Embed Size (px)
Citation preview

Papers in Press
“Papers in Press” includes peer-reviewed, accepted manuscripts of research
articles, reviews, and short notes to be published in Paleontological Research.
They have not yet been copy edited and/or formatted in the publication style of
Paleontological Research. As soon as they are printed, they will be removed from
this website. Please note they can be cited using the year of online publication
and the DOI, as follows:
Humblet, M. and Iryu, Y. 2014: Pleistocene coral assemblages on Irabu-jima,
South Ryukyu Islands, Japan. Paleontological Research,
doi: 10.2517/2014PR020.

Accepted manuscript
1
doi:10.2517/2017PR001
Radiolarian biostratigraphy from middle Miocene to late Pleistocene
in the Japan Sea
SHIN-ICHI KAMIKURI 1, TAKUYA ITAKI 2, ISAO MOTOYAMA 3 AND KENJI M.
MATSUZAKI 4
1Faculty of Education, Ibaraki University, 2-1-1 Bunkyo, Mito, Ibaraki 310-8512, Japan
([email protected]) 2AIST, Geological Survey of Japan, 1-1-1 Higashi, Tsukuba, Ibaraki 305-8567, Japan 3Department of Earth and Environmental Sciences, Yamagata University, 1-4-12
Kojirakawa-machi, Yamagata 990-8560, Japan
4Department of Earth and Planetary Science, Graduate School of Science, the
University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan
Abstract. In the Integrated Ocean Drilling Program (IODP) Exp. 346, sampling by
drilling was conducted at seven sites (U1422–U1427 and U1430) in the Japan Sea.
Radiolarians in moderately well preserved states were found in most samples
throughout the sequence in varying abundance. Forty-one radiolarian datum
events were identified in this study, and the radiolarian zonation that best divides
the middle Miocene to Pleistocene sequences with updated ages of radiolarian
datum events (estimates based on the geomagnetic polarity time scale (GTS) 2012)
was applied to the sedimentary sequences in the Japan Sea. Here, four new
radiolarian zones are proposed for the Quaternary of the Japan Sea, and one zone
is slightly revised to adjust for differences among other zones. The sequences
collected at the sites extended from the Pleistocene Ceratospyris borealis Zone to
progressively deeper zones as follows: Site U1427, four zones to the Pleistocene
Schizodiscus japonicus; Site U1422, six zones to the late Pliocene Hexacontium

Accepted manuscript
2
parviakitaense Zone; Sites U1423, U1424 and U1426, eight zones to the early
Pliocene Larcopyle pylomaticus Zone; and Sites U1425 and U1430, fourteen zones
to the middle Miocene Eucyrtidium inflatum Zone. The absence or extremely rare
occurrence of Stylatractus universus and E. matuyamai indicates that S. universus
lived in the deep water of the northwestern Pacific and had not been able to
migrate into the Japan Sea across the Tsugaru Strait since the Pliocene.
Key words: IODP Exp. 346, Japan Sea, middle Miocene to Quaternary,
radiolarian zonation, sedimentation rate
Introduction
The recently conducted Integrated Ocean Drilling Program (IODP) Expedition 346
recovered sedimentary sequences with a total length of 6135.3 m from cores taken from
the sea floor at seven sites in the Japan Sea (Tada et al., 2015). The sediment samples
offer an ideal record for understanding the atmosphere-ocean linkage during the late
Neogene and Quaternary that are related to the East Asian monsoonal system. The
drilling sites were selected to achieve a depth transect of cores off the Japanese Islands
in the Japan Sea in order to collect information on depth-dependent sedimentation,
ocean chemistry, and marine organisms from the middle Miocene to Quaternary.
Radiolarians are planktonic unicellular Protoctista, and inhabit shallow-to-deep
waters at depths as deep as 5,000 m in the modern ocean (Reshetnyak, 1955;
Petrushevskaya, 1971). The opal skeletons are well preserved in the sea-floor sediments
and have been widely used as a proxy to reconstruct paleoceanographic history and to
determine the geological age to a resolution of 1,000–10,000 years. The main objectives
of this study are to establish radiolarian biostratigraphy for more precise dating and to

Accepted manuscript
3
establish correlations among sedimentary sequences drilled at IODP Sites
U1422–U1427 and U1430 in the Japan Sea.
Material and methods
Techniques
In this study, a total of 559 core samples were collected for biostratigraphic analysis
from IODP Exp. 346 Sites U1422–U1427 and U1430 in the Japan Sea (Figure 1). The
lithology of these cores is described in detail by Tada et al. (2015). Here locality and
lithology are briefly described. Sample preparation was conducted following the
methods described in Itaki (2001a). All samples were freeze-dried, weighed, and then
treated with 10% H2O2 solution to remove organic matter and 10% HCl to remove
calcium carbonate. If the samples contained large quantities of lithologenic grains, opal
particles were extracted from the residues using the elutriation method (Itaki, 2003).
The residues were then wet sieved through a 45-µm mesh, dried on a slide, mounted
with Norland optical adhesive, and covered with a 22 mm × 40 mm cover glass. The
adhesive was solidified by placing the slide under a UV light (300–400 nm) for
approximately 15 minutes. Original data for Sites U1424, U1426 and U1427 were
reported by Tada et al. (2015).
Abbreviations for radiolarian abundances in a slide are A, abundant (>10,000
specimens per slide); C, common (2,000–10,000 specimens per slide); F, few
(500–2,000 specimens per slide); R, rare (50–500 specimens per slide); VR, very rare
(<50 specimens per slide); and B, barren (0 specimens per slide).
The relative abundance of individual radiolarian species is designated as A,
abundant (≥16% of the radiolarian assemblage); C, common (4–16% of the radiolarian
assemblage); F, few (1–4% of the radiolarian assemblage); R, rare (0.2–1% of the

Accepted manuscript
4
radiolarian assemblage); and P, present (<0.2%).
Preservation of the radiolarian specimens was recorded as G (good), only minor
fragmentation; M (moderate), obvious fragmentation but identification of species not
impaired; and P (poor), individual taxa exhibited considerable fragmentation, and
identification of some species was not possible.
Radiolarian zonation
Radiolarian zonation was first introduced by Nakaseko and Sugano (1973) based on
samples from the Japan Sea (Figure 2). Four assemblage zones extending from late
early Miocene to Pliocene were based on earlier studies of sediment samples collected
from onshore Japan by Nakaseko and co-workers (Nakaseko, 1954, 1960; Nakaseko et
al., 1965, 1972; Sugano and Nakaseko, 1968, 1971). Later, Funayama (1988)
distinguished four interval zones including two new zones and two new interval
subzones from the late early Miocene to early late Miocene in the Noto Peninsula
(central Japan). Ling (1975, 1992) and Alexandrovich (1992) reported occurrences of
radiolarians in Deep Sea Drilling Project (DSDP) Leg 31 and Ocean Drilling Program
(ODP) Legs 127 and 128 in the Japan Sea, but they proposed no radiolarian zonation for
the late Miocene to Quaternary. Tochilina (1985) and Vasilenko (2015) distinguished
four new assemblages from the late Miocene to the late Pliocene in the Japan Sea,
respectively. Motoyama (1996) defined eight radiolarian zones, including seven new
zones, for an interval between the upper Miocene and the lower Pleistocene based on
the DSDP Leg 31 Site 302 sequence (40°20.13′N and 136°54.01′E; water depth, 2399
m) in the Japan Sea. Itaki (2001b) recognized four new assemblage zones during the last
30 kyr based on piston cores taken off the Oga Peninsula in the Japan Sea. In this study,
the radiolarian zonation for the Japan Sea follows mainly Funayama (1988), Motoyama
(1996) and Itaki (2001b).

Accepted manuscript
5
Radiolarian datum levels
Not all radiolarian species identified in this study were used (Table 1). Most of the
radiolarian species selected for their stratigraphic usefulness were previously described
in Japan Sea and North Pacific sediments by other investigators (Foreman, 1973, 1975;
Kling, 1973; Ling, 1973; Sakai, 1980; Weaver et al., 1981; Wolfart, 1981; Popova,
1989; Spencer-Cervato et al., 1993; Vitukhin, 1993; Bak et al., 2012). In the subarctic
North Pacific, over 60 radiolarian datum levels have been directly tied to the
paleomagnetostratigraphy since the early Miocene (Morley and Nigrini, 1995; Shilov,
1995; Kamikuri et al., 2007; Kamikuri, 2010). In the temperate North Pacific, only
Pliocene and Pleistocene datum levels were directly tied to the
paleomagnetostratigraphy (Hays, 1970; Reynolds, 1980; Morley, 1985; Motoyama,
1996; Kamikuri et al., 2004). Recently, 17 radiolarian bioevents from the middle to late
Pleistocene were calibrated to the benthic foraminiferal oxygen isotope curves in the
temperate Northwest Pacific (Matsuzaki et al., 2014). In the Sea of Okhotsk, the late
Quaternary datum levels were precisely calibrated to the oxygen isotope curves
measured in the benthic foraminifera (Matul et al., 2002, 2009). In the Bering Sea,
radiolarian datum levels since the late Pliocene were tied to the
paleomagnetostratigraphy and the cyclostratigraphy of oxygen isotopes and Si/Al ratios
(Ikenoue et al., 2016). Only late Quaternary datum levels in the Japan Sea, on the other
hand, were indirectly correlated to the oxygen isotope stratigraphy (Itaki et al., 2007).
In this study, the correlation between magneto-oxygen isotope stratigraphy and
radiolarian datum levels mainly follows Matul et al. (2002), Kamikuri et al. (2004,
2007), Itaki et al. (2007), Kamikuri (2010), and Ikenoue et al. (2016) (Table 1). The
ages of the biohorizons used in this study are updated based on the geologic time-scale
(GTS) of Ogg (2012) by extrapolation of each horizon within each magnetic chron.

Accepted manuscript
6
Radiolarian datum levels and zonation in the Japan Sea
A summary of the radiolarian datum levels identified in this study is presented in
Table 1. The following abbreviations are used in this study to express events; FO, first
occurrence; LO, last occurrence; RI, rapid increase; RD, rapid decrease. The concept of
evolutionary transition (ET) follows that of Sanfilippo and Nigrini (1998).
The Japan Sea has developed a unique radiolarian fauna, especially since the late
Pliocene Epoch when the sea became more isolated from the Pacific Ocean due to
enhanced uplift of the Japanese archipelago (Kamikuri and Motoyama, 2007; Itaki,
2016). This makes it difficult to apply radiolarian zones established for the North
Pacific to the marginal sea. Thus, we here propose a local radiolarian zonal scheme for
the Quaternary Period in the Japan Sea.
The sedimentary sequences were divided into six zones from the Ceratospyris
borealis Zone to the Hexacontium parviakitaense Zone at Site U1422, eight zones from
the C. borealis Zone to the Larcopyle pylomaticus Zone at Sites U1423, U1424 and
U1426, fourteen zones from the C. borealis Zone to the Eucyrtidium inflatum Zone at
Sites U1425 and U1430, and four zones from the C. borealis to the Schizodiscus
japonicus Zone at Site U1427 (Figures 2–4, Table 1).
Ceratospyris borealis Zone (Itaki, 2001b; emend. herein)
Category.—Interval zone.
Definition.—Interval between the RD of Ceratospyris borealis (top) and the LO of
Lychnocanoma sakaii (base).
Age.—Late Pleistocene (54–15 ka).
Remarks.—Itaki (2001b) defined four assemblage zones: Larcopyle buetschlii Zone
(0–10.5 ka), Stylochlamydium venustum Zone (10.5–13 ka), Spongodiscus resurgens
Zone (13–15 ka), and Ceratospyris borealis Zone (15–28 ka). The C. borealis Zone was

Accepted manuscript
7
defined by abundant occurrence of C. borealis, but the lower limit of this zone was not
recognized (Itaki, 2001b). In the present study, the C. borealis Zone is employed as an
interval zone, and the LO of L. sakaii is taken as the lower limit of this zone.
Lychnocanoma sakaii Zone (herein)
Category.—Interval zone.
Definition.—Interval from the LO of Lychnocanoma sakaii (top) to the LO of
Amphimelissa setosa (base).
Age.—Late Pleistocene (0.085–0.054 Ma).
Remarks.—It is difficult to determine the LO of Amphimelissa setosa at Site 1424
because the species has a very sparse and sporadic occurrence.
Amphimelissa setosa Zone (herein)
Category.—Interval zone.
Definition.—Interval from the LO of Amphimelissa setosa (top) to the LO of
Schizodiscus japonicus (base).
Age.—Middle to Late Pleistocene (0.29–0.085 Ma).
Remarks.—Hays (1970) originally defined the Stylatractus universus Zone as an
interval zone from the LO of S. universus (top) to the LO of Eucyrtidium matuyamai
(base) for the subarctic North Pacific. Later, Ikenoue et al. (2016) changed the zonal
name from S. universus Zone to Amphimelissa setosa Zone because A. setosa was the
most abundant and characteristic species during the late Quaternary in the southern
Bering Sea. In the North Pacific, there are two radiolarian zones with the same
definition. ISSC (1994) clarified the nomenclature rules for zones, stating that the taxon
for which the zone is named does not have to be a common taxon within the zone or be
a particularly good guide-fossil among the members of the assemblage. Following this
rule, different names should not be assigned to the same biostratigraphic unit, and the A.

Accepted manuscript
8
setosa Zone of Ikenoue et al. (2016) is not accepted. Because the S. universus Zone has
been duly proposed as an unequivocal designation of the zonal limits by Hays (1970),
we follow the definition and define the A. setosa Zone as a new zone here.
Schizodiscus japonicus Zone (herein)
Category.—Interval zone.
Definition.—Interval from the LO of Schizodiscus japonicus (top) to the LO of
Axoprunum acquilonium (base).
Age.—Early to Middle Pleistocene (1.7–0.29 Ma).
Secondary events.—FO of Amphimelissa setosa and very short-ranging of
Eucyrtidium matuyamai.
Remarks.—In this study, Eucyrtidium matuyamai was not observed at Sites U1423,
U1425, U1427 and U1430, likely due to low stratigraphic resolution.
Hexacontium minerva Zone (herein)
Category.—Interval zone.
Definition.—Interval from the LO Axoprunum acquilonium (top) to the ET from
Cycladophora sphaeris to C. davisiana (base).
Age.—Early Pleistocene (2.6–1.7 Ma).
Secondary events.—LO of Cycladophora sphaeris.
Remarks.— This zone approximately correlates with the Spongodiscus osculosus
Zone of Tochilina (1985) and the Cycladophora davisiana Assemblage of Vasilenko
(2015).
Hexacontium parviakitaense Zone (Kamikuri, in press; emend. herein)
Category.—Interval zone.
Definition.—Interval between the ET from Cycladophora sphaeris to C. davisiana

Accepted manuscript
9
(top) and the LO of Dictyophimus bullatus (base).
Age.—Early to late Pliocene (3.9–2.6 Ma).
Secondary events.—FO and LO of Hexacontium parviakitaense and FO of
Cycladophora davisiana.
Remarks.—Motoyama (1996) defined the Cycladophora sphaeris Zone (=
Cycladophora sakaii Zone in the original paper) as an interval from the FO of
Eucyrtidium matuyamai (top) to the LO of Dictyophimus bullatus (base). However, E.
matuyamai shows sporadic occurrence, probably due to a very short biostratigraphic
range in the Japan Sea, making it difficult to recognize the FO and LO in Sites U1423,
U1425, U1427 and U1430. Hence, the C. sphaeris Zone was not assigned in the Japan
Sea.
The Cycladophora sphaeris Zone was subdivided into two subzones in the North
Pacific (Kamikuri, in press): Hexacontium dionysus and H. parviakitaense Subzones. In
this study, we use the latter subzone as a zone between the H. minerva Zone and
Dictyophimus bullatus Zone in the Japan Sea. Kamikuri (in press) defined the first
consistent occurrence of Cycladophora davisiana as the upper limit of the H.
parviakitaense Subzone (H. parviakitaense Zone in the present paper). As Kamikuri (in
press) already mentioned, the ET from Cycladophora sphaeris to C. davisiana is used
for the upper limit of this zone in the Japan Sea. The H. dionysus Subzone correlates the
lower part of the H. minerva Zone.
The LO of Hexacontium parviakitaense closely approximates to the top of this zone,
and the FO is also a good marker for the base of this zone (Kamikuri, 2010).
Dictyophimus bullatus Zone (Motoyama, 1996; rename. Kamikuri et al., 2004)
Category.—Taxon range zone.
Definition.—Taxon range of Dictyophimus bullatus.
Age.—Early Pliocene (4.4–3.9 Ma).

Accepted manuscript
10
Secondary events.—RD of Siphocampe arachnea group.
Remarks.—The FO and LO of Dictyophimus bullatus was not recognized at Sites
1424 and 1426, due to very sparse and sporadic occurrence.
Larcopyle pylomaticus Zone (Motoyama, 1996)
Category.—Interval zone.
Definition.—Interval from the FO of Dictyophimus bullatus (top) to the FO of
Larcopyle pylomaticus (base).
Age.—Late Miocene to early Pliocene (5.4–4.4 Ma).
Remarks.—This zone is divided into the next two subzones. Kamikuri (in press) used
the LO of L. redondoensis as a subzone marker (not zone), because of its usefulness as a
biostratigraphic marker in latitudes higher than 38°N of the North Pacific.
Lamprocyclas hannai Subzone (Kamikuri, in press)
Category.—Interval zone.
Definition.—Interval from the FO of Dictyophimus bullatus (top) to the LO of
Lipmanella redondoensis (base).
Age.—Early Pliocene (4.9–4.4 Ma).
Secondary events.—RI of Siphocampe arachnea group.
Remarks.—The RI of Siphocampe arachnea group is a good marker for the middle
part of this subzone (Kamikuri et al., 2004).
Lithelius klingi Subzone (Kamikuri, in press)
Category.—Interval zone.
Definition.—Interval from the LO of Lipmanella redondoensis (top) to the FO of
Larcopyle pylomaticus (base).
Age.—Late Miocene to early Pliocene (5.4–4.9 Ma).

Accepted manuscript
11
Remarks.—The coinciding recognition of the LO of Lipmanella redondoensis and the
FO of Larcopyle pylomaticus near 79.0 m at Site U1430 (Table 1, Figure 3) indicates
that the Lithelius klingi Subzone is absent in the studied core.
Axoprunum acquilonium Zone (Shilov, 1995; emend. Motoyama, 1996)
Category.—Interval zone.
Definition.—Interval from the FO of Larcopyle pylomaticus (top) to the LO of
Lychnocanoma parallelipes (base).
Age.—Late Miocene (6.1–5.4 Ma).
Secondary events.—RD of Lithelius barbatus.
Remarks.—The FO of Larcopyle pylomaticus and the LO of Lychnocanoma
parallelipes are located at the same level near 79.0 m at Site U1430 (Table 1, Figure 3).
Hence, the Axoprunum acquilonium Zone is absent in the studied core.
Lithelius barbatus Zone (Motoyama, 1996)
Category.—Interval zone.
Definition.—Interval from the LO of Lychnocanoma parallelipes (top) to the RI of
Lithelius barbatus (base).
Age.—Late Miocene (6.9–6.1 Ma).
Secondary events.—FO of Axoprunum acquilonium.
Remarks.—The FO of Axoprunum acquilonium coincides with the base of this zone
and can be used as an additional marker for recognizing the base of this zone (Kamikuri
et al., 2004). The Lithelius barbatus Zone is not designated at Site U1430 because no
peak event of L. barbatus is found in this studied core (Table 1, Figure 3).
Lychnocanoma parallelipes Zone (Motoyama, 1996)
Category.—Interval zone.

Accepted manuscript
12
Definition.—Interval from the RI of Lithelius barbatus (top) to the FO of
Lychnocanoma parallelipes (base).
Age.—Late Miocene (7.3–6.9 Ma).
Secondary events.—LO of Cycladophora nakasekoi.
Remarks.—The LO of Cycladophora nakasekoi is a good marker horizon for the
base of this zone (Motoyama, 1996; Kamikuri et al., 2004).
Cycladophora funakawai Zone (Motoyama, 1996; emend. Kamikuri, in press)
Category.—Interval zone.
Definition.—Interval from the FO of Lychnocanoma parallelipes (top) to the LO of
L. magnacornuta (base).
Age.—Late Miocene (9.1–7.3 Ma).
Secondary events.—FO of Cycladophora sphaeris.
Remarks.—The FO of Cycladophora sphaeris is a useful biohorizon for the middle
part of this zone. This zone approximately correlates the Spurioclathrocyclas urymensis
Assemblage of Vasilenko (2015). However S. urymensis was not encountered in this
study.
Lychnocanoma magnacornuta Zone (Funayama, 1988)
Category.—Taxon range zone.
Definition.—Taxon range of Lychnocanoma magnacornuta (base).
Age.—Middle to late Miocene (11.8–9.1 Ma).
Remarks.—This zone corresponds to the Lychnocanoma nipponica magnacorunuta
Assemblage of Vasilenko (2015) and the lower part of the Lychnocanium nipponicum
Zone of Tochilina (1985). The L. magnacorunuta Taxon range zone is divided into the
following two subzones.

Accepted manuscript
13
Collosphaera reynoldsi Subzone (Kamikuri, in press)
Category.—Interval subzone.
Definition.—Interval from the LO of Lychnocanoma magnacornuta (top) to the LO
of Cyrtocapsella japonica (base).
Age.—Late Miocene (10.0–9.1 Ma).
Secondary events.—FO of Cycladophora nakasekoi.
Remarks.—The FO of Cycladophora nakasekoi can be used as an additional marker
horizon for recognizing the base of this zone (Motoyama and Maruyama, 1998;
Kamikuri et al., 2004).
Cyrtocapsella japonica Subzone (Nakaseko and Sugano, 1972; emend. Kamikuri, in
press)
Category.—Interval subzone.
Definition.—Interval from the LO of Cyrtocapsella japonica (top) to the FO of
Lychnocanoma magnacornuta (base).
Age.—Middle to late Miocene (11.8–10.0 Ma).
Secondary events.—FO and LO of Dendrospyris uruyaensis.
Remarks.—This subzone was not encountered at Site U1425, probably due to low
stratigraphic resolution.
Eucyrtidium inflatum Zone (Reynolds, 1980; emend. Funayama, 1988)
Category.—Interval zone.
Definition.—Interval from the FO of Lychnocanoma magnacornuta (top) to the FO
of Eucyrtidium inflatum (base).
Age.—Middle Miocene (15.0–11.8 Ma).
Remarks.—This zone is divided into the following two subzones.

Accepted manuscript
14
Eucyrtidium inflatum b Subzone (Funayama, 1988)
Category.—Interval subzone.
Definition.—Interval from the FO of Lychnocanoma magnacornuta (top) to the RD
of Cyrtocapsella tetrapera (base).
Age.—Middle Miocene (12.7–11.8 Ma).
Secondary events.—LOs of Eucyrtidium inflatum, Lithopera renzae and
Calocycletta motoyamai.
Remarks.—The LO of Calocycletta motoyamai and the LO of Eucyrtidium inflatum
are good markers for the top of this zone (Kamikuri et al., 2007; Kamikuri, 2010).
Eucyrtidium inflatum a Subzone (Funayama, 1988)
Category.—Interval subzone.
Definition.—Interval from the RD of Cyrtocapsella tetrapera (base) to the FO of
Eucyrtidium inflatum (base).
Age.—Middle Miocene (15.0–12.7 Ma).
Secondary events.—FO of Calocycletta motoyamai and LO of these species:
Dendrospyris sakaii, Lipmanella hister, Stylosphaera magnaporulosa and Eucyrtidium
asanoi.
Remarks.—The RD of Cyrtocapsella tetrapera, FO of Calocycletta motoyamai, and
LO of Stylosphaera magnaporulosa occur at the same level between Site
1430A-29X-CC and 30X-CC. The LOs of Eucyrtidium asanoi, Dendrospyris sakaii and
Lipmanella hister are also found at the same horizon between Site 1430A-30X-CC and
31X-CC (Table 1, Figure 3). This suggests that the interval between 12.7 and 14.9 Ma
was recorded in the short sedimentary sequence from Site 1430A-29X-CC to 31X-CC
(264.43–270.13 m). The FO of Calocycletta motoyamai is a useful marker for the upper
part of this subzone (Kamikuri, 2010).

Accepted manuscript
15
Discussion
Motoyama (1996) presented the stratigraphic distribution of radiolarians at DSDP
Site 302 in central Japan in order to establish radiolarian zonation and reported that the
Quaternary radiolarian zones proposed by Hays (1970) and Foreman (1975) could not
be applied to DSDP Site 302.
The Quaternary radiolarian zones in the North Pacific were first developed by Hays
(1970). He defined four radiolarian zones from the Botryostrobus aquilonaris Zone
(=Eucyrtidium tumidulum in the original paper) to the Lamprocyrtis heteroporos Zone.
Later, Foreman (1975) revised Hays’s Eucyrtidium matuyamai Zone. The four zones
were distinguished by LOs of Stylatractus universus and E. matuyamai, and FO of E.
matuyamai. The zonation has been accepted as a standard zonal scheme for the middle
to high latitudes of the North Pacific (Kling, 1973; Sakai, 1980; Reynolds, 1980;
Weaver et al., 1981; Wolfart, 1981; Morley, 1985; Motoyama and Maruyama, 1998;
Kamikuri et al., 2007).
Stylatractus universus has been reported from the Southern Ocean (Hays, 1965),
North Pacific (Spencere-Cervato et al., 1993, Morley and Nigrini, 1995; Shilov, 1995;
Motoyama, 1996; Kamikuri et al., 2004), equatorial Pacific (Johnson and Knoll, 1974;
Moore, 1995; Kamikuri et al., 2009) and equatorial Atlantic and Indian oceans (Nigrini,
1967; Hays and Shackleton, 1976; Caulet et al., 1993). Based on these observations,
Hays and Shackleton (1976) suggested that S. universus effectively has a global
geographic distribution. They also showed that the LO of S. universus is globally
synchronous through comparison of the datum with oxygen isotope records over
widespread high- and low-latitude deep-sea sediments and that the isotopic transition is
between Stages 11 and 12 globally. Hays (1970) and Morley and Nigrini (1995)
suggested that the LO of S. universus is synchronous throughout the North Pacific. The

Accepted manuscript
16
cause of extinction for this species is unclear.
Hays (1970) documented the evolutionary history of Eucyrtidium matuyamai
during the early Pleistocene in the subarctic North Pacific. It evolved from Eucyrtidium
calvertense, which has a relatively small shell and a geographic distribution ranging
from the Bering Sea to 34°N. The LO of E. matuyamai within the Chron C1r.1n
(Jaramillo event) is a very good marker horizon for biostratigraphic correlation of the
middle- to high-latitude North Pacific (Hays, 1970; Kling, 1973; Foreman, 1975;
Kellogg and Hays, 1975; Reynolds, 1980; Sakai, 1980; Wolfart, 1981; Weaver et al.,
1981; Morley, 1985; Spencer-Cervato et al., 1993; Morley and Nigrini, 1995;
Motoyama, 1996; Kamikuri et al., 2004, 2007). The FO of E. matuyamai occurred near
the base of Chron C2n (Olduvai event) and is also synchronous throughout the North
Pacific.
In IODP Exp. 346, cores were taken at seven sites covering a wide latitudinal range
of the Japan Sea (Figure 1). In the cores recovered during this expedition, Stylatractus
universus was completely absent after the Pliocene (Appendix). In addition,
Eucyrtidium matuyamai occurred in only a single sample from each of Sites U1422,
U1424 and U1426. Hence, three bioevents (LOs of S. universus and E. matuyamai, and
FO of E. matuyamai) were either missing or had different ranges between the Japan Sea
and the open ocean. Although the LO of S. universus and E. matuyamai and FO of E.
matuyamai, which together define the Quaternary radiolarian zones for the subarctic
North Pacific, appear to be reliable synchronous biostratigraphic events throughout the
North Pacific, the absence or extremely rare occurrence of these species in this study
indicate that the usefulness of these markers are limited in the Japan Sea. The
occurrences were probably because S. universus and E. matuyamai are basically
intermediate to deep waters in vertical distribution, as mentioned below.
The modern Japan Sea is a semi-enclosed marginal sea in the Northwest Pacific and
is connected to other seas by four narrow and shallow straits with a maximum sill depth

Accepted manuscript
17
of 130 m. The Japan Sea opened in the early Miocene, and the position of straits has had
an influence on paleoceanographic changes, such as surface- and deep-water circulation
in the Japan Sea, since the Early Miocene (Jolivet and Tamaki, 1992; Tada, 1994). The
final restriction of the deeper water (probably intermediate water) connection between
the Japan Sea and Pacific is accompanied by uplift of the strait (paleo-Tsugaru Strait) in
northern Japan and an eustatic sea level drop during the late Pliocene (Tada, 1994;
Kamikuri and Motoyama, 2007; Itaki, 2016). Deeper-dwelling taxa, including
Cornutella profunda and Peripyramis circumtexta, may not have moved from the North
Pacific to the Japan Sea since the late Pliocene. Hence, the absence or extremely rare
occurrence of zonal markers Stylatractus universus and Eucyrtidium matuyamai likely
indicates that they inhabited deeper waters. Stylatractus universus was recognized at
ODP Sites 438 (40°37.79′N and 143°14.15′E; water depth, 1558 m) and 1150 (39°11′N
and 143°20′E; water depth, 2681 m) off the east coast of Honshu Island (Motoyama,
1996; Kamikuri et al., 2004) but did not occur at JAMSTEC Hole 902-C9001C
(41°10′N and 142°12′E; water depth, 1180 m) near the Tsugaru Strait (Matsuzaki et al.,
2014). Stylatractus universus appears to have inhabited deep water below 1200 m in the
northwestern Pacific and has not been able to migrate into the Japan Sea across the
Tsugaru Strait since the Pliocene. On the other hand, in the Bering Sea, which is the
northernmost marginal sea of the North Pacific, S. universus and E. matuyamai were
identified from the Late Quaternary sediment (Ikenoue et al., 2016), and this
distribution was likely made possible by the several deep straits, such as Kamchatka
Strait (maximum water depth, 4420 m), that connect this marginal sea to the North
Pacific.
Matul et al. (2002) identified four radiolarian datum events in the late Quaternary
sediment from the Sea of Okhotsk that they suggest are approximately synchronous in
comparison with those in the North Pacific—the LO of these species: Axoprunum
acquilonium (=Stylacontarium acquilonium in the original paper), Schizodiscus

Accepted manuscript
18
japonicus (=Spongodiscus sp. in the original paper), Amphimelissa setosa and
Lychnocanoma. sakaii (=L. nipponica sakaii in the original paper). These datum levels
are also isochronous between the Japan Sea and the North Pacific, except for the LO of
Ax. acquilonium (Motoyama, 1996; Itaki et al. 2007; Kamikuri and Motoyama, 2007).
We defined four new radiolarian zones for the Japan Sea based on four radiolarian
datum; the LOs of Am. setosa, S. japonicus and Ax. acquilonium, and the ET from
Cycladophora sphaeris to C. davisiana (Figures 2, 3). Of the new zonations for the
Japan Sea, the L. sakaii Zone and A. setosa Zone can be applied to the sediment of the
North Pacific, but the S. japonicus Zone and Hexacontium minerva Zone cannot be
applied to North Pacific sediment because the LO of Ax. acquilonium is diachronous
from the Japan Sea and the North Pacific. The LO of Ax. acquilonium is at 1.7 Ma in the
Japan Sea but is at 0.3 Ma in the North Pacific (Morley and Nigrini, 1995; Kamikuri et
al., 2004, 2007).
Matsuzaki et al. (2014) proposed eight new radiolarian zones based on 17
radiolarian datum levels in the Late Quaternary for the mixed water region of the
Northwest Pacific. Of these, four events (FO of Spongaster tetras irregularis,
Cyrtidosphaera reticulata, Spongoliva ellipsoids and Ceratospyris problematica),
which defined their respective zones, were not observed in the Japan Sea because these
species are basically subtropical or transitional in their geographic distribution.
Conclusions
Core samples were collected in the Japan Sea in IODP Exp. 346 in order to study
the atmosphere-ocean linkage related to the East Asian monsoonal system. More than
6000 m of Quaternary to middle Miocene sediments were recovered from seven sites.
Radiolarians, which are comprised of opal, are well preserved in the sea floor sediments

Accepted manuscript
19
of the Japan Sea and were used to establish biostratigraphy for more precise dating and
correlation of sedimentary sequences drilled at IODP Sites U1422–U1427 and U1430.
The ages of 41 radiolarian datum events recognized in this study were updated based on
the geologic time scale (GTS) of Ogg (2012) by extrapolation of each horizon within
each magnetic chron. In this study, we subdivided and dated the core materials mainly
using the radiolarian zonation of Funayama (1988), Motoyama (1996) and Itaki
(2001b); four newly established Quaternary radiolarian zones can be applied to the
Japan Sea. The sedimentary sequences were divided into six zones from the
Ceratospyris borealis Zone to the Hexacontium parviakitaense Zone at Site U1422,
eight zones from the C. borealis Zone to the Larcopyle pylomaticus Zone at Sites
U1423, U1424 and U1426, fourteen zones from the C. borealis Zone to the Eucyrtidium
inflatum Zone at Sites U1425 and U1430, and four zones from the C. borealis to the
Schizodiscus japonicus Zone at Site U1427. The absence or extremely rare occurrence
Stylatractus universus and E. matuyamai indicate that these species inhabited deeper
waters and have not been able to migrate into the Japan Sea across the Tsugaru Strait
since the Pliocene.
Faunal references
Figure 5
Amphimelissa setosa (Cleve, 1889). Kruglikova, 1989, p. 473, pl. 3, figs. 1–25.
Axoprunum acquilonium (Hays, 1970). Matsuzaki et al., 2015, p. 17, figs. 5.1–5.3.
Calocyclas motoyamai Kamikuri, 2010. Kamikuri, 2010, p. 100, pl. 7, fig. 1a–b.
Cycladophora davisiana Ehrenberg, 1862. Motoyama, 1997, p. 60, pl. 1, figs. 4–10.
Cycladophora nakasekoi Motoyama, 1996. Motoyama, 1996, p. 243, pl. 4, figs. 1a–3b.
Cycladophora sphaeris (Popova, 1989): Spuroclathrocyclas sphaeris Popova, 1989.

Accepted manuscript
20
Popova, 1989, p. 73, pl. 11, fig. 17, pl. 12, fig. 3: Cycladophora sakaii Motoyama,
1996. Motoyama, 1996, p. 246, pl. 4, figs. 4a–6b: Cycladophora sphaeris (Popova,
1989). Kamikuri, in press, fig. 8.5.
Cyrtocapsella tetrapera (Haeckel, 1887). Wolfart, 1981, p. 497, pl. 3, fig. 4.
Cyrtocapsella japonica Nakaseko, 1963. Nakaseko, 1963, p. 193, pl. 4, figs. 1–3.
Dendrospyris? sakaii Sugiyama and Furutani, 1992. Sugiyama and Furutani, 1992, p.
204, pl. 13, figs. 3, 6, pl. 20, figs. 3, 4c: Dendrospyris sachalinensis Vituchin, 1993.
Vituchin, 1993, p. 86, pl. 26, figs. 6–8, pl. 29, fig. 13.
Dendrospyris uruyaensis Kamikuri, 2010. Kamikuri, 2010, p. 100, pl. 5, figs. 7–9,
26–28, pl. 7, figs. 9–12.
Dictyophimus bullatus Morley and Nigrini, 1995. Morley and Nigrini, 1995, p. 79, pl. 4,
figs. 5, 9, 10: Dictyophimus robustus Motoyama, 1996. Motoyama, 1996, p. 246, pl.
6, figs. 1a–2c.
Eucyrtidium asanoi Sakai, 1980. Sakai, 1980, p. 709, pl. 7, figs. 12a–14b.
Eucyrtidium inflatum Kling, 1973. Kling, 1973, p. 636, pl. 11, figs. 7, 8, pl. 15, figs.
7–10.
Eucyrtidium matuyamai Hays, 1970. Hays, 1970, p. 213, pl. 1, figs. 7–9.
Hexacontium akitaense (Nakaseko, 1971): Hexacontium akitaensis (Nakaseko, 1971)
[sic]. Kamikuri, 2010, p. 98, pl. 1, figs. 7a–8b, 10a–12, pl. 4, fig. 16.
Hexacontium parviakitaense Kamikuri, 2010: Hexacontium parviakitaensis Kamikuri,
2010 [sic]. Kamikuri, 2010, p. 98, pl. 1, figs. 9a, b, 13a–14b, pl. 4, fig. 15.
Larcopyle pylomaticus (Riedel, 1958). Lazarus et al., 2005, p. 115, pl. 9, figs. 1–3, 6,
10?, 12? (not 4, 5, 7–9, 11): Spongurus? pylomaticus borealis Kruglikova, 1974.
Matsuzaki et al., 2015, p. 25, figs. 6.38, 6.39.
Lipmanella hister (Petrushevskaya, 1975). Sugiyama and Furutani, 1992, p. 209, pl. 13,
figs. 7, 8.
Lipmanella redondoensis (Campbell and Clark, 1944). Funakawa, 2000, p. 108, pl. 4,

Accepted manuscript
21
figs. 2a–3c, pl. 7, figs. 6a–6c, text-fig. 8.
Lithelius barbatus Motoyama, 1996. Motoyama, 1996, p. 243, pl. 3, figs. 11–14.
Lithopera renzae Sanfilippo and Riedel, 1970. Sanfilippo and Riedel, 1970, p. 454, pl. 1,
figs. 21–23, 27.
Lychnocanoma magnacornuta Sakai, 1980. Motoyama, 1996, p. 248, pl. 5, figs. 10a–11.
Lychnocanoma parallelipes Motoyama, 1996. Motoyama, 1996, p. 248, pl. 5, figs.
12a–14.
Lychnocanoma sakaii Morley and Nigrini, 1995. Kamikuri, 2010, p. 101, pl. 6, fig. 1a,
b.
Schizodiscus japonicus Matsuzaki and Suzuki, 2014. Matsuzaki et al., 2014, p. 209, pl.
2, figs. 27–30: Spongodiscus sp. Ling, 1973. Ling, 1973, p. 778, pl. 1, figs. 9, 10.
Matul et al., 2002, p. 30, figs. 4.3, 4.4. Ikenoue et al., 2016, p. 40, pl. 2, figs. 1–20.
Siphocampe arachnea (Ehrenberg, 1862) group. Nigrini, 1977, p. 255, pl. 3, figs. 7, 8.
Stylosphaera magnaporulosa (Clark and Campbell, 1942). Sugiyama and Furutani,
1992, p. 202, pl. 12, figs. 5–7, 10, pl. 15, figs. 3, 4, 6, 7.
Acknowledgments
The authors are grateful to John Rogers and Noritoshi Suzuki for reviewing the
manuscript. We are grateful to Ryuji Tada, Rick Murray, and the other scientists and
crew members of Exp. 346 of JOIDES Resolution for their support and prompt attention
to our requests. Samples were provided by the Integrated Ocean Drilling Program
(IODP). This work was also partly supported by Japan Society for the Promotion of
Science (JSPS) KAKENHI Grant Numbers 15K17780 (S. Kamikuri), 25400504 and
23221022 (T. Itaki); and JSPS Research Fellowship for Young Scientists (K.M.
Matsuzaki).

Accepted manuscript
22
References
Alexandrovich, J. M., 1992: Radiolarians from Sites 794, 795, 796, and 797 (Japan Sea).
In, Pisciotto, K. A., Ingle, J. C., Jr., von Breymann, M. T. and Barron, J. et al. eds.,
Proceedings of the Ocean Drilling Program, Scientific Results, vol. 127/128, no. 1,
p. 291–307. Ocean Drilling Program, College Station, TX.
Bak, Y. S., Lee, J. D., Yun, H., Oh, J. and Lee, S. J., 2012: Paleoceanographic
implications and radiolarians from the Pohang Basin, southeast Korea. Acta
Geologica Sinica (English Edition), vol. 86, p. 801–840.
Campbell, A. S. and Clark, B. L., 1944: Miocene radiolarian faunas from Southern
California. Geological Society of America, Special Paper, vol. 51, p. 1–76.
Caulet, J. -P., Nigrini, C. and Schneider, D. A., 1993: High resolution
Pliocene-Pleistocene radiolarian stratigraphy of the tropical Indian Ocean. Marine
Micropaleontology, vol. 22, p. 111–129.
Clark, B. L. and Campbell, A. S., 1942: Eocene radiolarian faunas from the Mt. Diablo
area, California. Geological Society of America, Special Paper, vol. 39, p. 1–112.
Cleve, P. T., 1889: Plankton collected by the Swedish Expedition to Spitzbergen in 1898.
Küngliga Svenska Vetenskaps Akademiens Handlingar, vol. 32, p. 1–51.
Ehrenberg, C. G., 1862: Über die Tiefgrund-Verhältnisse des Oceans am Einagange der
Monatsberichte der Königlich Preussischen Akademie
der Wissenschaften zu Berlin, vol. 1861, p. 275–315.
Foreman, H. P., 1973: Radiolaria from DSDP Leg 20. In, Heezen, B. C., MacGregor, I.
D. et al. eds., Initial Reports of the Deep Sea Drilling Project, vol. 20, p. 249–305.
U. S. Government Printing Office, Washington, DC.
Foreman, H. P., 1975: Radiolaria from the North Pacific, Deep Sea Drilling Project, Leg

Accepted manuscript
23
32. In, Larson, R. L., Moberly, R. et al. eds., Initial Reports of the Deep Sea
Drilling Project, vol. 32, p. 579–676. U. S. Government Printing Office,
Washington, DC.
Funakawa, S., 2000: Internal skeletal structures of the Cenozoic genera Gondwanaria,
Lipmanella and Lithomelissa (Plagiacanthidae, Nassellaria) and their taxonomy.
Micropaleontology, vol. 46, p. 97–121.
Funayama, M., 1988: Miocene radiolarian stratigraphy of the Suzu area, northwestern
part of the Noto Peninsula, Japan. Contributions of the Institute of Geology and
Paleontology, Tohoku University, vol. 91, p. 15–41. (in Japanese with English
abstract)
Haeckel, E., 1887: Report on the Radiolaria collected by H.M.S. Challenger during the
years 1873–1876. Report on Scientific Results of the Voyage of the H.M.S.
Challenger, Zoology, vol. 18, p. 1–1803.
Hays, J. D., 1965: Radiolaria and late Tertiary and Quaternary history of Antarctic seas.
Antarctic Research, vol. 5, p. 125–184.
Hays, J. D., 1970: Stratigraphy and evolutionary trends of Radiolaria in North Pacific
deep-sea sediments. Geological Society of America, Memoir, vol. 126, p. 185–218.
Hays, J. D. and Shackleton, N. J., 1976: Globally synchronous extinction of the
radiolarian Stylatractus universus. Geology, vol. 4, p. 649–652. Ikenoue, T., Okazaki, Y., Takahashi, K. and Sakamoto, T., 2016: Bering Sea radiolarian
biostratigraphy and paleoceanography at IODP Site U1341 during the last four
million years. Deep-Sea Research II, vol. 125/126, p. 38–55.
ISSC (International Subcommission on Stratigraphic Classification), 1994:
International Stratigraphic Guide–A Guide to Stratigraphic Classification,
Terminology, and Procedure (2nd edition), 214 p. The International Union of
Geological Sciences and The Geological Society of America, Inc.
Itaki, T., 2001a: Depth-related radiolarian assemblage in the water-column and surface
sediments of the Japan Sea. Marine Micropaleontology, vol. 47, p. 253–270.

Accepted manuscript
24
Itaki, T., 2001b: Radiolarian faunal changes in the eastern Japan Sea during the last 30
kyr. News of Osaka Micropaleontologists (NOM), Special Volume, no. 12, p.
359–374. (in Japanese with English abstract)
Itaki, T., 2003: Elutriation technique for the extraction of radiolarian skeletons from
sandy sediment. Fossils, vol. 73, p. 38–41. (in Japanese)
Itaki, T., 2016: Transitional changes in microfossill assemblages in the Japan Sea from
the Late Pliocene to Early Pleistocene related to global climatic and local tectonic
events. Progress in Earth and Planetary Science, vol. 3, no. 11, p. 1–21.
Itaki, T., Komatsu, N. and Motoyama, I., 2007: Orbital- and millennial-scale changes of
radiolarian assemblages during the last 220 kyrs in the Japan Sea. Palaeogeography,
Palaeoclimatology, Palaeoecology, vol. 247, p. 115–130.
Johnson, D. A. and Knoll, A. H., 1974: Absolute ages of Quaternary radiolarian datum
levels in the equatorial Pacific. Quaternary Research, vol. 5, p. 99–110.
Jolivet, L. and Tamaki, K., 1992: Neogene kinematics in the Japan Sea region and
volcanic activety of the Northeast Japan Arc. In, Tamaki, K., Suyehiro, K., Allan, J.,
McWilliams, M. et al. eds., Proceedings of the Ocean Drilling Program, Scientific
Results, vol. 127/128, no. 2, p. 1311–1331. Ocean Drilling Program, College
Station, TX.
Kamikuri, S., 2010: New late Neogene radiolarian species from the middle to high
latitudes of the North Pacific. Revue de Micropaleontologie, vol. 53, p. 85–106.
Kamikuri, S., in press: Late Neogene radiolarian biostratigraphy of the eastern North
Pacific ODP Sites 1020/1021. Paleontological Research.
Kamikuri, S. and Motoyama, I., 2007: Radiolarian assemblage and environmental
changes in the Japan Sea since the Late Miocene. Fossils, vol. 82, p. 35–42. (in
Japanese with English abstract)
Kamikuri, S., Motoyama, I., Nishi, H. and Iwai, M., 2009: Neogene radiolarian
biostratigraphy ad faunal evolution rates in the eastern equatorial Pacific ODP Sites

Accepted manuscript
25
845 and 1241. Acta Palaeontologica Polonica, vol. 54, p. 713–742.
Kamikuri, S., Nishi, H. and Motoyama, I., 2007: Effects of late Neogene climatic
cooling on North Pacific radiolarian assemblages and oceanographic conditions.
Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 249, p. 370–392.
Kamikuri, S., Nishi, H., Motoyama, I. and Saito, S., 2004: Middle Miocene to
Pleistocene radiolarian biostratigraphy in the Northwest Pacific Ocean, ODP Leg
186. Island Arc, vol. 13, p. 191–226.
Kellogg, D. E. and Hays, J. D., 1975: Microevolutionary patterns in late Cenozoic
Radiolaria. Paleobiology, vol. 1, p. 150–160.
Kling, S. A., 1973: Radiolaria from the eastern North Pacific, Deep Sea Drilling Project
Leg 18. In, Kulm, L. D., von Huene, R., et al. eds., Initial Reports of Deep Sea
Drilling Project, vol. 18, p. 617–671. U. S. Government Printing Office,
Washington, DC.
Kruglikova, S. B., 1974: The characteristic species of Radiolaria in bottom sediments of
the Pacific boreal zone. In, Zhuze, A. P. ed., Ocean and Marine Micropaleontology,
p. 187–196. Akademiya Nauk SSSR, Okeanograficheskaya Komissiya, Nauka,
Moskva. (in Russian with English abstract)
Kruglikova, S. B., 1989: Arctic Ocean radiolarians. In, Hermann, Y. ed., The Arctic
Seas: Climatology, Oceanography, Geology and Biology, p. 461–479. Van
Nostrand Reinhold Company, New York.
Lazarus, D., Faust, K. and Popova-Goll, I., 2005: New species of prunioid radiolarians
from the Antarctic Neogene. Journal of Micropalaeontology, vol. 24, p. 97–121.
Ling, H. Y., 1973: Radiolaria: Leg 19 of the Deep Sea Drilling Project. In, Creager, J.
S., Scholl D. W. et al. eds., Initial Reports of the Deep Sea Drilling Project, vol. 19,
p. 777–797. U. S. Government Printing Office, Washington, DC.
Ling, H. Y., 1975: Radiolaria: Leg 31 of the Deep Sea Drilling Project. In, Karig, D. E.,
Ingle, J. C., Jr., Bouma, A. H. et al. eds., Initial Reports of the Deep Sea Drilling

Accepted manuscript
26
Project, vol. 31, p. 703–761. U. S. Government Printing Office, Washington, DC.
Ling, H. Y., 1992: Radiolarians from the Sea of Japan: Leg 128. In, Pisciotto, K. A.,
Ingle, J. C., Jr., von Breymann, M. T., Barron, J. et al. eds., Proceedings of the
Ocean Drilling Program, Scientific Results, vol. 127/128, no. 1, p. 225–236. Ocean
Drilling Program, College Station, TX.
Matsuzaki, K. M. and Suzuki, N., in press. Quaternary radiolarian biostratigraphy in the
subarctic Northeastern Pacific (IODP Expedition 341 Site U1417) and synchroneity
of bioevents across the North Pacific. Journal of Micropalaeontology.
Matsuzaki, K. M., Suzuki, N. and Nishi, H., 2015: Middle to Upper Pleistocene
polycystine radiolarians from Hole 902-C9001C, northwestern Pacific.
Paleontological Research, vol. 19, p. 1–77.
Matsuzaki, K., Suzuki, N., Nishi, H., Takashima, R., Kawate, Y. and Sakaii, T., 2014:
Middle to Late Pleistocene radiolarian biostratigraphy in the water-mixed region of
the Kuroshio and Oyashio currents, northeastern margin of Japan (JAMSTEC Hole
902-C9001C). Journal of Micropalaeontology, vol. 33, p. 205–222.
Matul, A., Abelmann, A., Nürnberg, D. and Tiedemann, R., 2009: Stratigraphy and
major paleoenvironmental changes in the Sea of Okhotsk during the last million
years inferred from radiolarian data. Oceanography, vol. 49, p. 93–100.
Matul, A., Abelmann, A., Tiedemann, R., Kaiser, A. and Nürnberg, D., 2002: Late
Quaternary policystine radiolarian datum events in the Sea of Okhotsk. Geo-Marine
Letters, vol. 22, p. 25–32.
Moore, T. C., Jr., 1995: Radiolarian stratigraphy, Leg 138. In, Pisias, N. G. et al. eds.,
Proceedings of the Ocean Drilling Program, Scientific Results, vol. 138, p. 191–232.
Ocean Drilling Program, College Station, TX.
Morley, J. J., 1985: Radiolarians from the Northwest Pacific, Deep Sea Drilling Project
Leg 86. In, Heath, G. R., Burckle L. H. et al. eds., Initial Reports of the Deep Sea

Accepted manuscript
27
Drilling Project, vol. 86, p. 399–422. U. S. Government Printing Office,
Washington, DC.
Morley, J. J. and Nigrini, C., 1995: Miocene to Pleistocene radiolarian biostratigraphy
of North Pacific sites 881, 884, 885, 886 and 887. In, Rea, D. K., Basov, I. A.,
Scholl, D. W. and Allan, J. F. eds., Proceedings of the Ocean Drilling Program,
Scientific Results, vol. 145, p. 55–91. Ocean Drilling Program, College Station, TX.
Motoyama, I., 1996: Late Neogene radiolarian biostratigraphy in the subarctic
Northwest Pacific. Micropaleontology, vol. 42, p. 221–262.
Motoyama, I., 1997: Origin and evolution of Cycladophora davisiana Ehrenberg
(Radiolaria) in DSDP Site 192, Northwest Pacific. Marine Micropaleontology, vol.
30, p. 45–63.
Motoyama, I. and Maruyama, T., 1998: Neogene diatom and radiolarian biochronology
for the middle-to-high latitudes of the Northwest Pacific region: Calibration to the
Cande and Kent’s geomagnetic polarity time scales (CK 92 and CK 95). Journal of
the Geological Society of Japan, vol. 104, p. 171–183. (in Japanese with English
abstract)
Nakaseko, K., 1954: Preliminary report of Miocene radiolarian faunas from southern
Toyama Prefecture (Micro-biostratigraphic study of the Cenozoic strata in Japan,
part 5). Science Reports, South and North College, Osaka University, vol. 3, p.
107–124.
Nakaseko, K., 1960: Applied micropaleontologyical research by means of radiolarian
fossil in the oil bearing Tertiary, Japan (Mainly in Akita and Yamagata sedimentary
basins) Part 2. Radiolarian assemblage in Yamagata basin, discussion and
conclusion. Science Reports, South and North College, Osaka University, vol. 9, p.
149–185.

Accepted manuscript
28
Nakaseko, K., 1963: Neogene Cyrtoidea (Radiolaria) from the Isozaki Formation in
Ibaraki Prefecture, Japan. Science Reports, College of General Education, Osaka
University, vol. 12, p. 165–198.
Nakaseko, K., 1971: On some species of the Genus Thecosphaera from the Neogene
formations, Japan. Science Reports, College of General Education, Osaka
University, vol. 20, p. 59–66.
Nakaseko, K., Iwamoto, H. and Takahashi, K., 1965: Radiolarian stratigraphy in the oil
and gas bearing Tertiary and upper Cretaceous formations, Japan. Contribution
from the Government of Japan to Economic Commission for Asia and the Far East,
p. 3–14. Third Petroleum Symposium, Tokyo, Japan.
Nakaseko, K. and Sugano, K., 1972: Radiolarian biostratigraphy in the oil bearing
regions of Honshu, Japan. Journal of the Japanese Association for Petroleum
Technology, vol. 37, p. 375–384. (in Japanese)
Nakaseko, K. and Sugano, K., 1973: Neogene radiolarian zonation in Japan. Memoirs of
the Geological Society of Japan, no. 8, p. 23–34. (in Japanese with English
abstract)
Nakaseko, K., Sugano, K. and Ikeda, K., 1972: Some problems concerning the
radiolarian-stratigraphy in the Niigata Sedimentary Basin, Japan (Studies of fossil
radiolarian-stratigraphy of the Neogene formation in Niigata Prefecture, Japan, part
4). Journal of the Japanese Association of Petroleum Technologists, vol. 37, p.
7–22. (in Japanese with English abstract)
Nigrini, C., 1967: Radiolaria in pelagic sediments from the Indian and Atlantic Oceans.
Bulletin of the Scripps Institution of Oceanography, University of California, San
Diego, La Jolla, California, vol. 11, p. 1–125.
Nigrini, C., 1977: Tropical Cenozoic Artostrobiidae (Radiolaria). Micropaleontology,
vol. 23, p. 241–269.
Ogg, J. G., 2012: Geomagnetic polarity time scale. In, Gradstein, F. M., Ogg, J. G.,

Accepted manuscript
29
Schmitz, M. and Ogg, G. eds., The Geologic Time Scale 2012, p. 85–113. Elsevier,
BV.
Petrushevskaya, M. G., 1971: Spumellarian and nassellarian radiolaria in the plankton
and bottom sediments of the central Pacific. In, Funnell, B. M. and Riedel, W. R.
eds., The Micropaleontology of Oceans, p. 309–317. Cambridge University Press,
London.
Petrushevskaya, M. G., 1975: Cenozoic radiolarians of the Antarctic, Leg 29, DSDP. In,
Kennett, J. P., Houtz, R. E. et al. eds., Initial Reports of the Deep Sea Drilling
Project, vol. 29, p. 541–675. U. S. Government Printing Office, Washington, DC.
Popova, I., 1989: New insight on the systematics of some Theopiliinae (Radiolaria). In,
Anonymous ed., Paleontologo-Stratigraphicheskie Issledavaniya Fanerozoya,
Dal'nego Vostoka, p. 68–77, p. 131, and p. 143–145. Dal’nevostochniy Otdel
Akademii Nauka SSSR, Vladivostok. (in Russian; original title translated)
Reshetnyak, V. V., 1955: Vertical distribution of radiolarians from the
Kurilo-Kamchatka trench. Trudy Zoologicheskogo Instituta Academii Nauk SSSR,
vol. 21, p. 94–101.
Reynolds, R. A., 1980: Radiolarians from the western North Pacific, Leg 57, Deep Sea
Drilling Project. In, Scientific Party ed., Initial Reports of the Deep Sea Drilling
Project, vol. 56/57, p. 735–769. U. S. Government Printing Office, Washington,
DC.
Riedel, W. R., 1958: Radiolaria in Antarctic sediments. Reports of B.A.N.Z. Antarctic
Research Expedition, Series B, vol. 6, p. 247–255.
Sakai, T., 1980: Radiolarians from Sites 434, 435, and 436, Northwest Pacific, Leg 56,
Deep Sea Drilling Project. In, Scientific Party ed., Initial Reports of the Deep Sea
Drilling Project, vol. 56/57, p. 695–733. U. S. Government Printing Office,
Washington, DC.
Sanfilippo, A. and Nigrini, C., 1998: Code numbers for Cenozoic low latitude

Accepted manuscript
30
radiolarian biostratigraphic zones and GPTS conversion tables. Marine
Micropaleontology, vol. 33, p. 109–156.
Sanfilippo, A. and Riedel, W. R., 1970: Post-Eocene “closed” theoperid radiolarians.
Micropaleontology, vol. 16, p. 446–462.
Shilov, V. V., 1995: Miocene - Pleistocene radiolarians from Leg 145, North Pacific. In,
Rea, D. K. Basov, I. A., Scholl, D. W. and Allan, J. F. eds., Proceedings of the
Ocean Drilling Program, Scientific Results, vol. 145, p. 93–116. Ocean Drilling
Program, College Station, TX.
Spencer-Cervato, C., Lazarus, D. B., Beckmann, J. -P., von Salis Perch-Nielsen, K. and
Biolzi, M., 1993: New calibration of Neogene radiolarian events in the North Pacific.
Marine Micropaleontology, vol. 21, p. 261–293.
Sugano, K. and Nakaseko, K., 1968: On the assemblage of fossil radiolarians in the
neighbourhood of the Nishiyama Oil Field, Niigata Prefecture, Japan–Studies of
fossil radiolarian stratigraphy of the Niigata Prefecture, Japan (Part 1). Memoirs of
Osaka Kyouiku University, vol. 17, p. 1–14. (in Japanese with English abstract)
Sugano, K. and Nakaseko, K., 1971: On the assemblage of fossil Radiolaria in the
neighbord of Kitakanbara Plain, Niigata Prefecture, Japan–Studies of fossil
radiolarian stratigraphy of the Niigata Prefecture, Japan (Part 3). Memoirs of Osaka
Kyouiku University, vol. 20, 63–79. (in Japanese with English abstract)
Sugiyama, K. and Furutani, H., 1992: Middle Miocene radiolarians from the Oidawara
Formation, Mizunami Group, Gifu Prefecture, central Japan. Bulletin of the
Mizunami Fossil Museum, vol. 19, p. 199–213.
Tada, R., 1994: Paleoceanographic evolution of the Japan Sea. Palaeogeography,
Palaeoclimatology, Palaeoecology, vol. 108, p. 487–508.
Tada, R., Murray, R. W., Alvarez Zarikian, C. A., Anderson, W. T. Jr., Bassetti, M. -A.,
Brace, B. J., Clemens, S. C., da Costa Gurgel, M. H., Dickens, G. R., Dunlea, A. G.,
Gallagher, S. J., Giosan, L., Henderson, A. C. G., Holbourn, A. E., Ikehara, K.,

Accepted manuscript
31
Irino, T., Itaki, T., Karasuda, A., Kinsley, C. W., Kubota, Y., Lee, G. S., Lee, K. E.,
Lofi, J., Lopes, C. I. C. D., Peterson, L. C., Saavedra-Pellitero, M., Sagawa, T.,
Singh, R. K., Sugisaki, S., Toucanne, S., Wan, S., Xuan, C., Zheng, H. and Ziegler,
M., 2015: Expedition 346 summary. In, Tada, R., Murray, R.W., Alvarez Zarikian,
C.A., the Expedition 346 Scientists eds., Proceedings of Integrated Ocean Drilling
Program, vol. 346. College Station, TX (Integrated Ocean Drilling Program). doi:
10.2204/iodp.proc.346.101.2015
Tochilina, S. V., 1985: Biostratigraphy of the Cenozoic North-western Pacific Ocean.
Akademiya Nauk SSSR, Tikhookeanskiy Institut Okeanologii DVNTs, no. 114, p.
1–133. (in Russian with English abstract)
Vasilenko, L. N., 2015: Upper Cenozoic radiolarians from the sedimentary cover of
Peter the Great Seamount (Sea of Japan). Bulletin of Kamchatka Regional
Association “Educational-Scientific Center” Earth Sciences, vol. 4, p. 80–93. (in
Russian with English abstract)
Vitukhin, D. I., 1993: Subdivision of the Russian Far East Cenozoic sediments based on
radiolarians. Geologicheskii Institut, Rossiskaya Akademiya Nauk, Trudi, vol. 485, p.
1–104. (in Russian with English abstract)
Weaver, F. M., Casey, R. E. and Perez, A. M., 1981: Stratigraphic and
paleoceanographic significance of early Pliocene to middle Miocene radiolarian
assemblages from Northern to Baja California. In, Garrison, G. and Douglas, R. E.
eds., The Monterey Formation and Related Siliceous Rocks of California, p. 71–86.
Society of Economic Paleontologists and Mineralogists, Los Angels, California.
Wolfart, R., 1981: Neogene radiolarians from the eastern North Pacific (off Alta and
Baja California), Deep Sea Drilling Project Leg 63. In, Yeats, R. S., Haq, B. U. et al.
eds., Initial Reports of the Deep Sea Drilling Project, vol. 63, p. 473–506. U. S.
Government Printing Office, Washington, DC.

Accepted manuscript
32
Captions.
Figure 1. Map showing locations of Deep Sea Drilling Project (DSDP), Ocean Drilling
Program (ODP), and Integrated Ocean Drilling Program (IODP) sites and oceanic
currents. (From Tada et al., 2015)
Figure 2. Correlation of radiolarian zones for the middle to high latitude of North
Pacific and the Japan Sea. Geomagnetic polarity time scale is after Ogg (2012).
Figure 3. Intercorrelation of IODP Sites U1422–1427, and U1430 based on radiolarian
zones.
Figure 4. Age-depth plot of IODP Sites U1422–1427 and U1430. Age is adopted from
GTS2012 (Ogg, 2012).
Figure 5. Photographs of selected radiolarian species identified in this study. 1,
Cycladophora nakasekoi Motoyama, Sample 346-1430A-17H-CC, sl. 1, L44/1; 2,
Cycladophora sphaeris (Popova), Sample 346-1430A-9H-4, 8-10 cm, sl. 1, R42/2; 3,
Dendrospyris uruyaensis Kamikuri, Sample 346-1430A-25H-CC, sl. 1, N56/0; 4,
Hexacontium parviakitaense Kamikuri, Sample 346-1430A-8H-CC, sl. 1, R42/3; 5,
Hexacontium akitaense (Nakaseko), Sample 346-1430A-8H-CC, sl. 1, R42/3; 6,
Eucyrtidium inflatum Kling, Sample 346-1430A-29H-CC, sl. 1, S46/3; 7, Lithopera
renzae Sanfilippo and Riedel, Sample 346-1430A-28H-CC, sl. 1, Z53/3; 8,
Lipmanella redondoensis (Campbell and Clark), Sample 346-1430A-27H-CC, sl. 1,
U37/2; 9, Larcopyle pylomaticus (Riedel), Sample 346-1430A-9H-4, 8-10 cm, sl. 1,
G35/4; 10, Cyrtocapsella japonica Nakaseko, Sample 346-1430A-20H-CC, sl. 1,
X58/0; 11, Siphocampe arachnea (Ehrenberg) group, Sample 346-1430A-8H-CC, sl.

Accepted manuscript
33
1, S30/1; 12, Amphimelissa setosa (Cleve), Sample 346-1430A-3H-CC, sl. 1, M57/0;
13, Schizodiscus japonicus Matsuzaki and Suzuki, Sample 346-1425B-3H-4, 51-53
cm, sl. 1; 14, Axoprunum acquilonium (Hays), Sample 346-1430A-6H-CC, sl. 1,
U37/3; 15, Lychnocanoma magnacornuta Sakai, Sample 346-1430A-21H-CC, sl. 1,
N33/4; 16, Lychnocanoma sakaii Morley and Nigrini, Sample 346-1425B-1H-5,
98-100 cm, sl. 1.
Table 1. Summary of radiolarian datum events at IODP Sites U1422–1427, and U1430.
The following abbreviations are used in this study to express events; FO, first
occurrence; LO, last occurrence; RI, rapid increase; RD, rapid decrease. The
concept of evolutionary transition (ET) follows that of Sanfilippo and Nigrini
(1998). The definition of radiolarian zonations was indicated by the use of bold font.
Data source: a, Itaki et al., 2007; b, Matul et al., 2002; c, Motoyama, 1996; d,
Matsuzaki and Suzuki, in press; e, Itaki per. com.; f, Ikenoue et al., 2016; g,
Kamikuri et al., 2004; h, Kamikuri et al., 2007; i, Kamikuri, 2010; j, Funayama,
1988.
Appendix. Stratigraphic distribution of selected radiolarian taxa.

Figure 1

C5
C3A
C3B
C4A
C4
M i
o c
e n
eLa
teP
l i o
c e
n e La
teLa
teEa
rlyPl
eist
ocen
e
1
2
3
4
5
6
7
8
9
0
10
11
12
Early
C3
C2
C2A
C1
n
r
n
r
n
r
n
r
n
rnr
n
r
n
r
n
r
Mid
dle
13
14
15
n
n
n
n
n
n
r
r
r
r
r
rC5B
C5AD
C5ACC5ABC5AA
C5A
Schizodiscus japonicus
Amphimelissa setosa
LO L. sakaiiLO A. setosaLO S. japonicus
LO A. acquilonium
LO H. parviakitaense
LO D. bullatus
L. hannaiL. klingi
LO D. bullatus
FO L. pylomaticus
LO L. redondoensis
LO L. parallelips
LO L. magnacornuta
FO L. magnacornuta
LO Cyrtocapsella japonica
RD Cyrtocapsella tetrapera
RI L. barbatus
FO L. parallelips
Lychnocanoma sakaiiCeratospyris borealis
E. asanoi
Botryostrobus aquilonaris
Stylatractus universus
Eucyrtidium matuyamai
Dictyophimus bullatus
Litheliusbarbatus
Lychnocanoma parallelips
Cycladophora funakawai
Cycladophora funakawai
Lychnocanoma magnacornuta
Lychnocanoma nipponica magnacornuta
Eucyrtidium inflatum
Melittosphaeramagnaporulosa
Cyrtocapsella tetrapera
Lychnocanium nipponicum
Thecosphaera japonicaCycladophora
sphaeris
Cycladophora davisiana
Clathrocyclas bicornis - Spurioclath. sphaeris
Hexacontiumparviakitaense
Hexacontoium minerva
Dictyophimus bullatus
Larcopylepylomaticus
Larcopylepylomaticus
Lychnocanoma magnacornuta
Eucyrtidium inflatum
Axoprunumacquilonium
Litheliusbarbatus
Lychnocanoma parallelips
Axoprunumacquilonium
Chr
on
Epoc
h
Tim
e (M
a)
Pola
rity
GTS2012 Radiolarian zonation
Funayama (1988)
Vasilenko (2015) Kamikuri et al. (2004)
Motoyama (1996) Nakaseko &Sugano (1973)
Kamikuri (in press)
North Pacific Japan Sea
This study
Mid
dle
C. reynoldsi
C. japonica
b
a
Spurioclath. urymensis
Figure 2

Dep
th (m
ccs
f-a)
0
100
50
150
250
350
200
300
400
450
500
1422Site
1, 23
4
5
142312
3
4
5
6
6
6
7
78
9
101112
13,15
78-11
1427
1
1
2
2
312
13
14
1516
1425 1426
1, 21, 2
33 3
4
5
142413
4
4
4
5 5
5
1430
1 LO L. sakaii2 LO A. setosa3 LO S. japonincus4 LO A. acquilonium
6 LO D. bullatus13 LO L. magnacornuta
15 FO L. magnacornuta14 LO C. japonica
16 RD C. tetrapera7 FO D. bullatus
9 FO L. pylomaticus
8 LO L. redondoensis
12 FO L. parallelipes
10 LO L. parallelipes11 RI L. barbatus
5 ET C. davisiana
0.054
0.085
0.29
1.7
2.57
3.8/4.3
4.3/4.5
5.2/5.6
5.9/6.4
6.8/7.0
7.2/7.4
9.1
9.9/10.1
11.7/11.9
12.8
Schizodiscus japonicus
Amphimelissa setosa
Lychnocanoma sakaii
Ceratospyris borealis
Cycladophora funakawai
Hexacontiumparviakitaense
Hexacontium minerva
Dictyophimus bullatus
Larcopylepylomaticus
Lychnocanoma magnacornuta
Eucyrtidium inflatum
Litheliusbarbatus
Lychnocanoma parallelips
Axoprunumacquilonium
Radiolarian zonation
Time (Ma)
5.0/5.1
Figure 3

Epoch
Time (Ma)
Pleistocene Pliocene
Site U1427
MioceneLate Late MiddleEarlyEarlyMiddleLate
1 2 3 4 5 60 7 8 9 10 11 12 13 14 15 16
Dep
th (m
ccs
f-a)
0
100
50
150
250
350
200
300
400
450
500
Site U1430
Site U1425
Site U1426
Site U1424
Site U1423
Site U1422
Figure 4

100 µm1
3
87
6
15
5
10
9
2
4
11
14
12
16
13
Figure 5

Table 1a
U1422C U1423A U1423B U1424A
Datum Age (Ma) Source Sample CCSF-A (m) Sample CCSF-A (m) Sample CCSF-A (m) Sample CCSF-A (m) 1 LO Lychnocanoma sakaii 0.054 a 1H-CC/2H-CC 4.16/14.50 1H-3, 70-72cm/1H-4, 70-72cm 3.70/5.20 top/1H-CC 0.00/6.652 LO Amphimelissa setosa 0.085 a 1H-CC/2H-CC 4.16/14.50 1H-5, 72-74cm/1H-CC 6.72/7.103 LO Schizodiscus japonicus 0.29 b 2H-CC/3H-CC 14.50/24.81 3H-5, 70-72cm/3H-6, 70-72cm 24.36/25.86 1H-CC/2H-CC 6.65/16.234 Eucyrtidium matuyamai occurrence 1.1 c 7H5, 40-42 62.89 no data no data 5H-CC 44.6305 FO Amphimelissa setosa 1.48 d 8H-CC/9H-4, 70-72cm 77.47/82.846 LO Axoprunum acquilonium 1.7 e 10H-7, 40-42cm/10H-CC 95.31/95.49 9H-CC/10H-4, 70-72cm 87.20/92.69 4H-CC/5H-CC 35.23/44.637 LO Cycladophora sphaeris 2.18-2.21 f 13H-CC/14H-CC 124.94/135.58 12H-CC/13H-CC 117.24/127.68 8H-CC/9H-CC 73.00/82.758 ET C. sphaeris to C. davisiana 2.56-2.57 f 16H-CC/17H-3, 10-12cm 145.54/149.06 12H-CC/13H-CC 117.24/127.24 8H-CC/9H-CC 73.00/82.759 FO Cycladophora davisiana 2.62-2.66 f 18H-CC/19H-CC 155.69/160.51 12H-CC/13H-CC 115.87/126.31 8H-CC/9H-CC 73.00/82.75
10 LO Hexacontium parviakitaense 2.87-2.90 f 19H-CC/20H-3, 83-85cm 160.51/164.14 13H-CC/14H-CC 126.31/136.01 8H-CC/9H-CC 73.00/82.7511 FO Hexacontium parviakitaense 3.8-4.3 g 18H-CC/19H-CC 175.52/185.58 12H-CC/13H-CC 111.29/120.9412 LO Dictyophimus bullatus 3.8-4.3 g, h 19H-CC/20H-CC 185.58/195.16 no data no data13 RD Siphocampe arachnea group 4.4-4.5 g 19H-CC/20H-CC 185.58/195.16 13H-CC/14H-CC 120.94/130.3014 FO Dictyophimus bullatus 4.3-4.5 g, h 21H-CC/22H-CC 205.01/214.85 22H-CC/23H-CC 199.34/209.20 no data no data15 RI Siphocampe arachnea group 4.6-4.7 g 25H-CC/26H-CC 228.17/237.72 15H-CC/16H-CC 139.91/149.3916 LO Lipmanella redondoensis 5.0-5.1 g17 FO Larcopyle pylomaticus 5.2-5.6 g18 RD Lithelius barbatus 5.2-5.6 g19 LO Lychnocanoma parallelipes 5.9-6.4 g20 FO Axoprunum acquilonium 6.8-7.0 h21 RI Lithelius barbatus 6.8-7.0 h22 LO Cycladophora nakasekoi 6.7-7.7 g23 FO Lychnocanoma parallelipes 7.2-7.4 h24 FO Cycladophora sphaeris 8.5 g25 LO Lychnocanoma magnacornuta 9.1 h26 FO Cycladophora nakasekoi 9.7-10.2 g27 LO Cyrtocapsella japonica 9.9-10.1 h28 LO Dendrospyris uruyaensis 10.8-12.0 i29 LO Hexacontium akitaense 10.8-12.0 i30 FO Hexacontium akitaense 11.3-11.5 i31 FO Lychnocanoma magnacornuta 11.7-11.9 h32 FO Dendrospyris uruyaensis 11.7-11.9 i33 LO Eucyrtidium inflatum 11.7-11.9 h34 LO Calocyclas motoyamai 11.7-11.9 i35 LO Lithopera renzae 12.1-12.3 h36 RD Cyrticapsella tetrapera 12.8 j37 FO Calocyclas motoyamai 12.9-13.1 i38 LO Stylosphaera magnaporulosa 13.0-13.3 h39 LO Eucyrtidium asanoi 13.8-14.1 h40 LO Dendrospyris sakaii 14.6-14.8 h41 LO Lipmanella hister 14.8-15.0 h
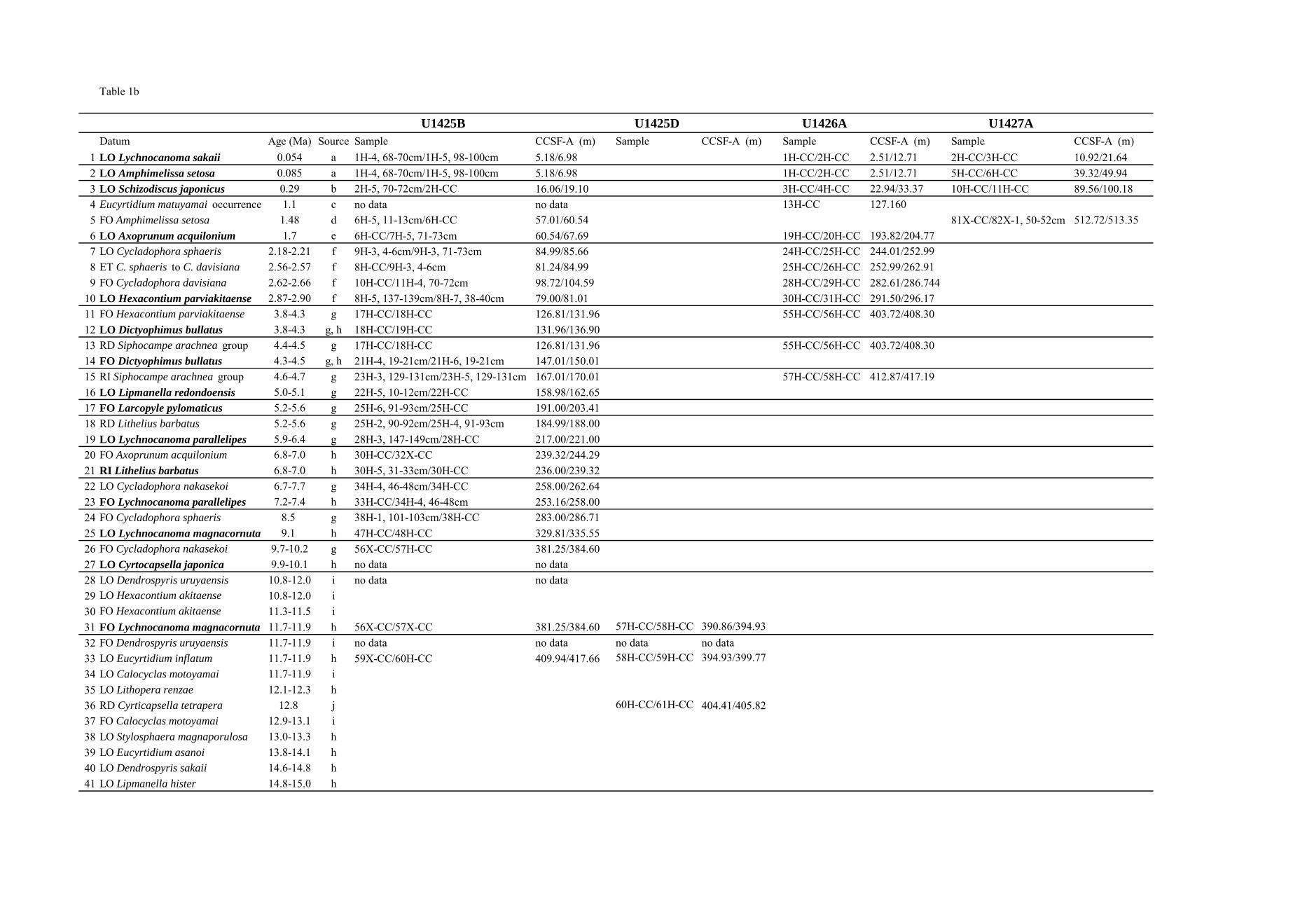
Table 1b
U1425B U1425D U1426A U1427A
Datum Age (Ma) Source Sample CCSF-A (m) Sample CCSF-A (m) Sample CCSF-A (m) Sample CCSF-A (m) 1 LO Lychnocanoma sakaii 0.054 a 1H-4, 68-70cm/1H-5, 98-100cm 5.18/6.98 1H-CC/2H-CC 2.51/12.71 2H-CC/3H-CC 10.92/21.642 LO Amphimelissa setosa 0.085 a 1H-4, 68-70cm/1H-5, 98-100cm 5.18/6.98 1H-CC/2H-CC 2.51/12.71 5H-CC/6H-CC 39.32/49.943 LO Schizodiscus japonicus 0.29 b 2H-5, 70-72cm/2H-CC 16.06/19.10 3H-CC/4H-CC 22.94/33.37 10H-CC/11H-CC 89.56/100.184 Eucyrtidium matuyamai occurrence 1.1 c no data no data 13H-CC 127.1605 FO Amphimelissa setosa 1.48 d 6H-5, 11-13cm/6H-CC 57.01/60.54 81X-CC/82X-1, 50-52cm 512.72/513.356 LO Axoprunum acquilonium 1.7 e 6H-CC/7H-5, 71-73cm 60.54/67.69 19H-CC/20H-CC 193.82/204.777 LO Cycladophora sphaeris 2.18-2.21 f 9H-3, 4-6cm/9H-3, 71-73cm 84.99/85.66 24H-CC/25H-CC 244.01/252.998 ET C. sphaeris to C. davisiana 2.56-2.57 f 8H-CC/9H-3, 4-6cm 81.24/84.99 25H-CC/26H-CC 252.99/262.919 FO Cycladophora davisiana 2.62-2.66 f 10H-CC/11H-4, 70-72cm 98.72/104.59 28H-CC/29H-CC 282.61/286.744
10 LO Hexacontium parviakitaense 2.87-2.90 f 8H-5, 137-139cm/8H-7, 38-40cm 79.00/81.01 30H-CC/31H-CC 291.50/296.1711 FO Hexacontium parviakitaense 3.8-4.3 g 17H-CC/18H-CC 126.81/131.96 55H-CC/56H-CC 403.72/408.3012 LO Dictyophimus bullatus 3.8-4.3 g, h 18H-CC/19H-CC 131.96/136.9013 RD Siphocampe arachnea group 4.4-4.5 g 17H-CC/18H-CC 126.81/131.96 55H-CC/56H-CC 403.72/408.3014 FO Dictyophimus bullatus 4.3-4.5 g, h 21H-4, 19-21cm/21H-6, 19-21cm 147.01/150.0115 RI Siphocampe arachnea group 4.6-4.7 g 23H-3, 129-131cm/23H-5, 129-131cm 167.01/170.01 57H-CC/58H-CC 412.87/417.1916 LO Lipmanella redondoensis 5.0-5.1 g 22H-5, 10-12cm/22H-CC 158.98/162.6517 FO Larcopyle pylomaticus 5.2-5.6 g 25H-6, 91-93cm/25H-CC 191.00/203.4118 RD Lithelius barbatus 5.2-5.6 g 25H-2, 90-92cm/25H-4, 91-93cm 184.99/188.0019 LO Lychnocanoma parallelipes 5.9-6.4 g 28H-3, 147-149cm/28H-CC 217.00/221.0020 FO Axoprunum acquilonium 6.8-7.0 h 30H-CC/32X-CC 239.32/244.2921 RI Lithelius barbatus 6.8-7.0 h 30H-5, 31-33cm/30H-CC 236.00/239.3222 LO Cycladophora nakasekoi 6.7-7.7 g 34H-4, 46-48cm/34H-CC 258.00/262.6423 FO Lychnocanoma parallelipes 7.2-7.4 h 33H-CC/34H-4, 46-48cm 253.16/258.0024 FO Cycladophora sphaeris 8.5 g 38H-1, 101-103cm/38H-CC 283.00/286.7125 LO Lychnocanoma magnacornuta 9.1 h 47H-CC/48H-CC 329.81/335.5526 FO Cycladophora nakasekoi 9.7-10.2 g 56X-CC/57H-CC 381.25/384.6027 LO Cyrtocapsella japonica 9.9-10.1 h no data no data28 LO Dendrospyris uruyaensis 10.8-12.0 i no data no data29 LO Hexacontium akitaense 10.8-12.0 i30 FO Hexacontium akitaense 11.3-11.5 i31 FO Lychnocanoma magnacornuta 11.7-11.9 h 56X-CC/57X-CC 381.25/384.60 57H-CC/58H-CC 390.86/394.9332 FO Dendrospyris uruyaensis 11.7-11.9 i no data no data no data no data33 LO Eucyrtidium inflatum 11.7-11.9 h 59X-CC/60H-CC 409.94/417.66 58H-CC/59H-CC 394.93/399.7734 LO Calocyclas motoyamai 11.7-11.9 i35 LO Lithopera renzae 12.1-12.3 h36 RD Cyrticapsella tetrapera 12.8 j 60H-CC/61H-CC 404.41/405.8237 FO Calocyclas motoyamai 12.9-13.1 i38 LO Stylosphaera magnaporulosa 13.0-13.3 h39 LO Eucyrtidium asanoi 13.8-14.1 h40 LO Dendrospyris sakaii 14.6-14.8 h41 LO Lipmanella hister 14.8-15.0 h

Table 1c
U1427B U1427C U1430A
Datum Age (Ma) Source Sample CCSF-A (m) Sample CCSF-A (m) Sample CCSF-A (m)1 LO Lychnocanoma sakaii 0.054 a 3H-4, 45-47cm/3H-5, 32-34cm 20.06/21.43 2H-6, 15-17cm/3H-4, 45-47cm 16.30/23.06 top/1H-CC 0.00/3.682 LO Amphimelissa setosa 0.085 a 4H-5, 48-50cm/5H-3, 48-50cm 30.63/37.16 4H-5, 45-47cm/5H-4, 45-47cm 33.79/42.90 2H-4, 68-70cm/2H-CC 9.14/13.523 LO Schizodiscus japonicus 0.29 b 12H-7, 46-48cm/14H-3, 50-52cm 102.72/109.75 12H-5, 45-47cm/14H-3, 45-47cm 109.02/124.54 2H-CC/3H-4, 68-70cm 13.52/19.284 Eucyrtidium matuyamai occurrence 1.1 c no data no data5 FO Amphimelissa setosa 1.48 d 5H-CC/6H-4, 68-70cm 43.69/49.156 LO Axoprunum acquilonium 1.7 e 6H-4, 68-70cm/6H-CC 49.15/53.567 LO Cycladophora sphaeris 2.18-2.21 f 6H-CC/7H-4, 68-70cm 53.56/59.208 ET C. sphaeris to C. davisiana 2.56-2.57 f 6H-CC/7H-4, 68-70cm 53.56/59.209 FO Cycladophora davisiana 2.62-2.66 f 7H-4, 68-70cm/7H-CC 59.20/63.61
10 LO Hexacontium parviakitaense 2.87-2.90 f 7H-4, 68-70cm/7H-CC 59.20/63.6111 FO Hexacontium parviakitaense 3.8-4.3 g 8H-CC/9H-1, 68-70cm 73.67/74.3312 LO Dictyophimus bullatus 3.8-4.3 g, h 9H-1, 68-70cm/9H-2, 68-70cm 74.33/75.8313 RD Siphocampe arachnea group 4.4-4.5 g 8H-4, 68-70cm/8H-CC 69.22/73.6714 FO Dictyophimus bullatus 4.3-4.5 g, h 9H-2, 68-70cm/9H-3, 68-70cm 75.83/77.3315 RI Siphocampe arachnea group 4.6-4.7 g 9H-2, 68-70cm/9H-3, 68-70cm 75.83/77.3316 LO Lipmanella redondoensis 5.0-5.1 g 9H-4, 68-70cm/9H-4, 120-122cm 78.83/79.3517 FO Larcopyle pylomaticus 5.2-5.6 g 9H-4, 68-70cm/9H-4, 120-122cm 78.83/79.3518 RD Lithelius barbatus 5.2-5.6 g no data no data19 LO Lychnocanoma parallelipes 5.9-6.4 g 9H-4, 68-70cm/9H-4, 120-122cm 78.83/79.3520 FO Axoprunum acquilonium 6.8-7.0 h 9H-4, 120-122cm/9H-5, 11-13cm 79.35/79.7621 RI Lithelius barbatus 6.8-7.0 h no data no data22 LO Cycladophora nakasekoi 6.7-7.7 g 9H-4, 120-122cm/9H-5, 11-13cm 79.35/79.7623 FO Lychnocanoma parallelipes 7.2-7.4 h 9H-5, 11-13cm/9H-5, 70-72cm 79.76/80.3524 FO Cycladophora sphaeris 8.5 g 10H-4, 68-70cm/10H-5, 70-72cm 89.23/90.7525 LO Lychnocanoma magnacornuta 9.1 h 16H-4, 70-72cm/16H-5, 68-70cm 149.94/151.4226 FO Cycladophora nakasekoi 9.7-10.2 g 19H-CC/20H-4, 68-70cm 185.41/191.0627 LO Cyrtocapsella japonica 9.9-10.1 h 20H-4, 68-70cm/20H-CC 191.06/194.5528 LO Dendrospyris uruyaensis 10.8-12.0 i 22H-4, 68-70cm/22H-CC 211.39/215.2629 LO Hexacontium akitaense 10.8-12.0 i 22H-4, 68-70cm/22H-CC 211.39/215.2630 FO Hexacontium akitaense 11.3-11.5 i 23H-4, 68-70cm/23H-CC 220.91/225.3431 FO Lychnocanoma magnacornuta 11.7-11.9 h 28H-1, 70-72cm/28H-3, 25-27cm 255.99/258.5632 FO Dendrospyris uruyaensis 11.7-11.9 i 28H-1, 70-72cm/28H-3, 25-27cm 255.99/258.5633 LO Eucyrtidium inflatum 11.7-11.9 h 28H-1, 70-72cm/28H-3, 25-27cm 255.99/258.5634 LO Calocyclas motoyamai 11.7-11.9 i 28H-1, 70-72cm/28H-3, 25-27cm 255.99/258.5635 LO Lithopera renzae 12.1-12.3 h 28H-3, 25-27cm/28H-CC 258.56/260.0036 RD Cyrticapsella tetrapera 12.8 j 29H-CC/30X-CC 264.43/264.6337 FO Calocyclas motoyamai 12.9-13.1 i 29H-CC/30H-CC 264.43/264.6338 LO Stylosphaera magnaporulosa 13.0-13.3 h 29X-CC/30X-CC 264.43/264.6339 LO Eucyrtidium asanoi 13.8-14.1 h 30X-CC/31X-CC 264.63/270.1340 LO Dendrospyris sakaii 14.6-14.8 h 30H-CC/31X-CC 264.63/270.1341 LO Lipmanella hister 14.8-15.0 h 30H-CC/31X-CC 264.63/270.13

Hol
e
Cor
e
Type
Sec
t
Inte
rval
Top
(cm
)
Inte
rval
Bot
tom
(cm
)
CC
SF-
A(m
)
Pre
serv
atio
n
Gro
up A
bund
ance
Am
ph
ime
lissa
se
tosa
Axo
pru
nu
m a
cq
uilo
niu
m
Cycla
do
ph
ora
da
vis
ian
a
Cycla
do
ph
ora
sp
ha
eri
s
Eu
cyrt
idiu
m m
ats
uya
ma
i
He
xa
co
ntiu
m p
arv
iakita
en
se
La
rco
pyle
pylo
ma
ticu
s
Sip
ho
ca
mp
e a
rach
ne
a gr
oup
Sch
izo
dis
cu
s ja
po
nis
cu
s
Sty
latr
actu
s u
niv
ers
us
U1422C 1 H CC 0 5 4.15 M F AU1422C 2 H CC 0 3 14.49 G A P PU1422C 3 H CC 0 5 24.80 G A R P AU1422C 4 H CC 0 5 35.09 M R RU1422C 5 H CC 0 4 45.06 G A A CU1422C 6 H CC 0 4 54.94 BU1422C 7 H 5 40 42 62.89 G R PU1422C 7 H CC 0 7 66.21 BU1422C 8 H 7 41 43 76.05 P R P AU1422C 8 H CC 0 5 76.23 P RU1422C 9 H 3 111 113 81.57 P R CU1422C 9 H CC 0 5 83.94 BU1422C 10 H 2 20 22 87.52 P R PU1422C 10 H 4 70 72 91.06 M R RU1422C 10 H 7 40 42 95.31 P R CU1422C 10 H CC 0 6 95.49 M F FU1422C 11 H CC 0 5 105.32 G C C C RU1422C 12 H CC 0 5 115.62 G A F A R RU1422C 13 H CC 0 5 124.94 G F R R RU1422C 14 H CC 0 5 135.58 G C F R R FU1422C 15 H 3 71 73 139.23 G R P CU1422C 15 H CC 0 5 140.59 G C R C CU1422C 16 H 2 50 52 142.68 G C P R C CU1422C 16 H CC 0 5 145.54 G A R F R FU1422C 17 H 3 10 12 149.06 G C RU1422C 17 H CC 0 5 150.82 M R C P RU1422C 18 H 3 73 75 154.65 G R R RU1422C 18 H CC 0 5 155.69 G C P R CU1422C 19 H 3 65 67 159.25 M C RU1422C 19 H CC 0 5 160.51 G A R R F RU1422C 20 H 3 83 85 164.14 G C P C P R AU1422C 20 H CC 0 5 165.12 G C F R FU1422C 21 H CC 0 5 169.85 G A A F R RU1422C 22 H CC 0 5 174.41 G A R A R FU1422C 23 H CC 0 5 179.15 G A C A CU1422C 24 H CC 0 5 183.82 G A C F CU1422C 25 H CC 0 6 188.65 G A A R F FU1422C 26 H CC 0 6 193.23 G C R C R R FU1422C 27 H CC 0 6 197.93 G C R C F R R PU1422C 28 H CC 0 6 202.75 G C R F F RU1422C 29 H CC 0 6 207.31 G C R F F C FU1422C 30 H CC 0 5 212.17 G C F F FU1422C 31 H CC 0 5 216.75 G C R C C C F

No. 1 2 3 4 5 6 7 8 9 10 11 12H
ole
Co
re
Typ
e
Se
ct
Inte
rva
l T
op
(cm
)
Inte
rva
l B
ott
om
(cm
)
CC
SF
-A
(m)
Pre
se
rva
tio
n
Gro
up
Ab
un
da
nce
Am
ph
ime
lissa
se
tosa
Axo
pru
nu
m a
cq
uilo
niu
m
Cycla
do
ph
ora
da
vis
ian
a
Cycla
do
ph
ora
na
ka
se
ko
i
Cycla
do
ph
ora
sp
ha
eri
s
Dic
tyo
ph
imu
s b
ulla
tus
Eu
cyrt
idiu
m m
ats
uya
ma
i
He
xa
co
ntiu
m p
arv
iakita
en
se
La
rco
pyle
pylo
ma
ticu
s
Lych
no
ca
no
ma
sa
ka
ii
Sip
ho
ca
mp
e a
rach
ne
a g
rou
p
Sch
izo
dis
cu
s ja
po
nis
cu
s
U1423A 1 H 1 70 72 0.70 G A A
U1423A 1 H 2 70 72 2.20 G C F
U1423A 1 H 3 70 72 3.70 P R F
U1423A 1 H 4 70 72 5.20 G F C
U1423A 1 H 5 72 74 6.72 G F A R
U1423A 1 H CC 0 4 7.10 G F A F
U1423A 2 H 4 130 132 13.57 G C A C P
U1423A 2 H 5 70 72 14.07 G F A C
U1423A 2 H 6 70 72 15.97 G C A F
U1423A 2 H CC 0 5 17.33 M A C F A
U1423A 3 H 5 70 72 24.36 G A A
U1423A 3 H 6 70 72 25.86 G A F A R
U1423A 3 H CC 0 5 27.31 M R R F R F
U1423A 4 H 5 70 72 34.49 G C C P A
U1423A 4 H 6 70 72 35.99 G A A F
U1423A 4 H CC 0 5 37.32 M A F P A
U1423A 5 H 4 70 72 42.92 G A A
U1423A 5 H 5 70 72 44.64 P R F
U1423A 5 H 6 70 72 46.14 G A A F
U1423A 5 H 7 10 12 46.85 M F C C F F
U1423A 5 H CC 0 5 47.32 G C R P P F F
U1423A 6 H 4 70 72 52.84 G A A P
U1423A 6 H 5 70 72 54.34 G C A R
U1423A 6 H 6 70 72 55.84 G C F C
U1423A 6 H CC 0 5 57.13 G A A F R
U1423A 7 H 4 70 72 63.00 G C C A R F
U1423A 7 H 5 70 72 64.50 G A C F C
U1423A 7 H 6 70 72 66.00 G C C F
U1423A 7 H CC 0 5 67.40 G A C F P A
U1423A 8 H 4 70 72 73.10 G C A
U1423A 8 H 5 70 72 74.60 G A F F R
U1423A 8 H 6 70 72 76.10 G C F F
U1423A 8 H CC 0 5 77.47 G A C
U1423A 9 H 4 70 72 82.84 M F C
U1423A 9 H 5 70 72 84.34 G C A
U1423A 9 H 6 70 72 85.84 G C C P
U1423A 9 H CC 0 4 87.20 P R P P
U1423A 10 H 4 70 72 92.68 G A P C P P
U1423A 10 H 5 70 72 94.18 G C R A F R
U1423A 10 H 6 70 72 95.68 G C A F
U1423A 10 H 7 58 60 97.06 G C C R
U1423A 10 H CC 0 4 97.09 G A C A F F
U1423A 11 H 1 70 72 97.99 P R C P P
U1423A 11 H 3 70 72 100.99 G C A F
U1423A 11 H 5 70 72 103.99 M C A R

No. 1 2 3 4 5 6 7 8 9 10 11 12U1423A 11 H CC 0 5 106.84 G A F C RU1423A 12 H 1 70 72 107.01 M A CU1423A 12 H 3 70 72 110.01 M A C RU1423A 12 H 5 70 72 113.01 P R RU1423A 12 H CC 0 5 115.87 G A F F C RU1423A 13 H 1 70 72 117.40 G C C F PU1423A 13 H 3 70 72 120.40 P C F PU1423A 13 H 5 70 72 123.40 G C F CU1423A 13 H CC 0 4 126.31 G A C F FU1423A 14 H 1 70 72 126.98 G C A CU1423A 14 H 3 70 72 129.98 G C AU1423A 14 H 5 70 72 132.98 G C FU1423A 14 H CC 0 5 136.01 G A C R F FU1423A 15 H 1 70 72 136.94 G C C RU1423A 15 H 3 70 72 139.94 G C C RU1423A 15 H 5 70 72 142.94 G C C RU1423A 15 H CC 0 5 145.86 G C P F F P FU1423A 16 H 1 70 72 147.32 G R CU1423A 16 H 3 70 72 150.32 G C C RU1423A 16 H 5 70 72 153.32 G C C FU1423A 16 H CC 0 4 156.38 G C R F CU1423A 17 H 1 70 72 156.84 G C A P FU1423A 17 H 3 70 72 159.84 M R C FU1423A 17 H 5 70 72 162.84 M C C R CU1423A 17 H CC 0 5 165.29 G C R F C F FU1423A 18 H 1 70 72 166.63 M C C CU1423A 18 H 3 70 72 169.63 M C C CU1423A 18 H 5 70 72 172.63 P F CU1423A 18 H CC 0 5 175.52 G A F C F FU1423A 19 H 1 70 72 176.78 P R C P P RU1423A 19 H 3 70 72 179.78 P R P P PU1423A 19 H 5 70 72 182.78 M F AU1423A 19 H CC 0 5 185.58 G C P C CU1423A 20 H 2 70 72 187.91 M F F FU1423A 20 H 4 70 72 190.91 M C C AU1423A 20 H 6 70 72 193.91 M C C AU1423A 20 H CC 0 4 195.15 G A P A P C AU1423A 21 H 2 70 72 197.71 M R F P AU1423A 21 H 4 70 72 200.71 M F AU1423A 21 H 6 70 72 203.71 P F R AU1423A 21 H CC 0 5 205.01 G A P C R AU1423A 22 H 2 70 72 207.39 M C C AU1423A 22 H 4 70 72 210.39 M C CU1423A 22 H 6 100 102 213.69 M A P C AU1423A 22 H CC 0 5 214.85 G A P A AU1423B 2 H 3 70 72 8.23 G C A AU1423B 2 H 4 70 72 9.73 G A AU1423B 2 H 5 70 72 11.23 G A F R CU1423B 2 H 6 70 72 12.73 M F F C CU1423B 3 H 2 70 72 16.60 G C A FU1423B 3 H 3 70 72 18.10 G C A F CU1423B 3 H 4 70 72 19.60 G C A AU1423B 3 H 5 70 72 21.10 G C A CU1423B 3 H 6 70 72 22.60 M F A FU1423B 3 H 7 20 22 23.60 G C A

No. 1 2 3 4 5 6 7 8 9 10 11 12
U1423B 4 H 2 100 102 26.76 G A A P
U1423B 4 H 3 70 72 27.93 P VR P
U1423B 4 H 4 70 72 29.43 G C A R C
U1423B 4 H 5 70 72 30.93 G A R C
U1423B 4 H 6 68 70 32.41 G C A A R
U1423B 4 H 7 40 42 33.63 M F A F R R
U1423B 5 H 2 120 122 36.73 G C A F
U1423B 5 H 3 70 72 37.73 G C A
U1423B 5 H 4 70 72 39.23 G A A R C
U1423B 5 H 5 69 71 40.72 G C P A
U1423B 5 H 6 70 72 42.23 P R
U1423B 6 H 3 70 72 47.94 M F A F
U1423B 6 H 4 70 72 48.94 G C C
U1423B 6 H 5 70 72 50.94 G C F
U1423B 6 H 6 58 60 52.32 G C C R C
U1423B 7 H 2 140 142 56.60 G C F
U1423B 7 H 3 70 72 57.40 G A A
U1423B 7 H 4 70 72 58.90 G C C C F F
U1423B 7 H 5 70 72 60.40 G A A C C
U1423B 7 H 6 70 72 61.90 G A A A F
U1423B 8 H 3 70 72 67.09 G A
U1423B 8 H 4 70 72 68.59 M C P
U1423B 8 H 5 70 72 70.09 P R P
U1423B 8 H 6 70 72 71.59 G M
U1423B 9 H 3 70 72 77.11 G A R P
U1423B 9 H 4 70 72 78.61 M A F P P
U1423B 9 H 5 70 72 80.11 G A F R
U1423B 9 H 6 70 72 81.61 P R
U1423B 10 H 3 70 72 87.31 P R A
U1423B 10 H 4 74 76 88.85 P R F
U1423B 10 H 5 70 72 90.31 M C A
U1423B 10 H 6 30 32 91.41 P C P
U1423B 22 H CC 0 5 199.34 G A A P P A
U1423B 23 H CC 0 5 209.20 G A C P A
U1423B 24 H 4 140 142 215.12 M C F C
U1423B 24 H 6 70 72 217.42 M C C A
U1423B 24 H CC 0 5 218.86 G A R A P A
U1423B 25 H 2 70 72 220.92 M C F A
U1423B 25 H 4 70 72 223.92 M C F A
U1423B 25 H 6 70 72 226.92 M C F A
U1423B 25 H CC 0 5 228.17 G A R C R A
U1423B 26 H 2 70 72 230.42 M C F F C
U1423B 26 H 4 70 72 233.42 G C F A
U1423B 26 H 6 70 72 236.42 G C F C
U1423B 26 H CC 0 5 237.72 G A F C R F
U1423B 27 H 2 70 72 239.92 G A R F F
U1423B 27 H 4 70 72 242.92 G A F P F
U1423B 27 H 6 70 72 245.92 G A F R
U1423B 27 H CC 0 5 247.34 G A P C R F
U1423B 28 H 2 70 72 249.42 G A F R
U1423B 28 H 4 70 72 252.42 M A F
U1423B 28 H 6 70 72 255.42 G A P R
U1423B 28 H CC 0 5 256.92 G A P C F R

No. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19H
ole
Cor
e
Type
Sec
t
Inte
rval
Top
(cm
)
Inte
rval
Bot
tom
(cm
)
CC
SF-
A(m
)
Pre
serv
atio
n
Gro
up A
bund
ance
Am
ph
ime
lissa
se
tosa
Axo
pru
nu
m a
cq
uilo
niu
m
Ca
locycla
s m
oto
ya
ma
i
Cycla
do
ph
ora
da
vis
ian
a
Cycla
do
ph
ora
na
ka
se
ko
i
Cycla
do
ph
ora
sp
ha
eri
s
Cyrt
oca
pse
lla t
etr
ap
era
Dic
tyo
ph
imu
s b
ulla
tus
Eu
cyrt
idiu
m in
fla
tum
He
xa
co
ntiu
m a
kita
en
sis
e
He
xa
co
ntiu
m p
arv
iakita
en
se
La
rco
pyle
pylo
ma
ticu
s
Lip
ma
ne
lla r
ed
on
do
en
sis
Lith
eliu
s b
arb
atu
s
Lych
no
ca
no
ma
ma
gn
aco
rnu
ta
Lych
no
ca
no
ma
pa
ralle
lipe
s
Lych
no
ca
no
ma
sa
ka
ii
Sip
ho
ca
mp
e a
rach
ne
a gr
oup
Sch
izo
dis
cu
s ja
po
nic
us
U1425B 1 H 4 68 70 5.18 M R A
U1425B 1 H 5 98 100 6.98 M A C F C
U1425B 1 H CC 0 5 8.70 P R F R F F
U1425B 2 H 5 64 66 16.00 G A A P
U1425B 2 H 5 70 72 16.06 M VR
U1425B 2 H CC 0 5 19.05 M R F F P F R P R
U1425B 3 H 4 51 53 24.89 M A C R R A
U1425B 3 H 4 68 70 25.06 M F
U1425B 3 H 5 112 114 27.00 M C A P F
U1425B 3 H CC 0 5 28.38 M C R P A
U1425B 4 H 5 68 70 36.58 M F A
U1425B 4 H 7 31 33 39.02 M C F F R
U1425B 4 H CC 0 5 39.39 G A R A F
U1425B 5 H 5 31 33 46.02 M A P F
U1425B 5 H 5 71 73 46.42 M F
U1425B 5 H 6 81 83 48.02 G A C R F F R
U1425B 5 H CC 0 5 49.25 G C R R
U1425B 6 H 2 61 63 53.01 G A A P
U1425B 6 H 3 111 113 55.01 G C F F P
U1425B 6 H 4 68 70 56.08 M F C
U1425B 6 H 5 11 13 57.01 G A R F R
U1425B 6 H CC 0 5 60.49 M C R
U1425B 7 H 5 71 73 67.69 M VR F A F R
U1425B 7 H 5 103 105 68.01 P C R R
U1425B 7 H CC 0 5 70.57 P R C F R P
U1425B 8 H 3 137 139 76.00 M A R A P A
U1425B 8 H 4 71 73 76.84 M F P A F R
U1425B 8 H 5 137 139 79.00 M A R A F C
U1425B 8 H 7 38 40 81.01 M A R F R F A
U1425B 8 H CC 0 5 81.19 M C R R R F F
U1425B 9 H 3 4 6 84.99 P R R
U1425B 9 H 3 71 73 85.66 M F F A F
U1425B 9 H 4 54 56 86.99 M A P R F R
U1425B 9 H CC 0 5 91.54 G A R P F C F
U1425B 10 H 5 71 73 95.78 M R P A P P
U1425B 10 H 5 93 95 96.00 G A A R R
U1425B 10 H CC 0 5 98.67 G A F C F R P
U1425B 11 H 4 70 72 104.59 M F A F
U1425B 11 H 5 61 63 106.00 G C C R R F
U1425B 11 H CC 0 5 108.90 G C R R C P R
U1425B 12 H CC 0 5 #N/A G A R C F C R
U1425B 13 X CC 0 5 111.38 G A F C C C F
U1425B 14 H 3 58 60 115.00 G A P A R R F
U1425B 14 H 4 70 72 116.65 M F F A F R
U1425B 14 H 5 55 57 118.00 G A R A F R F
U1425B 14 H 6 105 107 120.00 G A R C P R F
U1425B 14 H CC 0 5 121.03 G A R C R C F F
U1425B 15 H CC 0 3 120.92 G A P F F C P
U1425B 16 X CC 0 4 122.05 G A C A C R F
U1425B 17 H 2 123 125 124.99 G C A R R R
U1425B 17 H 3 70 72 125.96 M R A F P
U1425B 17 H CC 0 5 126.76 G A F R A R
U1425B 18 H CC 0 5 131.91 G A C C C
U1425B 19 H CC 0 6 136.84 G C P C F F C
U1425B 20 H CC 0 6 143.10 G C P C R F
U1425B 21 H 2 20 22 144.02 G A R A 2 P A
U1425B 21 H 4 19 21 147.01 G A R A 2 R R A
U1425B 21 H 6 19 21 150.01 G A A R A
U1425B 21 H CC 0 5 151.97 G A R C C A
U1425B 22 H 3 110 112 156.98 G A P C R A
U1425B 22 H 5 10 12 158.98 G A P C F A
U1425B 22 H CC 0 5 162.60 G A P F F P F
U1425B 23 H 3 129 131 167.01 G A C F F
U1425B 23 H 5 129 131 170.01 G A R A F P R
U1425B 23 H CC 0 5 172.33 G A R C F
U1425B 24 H 2 144 146 175.99 G A C F F
U1425B 24 H CC 0 5 182.78 G A R P F P R
U1425B 25 H 2 90 92 184.99 M A F A R R
U1425B 25 H 4 91 93 188.00 G A P A P F
U1425B 25 H 6 91 93 191.00 M A F A R F F

No. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
U1425B 25 H CC 0 5 192.27 G A P C P F
U1425B 26 H CC 0 5 203.41 G A P C C F P
U1425B 27 H 4 27 29 207.00 G A C R A
U1425B 27 H 6 26 28 209.99 G A F C R A
U1425B 27 H CC 0 5 211.93 G C F F R F
U1425B 28 H 3 147 149 217.00 G A R A R A
U1425B 28 H CC 0 5 221.00 G R C R R R
U1425B 29 H 1 81 83 222.01 G A R C P A F R
U1425B 29 H 3 81 83 225.01 G A R C R A F F
U1425B 29 H 5 82 84 228.02 G A P A R A F F
U1425B 29 H CC 0 5 230.70 G A P C R A C
U1425B 30 H 3 31 33 233.00 G A R A F A R R
U1425B 30 H 5 31 33 236.00 G A C R A F P
U1425B 30 H CC 0 5 239.32 G A R A F R F
U1425B 32 X CC 0 5 244.29 G A A R F
U1425B 33 H 3 39 41 246.99 G A C P R R R
U1425B 33 H 4 89 91 248.99 G A A R R
U1425B 33 H 6 90 92 252.00 G A A R C R
U1425B 33 H CC 0 5 253.16 G A C F F F
U1425B 34 H 4 46 48 258.00 M A F R R
U1425B 34 H CC 0 6 262.64 M C A R F P
U1425B 35 H 3 47 49 266.00 M A A C F P F
U1425B 35 H 5 47 49 269.00 M A A C
U1425B 35 H CC 0 5 271.74 G A C F
U1425B 36 H CC 0 5 277.60 G A A
U1425B 37 H 1 128 130 279.00 M A A R R
U1425B 37 H CC 0 5 282.45 G C A P
U1425B 38 H 1 101 103 283.00 M A C C R P
U1425B 38 H CC 0 5 286.71 G A A F
U1425B 39 H 2 125 127 290.00 G A A R
U1425B 39 H CC 0 6 292.01 G A A F
U1425B 40 X 1 109 111 293.00 M C R R
U1425B 40 X 3 9 11 295.00 M A C R
U1425B 40 X CC 0 5 297.05 G A A P
U1425B 41 H CC 0 5 301.38 G A A F
U1425B 42 H CC 0 5 307.10 G A A P
U1425B 43 H CC 0 5 311.82 G A A P
U1425B 44 H 4 4 6 316.00 G A C F
U1425B 44 H CC 0 5 316.51 G A A F
U1425B 45 H CC 0 5 320.57 G A A F
U1425B 46 H CC 0 5 325.22 M A C C
U1425B 47 H 1 91 93 326.01 G A A F
U1425B 47 H 3 91 93 329.01 G A A P
U1425B 47 H CC 0 5 329.81 G F P
U1425B 48 H CC 0 5 335.55 G F F F
U1425B 49 H 2 129 131 339.01 G A A P P
U1425B 49 H CC 0 5 340.94 G F C
U1425B 50 H 2 66 68 343.98 G A C F C
U1425B 50 H 3 117 119 345.99 M A A R F
U1425B 50 H CC 0 5 346.57 G A A P
U1425B 51 H CC 0 5 352.15 M C A F P
U1425B 52 H CC 0 5 356.82 M C A P R
U1425B 53 H CC 0 5 361.58 M C F F C
U1425B 54 H CC 0 5 366.18 P R F P P
U1425B 55 X CC 0 5 375.41 P R P F
U1425B 56 X CC 0 5 381.25 P R F P
U1425B 57 X CC 0 5 384.60 P VR
U1425B 58 X CC 0 5 393.63 P VR
U1425B 59 X CC 0 2 409.94 B
U1425B 60 H CC 0 5 417.66 M R P F P
U1425B 61 X CC 0 4 419.15 P R
U1425D 57 H CC 0 5 390.86 P R R F F
U1425D 58 H CC 0 5 394.93 P R R
U1425D 59 H CC 0 5 399.77 P R P C C
U1425D 60 H CC 0 5 404.41 P R P P
U1425D 61 H CC 0 5 405.82 P R A F C
U1425D 62 H CC 0 5 408.67 P VR C C
U1425D 63 H CC 0 5 413.06 P VR P R P
U1425D 64 H CC 0 5 416.41 P VR A P
U1425D 65 H CC 0 5 419.82 P R C R
U1425D 66 H CC 0 5 423.42 P R
U1425D 67 H CC 0 5 424.26 P VR F P
U1425D 69 H CC 0 5 429.40 P VR
U1425D 70 H CC 0 1 430.40 B

No. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26H
ole
Core
Typ
e
Sec.
Inte
rval
Top
(cm
)
Inte
rval
Bot
tom
(cm
)
CC
SF-A
(m
)
PR
ESER
VA
TIO
N
AB
UN
DA
NC
E
Am
phim
elis
sa s
etos
a
Axo
prun
um a
cqui
loni
um
Cal
ocyc
las
mot
oyam
ai
Cyc
lado
phor
a da
visi
ana
Cyc
lado
phor
a na
kase
koi
Cyc
lado
phor
a sp
haer
is
Cyr
toca
psel
la ja
poni
ca
Cyr
toca
psel
la t
etra
pera
Dic
tyop
him
us b
ulla
tus
Den
dros
pyris
saka
ii
Den
dros
pyris
uruy
aens
is
Euc
yrtidi
um a
sano
i
Euc
yrtidi
um in
flatu
m
Hex
acon
tium
aki
taen
se
Hex
acon
tium
par
viak
itae
nse
Lar
copy
le p
ylom
atic
us
Lip
man
ella
his
ter
Lip
man
ella
red
ondo
ensi
s
Lithe
lius
barb
atus
Litho
pera
ren
zae
Lyc
hnoc
anom
a m
agna
corn
uta
Lyc
hnoc
anom
a pa
ralle
lipes
Lyc
hnoc
anom
a sa
kaii
Sip
hoca
mpe
ara
chne
a gr
oup
Sch
izod
iscu
s jo
poni
scus
Sty
losp
haer
a m
agna
poru
losa
U1430A 1 H CC 3.74 P R F R
U1430A 2 H 4 68 70 9.14 P vR R R
U1430A 2 H CC 13.63 P R R
U1430A 3 H 4 68 70 19.28 M F R R
U1430A 3 H CC 23.88 G A C F
U1430A 4 H 4 68 70 29.59 M F C
U1430A 4 H CC 33.99 G A R A
U1430A 5 H 4 68 70 39.23 G F R R R
U1430A 5 H CC 44.00 G F F R
U1430A 6 H 4 68 70 49.15 M F R R
U1430A 6 H CC 53.74 G A F C
U1430A 7 H 4 68 70 59.20 G F R R F R
U1430A 7 H CC 63.92 G F R F R R F
U1430A 8 H 4 68 70 69.22 M F R R R F R
U1430A 8 H CC 73.83 G C F C R F C
U1430A 9 H 1 68 70 74.33 G F R C C
U1430A 9 H 2 68 70 75.83 G F R C R C
U1430A 9 H 3 68 70 77.33 G F R F F
U1430A 9 H 4 8 10 78.23 G C R F R
U1430A 9 H 4 68 70 78.83 G F R F R
U1430A 9 H 4 120 122 79.35 G F R F R R
U1430A 9 H 5 11 13 79.76 G F R C R R
U1430A 9 H 5 70 72 80.35 G C C R
U1430A 9 H 6 70 72 81.85 G F R C
U1430A 9 H CC 83.45 G C F C F
U1430A 10 H 1 70 72 84.75 G F F C
U1430A 10 H 2 70 72 86.25 G F F C R
U1430A 10 H 3 68 70 87.73 G F F F R
U1430A 10 H 4 68 70 89.23 G F F R
U1430A 10 H 5 70 72 90.75 G F F R
U1430A 10 H 6 70 72 92.25 G F F R
U1430A 10 H CC 93.91 G C C F
U1430A 11 H 4 68 70 99.35 G F F R
U1430A 11 H CC 103.91 G F A R R
U1430A 12 H 4 68 70 109.56 G F C R
U1430A 12 H CC 114.19 G F A R R
U1430A 13 H 4 68 70 119.82 G F F
U1430A 13 H CC 124.47 G C A R R
U1430A 14 H 4 68 70 129.73 G R C
U1430A 14 H CC 134.37 G C A F F
U1430A 15 H 4 68 70 139.65 G C C R
U1430A 15 H CC 144.28 G A A R F
U1430A 16 H 1 70 72 145.44 G F R
U1430A 16 H 2 70 72 146.94 G F R R
U1430A 16 H 3 70 72 148.44 G F R
U1430A 16 H 4 70 72 149.94 G F R
U1430A 16 H 5 68 70 151.42 G F R R R
U1430A 16 H 6 70 72 152.94 G F F R
U1430A 16 H CC 154.61 G C F F R
U1430A 17 H 1 70 72 156.81 M F R R R
U1430A 17 H 2 70 72 158.31 G F F R
U1430A 17 H 3 68 70 159.79 G C C R
U1430A 17 H 4 70 72 161.33 G F C R
U1430A 17 H 5 68 70 162.83 G R A
U1430A 17 H 6 70 72 164.33 G F A
U1430A 17 H CC 165.97 G C A
U1430A 18 H 4 68 70 170.59 G F F
U1430A 18 H CC 175.25 G F A R R R
U1430A 19 H 4 70 72 180.94 M F F R
U1430A 19 H CC 185.60 G F F C F
U1430A 20 H 4 68 70 191.06 M R F
U1430A 20 H CC 194.76 G C F R F
U1430A 21 H 4 68 70 200.65 M R R
U1430A 21 H CC 204.87 M C F
U1430A 22 H 4 68 70 211.39 G F R
U1430A 22 H CC 215.41 G C R R F F
U1430A 23 H 4 68 70 220.91 M F R R F
U1430A 23 H CC 225.34 G C R F F
U1430A 24 H 4 68 70 230.83 H F R F
U1430A 24 H CC 235.36 G C R F R
U1430A 25 H 1 70 72 235.61 M F R R
U1430A 25 H 2 70 72 237.11 G F R R
U1430A 25 H 3 70 72 238.61 G R R R
U1430A 25 H 4 70 72 240.11 G R R R
U1430A 25 H 5 68 70 241.59 G R F R R

No. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26H
ole
Core
Typ
e
Sec.
Inte
rval
Top
(cm
)
Inte
rval
Bot
tom
(cm
)
CC
SF-A
(m
)
PR
ESER
VA
TIO
N
AB
UN
DA
NC
E
Am
phim
elis
sa s
etos
a
Axo
prun
um a
cqui
loni
um
Cal
ocyc
las
mot
oyam
ai
Cyc
lado
phor
a da
visi
ana
Cyc
lado
phor
a na
kase
koi
Cyc
lado
phor
a sp
haer
is
Cyr
toca
psel
la ja
poni
ca
Cyr
toca
psel
la t
etra
pera
Dic
tyop
him
us b
ulla
tus
Den
dros
pyris
saka
ii
Den
dros
pyris
uruy
aens
is
Euc
yrtidi
um a
sano
i
Euc
yrtidi
um in
flatu
m
Hex
acon
tium
aki
taen
sisi
s
Hex
acon
tium
par
viak
itae
nsis
Lar
copy
le p
ylom
atic
us
Lip
man
ella
his
ter
Lip
man
ella
red
ondo
ensi
s
Lithe
lius
barb
atus
Litho
pera
ren
zae
Lyc
hnoc
anom
a m
agna
corn
uta
Lyc
hnoc
anom
a pa
ralle
lipes
Lyc
hnoc
anom
a sa
kaii
Sip
hoca
mpe
ara
chne
a gr
oup
Sch
izod
iscu
s jo
poni
scus
Sty
losp
haer
a m
agna
poru
losa
U1430A 25 H 6 70 72 243.11 G R F
U1430A 25 H CC 244.40 M C C F R
U1430A 26 H 1 70 72 246.00 M R F R R
U1430A 26 H 2 68 70 247.48 M F R R
U1430A 26 H 3 70 72 249.00 M R F
U1430A 26 H CC 250.24 M C F C
U1430A 27 H 3 68 70 253.69 G F F
U1430A 27 H CC 254.99 G A F C C R
U1430A 28 H 1 70 72 255.99 M R F R R
U1430A 28 H 3 25 27 258.56 P R R R R
U1430A 28 H CC 260.25 M A R R C R
U1430A 29 H CC 264.63 P F R R C A
U1430A 30 X CC 264.63 P R C R
U1430A 31 X CC 3 5 270.16 P R R R R R
U1430A 32 X CC 280.33 B B
U1430C 29 H CC 254.36 M C R R F
U1430C 30 H CC 259.30 M F R R
U1430C 31 H CC 260.69 P R C R F
U1430C 32 H CC 265.32 P R C F
U1430B 32 H 1 111 113 267.45 P F C R
U1430B 33 X 1 59 61 272.63 P C F





























