Embed Size (px)
Citation preview

JournalofEndocrinology
ResearchA DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 327–339
PACAP is essential for the adaptivethermogenic response of brownadipose tissue to cold exposure
Abdoulaye Diane, Nikolina Nikolic*, Alexander P Rudecki*, Shannon M King,
Drew J Bowie and Sarah L Gray
Northern Medical Program, University of Northern British Columbia, 3333 University Way,
Prince George, British Columbia, Canada V2N 4Z9*(N Nikolic and A P Rudecki contributed equally to this work)
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
Published by Bioscientifica Ltd.
Correspondence
should be addressed
to S L Gray
Abstract
Pituitary adenylate cyclase-activating polypeptide (PACAP) is a widely distributed neuro-
peptide that acts as a neurotransmitter, neuromodulator, neurotropic factor, neuroprotectant,
secretagogue, and neurohormone. Owing to its pleiotropic biological actions, knockout of
Pacap (Adcyap1) has been shown to induce several abnormalities in mice such as impaired
thermoregulation. However, the underlying physiological and molecular mechanisms remain
unclear. A previous report has shown that cold-exposed Pacap null mice cannot supply
appropriate levels of norepinephrine (NE) to brown adipocytes. Therefore, we hypothesized
that exogenous NE would rescue the impaired thermogenic response of Pacap null mice
during cold exposure. We compared the adaptive thermogenic capacity of PacapK/K to
PacapC/C mice in response to NE when housed at room temperature (24 8C) and after a
3.5-week cold exposure (4 8C). Biochemical parameters, expression of thermogenic genes, and
morphological properties of brown adipose tissue (BAT) and white adipose tissue (WAT) were
also characterized. Results showed that there was a significant effect of temperature, but no
effect of genotype, on the resting metabolic rate in conscious, unrestrained mice. However,
the normal cold-induced increase in the basal metabolic rate and NE-induced increase in
thermogenesis were severely blunted in cold-exposed PacapK/K mice. These changes were
associated with altered substrate utilization, reduced b3-adrenergic receptor (b3-Ar (Adrb3))
and hormone-sensitive lipase (Hsl (Lipe)) gene expression, and increased fibroblast growth
factor 2 (Fgf2) gene expression in BAT. Interestingly, PacapK/Kmice had depletedWAT depots,
associated with upregulated uncoupling protein 1 expression in inguinal WATs. These results
suggest that the impairment of adaptive thermogenesis in Pacap null mice cannot be rescued
by exogenous NE perhaps in part due to decreased b3-Ar-mediated BAT activation.
Key Words
" PACAP
" thermogenesis
" cold
" gene expression
" mice
Journal of Endocrinology
(2014) 222, 327–339
Introduction
All mammals, including humans, are homeotherms – they
maintain euthermia regardless of environmental tempera-
ture by hormonal and neuronal control of heat pro-
duction and dissipation. This essential task is mainly
performed by the adipose tissues (Cannon & Nedergaard
2004, Cypess et al. 2009, Ouellet et al. 2011) although
skeletal muscle can also contribute to increasing heat
production via shivering thermogenesis in mammals

JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 328
(Dubois-Ferriere and Chinet 1981). The adipose tissue pool
in mammals is composed of at least two functionally
different types of fat: white adipose tissue (WAT) and
brown adipose tissue (BAT). WAT is the primary site of
energy storage and releases hormones and cytokines that
modulate whole-body metabolism (Richard & Picard
2011). BAT, on the other hand, contributes to overall
energy expenditure in small mammals and neonates
through the process of non-shivering, adaptive thermo-
genesis. In the last few years, several studies have reported
the existence of a third type of adipose cell, the brown in
white (‘brite’) or ‘beige’ adipocyte. As in canonical BAT,
the recruitment of brite adipocytes or ‘browning’ of WATs
is induced by thermogenic stimuli such as cold as well as
by pharmacological treatments such as b-adrenergic
agonists or thiazolidinediones (Ohno et al. 2012, Wu
et al. 2012, Schulz & Tseng 2013). Brite adipocytes not only
present gene expression signatures similar to those of
canonical brown adipocytes (i.e. uncoupling protein 1
(Ucp1)) but also express unique genes such as Hoxc9,
Tmem26, and Tbx1 (Walden et al. 2012, Wu et al. 2012).
In rodents, BAT generates heat for two principal
reasons: i) to protect against cold exposure and ii) to
burn off excess calories in response to excess caloric intake
(Lowell et al. 1993, Ghorbani et al. 1997, Guerra et al. 1998,
Clapham & Arch 2011). The exceptional thermogenic
capacity of BAT relies on its numerous, densely packed
mitochondria containing the BAT-specific inner mito-
chondrial membrane protein UCP1. BAT is highly
vascularized and richly innervated by postganglionic
nerve terminals of the sympathetic nervous system (SNS;
Baron et al. 2012, Vaughan et al. 2014). Thermoregulatory
pathways are induced by chemical messengers of the SNS,
the catecholamines (Thomas & Palmiter 1997); although
circulating thyroid hormones (thyroxine (T4)) as well as
bone morphogenetic protein 8B are also known to
regulate or enhance obligatory thermogenesis by acting
centrally to increase sympathetic output to BAT (Ricquier
et al. 2000, Lopez et al. 2010, Whittle et al. 2012). Over the
past 7 years, a number of molecules including fibroblast
growth factor 21 (FGF21) and brain natriuretic peptides
(BNPs) have also been shown to activate thermogenic
machinery in BAT, independent of adrenergic receptors
(ARs) (Tseng et al. 2008, Hondares et al. 2010).
The hypothalamus responds to afferent signals from
cutaneous and core thermoreceptors to detect cold and
then activates adaptive thermogenesis contributing to the
maintenance of body temperature (Perkins et al. 1981,
Cannon & Nedergaard 2004). Cumulative evidence has
shown various hypothalamic neuropeptides to be
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
important regulators of BAT thermogenesis through the
SNS (Bi & Li 2013, Zengin et al. 2013). The hypothalamic
neuropeptide pituitary adenylate cyclase-activating poly-
peptide (PACAP) is known to regulate sympathetic nerve
activity, yet its role in adaptive thermogenesis requires
further characterization.
PACAP belongs to the secretin/glucagon/vasoactive
intestinal (VIP) family (Miyata et al. 1989, Vaudry et al.
2009) and has been implicated in the regulation of energy
homeostasis including both appetite and thermogenesis
(Gray et al. 2001, Mounien et al. 2009, Inglott et al. 2011).
Hypothalamic nuclei such as the arcuate nucleus, para-
ventricular nucleus, and ventromedial nucleus highly
express PACAP (ADCYAP1) and PACAP receptors (Segal
et al. 2010, Kohno & Yada 2012, Resch et al. 2013),
suggesting that PACAP may be critical for the regulation
of energy balance. Consistently, i.c.v. injection of PACAP
decreased food intake and increased core body temperature
in rodents (Mounien et al. 2009) with a concurrent increase
in BAT UCP1 expression (Resch et al. 2013). In parallel, mice
lacking PACAP showed decreased BAT thermogenic
capacity and reduced sympathetic outflow to this organ
(Gray et al. 2001, 2002). These studies are supported by the
temperature-sensitive phenotype of Pacapnull pups, which
display reduced survival at a lower housing temperature
(Gray et al. 2002); however, the underlying mechanisms by
whichPacapnull mice are cold intolerant remain unknown.
A previous report (Gray et al. 2002) has shown that during
cold exposure, adult Pacap null mice cannot supply
appropriate levels of norepinephrine (NE) to BAT. NE
released from sympathetic nerves innervating BAT binds
b3-ARs increasing cAMP and hormone-sensitive lipase
(HSL)-mediated lipolysis. Free fatty acids are oxidized in
the mitochondria where UCP1 uncouples ATP production
releasing oxidative energy as heat (Enerback et al. 1997,
Lowell & Spiegelman 2000, Matthias et al. 2000, Cannon &
Nedergaard 2004, Inokuma et al. 2006). Interestingly, due to
the substantial demand for nutrients and oxygen by BAT
during cold exposure (Golozoubova et al. 2004, Walden
et al. 2012), extensive angiogenesis occurs within BAT to
provide sufficient blood supply (Xue et al. 2009). As PACAP
is known as a non-classical regulator of angiogenesis
(Castorina et al. 2010), the highly cold-sensitive phenotype
of Pacap-deficient mice may be related to decreased
vascularization in adipose tissues.
To determine the mechanism by which PACAP
regulates thermogenesis, we hypothesized that adminis-
tration of exogenous NE would rescue the impaired
thermogenic response of Pacap null mice to cold exposure.
We also hypothesized that reduced browning of WATs
Published by Bioscientifica Ltd.

JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 329
(as a consequence of inappropriate NE release) as well as
diminished vascularization in BATs and WATs during
chronic cold exposure would contribute to the impaired
adaptive thermogenic response of Pacap-deficient mice.
Materials and methods
Animals
Cohorts of young–adult (8-week-old) PacapK/K and
PacapC/C male or female littermate mice were generated
from the breeding colony at the University of Northern
British Columbia. Mice used in this study are derived from
the Pacap null line generated by Gray et al. (2001) and have
been backcrossed by more than ten times onto a C57/BL6
background. Mice were housed at a density of two per cage
with sterile corncob bedding and placed on a normal 12 h
light:12 h darkness cycle (lights on 0700–1900 h).
Throughout the experiment, animals had free access to
water and standard rodent chow diet (LabDiet 5001,
LabDiet, Inc., Brentwood, Leduc, AB, Canada; metaboliz-
able energyZ3.02 kcal/g). Measures of body weight (g)
were taken weekly. Care and treatment of mice was in
accordance with the guidelines of the Canadian Council
on Animal Care, and protocols for the study were
approved by the University of Northern British Colum-
bia’s Animal Care and Use Committee.
Experimental procedures
NE-induced thermogenesis in PacapK/K mice PacapK/K
and PacapC/C mice were paired and housed together
according to age and gender. Resting and maximal
metabolic rates (MMR) were measured in 8-week-old
mice reared at 24 8C. These mice were then acclimated at
18 8C for 1 week before being housed at 4 8C for 3.5 weeks.
After 3.5 weeks of housing at 4 8C, resting and MMR were
measured again in the same mice.
Metabolic rates of mice were measured via oxygen
consumption (ml O2/min) in an open circuit, indirect
calorimeter (Oxymax machine, Columbus Instruments,
Columbus, OH, USA). Body weights were not different
between genotypes. A standard Oxymax housing chamber
(2625 ml) was allowed to equilibrate for 20 min before the
resting metabolic rate (RMR) was measured in conscious,
unrestrained animals for 4–5 h at room temperature
(24 8C). The RMR was defined as the mean oxygen
consumption during the last 3 h of the experiment.
Basal metabolic rate (BMR) and MMR were measured
in anesthetized mice (sodium pentobarbital, 60–65 mg/kg)
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
in 10 s intervals, during the light phase of the cycle, before
and after s.c. injection of NE (1 mg/kg) into interscapular
BAT (iBAT). NE injection was administered via cannulae to
avoid opening and disequilibrium of the chamber during
NE administration (protocol adapted from Golozoubova
et al. (2006) and Virtue & Vidal-Puig (2013)). During BMR
and MMR measurements, a heating pad was placed under
the smaller metabolic chamber (260 ml) to maintain the
chamber at 30 8C. BMR was defined as the mean oxygen
consumption 5 min before the NE injection; while MMR
was the mean of the five highest oxygen consumption
values after NE administration. Additionally, the relative
exchange ratio or RER, which is the ratio of CO2 produced
to O2 consumed, was calculated.
Molecular mechanisms of impaired adaptive thermo-
genesis in Pacap-deficient mice following cold
exposure Molecular experiments were performed only
on male mice, as estrous cycle patterns and sexual
hormones in female rodents are known to impact
thermoregulatory responses (Bu & Lephart 2005, Uchida
et al. 2010). This experiment was conducted in two
independent batches of male PacapK/K and PacapC/C
mice. Within each genotype, 9-week-old mice
were randomly assigned to four experimental groups:
PacapK/K and PacapC/C mice housed at 24 8C (PacapK/K,
24 8C and PacapC/C, 24 8C) and PacapK/K and PacapC/C
mice housed at 4 8C for 3.5 weeks after they have been
acclimatized for 1 week at 18 8C (PacapK/K, 4 8C and
PacapC/C, 4 8C) as described above.
Postmortem analysis Mice (non-fasted) were killed with
Euthanyl. Blood was taken by cardiac puncture, from
which plasma was isolated and stored at K20 8C until
analysis. The following tissues were collected and
weighed: brain, liver, heart, spleen, lung, pancreas, iBAT,
inguinal WAT (ingWAT), and gonadal WAT (gWAT). For
each BAT and WAT depot, half was flash frozen in liquid
nitrogen and stored at K80 8C for RNA extractions, and
the other half was fixed in 10% formalin for histological
analysis.
Extraction of RNA and generation of cDNA iBAT
(w50 mg) was homogenized in TRIzol (Life Technologies)
and RNA was extracted according to the manufacturer’s
protocol. DNA contamination was removed by treating
iBAT RNA with TURBO DNase (Life Technologies) accor-
ding to the manufacturer’s protocol. RNA from ingWAT
and gWAT was extracted using an RNeasy Kit (Qiagen),
which included DNase treatment. Concentration and
Published by Bioscientifica Ltd.

JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 330
purity of RNA were assessed by spectrophotometry
(Nanodrop ND-1000; Thermo Scientific, Rockford, IL,
USA), while RNA integrity was assessed by visualizing
intact 18S and 28S rRNA bands on native 1.5% agarose gel.
RNA (500 ng) was reverse transcribed into cDNA (Super-
script III; Invitrogen) according to the manufacturer’s
protocol.
Quantitative real-time PCR
Expression levels of thermogenic genes (Ucp1, Hsl (Lipe),
b1-Ar (Adrb1), b2-Ar (Adrb2), b3-Ar (Adrb3), Fgf2, homeobox
C9 (Hoxc9), and vascular endothelial growth factor A
(Vegfa)) in each fat depot from mice housed at 24 and 4 8C
were assessed using quantitative real-time PCR (qPCR)
(Table 1). Reference gene stability across treatments and
genotypes was assessed via geNorm experiments, and a
maximum number of stable reference genes were used for
each gene expression analysis. Fold change in thermo-
genic gene expression from 24 to 4 8C samples was
calculated for each genotype.
Primers and probes (IDT, Coralville, IA, USA) were
either designed using the Beacon Designer Software
Table 1 List of the endogenous control (*) and target genes analyz
each gene were optimized using SYBR Green chemistry or TaqMan
Genes Gene abbreviat
18S rRNA 18S*
Glyceraldehyde 3-phosphate dehydrogenase Gapdh*
b-actin b-actin*
TATA-binding protein Tbp*
Ribosomal protein L19 Rpl19*
b1-adrenergic receptor b1-Ar
b2-adrenergic receptor b2-Ar
b3-adrenergic receptor b3-Ar
Hormone-sensitive lipase Hsl
Uncoupling protein 1 Ucp1
Fibroblast growth factor 2 Fgf2
Vascular endothelial growth factor A Vegfa
Homeobox C9 Hoxc9
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
(Premier Biosoft, Palo Alto, CA, USA) or taken from the
literature (sequences available upon request). An iQ5
thermocycler (Bio-Rad Laboratories) was used to conduct
25 ml reactions, which contained forward and reverse
primers (300 nM; Sigma), nuclease-free H2O, and 1/10
cDNA (3 ml). For genes utilizing SYBR Green chemistry, iQ
SYBR Green Supermix (1!) was included, while those using
the TaqMan method included iQ Supermix (1!) and probe
(150 nM). Primers and probes were optimized before qPCR
experiments, which conformed to the Minimum Infor-
mation for Publication of Quantitative Real-time PCR
Experiments (MIQEs) guidelines (Bustin et al. 2009).
Morphometric analysis
Formalin-fixed iBAT, ingWAT, and gWAT from PacapK/K
(nZ5) and PacapC/C (nZ6) mice were sent to Wax-it
Histology Services (Vancouver, BC, Canada) for paraffin
embedding, slide fixation (5 mm thick), and hematoxylin
and eosin staining.
Ten representative micrographs at 60! magnification
were taken (Olympus BX61) per hematoxylin and eosin-
stained iBAT slide. Using the CellSens Software (Olympus),
ed by qPCR in adipose depots. Primer and primer/probe sets for
respectively
ion Primer sequences (5 0–3 0)
F-CGGCTACCACATCCAAGGAAR-GCTGGAATTACCGCGGCTF-TGCACCACCAACTGCTTAGR-GGATGCAGGGATGATGTTCF-GCTCTGGCTCCTAGCACCATR-GCCACCGATCCACACAGAGTF-CACCAATGACTCCTATGAR-CCAAGATTCACGGTAGATAP-/5FAM/CCTATCACTCCTGCCACACCA/31ABkFQ/F-GAAGCTGATCAAGGATGGR-CTTCCCTATGCCCATATGP-/56FAM/CATCCGCAAGCCTGTGACTG/31ABkFQ/F-GTCGTCTCCTTCTACGTGR-GCTGTCGATCTTCTTTACCF-GTTCGAGCGACTACAAACR-TAGGCCCATGACTAGATCF-CAACCCGGTCATCTACTGR-ACCGTAGCTACACAGAAGF-GGAGCACTACAAACGCAACGAR-TCGGCCACCGGTAAAGAGF-CCTGGCAGATATCATCACR-TCACCTTGGATCTGAAGGF-AACCGGTACCTTGCTATGAAGR-GTTCGTTTCAGTGCCACATACF-AGACAGAACAAAGCCAGAAATCACR-CACGTCTGCGGATCTTGGACF-GCAGCAAGCACAAAGAGGAR-CGTCTGGTACTTGGTGTAGGG
Published by Bioscientifica Ltd.
JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 331
bright-field images were converted into gray scale. An
intensity threshold (gray channel: minZ145 and maxZ
256) was set that best represented lipid droplets (white area)
in iBAT sections, and these areas were quantified. Theses
areas were normalized to their respective image areas.
24 °CFemales
P=0.09 P=0.2
RM
Rsu
mpt
ion;
ml/m
in)
4
3
2
4 °C
Immunohistochemistry
Sections of iBAT and ingWAT (paraffin, 5 mm thick;
nZ5/group, Wax-it Histological Services) were deparaf-
finized and rehydrated as described previously (Riedel
2010). Antigen retrieval was completed in Tris–EDTA
buffer (pH 9) for 20 min at 95 8C. Sections were blocked
with serum-free solution (DAKO, Burlington, ON, Canada)
before being incubated at 4 8C overnight with a rabbit
anti-CD31 primary antibody (1:100; ab28364; Abcam,
Toronto, ON, Canada). Sections were then incubated for
an hour with the Alexa Fluor 594 donkey anti-rabbit
antibody (1:1000; A21207; Invitrogen) and fixed using
VECTRASHIELD Hardset Mounting Medium with DAPI
(Vector Labs, Burlington, ON, Canada). Samples were
visualized using a fluorescent light microscope (Olympus
BX61) and images for iBAT were analyzed using the
CellSens Software (Olympus).
Vasculature was quantified in CD31-stained iBAT
by taking nine representative micrographs at 20!
magnification for each sample. An average proportion of
CD31-positive area was quantified using the automatic
threshold option in CellSens (Olympus).
* P=0.1
Pacap+/+
Males
Pacap–/–
(O2
con
min
)
1
0
4
3
24 °C
4 °C
Plasma biochemical analysis
Total plasma free fatty acid concentrations were measured
using direct colorimetric enzymatic reactions (Cell
Biolabs, Inc., San Diego, CA, USA). Plasma BNPs and
T4 were measured using specific ELISA (Elabscience
Biotechnology Co. Ltd, Beijing, China) kits for mice.
Pacap+/+ Pacap–/–
RM
R(O
2 co
nsum
ptio
n; m
l/
2
1
0
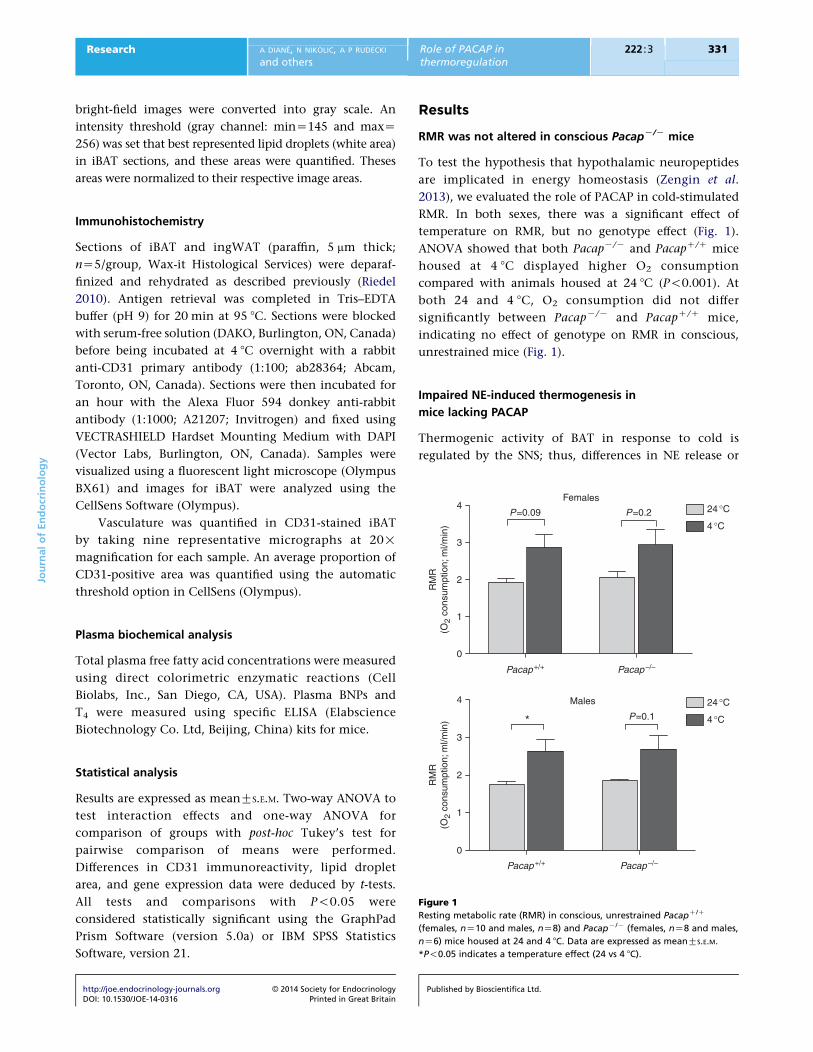
Figure 1
Resting metabolic rate (RMR) in conscious, unrestrained PacapC/C
(females, nZ10 and males, nZ8) and PacapK/K (females, nZ8 and males,
nZ6) mice housed at 24 and 4 8C. Data are expressed as meanGS.E.M.
*P!0.05 indicates a temperature effect (24 vs 4 8C).
Statistical analysis
Results are expressed as meanGS.E.M. Two-way ANOVA to
test interaction effects and one-way ANOVA for
comparison of groups with post-hoc Tukey’s test for
pairwise comparison of means were performed.
Differences in CD31 immunoreactivity, lipid droplet
area, and gene expression data were deduced by t-tests.
All tests and comparisons with P!0.05 were
considered statistically significant using the GraphPad
Prism Software (version 5.0a) or IBM SPSS Statistics
Software, version 21.
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
Results
RMR was not altered in conscious PacapK/K mice
To test the hypothesis that hypothalamic neuropeptides
are implicated in energy homeostasis (Zengin et al.
2013), we evaluated the role of PACAP in cold-stimulated
RMR. In both sexes, there was a significant effect of
temperature on RMR, but no genotype effect (Fig. 1).
ANOVA showed that both PacapK/K and PacapC/C mice
housed at 4 8C displayed higher O2 consumption
compared with animals housed at 24 8C (P!0.001). At
both 24 and 4 8C, O2 consumption did not differ
significantly between PacapK/K and PacapC/C mice,
indicating no effect of genotype on RMR in conscious,
unrestrained mice (Fig. 1).
Impaired NE-induced thermogenesis in
mice lacking PACAP
Thermogenic activity of BAT in response to cold is
regulated by the SNS; thus, differences in NE release or
Published by Bioscientifica Ltd.
0.0
0.5
1.0
1.5
2.0
2.5Pacap–/–, 24 °CPacap–/–, 4 °CPacap+/+, 24 °CPacap+/+, 4 °C
Pacap–/–, 24 °CPacap–/–, 4 °CPacap+/+, 24 °CPacap+/+, 4 °C
NE injection
Females
**
Time (min)
O2
cons
umpt
ion
(ml/m
in)
O2
cons
umpt
ion
(ml/m
in)
Males
2 10 20 30 40 50 60 70
2 10 20 30 40 50 60 700.0
0.5
1.0
1.5
2.0
2.5
NE injection *
*
Time (min)
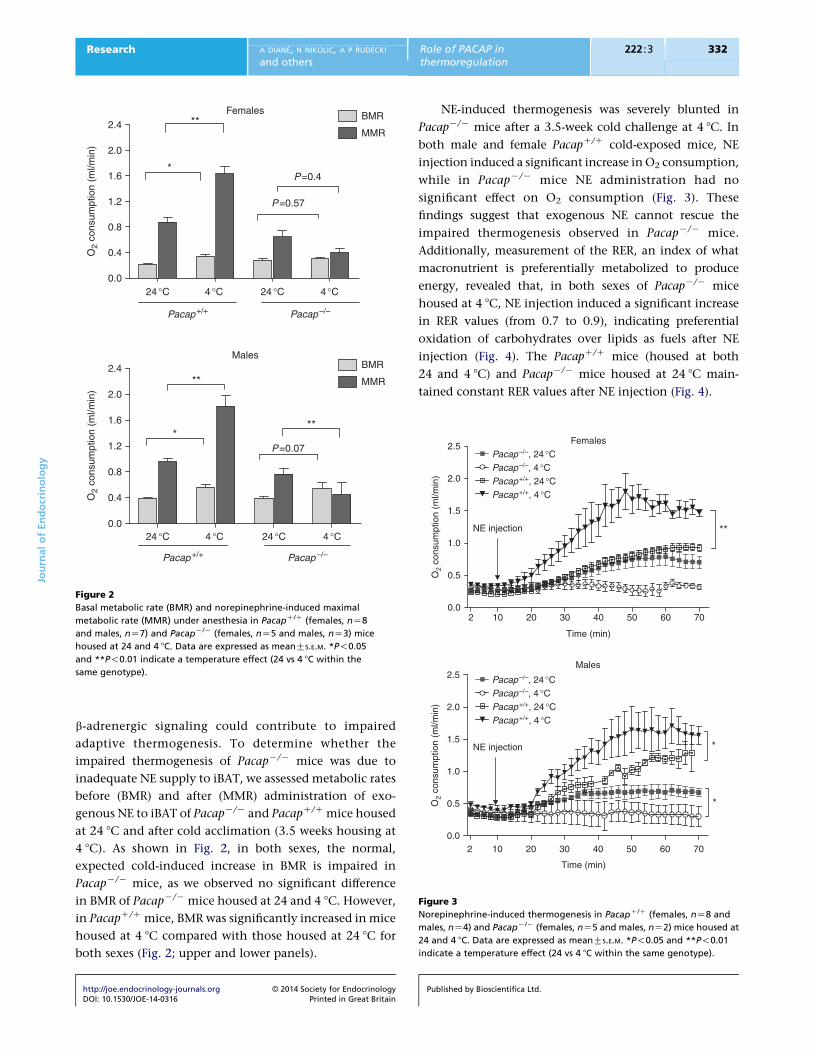
Figure 3
Norepinephrine-induced thermogenesis in PacapC/C (females, nZ8 and
males, nZ4) and PacapK/K (females, nZ5 and males, nZ2) mice housed at
24 and 4 8C. Data are expressed as meanGS.E.M. *P!0.05 and **P!0.01
indicate a temperature effect (24 vs 4 8C within the same genotype).
Females
24 °C 4 °C 24 °C 4 °C
24 °C 4 °C 24 °C 4 °C
0.0
0.4
0.8
1.2
1.6
2.0
2.4BMR
MMR
BMR
MMR
*
**
P=0.4
P=0.57
P=0.07
Pacap+/+ Pacap–/–
Pacap+/+ Pacap–/–
O2
cons
umpt
ion
(ml/m
in)
O2
cons
umpt
ion
(ml/m
in)
Males
0.0
0.4
0.8
1.2
1.6
2.0
2.4
*
**
**
Figure 2
Basal metabolic rate (BMR) and norepinephrine-induced maximal
metabolic rate (MMR) under anesthesia in PacapC/C (females, nZ8
and males, nZ7) and PacapK/K (females, nZ5 and males, nZ3) mice
housed at 24 and 4 8C. Data are expressed as meanGS.E.M. *P!0.05
and **P!0.01 indicate a temperature effect (24 vs 4 8C within the
same genotype).
JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 332
b-adrenergic signaling could contribute to impaired
adaptive thermogenesis. To determine whether the
impaired thermogenesis of PacapK/K mice was due to
inadequate NE supply to iBAT, we assessed metabolic rates
before (BMR) and after (MMR) administration of exo-
genous NE to iBAT of PacapK/K and PacapC/C mice housed
at 24 8C and after cold acclimation (3.5 weeks housing at
4 8C). As shown in Fig. 2, in both sexes, the normal,
expected cold-induced increase in BMR is impaired in
PacapK/K mice, as we observed no significant difference
in BMR of PacapK/K mice housed at 24 and 4 8C. However,
in PacapC/C mice, BMR was significantly increased in mice
housed at 4 8C compared with those housed at 24 8C for
both sexes (Fig. 2; upper and lower panels).
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
NE-induced thermogenesis was severely blunted in
PacapK/K mice after a 3.5-week cold challenge at 4 8C. In
both male and female PacapC/C cold-exposed mice, NE
injection induced a significant increase in O2 consumption,
while in PacapK/K mice NE administration had no
significant effect on O2 consumption (Fig. 3). These
findings suggest that exogenous NE cannot rescue the
impaired thermogenesis observed in PacapK/K mice.
Additionally, measurement of the RER, an index of what
macronutrient is preferentially metabolized to produce
energy, revealed that, in both sexes of PacapK/K mice
housed at 4 8C, NE injection induced a significant increase
in RER values (from 0.7 to 0.9), indicating preferential
oxidation of carbohydrates over lipids as fuels after NE
injection (Fig. 4). The PacapC/C mice (housed at both
24 and 4 8C) and PacapK/K mice housed at 24 8C main-
tained constant RER values after NE injection (Fig. 4).
Published by Bioscientifica Ltd.

0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1.1
1.2
NE injection
Females
*
Time (min)
RE
R, V
CO
2/V
O2
RE
R, V
CO
2/V
O2
Males
10 20 30 40
10 20 30 400.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1.1
1.2
NE injection
*
Time (min)
Pacap–/–, 24 °CPacap–/–, 4 °CPacap+/+, 24 °CPacap+/+, 4 °C
Pacap–/–, 24 °CPacap–/–, 4 °CPacap+/+, 24 °CPacap+/+, 4 °C
Figure 4
Relative exchange ratio (RER) at 24 8C and after 3.5 weeks of cold exposure
(4 8C) in PacapC/C (females, nZ8 and males, nZ4) and PacapK/K (females,
nZ5 and males, nZ2) mice. Data are expressed as meanGS.E.M. *P!0.05
indicates a temperature effect (24 vs 4 8C within the same genotype).
Table 2 Body composition and plasma circulating factors of
male PacapC/C (nZ9) and PacapK/K (nZ8) mice after 3.5-week
cold challenge. Data are expressed as meanGS.E.M.
PacapC/C PacapK/K
Body weight (g) 26.86G0.68 26.93G0.59iBAT (g) 0.10G0.01 0.09G0.001
JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 333
Cold exposure did not induce changes in body weight or
fat histology but significantly reduced s.c. and
intra-abdominal WAT depots in mice lacking PACAP
Body composition was assessed in PacapK/K and PacapC/C
male mice after 3.5 weeks of cold exposure. There were no
significant differences in body weight, and iBAT, pancreas,
and liver weights between the two genotypes. However,
ingWAT and gWAT depots were significantly lower in
cold-exposed PacapK/K mice compared with PacapC/C
mice (P!0.05; Table 2).
ingWAT (g) 0.21G0.02 0.16G0.01*gWAT (g) 0.21G0.02 0.12G0.02*Liver (g) 1.66G0.08 1.60G0.05Pancreas (g) 0.38G0.04 0.35G0.03Glucose (mmol/l) 10.11G0.71 7.92G1.29Free fatty acids (mM/ml) 154.34G32.12 222.15G55.73BNP (ng/ml) 31.99G1.24 34.60G1.46T4 (ng/ml) 1.91G0.33 1.83G0.28
*P!0.05 indicates a genotype effect (PacapC/C vs PacapK/K).
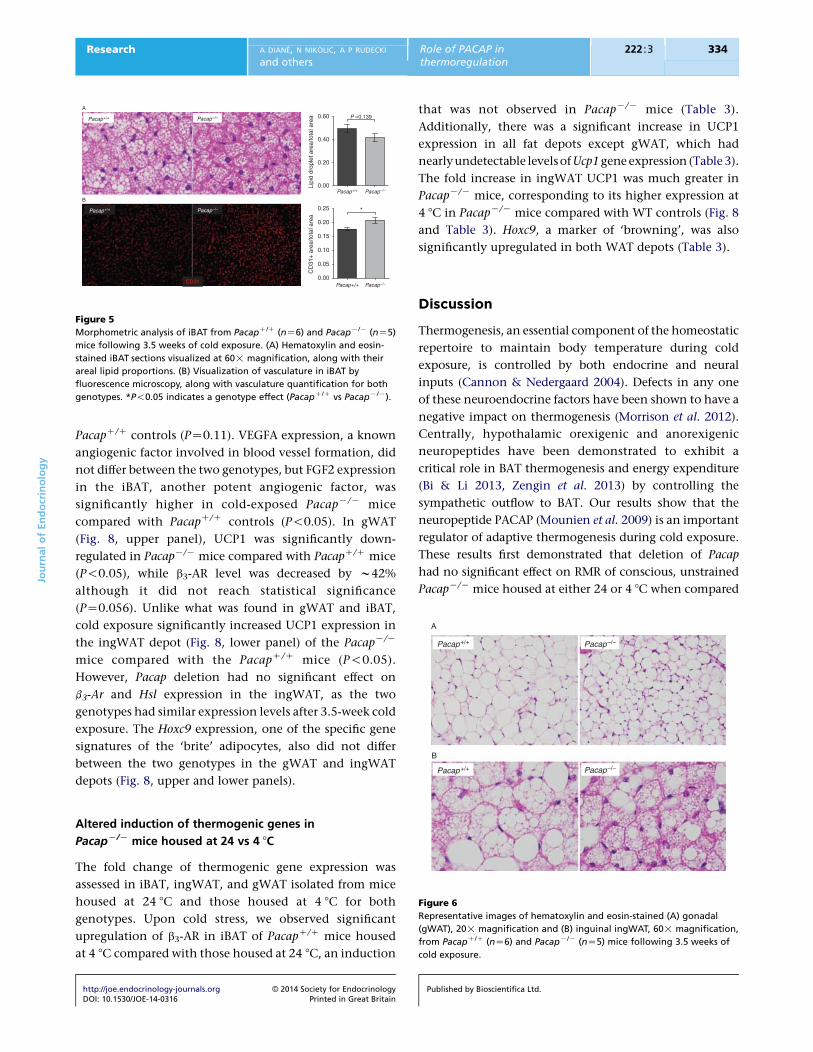
Lipid content is unaltered in iBAT of
cold-exposed PacapK/K mice
To determine if impaired BAT thermogenesis was due to,
or caused, alterations in lipid stores, intracellular lipid was
quantified in BAT as described above. Lipid droplets
appeared smaller and less prevalent in PacapK/K BAT,
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
but quantification yielded no statistical difference in lipid
content between the genotypes (Fig. 5A). Such analyses
could not be performed on ingWAT or gWAT due to
heterogeneity of sections, although the morphology
looked similar for both genotypes (Fig. 6).
Increased angiogenesis in iBAT of cold-exposed
PacapK/K mice
iBAT from cold-exposedPacapK/Kmice showed significantly
increased immunoreactivity for the angiogenic marker,
CD31, compared with iBAT from PacapC/C control mice
(P!0.05; Fig. 5B). Visually, we did not detect a difference in
CD31 immunoreactivity in PacapK/K ingWAT sections
comparedwithPacapC/C ingWATsections (datanot shown).
No change in circulating factors associated with
thermoregulation in PacapK/K mice
Assessment of a number of circulating factors in the
plasma known to affect thermogenesis, such as free fatty
acids, T4, and BNPs, showed no difference between the two
genotypes after 3.5 weeks of cold exposure (Table 2).
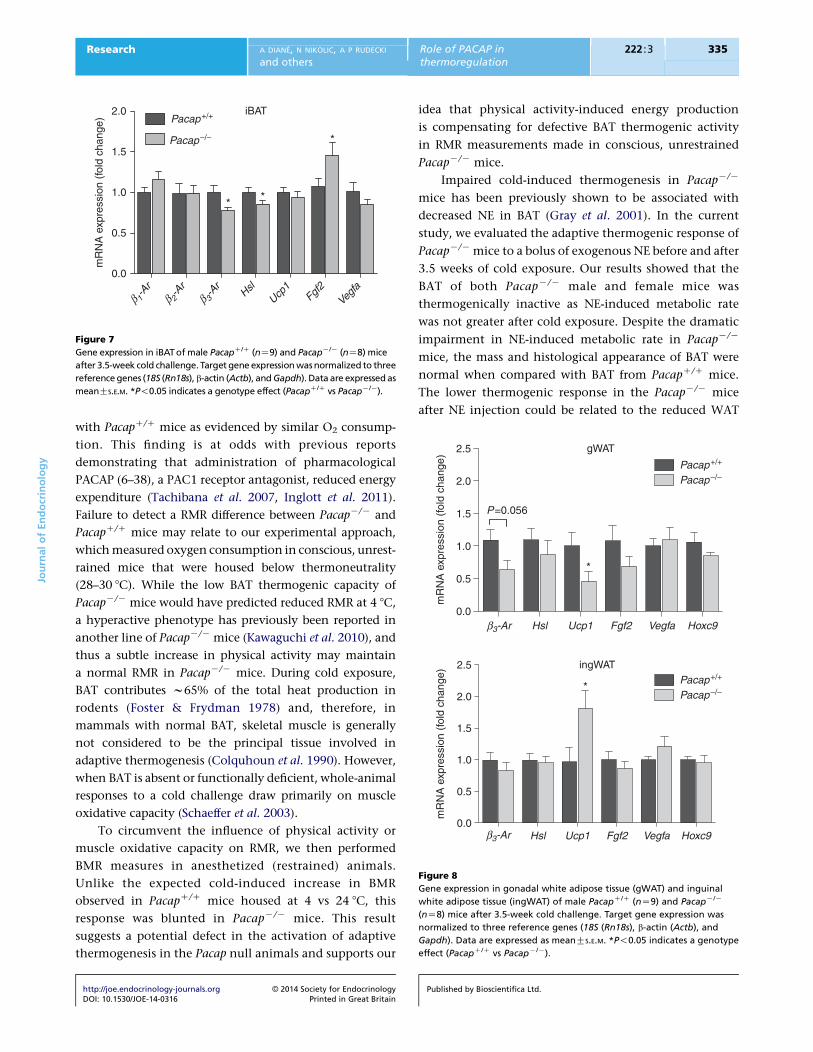
Altered expression of thermogenic genes in
cold-exposed PacapK/K mice
To evaluate the contribution of PACAP to energy
metabolism, we compared the mRNA levels of thermo-
genic genes in BATs and WATs of male PacapK/K and
PacapC/C mice. In iBAT (Fig. 7), Hsl and b3-Ar mRNA levels
were significantly decreased in PacapK/K mice compared
with their PacapC/C littermates (P!0.05). Ucp1 mRNA
expression did not differ between PacapK/K mice and
Published by Bioscientifica Ltd.
0.00
0.05
0.10
0.15
0.20
0.25 *
0.00
0.20
0.40
0.60 P =0.139Pacap+/+
A
B
Pacap–/–
Pacap+/+ Pacap–/–
Pacap+/+ Pacap–/–
Pacap–/–
CD31
Pacap+/+
Lipi
d dr
ople
t are
a/to
tal a
rea
CD
31+
are
a/to
tal a
rea
Figure 5
Morphometric analysis of iBAT from PacapC/C (nZ6) and PacapK/K (nZ5)
mice following 3.5 weeks of cold exposure. (A) Hematoxylin and eosin-
stained iBAT sections visualized at 60! magnification, along with their
areal lipid proportions. (B) Visualization of vasculature in iBAT by
fluorescence microscopy, along with vasculature quantification for both
genotypes. *P!0.05 indicates a genotype effect (PacapC/C vs PacapK/K).
Pacap+/+
A
B
Pacap–/–
Pacap+/+ Pacap–/–
JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 334
PacapC/C controls (PZ0.11). VEGFA expression, a known
angiogenic factor involved in blood vessel formation, did
not differ between the two genotypes, but FGF2 expression
in the iBAT, another potent angiogenic factor, was
significantly higher in cold-exposed PacapK/K mice
compared with PacapC/C controls (P!0.05). In gWAT
(Fig. 8, upper panel), UCP1 was significantly down-
regulated in PacapK/K mice compared with PacapC/C mice
(P!0.05), while b3-AR level was decreased by w42%
although it did not reach statistical significance
(PZ0.056). Unlike what was found in gWAT and iBAT,
cold exposure significantly increased UCP1 expression in
the ingWAT depot (Fig. 8, lower panel) of the PacapK/K
mice compared with the PacapC/C mice (P!0.05).
However, Pacap deletion had no significant effect on
b3-Ar and Hsl expression in the ingWAT, as the two
genotypes had similar expression levels after 3.5-week cold
exposure. The Hoxc9 expression, one of the specific gene
signatures of the ‘brite’ adipocytes, also did not differ
between the two genotypes in the gWAT and ingWAT
depots (Fig. 8, upper and lower panels).
Figure 6
Representative images of hematoxylin and eosin-stained (A) gonadal
(gWAT), 20! magnification and (B) inguinal ingWAT, 60! magnification,
from PacapC/C (nZ6) and PacapK/K (nZ5) mice following 3.5 weeks of
cold exposure.
Altered induction of thermogenic genes in
PacapK/K mice housed at 24 vs 4 8C
The fold change of thermogenic gene expression was
assessed in iBAT, ingWAT, and gWAT isolated from mice
housed at 24 8C and those housed at 4 8C for both
genotypes. Upon cold stress, we observed significant
upregulation of b3-AR in iBAT of PacapC/C mice housed
at 4 8C compared with those housed at 24 8C, an induction
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
that was not observed in PacapK/K mice (Table 3).
Additionally, there was a significant increase in UCP1
expression in all fat depots except gWAT, which had
nearlyundetectable levelsofUcp1gene expression (Table 3).
The fold increase in ingWAT UCP1 was much greater in
PacapK/K mice, corresponding to its higher expression at
4 8C in PacapK/K mice compared with WT controls (Fig. 8
and Table 3). Hoxc9, a marker of ‘browning’, was also
significantly upregulated in both WAT depots (Table 3).
Discussion
Thermogenesis, an essential component of the homeostatic
repertoire to maintain body temperature during cold
exposure, is controlled by both endocrine and neural
inputs (Cannon & Nedergaard 2004). Defects in any one
of these neuroendocrine factors have been shown to have a
negative impact on thermogenesis (Morrison et al. 2012).
Centrally, hypothalamic orexigenic and anorexigenic
neuropeptides have been demonstrated to exhibit a
critical role in BAT thermogenesis and energy expenditure
(Bi & Li 2013, Zengin et al. 2013) by controlling the
sympathetic outflow to BAT. Our results show that the
neuropeptide PACAP (Mounien et al. 2009) is an important
regulator of adaptive thermogenesis during cold exposure.
These results first demonstrated that deletion of Pacap
had no significant effect on RMR of conscious, unstrained
PacapK/K mice housed at either 24 or 4 8C when compared
Published by Bioscientifica Ltd.
gWAT
β3-Ar0.0
0.5
1.0
1.5
2.0
2.5
Pacap+/+
Pacap–/–
Pacap+/+
Pacap–/–
*
P=0.056
mR
NA
exp
ress
ion
(fol
d ch
ange
)
ingWAT
Hsl Ucp1 Fgf2 Vegfa Hoxc9
β3-Ar Hsl Ucp1 Fgf2 Vegfa Hoxc90.0
0.5
1.0
1.5
2.0
2.5
*
mR
NA
exp
ress
ion
(fol
d ch
ange
)
Figure 8
Gene expression in gonadal white adipose tissue (gWAT) and inguinal
white adipose tissue (ingWAT) of male PacapC/C (nZ9) and PacapK/K
(nZ8) mice after 3.5-week cold challenge. Target gene expression was
normalized to three reference genes (18S (Rn18s), b-actin (Actb), and
Gapdh). Data are expressed as meanGS.E.M. *P!0.05 indicates a genotype
effect (PacapC/C vs PacapK/K).
iBAT
β 1-A
rβ 2
-Ar
β 3-A
rHsl
Ucp1
Fgf2
Vegfa
0.0
0.5
1.0
1.5
2.0Pacap+/+
Pacap–/– *
* *
mR
NA
exp
ress
ion
(fol
d ch
ange
)
Figure 7
Gene expression in iBATof male PacapC/C (nZ9) and PacapK/K (nZ8) mice
after 3.5-week cold challenge. Target gene expressionwasnormalized to three
reference genes (18S (Rn18s), b-actin (Actb), andGapdh). Data are expressedas
meanGS.E.M. *P!0.05 indicates a genotype effect (PacapC/C vs PacapK/K).
JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 335
with PacapC/C mice as evidenced by similar O2 consump-
tion. This finding is at odds with previous reports
demonstrating that administration of pharmacological
PACAP (6–38), a PAC1 receptor antagonist, reduced energy
expenditure (Tachibana et al. 2007, Inglott et al. 2011).
Failure to detect a RMR difference between PacapK/K and
PacapC/C mice may relate to our experimental approach,
which measured oxygen consumption in conscious, unrest-
rained mice that were housed below thermoneutrality
(28–30 8C). While the low BAT thermogenic capacity of
PacapK/K mice would have predicted reduced RMR at 4 8C,
a hyperactive phenotype has previously been reported in
another line of PacapK/K mice (Kawaguchi et al. 2010), and
thus a subtle increase in physical activity may maintain
a normal RMR in PacapK/K mice. During cold exposure,
BAT contributes w65% of the total heat production in
rodents (Foster & Frydman 1978) and, therefore, in
mammals with normal BAT, skeletal muscle is generally
not considered to be the principal tissue involved in
adaptive thermogenesis (Colquhoun et al. 1990). However,
when BAT is absent or functionally deficient, whole-animal
responses to a cold challenge draw primarily on muscle
oxidative capacity (Schaeffer et al. 2003).
To circumvent the influence of physical activity or
muscle oxidative capacity on RMR, we then performed
BMR measures in anesthetized (restrained) animals.
Unlike the expected cold-induced increase in BMR
observed in PacapC/C mice housed at 4 vs 24 8C, this
response was blunted in PacapK/K mice. This result
suggests a potential defect in the activation of adaptive
thermogenesis in the Pacap null animals and supports our
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
idea that physical activity-induced energy production
is compensating for defective BAT thermogenic activity
in RMR measurements made in conscious, unrestrained
PacapK/K mice.
Impaired cold-induced thermogenesis in PacapK/K
mice has been previously shown to be associated with
decreased NE in BAT (Gray et al. 2001). In the current
study, we evaluated the adaptive thermogenic response of
PacapK/K mice to a bolus of exogenous NE before and after
3.5 weeks of cold exposure. Our results showed that the
BAT of both PacapK/K male and female mice was
thermogenically inactive as NE-induced metabolic rate
was not greater after cold exposure. Despite the dramatic
impairment in NE-induced metabolic rate in PacapK/K
mice, the mass and histological appearance of BAT were
normal when compared with BAT from PacapC/C mice.
The lower thermogenic response in the PacapK/K mice
after NE injection could be related to the reduced WAT
Published by Bioscientifica Ltd.

Table 3 Fold change in iBAT, ingWAT, and gWAT gene expression from 24 8C housed to 4 8C housed PacapC/C and PacapK/K mice
(PacapC/C, 24 8C (nZ5); PacapK/K, 24 8C (nZ5); PacapC/C, 4 8C (nZ9); and PacapK/K, 4 8C (nZ8)). mRNA expression data were
normalized to reference genes that remained stable across treatments and genotypes for BAT (b-actin (Actb)), ingWAT (18S (Rn18s),
b-actin, and Gapdh), and gWAT (Tbp and b-actin)
Genes
iBAT ingWAT gWAT
PacapC/C PacapK/K PacapC/C PacapK/K PacapC/C PacapK/K
b1-Ar 0.95 1.02 NA NA NA NAb2-Ar 0.60 0.87 NA NA NA NAb3-Ar 3.34** 1.43 1.53 0.70 0.22*** 0.25*Hsl 1.57 1.38 1.07 0.68 0.19*** 0.22*Ucp1 2.83** 2.37** 169.65** 755.54*** 1.99 2.29Fgf2 6.48** 6.31** 0.54* 0.29*** 0.14*** 0.15**Vegfa 1.63** 1.45 0.88 0.66* 0.35*** 0.43*Hoxc9 NA NA 3.35*** 2.16** 3.68** 5.04**
Significant differences betweenmRNA expression in 24 and 4 8C samples for each genotype are denoted by *P!0.05, **P!0.0, or ***P!0.001. NA, gene notmeasured.
JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 336
found in these mice after cold exposure as a similar
thermogenic pattern has been previously reported in
A-ZIP/F-1 lipodystrophic mice (Gavrilova et al. 2000).
Differential use of physiological fuels (carbohydrates,
fat, and protein) is another strategy to adapt and survive
during challenge of low environmental temperature
(Doubt 1991, Schaeffer et al. 2003). Following 3.5 weeks of
cold exposure (4 8C), RER increased from 0.7 to 0.9 in
PacapK/K mice, but not PacapC/C mice, after NE injection,
indicating a preferential use of carbohydrates to produce
energy by the Pacap null mice. This switch from fat to
carbohydrate utilization in cold-exposed PacapK/K mice
could indicate the impairment of mechanisms of lipid
mobilization from adipose tissue during cold exposure
(Doubt 1991) or a physiological adaptation to withstand
the high energy demands associated with NE infusion, as
oxidation of carbohydrates produces more energy per mole
of oxygen than fat oxidation (Virtue & Vidal-Puig 2013).
The capacity for NE-induced thermogenesis in BAT will
be influenced by the level of b3-AR available for NE binding
on brown adipocytes to regulate the expression and activity
of HSL and UCP1 (Cannon & Nedergaard 2004). Gene
expression data revealed a significant reduction in b3-Ar
mRNA in PacapK/K BAT compared with PacapC/C BAT
isolated from cold-exposed mice. Comparison of expression
levels of b3-Ar mRNA in mice housed at 24 vs 4 8C revealed
that b3-Ar mRNA is not induced in the BAT of PacapK/K
mice in response to cold, unlike WT mice where it is
induced significantly. The failure to observe increased O2
consumption after NE injection may be associated with an
inability to upregulate b3-Ar mRNA in PacapK/K BAT. The
fundamental role of b3-AR in BAT thermogenic machinery
is well substantiated (Tachibana et al. 2003, Ueta et al. 2012)
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
and these results demonstrate for the first time that PACAP
plays an important role in regulating the expression of
b3-AR during cold exposure. PACAP deficiency has been
previously shown to reduce endogenous levels of NE in
BAT. The tonic suppression of NE in PacapK/K BAT may
suppress b3-AR expression in BAT, contributing to the
impaired NE-induced metabolic rate in cold-exposed Pacap
null mice. Subsequent studies either measuring the b3-AR
protein levels from the BAT of the two genotypes after cold
exposure or using b3-AR agonists to stimulate metabolic
rate should be performed to support this finding.
Despite the lower b3-AR expression within the
canonical BAT, the adult male PacapK/K mice survived a
3.5-week period of cold exposure and were able to induce
Ucp1 gene expression in BAT. This suggests that other
mechanisms to activate the thermogenic machinery,
independent of ARs, also exist. In addition to increased
physical activity, this may include the recruitment of ‘brite’
adipocytes into WAT depots as evidenced by the higher
induction of Ucp1 gene expression in ingWAT of PacapK/K
mice compared with PacapC/C animals. The disparate
expression of thermogenic genes in iBAT and ingWAT
depots of PacapC/C and PacapK/K mice suggests that PACAP
may differentially regulate the central innervation of these
depots. Future studies using retrograde viral transneuronal
tract tracers to label neuronal circuits that originate in the
hypothalamus and terminate in brown and WAT depots
will help to clarify as to how PACAP mediates these effects
in different adipose tissue depots.
In addition to sympathetic innervation of brown fat,
thermoregulation of BAT is made possible by extensive
vascularization enabling rapid access of circulating metab-
olites to the brown adipocytes (Asano et al. 1999). As basic
Published by Bioscientifica Ltd.

JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 337
FGF (or FGF2) is shown to induce mitogenic activity in
endothelial cells in vitro and angiogenesis in vivo
(Montesano et al. 1986, Gualandris et al. 1996), it is quite
likely that FGF2 is involved in the blood vessel formation
associated with the cold-induced BAT growth. Our results
showed that mRNA expression of Fgf2, as well as CD31
immunoreactivity, was higher in BAT of cold-exposed
PacapK/K mice compared with PacapC/C mice. This
heightened angiogenic induction in BAT of PacapK/K
mice, despite their low thermogenic capacity, may indicate
a physiological adaptive response that attempts to increase
the supply of nutrients and oxygen to boost thermogenesis.
This adaptation, along with the above-mentioned
mechanisms might be an attempt to activate thermogenic
machinery independent of ARs as a compensatory
approach for survival during chronic cold exposure.
In conclusion, our data show for the first time that
exogenous NE administration cannot rescue the impaired
adaptive thermogenesis in PacapK/K mice and the reduced
induction of b3-AR expression in BAT in response to cold
may contribute to the impaired sympathetic-induced
adaptive thermogenesis in these mice. These results
contribute to our understanding of the basic mechanisms
regulating energy metabolism, demonstrating PACAP as
an important neuroendocrine mediator of the sym-
pathetic regulation of adaptive thermogenesis. The
PacapK/K mouse is thus a suitable model for further
studies investigating the molecular and neuroendocrine
mechanisms involved in thermoregulation.
Declaration of interest
The authors declare that there is no conflict of interest that could be
perceived as prejudicing the impartiality of the research reported.
Funding
This study was funded by a grant from the Natural Sciences and
Engineering Research Council of Canada (NSERC) to Dr S L G.
Acknowledgements
The authors thank Lydia Troc, Dee Jones, and K-Lynn Hogh for their
exceptional technical assistance and dedication to the care and mainten-
ance of our PACAP null mouse colony.
References
Asano A, Kimura K & Saito M 1999 Cold-induced mRNA expression of
angiogenic factors in rat brown adipose tissue. Journal of Veterinary
Medical Science 61 403–409. (doi:10.1292/jvms.61.403)
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
Baron DM, Clerte M, Brouckaert P, Raher MJ, Flynn AW, Zhang H, Carter EA,
Picard MH, Bloch KD, Buys ES et al. 2012 In vivo noninvasive
characterization of brown adipose tissue blood flow by contrast
ultrasound in mice. Circulation. Cardiovascular Imaging 5 652–659.
(doi:10.1161/CIRCIMAGING.112.975607)
Bi S & Li L 2013 Browning of white adipose tissue: role of hypothalamic
signaling. Annals of the New York Academy of Sciences 1302 30–34.
(doi:10.1111/nyas.12258)
Bu LH & Lephart ED 2005 Effects of dietary phytoestrogens on core body
temperature during the estrous cycle and pregnancy. Brain Research
Bulletin 65 219–223. (doi:10.1016/j.brainresbull.2005.01.008)
Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R,
Nolan T, Pfaffl MW, Shipley GL et al. 2009 The MIQE guidelines:
minimum information for publication of quantitative real-time PCR
experiments. Clinical Chemistry 55 611–622. (doi:10.1373/clinchem.
2008.112797)
Cannon B & Nedergaard J 2004 Brown adipose tissue: function and
physiological significance. Physiological Reviews 84 277–359.
(doi:10.1152/physrev.00015.2003)
Castorina A, Giunta S, Mazzone V, Cardile V & D’Agata V 2010 Effects of
PACAP and VIP on hyperglycemia-induced proliferation in murine
microvascular endothelial cells. Peptides 31 2276–2283. (doi:10.1016/j.
peptides.2010.08.013)
Clapham JC & Arch JR 2011 Targeting thermogenesis and related pathways
in anti-obesity drug discovery. Pharmacology & Therapeutics 131
295–308. (doi:10.1016/j.pharmthera.2011.04.004)
Colquhoun EQ, Hettiarachchi M, Ye JM, Rattigan S & Clark MG 1990
Inhibition by vasodilators of noradrenaline and vasoconstrictor-mediated,
butnot skeletalmuscle contraction-inducedoxygen uptake inthe perfused
rat hindlimb; implications for non-shivering thermogenesis in muscle
tissue. General Pharmacology 21 141–148.
Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC,
Palmer EL, Tseng YH, Doria A et al. 2009 Identification and importance
of brown adipose tissue in adult humans. New England Journal of
Medicine 360 1509–1517. (doi:10.1056/NEJMoa0810780)
Doubt TJ 1991 Physiology of exercise in the cold. Sports Medicine 1 367–381.
(doi:10.2165/00007256-199111060-00003)
Dubois-Ferriere R & Chinet AE 1981 Contribution of skeletal muscle to the
regulatory non-shivering thermogenesis in small mammals. Pflugers
Archiv: European Journal of Physiology 390 224–229. (doi:10.1007/
BF00658266)
Enerback S, Jacobsson A, Simpson EM, Guerra C, Yamashita H, Harper ME
& Kozak LP 1997 Mice lacking mitochondrial uncoupling protein are
cold-sensitive but not obese. Nature 387 90–94. (doi:10.1038/
387090a0)
Foster DO & Frydman ML 1978 Brown adipose tissue: the dominant site of
nonshivering thermogenesis in the rat. Experientia. Supplementum 32
147–151.
Gavrilova O, Marcus-Samuels B & Reitman ML 2000 Lack of responses to a
b3-adrenergic agonist in lipoatrophic A-ZIP/F-1 mice. Diabetes 49
1910–1916. (doi:10.2337/diabetes.49.11.1910)
Ghorbani M, Claus TH & Himms-Hagen J 1997 Hypertrophy of brown
adipocytes in brown and white adipose tissues and reversal of
diet-induced obesity in rats treated with a b3-adrenoceptor agonist.
Biochemical Pharmacology 54 121–131. (doi:10.1016/S0006-
2952(97)00162-7)
Golozoubova V, Gullberg H, Matthias A, Cannon B, Vennstrom B &
Nedergaard J 2004 Depressed thermogenesis but competent brown
adipose tissue recruitment in mice devoid of all hormone-binding
thyroid hormone receptors. Molecular Endocrinology 18 384–401.
(doi:10.1210/me.2003-0267)
Golozoubova V, Cannon B & Nedergaard J 2006 UCP1 is essential for
adaptive adrenergic nonshivering thermogenesis. American Journal of
Physiology. Endocrinology and Metabolism 291 E350–E357. (doi:10.1152/
ajpendo.00387.2005)
Published by Bioscientifica Ltd.

JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 338
Gray SL, Cummings KJ, Jirik FR & Sherwood NM 2001 Targeted disruption
of the pituitary adenylate cyclase-activating polypeptide gene results in
early postnatal death associated with dysfunction of lipid and
carbohydrate metabolism. Molecular Endocrinology 15 1739–1747.
(doi:10.1210/mend.15.10.0705)
Gray SL, Yamaguchi N, Vencova P & Sherwood NM 2002 Temperature-
sensitive phenotype in mice lacking pituitary adenylate cyclase-
activating polypeptide. Endocrinology 143 3946–3954. (doi:10.1210/en.
2002-220401)
Gualandris A, Rusnati M, Belleri M, Nelli EE, Bastaki M, Molinari-Tosatti MP,
Bonardi F, Parolini S, Albini A, Morbidelli L et al. 1996 Basic fibroblast
growth factor overexpression in endothelial cells: an autocrine
mechanism for angiogenesis and angioproliferative diseases. Cell
Growth & Differentiation 7 147–160.
Guerra C, Koza RA, Yamashita H, Walsh K & Kozak LP 1998 Emergence of
brown adipocytes in white fat in mice is under genetic control. Effects
on body weight and adiposity. Journal of Clinical Investigation 102
412–420. (doi:10.1172/JCI3155)
Hondares E, Rosell M, Gonzalez FJ, Giralt M, Iglesias R & Villarroya F 2010
Hepatic FGF21 expression is induced at birth via PPARa in response
to milk intake and contributes to thermogenic activation of neonatal
brown fat. Cell Metabolism 11 206–212. (doi:10.1016/j.cmet.
2010.02.001)
Inglott MA, Farnham MM & Pilowsky PM 2011 Intrathecal PACAP-38
causes prolonged widespread sympathoexcitation via a spinally
mediated mechanism and increases in basal metabolic rate in
anesthetized rat. American Journal of Physiology. Heart and Circulatory
Physiology 300 H2300–H2307.
Inokuma K, Okamatsu-Ogura Y, Omachi A, Matsushita Y, Kimura K,
Yamashita H & Saito M 2006 Indispensable role of mitochondrial UCP1
for antiobesity effect of b3-adrenergic stimulation. American Journal of
Physiology. Endocrinology and Metabolism 290 E1014–E1021. (doi:10.
1152/ajpendo.00105.2005)
Kawaguchi C, Isojima Y, Shintani N, Hatanaka M, Guo X, Okumura N,
Nagai K, Hashimoto H & Baba A 2010 PACAP-deficient mice exhibit
light parameter-dependent abnormalities on nonvisual photoreception
and early activity onset. PLoS ONE 5 e9286. (doi:10.1371/journal.pone.
0009286)
Kohno D & Yada T 2012 Arcuate NPY neurons sense and integrate
peripheral metabolic signals to control feeding. Neuropeptides 46
315–319. (doi:10.1016/j.npep.2012.09.004)
Lopez M, Varela L, Vazquez MJ, Rodrıguez-Cuenca S, Gonzalez CR,
Velagapudi VR, Morgan DA, Schoenmakers E, Agassandian K, Lage R
et al. 2010 Hypothalamic AMPK and fatty acid metabolism mediate
thyroid regulation of energy balance. Nature Medicine 16 1001–1008.
(doi:10.1038/nm.2207)
Lowell BB & Spiegelman BM 2000 Towards a molecular understanding of
adaptive thermogenesis. Nature 404 652–660.
Lowell BB, S-Susulic V, Hamann A, Lawitts JA, Himms-Hagen J, Boyer BB,
Kozak LP & Flier JS 1993 Development of obesity in transgenic mice
after genetic ablation of brown adipose tissue. Nature 366 740–742.
(doi:10.1038/366740a0)
Matthias A, Ohlson KB, Fredriksson JM, Jacobsson A, Nedergaard J &
Cannon B 2000 Thermogenic responses in brown fat cells are fully
UCP1-dependent. UCP2 or UCP3 do not substitute for UCP1 in
adrenergically or fatty acid-induced thermogenesis. Journal of Biological
Chemistry 275 25073–25081. (doi:10.1074/jbc.M000547200)
Miyata A, Arimura A, Dahl RR, Minamino N, Uehara A, Jiang L, Culler MD
& Coy DH 1989 Isolation of a novel 38 residue-hypothalamic
polypeptide which stimulates adenylate cyclase in pituitary cells.
Biochemical and Biophysical Research Communications 164 567–574.
(doi:10.1016/0006-291X(89)91757-9)
Montesano R, Vassalli JD, Baird A, Guillemin R & Orci L 1986 Basic
fibroblast growth factor induces angiogenesis in vitro. PNAS 83
7297–7301. (doi:10.1073/pnas.83.19.7297)
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
Morrison SF, Madden CJ & Tupone D 2012 Central control of brown
adipose tissue thermogenesis. Frontiers in Endocrinology 3 pii: 00005.
(doi:10.3389/fendo.2012.00005)
Mounien L, Do Rego JC, Bizet P, Boutelet I, Gourcerol G, Fournier A, Brabet P,
Costentin J, Vaudry H & Jegou S 2009 Pituitary adenylate cyclase-
activating polypeptide inhibits food intake in mice through activation
of the hypothalamic melanocortin system. Neuropsychopharmacology 34
424–435. (doi:10.1038/npp.2008.73)
Ohno H, Shinoda K, Spiegelman BM & Kajimura S 2012 PPARg agonists
induce a white-to-brown fat conversion through stabilization of
PRDM16 protein. Cell Metabolism 15 395–404. (doi:10.1016/j.cmet.
2012.01.019)
Ouellet V, Routhier-Labadie A, Bellemare W, Lakhal-Chaieb L, Turcotte E,
Carpentier AC & Richard D 2011 Outdoor temperature, age, sex, body
mass index, and diabetic status determine the prevalence, mass, and
glucose-uptake activity of 18F-FDG-detected BAT in humans. Journal of
Clinical Endocrinology and Metabolism 96 192–199. (doi:10.1210/jc.
2010-0989)
Perkins MN, Rothwell NJ, Stock MJ & Stone TW 1981 Activation of brown
adipose tissue thermogenesis by the ventromedial hypothalamus.
Nature 289 401–402. (doi:10.1038/289401a0)
Resch JM, Maunze B, Gerhardt AK, Magnuson SK, Phillips KA & Choi S
2013 Intrahypothalamic pituitary adenylate cyclase-activating poly-
peptide regulates energy balance via site-specific actions on feeding and
metabolism. American Journal of Physiology. Endocrinology and
Metabolism 305 E1452–E1463. (doi:10.1152/ajpendo.00293.2013)
Richard D & Picard F 2011 Brown fat biology and thermogenesis. Frontiers
in Bioscience 16 1233–1260. (doi:10.2741/3786)
Ricquier D, Miroux B, Larose M, Cassard-Doulcier AM & Bouillaud F 2000
Endocrine regulation of uncoupling proteins and energy expenditure.
International Journal of Obesity and Related Metabolic Disorders 24
S86–S88. (doi:10.1038/sj.ijo.0801286)
Riedel D 2010 Single molecule manipulation at low temperature and laser
scanning tunnelling photo-induced processes analysis through time-
resolved studies. Journal of Physics. Condensed Matter 22 264009.
(doi:10.1088/0953-8984/22/26/264009)
Schaeffer PJ, Villarin JJ & Lindstedt SL 2003 Chronic cold exposure
increases skeletal muscle oxidative structure and function in
Monodelphis domestica, a marsupial lacking brown adipose tissue.
Physiological and Biochemical Zoology 76 877–887. (doi:10.1086/378916)
Schulz TJ & Tseng YH 2013 Systemic control of brown fat thermogenesis:
integration of peripheral and central signals. Annals of the New York
Academy of Sciences 1302 35–41. (doi:10.1111/nyas.12277)
Segal JP, Stallings NR, Lee CE, Liping Z, Socci N, Viale A, Harris TM,
Soares MB, Childs G, Elmquist JK et al. 2010 Use of laser-capture
microdissection for the identification of marker genes for the
ventromedial hypothalamic nucleus. Journal of Neuroscience 25
4181–4188. (doi:10.1523/JNEUROSCI.0158-05.2005)
Tachibana T, Takagi T, Saito ES, Tomonaga S, Zhang R, Koga Y, Kido Y,
Michael Denbow D & Furuse M 2003 b3-Adrenergic receptor is involved
in feeding regulation in chicks. Comparative Biochemistry and Physiology.
Part A, Molecular & Integrative Physiology 135 403–409.
Tachibana T, Oikawa D, Adachi N, Boswell T & Furuse M 2007 Central
administration of vasoactive intestinal peptide and pituitary adenylate
cyclase-activating polypeptide differentially regulates energy metab-
olism in chicks. Comparative Biochemistry and Physiology. Part A,
Molecular & Integrative Physiology 147 156–164.
Thomas SA & Palmiter RD 1997 Thermoregulatory and metabolic
phenotypes of mice lacking noradrenaline and adrenaline. Nature 387
94–97. (doi:10.1038/387094a0)
Tseng YH, Kokkotou E, Schulz TJ, Huang TL, Winnay JN, Taniguchi CM,
Tran TT, Suzuki R, Espinoza DO, Yamamoto Y et al. 2008 New role of
bone morphogenetic protein 7 in brown adipogenesis and energy
expenditure. Nature 454 1000–1004. (doi:10.1038/nature07221)
Uchida Y, Kano M, Yasuhara S, Kobayashi A, Tokizawa K & Nagashima K
2010 Estrogen modulates central and peripheral responses to cold in
Published by Bioscientifica Ltd.

JournalofEndocrinology
Research A DIANE, N NIKOLIC, A P RUDECKI
and othersRole of PACAP inthermoregulation
222 :3 339
female rats. Journal of Physiological Sciences 60 151–160. (doi:10.1007/
s12576-009-0079-x)
Ueta CB, Fernandes GW, Capelo LP, Fonseca TL, Maculan FD, Gouveia CH,
Brum PC, Christoffolete MA, Aoki MS, Lancellotti CL et al. 2012 b(1)
Adrenergic receptor is key to cold- and diet-induced thermogenesis in
mice. Journal of Endocrinology 214 359–365. (doi:10.1530/JOE-12-0155)
Vaudry D, Falluel-Morel A, Bourgault S, Basille M, Burel D, Wurtz O,
Fournier A, Chow BK, Hashimoto H, Galas L et al. 2009 Pituitary
adenylate cyclase-activating polypeptide and its receptors: 20 years
after the discovery. Pharmacological Reviews 61 283–357. (doi:10.1124/
pr.109.001370)
Vaughan CH, Zarebidaki E, Ehlen JC & Bartness TJ 2014 Analysis and
measurement of the sympathetic and sensory innervation of white and
brown adipose tissue. Methods in Enzymology 537 199–225.
Virtue S & Vidal-Puig A 2013 Assessment of brown adipose tissue function.
Frontiers in Physiology 4 128–156. (doi:10.3389/fphys.2013.00128)
Walden TB, Hansen IR, Timmons JA, Cannon B & Nedergaard J 2012
Recruited vs. nonrecruited molecular signatures of brown, “brite”, and
http://joe.endocrinology-journals.org � 2014 Society for EndocrinologyDOI: 10.1530/JOE-14-0316 Printed in Great Britain
white adipose tissues. American Journal of Physiology. Endocrinology and
Metabolism 302 E19–E31. (doi:10.1152/ajpendo.00249.2011)
Whittle AJ, Carobbio S, Martins L, Slawik M, Hondares E, Vazquez MJ,
Morgan D, Csikasz RI, Gallego R, Rodriguez-Cuenca S et al. 2012 BMP8B
increases brown adipose tissue thermogenesis through both central and
peripheral actions. Cell 149 871–885. (doi:10.1016/j.cell.2012.02.066)
Wu J, Bostrom P, Sparks LM, Ye L, Choi JH, Giang AH, Khandekar M,
Virtanen KA, Nuutila P, Schaart G et al. 2012 Beige adipocytes are a
distinct type of thermogenic fat cell in mouse and human. Cell 150
366–376. (doi:10.1016/j.cell.2012.05.016)
Xue Y, Petrovic N, Cao R, Larsson O, Lim S, Chen S, Feldmann HM, Liang Z,
Zhu Z, Nedergaard J et al. 2009 Hypoxia-independent angiogenesis in
adipose tissues during cold acclimation. Cell Metabolism 9 99–109.
(doi:10.1016/j.cmet.2008.11.009)
Zengin A, Nguyen AD, Wong IP, Zhang L, Enriquez RF, Eisman JA, Herzog H,
Baldock PA & Sainsbury A 2013 Neuropeptide Y mediates the short-
term hypometabolic effect of estrogen deficiency in mice. International
Journal of Obesity 37 390–398. (doi:10.1038/ijo.2012.71)
Received in final form 11 July 2014Accepted 22 July 2014Accepted Preprint published online 23 July 2014
Published by Bioscientifica Ltd.





























