUSSION In this report, we found that extracts from gently
crushed adult mouse muscles contain mitogens for pri- mary newborn mouse myoblasts and C2 cells, a myo- genic cell line from adult mouse muscle. Both cell counts and 'H-TdR incorporation assays showed that when CME was added to cultures of newborn mouse myoblasts or to C2 cells, cell proliferation was in- creased significantly over controls. In addition, when examined on day 5, newborn mouse myogenic cultures treated with CME showed higher numbers of termi- nally differentiated cells than cultures not treated with CME. We also compared the activity of CME with that of bFGF and IGF-I, the two most extensively docu- mented growth factors which act as mitogens for pri- mary myoblasts (Florini and Magri, 1989). We rea- soned that if the presence of bFGF and IGF-I could account for all the activity in CME, then the activity of CME plus saturating amounts of bFGF and IGF-I should be no greater than the activity of combinations of the two growth factors alone. Our data indicate that CME, with the presence of saturating amounts of bFGF and IGF-I, produced a greater stimulation of myoblast proliferation and differentiation than saturating doses of bFGF, IGF-I, or the combination of the two. These findings indicate that the full activity of CME cannot

CRUSHED MUSCLE EXTRACT 569
A h -- I ---P
with CME
I /I---$ -1- without CME
I I I 1 I A
O Z v : VI P:’?N w Vlovlo 0
bFGF (ng/rnl)
w 200
a u J
LL
CME + FGF
T
Fig. 4. A Dose-response curves of newborn mouse myogenic cultures to bFGF, with or without 240 yg/ml mouse CME, determined by “H- TdR incorporation assay. Cells were cultured in 10% FCS. B: The effects of CME (240 yg/ml), bFGF (30 ng/ml), and the combination of CME with bFGF on the proliferation of newborn mouse myogenic cultures, determined by counts of total nuclei per microscopic field after 5 days in culture. The numbers of nuclei (“cells”) in cultures treated with CME, bFGF, or the combination of CME plus bFGF were statistically different from each other (P < 0.0005; t-test). C The ef- fects of CME (240 ygiml), bFGF (30 ng/ml), and the combination of CME with bFGF on newborn mouse myogenic cultures determined by the counts of nuclei in MF-20 positive cells IMF-20 positive “cells”) after 5 days in culture. The numbers of nuclei in differentiated cells in
be due solely to the combination of bFGF and IGF-I. Factors other than bFGF and IGF-I must be present in CME.
We also investigated whether other known mitogens for myoblasts could account for the mitogenic effect of CME on myoblasts. Besides FGF and IGF-I, factors re- ported to stimulate mouse skeletal myoblast prolifera- tion are IGF-11, PDGF, Tf, ACTH, and M-CSF (Dodson et al., 1985; Yablonka-Reuveni et al., 1990a; Cossu et a]., 1989; Jones et a]., 1991). EGF has been shown to act as a progression factor to stimulate proliferation of BC3H1 myoblasts, which, however, have some proper- ties uncharacteristic of other widely studied skeletal muscle cell lines (Kelvin et al., 1989). Additionally, we have observed that EGF could stimulate ’H-TdR incor- poration of C2 myoblasts. Of the three isoforms of PDGF (PDGF-AA, PDGF-BB, PDGF-AB), only PDGF-AB and PDGF-BB are mitogenic for C2 cells, with PDGF-BB being the most potent (Yablonka-Reu-
CME + FGF B
500
n CME + FGF .,, .,
0
X - 170
I
140 v
a a:
Z
E
I I
r)
2 110 -
2 80
50
cultures treated with the combination of CME plus bFGF were signif- icantly different from the counts in cultures treated with bFGF alone, or CME alone (P < 0.0005; t-test). The numbers of nuclei in differenti- ated cells in CME treated cultures were significantly higher than in bFGF treated cultures (P .: 0.025; t-test). D: Mitogenic response of mouse C2 cells to 240 kg/ml CME, 30 ng/ml bFGF, and the combina- tion of the two, determined by ‘H-TdR incorporation assays. C2 cells wcrc culturedin 2% FCS. Label incorporation in CME treated cultures was significantly higher than in bFGF treated cultures (P < 0.025; t-test); incorporation in cultures treated with bFGF plus CME was statistically different from in cultures treated with CME alone, or bPGF alone (P < 0.025; t-test).
veni et al., 1990). We first examined whether IGF-I1 could explain the additivity of CME to combinations of bFGF and IGF-I. In agreement with earlier reports that IGF-I1 utilizes the type-I receptor to exert its mitogenic activity (Florini, 1987), we found that IGF-I1 stimu- lated ‘H-TdR incorporation of C2 cells to the same level as IGF-I, though requiring 10-fold higher doses of IGF-I1 to do so. We also found that saturating doses of IGF-I and IGF-I1 did not act additively. We concluded that IGF-I1 acts through the type-I IGF receptor to elicit mitogenic responses of C2 cells (Ewton et al., 1987; Duclos et al., 1991), so IGF-I1 could not explain the additivity of CME with combinations of bFGF and IGF-I. We also examined the mitogenic activity of M-CSF and ACTH. In contrast to the report of Jones et al. (19911, we did not observe any response in ’H-TdR incorporation of C2 myoblasts to M-CSF, either with or without the addition of IGF-I or CME to the cultures. ACTH was also not mitogenic for C2 myoblasts. We

570 CHEN AND QUINN
rr) - c
1507
IGF-I IGF-I1 IGF-I + IGF-II
m- 1 A o 130
0 5 70
T+--p----P
w
I/------ 5 0 1 without CMt m
30 ' I I I I I 1 1
0 3 5 10 50 100 200 IGF-I (ng/ml)
100 , I
w 7 *oJ
nl I - I
0 500 1000 1500 2000 IGF-I1 (ng/ml)
Fig. 5. Dose-response curves of C2 cells to IGF-I, with or without 240 pgiml CME (A), to IGF-I1 (B), and mitogenic activities of saturating amounts of IGF-I (100 ngiml), IGF-II(1,OOO ngimli, and the combina- tion of the two on C2 cells (C). In all cases, C2 cells were cultured in 2 8 FCS. In B and C, each data point represents one culture.
then examined the additivity of CME with saturating doses of bFGF, IGF-I, PDGF-BB, EGF, Tf, plus ACTH (100 ngiml) and M-CSF (50 ngiml). Our results show that addition of CME plus saturating amounts of these seven mitogens to C2 myoblast cultures caused a higher level of stimulation in 3H-TdR incorporation than the combination of the seven mitogens without CME, We concluded that CME must contain factors other than bFGF, the IGFs, PDGF, EGF, ACTH, M-CSF, and Tf. The identities of the active factors in
6oo I LL 400 Y
a n 500 t 0 -I
W
0 I
A CME + FGF + IGF
1
FGF + IGF-I
. CONTROL
250 T
CME + FGF + IGF
1 W n
! 150 FGF + IGF-I W 0 W > v)
100
2 8 50 I L I
0
LL Y
L
+ 0
L e + W I V
n 0 200 X
I
8 150
d
z
7 50 r
v
K 8 100
e U
rr)
0
Fig. 6. The effects of CME (240 Fg/ml), bFGF (30 ngiml) plus IGF-I (100 ngiml!, and the combination of the three on total cell number (A) and number of nulei in differentiated cells (MF-20 positive "cells" (B) of newborn mouse myogenic cultures, and 'H-TdR incorporation of C2 cells (C). Primary cells were cultured in 10% FCS, and C2 cells were cultured in 2%. FCS. The numbers obtained with the combination of CME, bFGF, and IGF-I were significantly higher than the numbers obtained with bFGP plus IGF-I in all cases CP <: 0.0005 in A and B, P s( 0.025 in C; t-test).
CME and the mechanisms employed by CME to stimu- late myoblasts are unknown. The extra factors in CME could act independently of known growth factors, or could act by potentiation of known growth factor activ- ities.

CRUSHED MUSCLE EXTRACT 57 1
h 75 M 0 - x 60 z 0 v 45 d K 0 y 3c
7 15
a
- a D
r rr)
A
5 10 20 50
EGF (ng/rnl)
d 150
u z K -0 1 0 0 1
I-
& 50 M
without CMI
o---
I I I I 1 2 5 10 20 50
PDGF-88 (ng/rnl)
120.’
x l a , u 80t/” J 1 1
- I V
Z 40 a D ll
rrl 2ot 0’11 I I I I I
1 2 5 10 20 50 100 TRANSFERRIN (pg/rnl)
Fig. 7. Dose-response curves of EGF (A), PDGF-BB (B), and Tf (C) on ‘H-TdR incorporation by C2 cells. The PDGF-BB dose-response curves were done with or without 240 p,g/ml CME. EGF dose-response curve was performed using C2 cells cultured in 2% FCS. PDGF-BB and Tf dose-responses were done on C2 cells in 0.5% FCS. Varying culture conditions were used in order to maximize the mitogenic effects of growth factors.
Besides the presence of unknown factors, CME also contains a mixture of some known growth factor activi- ties. Earlier studies have shown that Tf is present in
I C M F
TfPFElMA CME+ 1
400 B
CHE 200 -
CONTROL (107 FCS)
~
C M E t TfPFElMA
Fig. 8. Stimulation of proliferation ofC2 cells (A) and primary mouse mvoeenic cultures (B) bv CME (240 udml). combinations of saturat- ing Ydoses Tf (10 pgimlj, PDGF-BB ‘(‘‘P’, 10 ngiml), bFGF YF”, 30 ng/ml), EGF (“E”, 10 ng/mli, IGF-I ?I”, 100 nglml), M-CSF (‘‘M’, 50 nglml), and ACTH (“A> 100 ngiml), and CME plus the combined growth factors. C2 cell proliferation was quantitated using ”H-TdR incorporation assay. Proliferation of primary cultures was quantitated by counting the number of cells per microscopic field. C2 cells and primary mouse myogenic cultures were cultured in 0.5% FCS and 10% FCS, respectively. TPFEIMA + CME was significantly higher than TfPFEIMA (P < 0.01 for A and P < 0.0005 for B; t-test).
muscle tissue (Matsuda et al., 1984; Summers et al., 1985). Our data suggest that CME contains significant amounts of Tf. Using Western blot analysis, we ob- served Tf was present in CME and estimated the con- centration of Tf in CME was approximately 30 Fgimg CME protein (not shown). The amounts of CME used in the additivity studies contained saturating or close to saturating amounts of Tf for C2 myoblast cultures. In agreement with the Western blot data, CME showed no additivity to Tf, and greater additivity to the combina- tion of bFGF, IGF-I, EGF, ACTH, M-CSF, and PDGF-BB than to the combination of Tf, bFGF, IGF-I, EGF, ACTH, M-CSF, and PDGF-BB. Additionally, pre- liminary studies using radioimmunoassays indicate that CME contains a physiologically significant amount of IGF-I (not shown). Our studies did not ad- dress the sources of Tf and IGF-I present in CME. Ear- lier studies have examined the distributions of growth factors in various tissues. Tf has been found in blood circulation (Kohania and Ozawa, 1978), nerves (Mar- kelonis and Oh, 1979), interstitial cells present in mus- cle tissues, and to a lesser extent, in muscle fibers (Mat-

572 CHEN AND QUINN
- c) 0 - 320.- x
A CME + PFEIMA 200
h
*) 0 - 160 X
I Q
120 v
ti 8 80 z E
CL
40 I M
0
B CME + Tf CME
Fig 9. A Stimulation of 3H-TdR incorporation ofC2 cells by CME (240 p.g/ml), combinations ofsaturat- ing doses of PDGF-BB (“P, 10 ng/mlj, bFGF (“F, 30 ngiml), EGF (“E”, 10 ngirnl), IGF-I (“I”, 100 ngiml), M-CSF ( “ M , 50 ng/ml), and ACTH (“A”, 100 ng/ml), and CME plus the combined growth factors B: Stimulation of %TdR incorporation of C2 cells by Tf (10 pg/mll, CME (240 p.g/ml), or CME (240 pg/ml) plus Tf (10 pg/ml)
suda et al., 1984). These are all potential sources of Tf present in CME. IGF-I has been found in blood (Klap- per et al., 19831, muscle spindles, and nerve terminals and Schwann cells present in muscle tissue (Jennische et al., 1987). IGF-I present in CME could come from any of these sources. Jennische and Hansson (1987) did not detect IGF-I in uninjured adult mouse muscle fibers and satellite cells using a IGF-I specific antibody, and detectable IGF-I immunoreactivity did not appear until 15 hours after injury (Jennische and Hansson, 1987; Jennische et al., 1987). Therefore, muscle fibers and satellite cells are unlikely to be the primary source of IGF-I in CME collected immediately following muscle injury.
Besides Tf and IGF-I, CME is likely to contain small amounts of PDGF and FGF. PDGF has been shown to be released from platelets (Raines and Ross, 1982; Ross et al., 1986) present in the microvasculature in the muscle tissue from which CME was made. Our prelimi- nary studies using radioreceptor assays indicated the presence of PDGF in CME (not shown). However, the radioreceptor assays did not distinguish between the three isoforms of PDGF (AA, BB, and AB isoforms). The three isoforms have been shown to have different abili- ties to bind and stimulate C2 cell proliferation, with the BB isoform being the most potent one (Yablonka-Reu- veni et al., 1990a). CME may also contain bFGF, which is present in muscle tissue homogenates (Anderson et al., 1991). Immunocytochemical studies have localized bFGF to muscle fiber peripheries and muscle satellite cells (DiMario et al., 1989; Anderson et al., 1991). Our data presented here did not assay for bFGF. CME could stimulate proliferation of MM-14 cells, a myoblast cell line which requires bFGF to proliferate in serum free medium (Hauschka, personal communication). The amount of bFGF present in CME is likely to be small, however, as 240 pgiml CME did not stimulate prolifer- ation of mouse muscle-derived fibroblasts and chick myoblasts, whereas bFGF was mitogenic in sister cul- tures (data on chick myoblasts not shown).
As with all primary myogenic cultures, our mouse myogenic cultures contained nonmyogenic cells or fi- broblasts. Our data indicate that CME acts specifically
on myoblasts. CME caused an increase in the number of differentiated muscle nuclei. Also, CME did not stimu- late ‘H-TdR incorporation of muscle-derived fibro- blasts, but did stimulate proliferation of C2 myoblasts, presumably a homogeneous population of myogenic cells. Since bFGF was able to stimulate proliferation of primary fibroblasts, this lends further support to our findings that CME contains mitogens for myoblasts which are distinct from bFGF.
We did not observe a consistent effect of IGF-I on the proliferation of newborn mouse myogenic cultures. Greene and Allen (1991) reported that IGF-I did not significantly stimulate the proliferation of primary bo- vine myogenic cultures unless bFGF was present. Dod- son et al. (1985) also reported that dexamethasone could potentiate the effects of IGF-I on the proliferation of primary rat myogenic cultures. However, we could not observe mitogenic effects of IGF-I on the prolifera- tion of newborn mouse myogenic cultures in the pres- ence of bFGF or dexamethasone. The differences in the responsiveness to IGF-I among C2 cells, newborn mouse myoblasts, rat myoblasts, and bovine primary myoblasts could be caused by intrinsic differences in cells. C2 myoblasts, L6 myoblasts, and primary rat myogenic cultures have been shown to produce IGF (Tollefsen et al., 1989; Florini et al., 1991; HI11 et al., 1985). Therefore, primary mouse myoblasts may pro- duce IGF which can act in an autocrine manner. Activ- ity of exogeneous IGF would then be “masked” by the endogeneously produced IGF. Alternatively, serum- containing culture medium may contain high levels of IGF, or IGF inhibitors such as IGF binding proteins that act in a species specific manner (De Mellow and Baxter, 1988). In pilot experiments, we observed that IGF-I, which is mitogenic for C2 cells grown in 2% FCS, did not significantly stimulate the proliferation of C2 cells in 10% FCS (data not shown).
Our data are in general agreement with those re- ported by Bischoff (1986b, 1989, 1990) for rat muscle extract. Bischoff found that extracts from gently crushed adult rat muscles are mitogenic for embryonic rat myoblast cultures, as well as for single myofiber cultures. In the rat single myofiber culture system, sat-

CRUSHED MUSC :LE EXTRACT 573
(1989) suggested that multiple factors could be present in ra t CME. Bischoff did not test the mitogenic re- sponses of myoblasts to combinations of growth factors and CME. In this report, we found that the combination of CME with saturating doses of bFGF and IGF-I caused greater stimulation of myoblast proliferation than bFGF and IGF-I used together. Using a different strategy, our conclusions on mouse CME are in agree- ment with Bischoff s conclusion concerning rat CME; that is, CME contains factors other than bFGF and
Growth factors are thought to play a pivotal role in vivo in regulating the proliferation and differentiation of embryonic myoblasts and satellite cells following tis- sue injury. However, direct evidence showing that growth factors which are active in tissue culture condi- tions play physiological roles in vivo is lacking. CME provides a good model system to investigate the factors released during muscle tissue injury. In this study, we have shown that CME contains a combination of growth factors, some of which are known (TO and some of which are unknown. Each factor may play a different role in the process of muscle regeneration. Bischoff (198613) has reported that extracts from crushed mus- cle, but not from uncrushed muscle, are mitogenic for satellite cells. Thus “crushing” may mimic tissue injury processes, and factors present in crushed muscle ex- tract are likely to be the factors released during normal tissue injury. Further identification and characteriza- tion of the factors present in crushed muscle extracts should allow better understanding of the roles t.he growth factors play in vivo in muscle injury and repair.
IGF-I.
1 2 0 T I* h
R
E l ’ x
I P 0 v
6o t ti
z_
e 0 V “1 CONTROL Tf + Ab
- 2o 0 t 300 T
FEI + 11 R 0
X
-
150 t I p. V
Q
r K 0 V
FEI + Ab
FEI
C 0 N T R 0 L ’ O O L 50
0
CL u #-
I c)
- W LL
+ Q 4 +
300 T h
n x 4 W
u 9 2 + E
c)
0 W I u
X
I 0 V Y
ACKNOWLEDGMENTS We thank Dr. Z. Yablonka-Reuveni for C2 cells
(through Dr. D. Yaffe), Rebecca Hartley for procedures for MF-20 antibody staining, and Dr. D. Bowen-Pope for providing PDGF. We are also grateful to Dr. 0. Mejino who helped with the initial mouse studies. The MF-20 antibody was obtained from the Developmental Studies Hybridoma Bank maintained by the Depart- ment of Pharmacology and Molecular Sciences, Johns Hopkins University School of Medicine, Baltimore, MD, and the Department of Biology, University of Iowa, Iowa City, IA, under contract N01-HD-6-2915 from the NICHD. This work was supported by grants to L.S.Q. from Eli Lilly and Company, Association Francaise Contre Les Myopathies, Institutional Cancer Grant IN-26-21 from the American Cancer Society, and the Cooperative State Research, U S . Department of Agriculture (Agreement #90-37265-5456).
150 *0° t o w
- W
Q 0 I- I I
c)
9-
Fig. 10. Effects of‘ anti-Tf Ab (150) on the stimulation of “H-TdR incorporation of C2 cells by Tf (10 pgiml) (A), combinations of bFGF (“F, 30 ngimt), EGF (“E”, 10 ngiml), and IGF-I (“I”, 100 ngiml) (B), and CME (240 pgiml) (C). C also shows the effect of a higher dose of anti-Tf Ab (1:38) on CME stimulation, and the additivity of CME to combinations of bFGF (30 nglml), EGF (10 ngiml), and IGF-I (100 ngiml) in the presence of anti-Tf Ab (1:50). All experiments were done on C2 cells in 0.56 FCS.
ellite cells remain in a position between the basal lam- ina and the myofiber, and remain mitotically quiescent in 5% HS for up to 4 days. Upon addition of CME, rat satellite cells are stimulated to enter the cell cycle and proliferate. Among many known growth factors, only bFGF was found to be mitogenic for rat satellite cells in the single myofiber cultures (Bischoff, 1986b). How- ever, the properties of rat CME differ from those of bFGF in terms of molecular weight and the salt concen- tration required to elute the activity from heparin-af- finity columns (Bischoff, 1989). The identity of the ac- tive factors in rat CME is not known, although Bischoff
LITERATURE CITED Allen, R.E., and Boxhorn, L.K. (79891 Regulation of skeletal muscle
satellite cell proliferation and differentiation by transforming growth factor-beta, insulin-like growth factor I, and fibroblast growth factor. J. Cell. Physiol., 138.311-315.
Allen, R.E., Dodson, M.V., and Lutein, L.S. (19841 Regulation ofskel- eta1 muscle satellite cell proliferation by bovine pituitary fibroblast growth factor. Exp. Cell. Res., 152:154-160.
Anderson, J.E., Liu, L., and Kardami, E. (19911 Distinctive patterns of basic fibroblast growth factor (bFGF) distribution in degenerating and regenerating areas of dystrophic (mdx1 striated muscles. Dev. Biol., 147.96103.
Bader, D., Masaki, T., and Fischman, D.A. (1982) Immunochemical

574 CHEN AND QUINN
analysis of myosin heavy chain dring avian myogenesis in vivo and in vitro. J. Cell. Biol., 95r763-770.
Bischoff, R. (1986a) Proliferation of muscle satellite cells on intact myofibers in culture. Dev. Biol., 115t129-139.
Bischoff, R. (198613) A satellite cell mitogen from crushed adult mus- cle. Dev. Biol., 115t140-147.
Bischoff, R. (1989) Analysis of muscle regeneration using single myo- fibers in culture. Med. Sci. Sports. Exerc. 21 :S164-S172.
Bischoff, R. (1990) Cell cycle commitment of rat satellite cells. J. Cell. Biol., 111r201-207.
Bradford, M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principal of protein- dye binding. Anal. Biochem., 72:248-254.
Brown, B.W., and Hollander, M. (1977) Statistics: A Biomedical Intro- duction. John Wiley & Sons, Inc., New York.
Campion, D.R. (1984) The muscle satellite cells: A review. Int. Rev. Cytol., 87225-251.
Carlson, B.M., and Faulkner, J.A. (1983) The regeneration of skeletal muscle fibers following injury: A review. Med. Sci. Sports Exerc., 15r187-198.
Cossu, G., Cuselia-De Angelis, M.G., Senni, M.I., De Angelis, L., Vi- varelli, E., Vella, S., Bouche, M., Boitani, C., and Molinaro, M. (1989) Adrenocorticotrophin is a specific mitogen for mammalian myogenic cells. Dev. Biol., 131:331-336.
De Mellow, J.S.M., and Baxter R.C. (1988) Growth hormone-depen- dent insulin-like growth factor (IGF) binding proteins both inhibit and potentiate IGF-I stimulated DNA synthesis in human skin fi- broblasts. Biochem. Biophys. Res. Cornmun., 156:199-204.
DeMario, J., and Strohman, R.C. (1988) Satellite cells from dystrophic (mdx) mouse muscle are stimulated by fibroblast growth factors in vitro. Differentiation, 39r42-49.
DiMario, J . , Buffinger, N., and Strohman, R.C. (1989) Fibroblast growth factor in the extracellular matrix of dystrophic (mdx) mouse muscle. Science, 244:68%690.
Dodson, M.V., Allen, R.E., and Hossner, K.L. (1985) Ovine somatome- din, multiplication-stimulating activity, and insulin promote skele- tal muscle satellite cell proliferation in vitro. Endocrinology, 11 7:2357-2363.
Duclos, M.J., Wilkie, R.S., and Goddard, C. (1991) Stimulation of DNA synthesis in chick muscle satellite cells by insulin and insulin-like growth factors: Evidence for exclusive mediation by a type-I insulin- like growth factor receptor. J. Endocrinol., 128:3542.
Ewton, D.Z., Falen, S.L., and Florini, J.R. (1987) The type I1 IGF receptor has low affinity for IGF-I analogs: Pleiotypic actions of IGFs on myoblasts are apparently mediated by the type I receptor. Endocrinology, 120: 115-124.
Florini, J.R. (1987) Hormonal control of muscle growth. Muscle Nerve, l0:577-598.
Florini, J.R., and Magri, K.A. (1989) Effects of growth factors on myo- genic differentiation. Am. J. Physiol., 256tC701-C711.
Florini, J.R., Magri, K.A., Ewton, D.Z., James, P.L., Grindstaff, K., and Rotwein, P.S. (1991) “Spontaneous” differentiation of skeletal myoblasts is dependent upon autocrine secretion of insulin-like growth factor 11. J. Biol. Chem., 266t15917-16923.
Greene, E.A., and Allen, R.E. (1991) Growth factor regulation of bo- vine satellite cell growth in vitro. J. Anim. Sci., 69r14G152.
Hill, D.J., Crace, C.J., Nissley, S.P., Morell, D., Holder, A.T., and Milner, R.D.G. (1985) Fetal rat myoblasts release both rat so- matomedin-C EM-C)/insulin-like growth factor I (IGF-I) and multi- plication-stimulating activity in vitro: Partial characterization and biological activity of myohlast-derived SM-CIIGF-I. Endocrinology, 117t2061-2072.
Jennische, E., and Hansson, H.-A. (1987) Regenerating skeletal mus-
cle cells express insulin-like growth factor I. Acta Physiol. Scand., 190327-331.
Jennische, Ey,Skottner, A., and Hansson, H-A. (1987) Satellite cells express the trophic factor IGF-I in regenerating skeletal muscle. Acta Physiol. Scand., 129:9-15.
Jin, P., Rahm, M., Claesson-Welsh, L., Heldin, C.H., and Sejerson, T. (1990) Expression of PDGF A-chain and B-receptor genes during rat myoblast differentiation. J. Cell Biol., 110:1665-1672.
Jones, G.E., Murphy, S.J., Wise, C., and Watt, D.J. (1991) Macro- phage-colony-stimulating factor (CSF-1) stimulates proliferation of myogenic cells. J . Cell. Biochem., Supplement 15Cr39.
Kelvin, D.J., Simard, G., and Connolly, J.A. (1989) FGF and EGF act synergistically to induce proliferation in BC3H1 myoblasts. J. Cell. Physiol., 138:267-272.
Klapper, D.G., Svoboda, M.E., and Van Wyk, J.J. (1983) Sequence analysis of somatomedin-C: Confirmation of identity with insulin- like growth factor-I. Endocrinology, 112r2215-2217.
Kohama, K., and Ozawa, E. (1978) Muscle trophic factor: 11. Ontogenic development of activity of a muscle trophic activity in chicken se- rum. Muscle Nerve, 1:236-241.
Linkhart, T.A., Clegg, C.H., and Hauschka, S.D. (1981) Myogenic differentiation in permanent clonal mouse myoblast cell lines: Reg- ulation by macromolecular growth factors in culture medium. Dev. Biol., 86:1930.
Markelonis. G., and Oh, T.H. (1979) A sciatic nerve Drotein has a trophic effect on development and maintenance of skeletal muscle cells in culture. Proc. Natl. Acad. Sci. USA, 76r2470-2474.
Matsuda, R., Spector, D., Micou-Eastwood, J., and Strohman R.C. (1984) There is selective accumulation of a growth factor in chicken skeletal muscle. Dev. Biol., 103276-284.
Mauro, A. (1961) Satellite cells of skeletal muscle fibers. J. Biophys. Biochem. Cytol., 9t493495.
Raines, E.W., and Ross, R. (1982) Platelet-derived growth factor. I. High yield purification and evidence for multiple forms. J. Biol. Chem., 257515P5160.
Ross. R., Raines, E.W., and Bowen-Pow, D.F. (1986) The biolom of I”
platelet-derived growth factor. Cell, 26155-169. Schultz, E., Gibson, M.C., and Champion, T. (1978) Satellite cells are
mitotically quiescent in mature mouse muscle: An EM and radioau- tographic study. J. Exp. Zool., 206t451-455.
Shimizu, M., Webster, C., Morgan, D.O., Blau, H.M., and Roth. R.A. (1986) Insulin and insulinlike growth factor receptors and responses in cultured human muscle cells. Am. J. Physiol., 251:E611-E615.
Shimo-Oka, T., Hagiwara, Y. , and Ozawa, E. (1986) Class specificity of transferrin as a muscle trophic factor. J. Cell. Physiol., 126r341- 351.
Spizz, G., Roman, D., Straws, A., and Olson, E.N. (1986) Serum and fibroblast growth factor inhibit myogenic differentiation through a mechanism dependent on protein synthesis and independent of cell proliferation. J. Biol. Chem., 261 :9483-9488.
Summers, P.J., Ashmore, C.R., Lee, Y.B., and Ellis S. (1985) Stretch- induced growth in chicken wing muscles: Role of soluble growth- promoting factors. J. Cell. Physiol., 125.288-294.
Tollefsen, S.E., Lajara, R., McCusker, R.H., Clemmons, D.R., and Rot- wein. P. (1989) Insulin-like growth factors (IGF) in muscle develou- ment, J. Biol. Chem., 264t13Y810-13817.
Yablonka-Reuveni, Z. , Balestreri, T.M., and Bowen-Pope, D.F. (1990) Regulation of proliferation of myoblasts derived from adult mouse skeletal muscles by specific isoforms of PDGF. J. Cell. Biol., 111 :1623-1629.
Yaffe, D., and Saxel, 0. (1977) Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature, 270325-727.