Embed Size (px)
Citation preview

P2X3 Receptor Localizes Into Lipid Rafts inNeuronal Cells
Fabrizio Vacca,1,2 Susanna Amadio,1,3 Giuseppe Sancesario,3 Giorgio Bernardi,1,3 andCinzia Volonte1,4*1Fondazione Santa Lucia, Rome, Italy2University of Rome “La Sapienza,” Department of Human Physiology and Pharmacology, Rome, Italy3University of Rome Tor Vergata, Department of Neuroscience, Rome, Italy4C.N.R. Institute of Neurobiology and Molecular Medicine, Rome, Italy
P2X receptors are a family of seven (P2X1–7) cation chan-nels gated by extracellular ATP, widely expressed inneurons and nonneuronal cells. Lipid rafts arecholesterol/sphingolipid-rich membrane domains, in-volved in many cellular processes, including transmem-brane receptor signaling, vesicle traffic, and protein sort-ing. We provide direct biochemical evidence that P2X3receptor localizes into lipid rafts, in primary cultures ofcerebellar granule neurons as well as in brain and dorsalroot ganglia extracts. We show that P2X3 exhibits all thecharacteristics distinctive of a protein associated withlipid rafts. These characteristics include resistance todetergent extraction at 4°C, solubility after extraction ofcholesterol from membranes with either saponin ormethyl-�-cyclodextrin, and partitioning to low buoyantdensity fractions after sucrose gradient centrifugation inboth detergent-containing and detergent-free condi-tions. Furthermore, P2X3 localizes in raft-containing frac-tions in transiently transfected SH-SY5Y neuroblastomacells. The present finding contributes to the character-ization of the functional localization of P2X3 in neuronsand provides a novel potential mechanism for correcttargeting and dynamic activation of this receptor.© 2004 Wiley-Liss, Inc.
Key words: detergent-insoluble proteins; P2 receptor;cerebellar granule neurons; dorsal root ganglia
P2X receptors are a family of seven (P2X1–7) cationchannels gated by extracellular ATP. They are expressedin many excitable and nonexcitable cells and mediate avariety of physiologic actions, including smooth musclecontractility, neuroendocrine secretion, and synaptictransmission (Ralevic and Burnstock, 1998). They arepresent in the central nervous system (CNS) in both pre-and postsynaptic terminals, thereby modulating neuro-transmitter release and fast excitatory currents (Robertsonet al., 2001). Among them, the P2X3 subunit is expressedin cell bodies as well as in peripheral and central terminalsof sensory neurons in dorsal root ganglia (DRG) (Vulch-anova et al., 1997; Llewellyn-Smith and Burnstock, 1998),where it carries out a well-defined role in pain perception
(Cook et al., 1997; Cockayne et al., 2000; Souslova et al.,2000). It is expressed also in various discrete regions duringrat brain development (Kidd et al., 1998). Although P2Xreceptors are finely targeted in specific synaptic sites inboth CNS and peripheral nervous system (PNS) neurons,they do not have any obvious consensus sequence forinteracting with known scaffolding proteins and the mo-lecular basis of their localization remains to be determined.
In addition to the well-established role of scaffoldingproteins (Garner et al., 2000), it has been proposed re-cently that the association of receptors with particulardomains of the lipid bilayer, called lipid rafts, can allowspecific localization of membrane proteins. These domainsconstitute an ordered phase of the lipid bilayer (liquid-ordered phase), are highly enriched in sphingolipids andcholesterol and are characterized biochemically by resis-tance to nonionic detergent extraction and by low buoy-ant density in sucrose gradient centrifugation (Simons andIkonen, 1997; Brown and London, 1998; Brown andLondon, 2000). Lipid rafts are enriched in lipid secondmessengers and in specific membrane proteins, particularlythose containing a glycosylphosphatidylinositol anchor, orsignaling molecules, including transmembrane receptors,Src family kinases, and heterotrimeric G proteins (Smart etal., 1999). Growing evidences indicate that rafts are crucialfor many functions in neurons, including neurotrophicfactor signaling, adhesion, axon guidance, vesicular traf-ficking and protein sorting (Tsui-Pierchala et al., 2002).The insertion of proteins into sphingolipid/cholesterol-rich domains at the level of the Golgi apparatus (Gkanti-ragas et al., 2001) is an appealing mechanism for proteintargeting to specific membrane compartments in neurons.Indeed, lipid rafts have been suggested to play an impor-
Contract grant sponsor: Italian Health Ministry; Contract grant number:RA00.86V; Contract grant sponsor: Cofinanziamento MURST 2002.
*Correspondence to: Cinzia Volonte, Fondazione Santa Lucia, ViaArdeatina 354, 00179 Rome, Italy. E-mail: [email protected]
Received 19 August 2003; Revised 29 December 2003; Accepted 13January 2004
Published online 20 April 2004 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jnr.20069
Journal of Neuroscience Research 76:653–661 (2004)
© 2004 Wiley-Liss, Inc.

tant role in axonal versus dendritic sorting in hippocampalneurons (Ledesma et al., 1998, 1999). Lipid rafts are alsoessential for the maintenance of �7 nicotinic acetylcholinereceptor clusters in somatic spines, in ciliary ganglionneurons (Bruses et al., 2001) and for the synaptic stabilityof the AMPA-type glutamate receptors in hippocampalneurons (Hering et al., 2003). Moreover, it has beenobserved that sorting and targeting of the muscular nico-tinic acetylcholine receptor, together with its binding pro-tein rapsyn, is mediated by raft association (Marchand etal., 2002). Additional neurotransmitter receptors and ionchannels are associated with lipid rafts, including the�-aminobutyric acid (GABA) receptor B (Becher et al.,2001), the voltage-gated potassium channels Kv2.1 (Mar-tens et al., 2000) and Kv1.5 (Martens et al., 2001), and theG protein-activated inwardly rectifying (kir3.1/3.2) potas-sium channel (Delling et al., 2002).
We analyze the association of several P2X familymembers with lipid rafts and consider the hypothesis thatlipid rafts could contribute to the specific subcellular tar-geting and functional activation of P2X receptors. As acellular model system, we utilize mainly primary culturesof cerebellar granule neurons (CGNs), a cell type suitablefor biochemical analyses, as they are highly homogeneousand available in large quantities. Furthermore, they expressfunctional P2X and P2Y receptors in CGNs active duringneuronal cell death (Volonte and Merlo, 1996; Merlo andVolonte, 1996; Amadio et al., 2002; Cavaliere et al., 2002,2003, 2004). We found that P2X3 was the only subtypeassociated with lipid rafts among the P2 receptors ana-lyzed. To evaluate whether the localization of P2X3 wasexclusive to cultured CGNs, we confirmed our results inyoung rat brain, cerebellum, and dorsal root ganglia(DRG). Moreover, we utilized SH-SY5Y neuroblastomacells transiently transfected with the P2X3 receptor toevaluate whether lipid raft association was strictly depen-dent on the cell context or is indeed an intrinsic molecularproperty of the protein.
MATERIALS AND METHODSAntibodies and Reagents
P2X3 rabbit polyclonal antibody was purchased fromNeuromics (Northfield, MN). P2X1, P2X2, P2X4, P2X7, andP2Y4 antibodies were purchased from Alomone Labs (Jerusa-lem, Israel). Caveolin rabbit polyclonal and flotillin-2 mousemonoclonal antibodies were obtained from BD Biosciences (SanJose, CA) and �-actin and anti-glial fibrillary acidic protein(GFAP) antibodies were from Sigma-Aldrich (Milano, Italy). Allother reagents, unless indicated, were purchased from Sigma-Aldrich.
Treatment of Experimental Animals
All animal handling and experimental procedures werecarried out in accordance with guidelines published in the NIHGuide for the Care and Use of Laboratory, the principles pre-sented in the “Guidelines for the Use of Animals in Neuro-science Research” by the Society for Neuroscience, and Italianlegislation on animal experimentation (Decreto legislativo 116/92).
Cell Cultures and Transfections
Primary cell cultures CGNs were prepared from 8-day-old Wistar rats as described previously (Levi et al., 1989) andseeded on dishes coated with poly-L-lysine, in Eagle’s basalmedium (BME; Sigma), supplemented with 25 mM KCl, 2 mML-glutamine, 0.1 M gentamycin, and 10% heat-inactivated calfserum (Invitrogen, San Giuliano Milanese, Italy). To preventglial cell proliferation, cultures were supplemented with 10 �Mcytosine arabinoside 18–24 hr after plating and kept for 7 days,without replacing the culture medium.
The rat pheochromocytoma PC12 cell line was culturedon collagen-coated dishes in RPMI 1640 medium supple-mented with 10% heat-inactivated horse serum (HS) and 5%fetal calf serum (FCS).
The human neuroblastoma cell line SH-SY5Y was cul-tured in a 1:1 mixture of Dulbecco’s modified Eagle mediumand nutrient mixture F-12 (D-MEM/F-12), supplemented with2 mM L-glutamine, 100 U/ml penicillin, 100 �g/ml strepto-mycin and 10% heat inactivated calf serum (Invitrogen). Cellswere transfected with full-length cDNA coding for rat P2X3receptor inserted in the pcDNA3 expressing vector (kindlyprovided by A. North, Sheffield, UK); control cells were trans-fected with the empty pcDNA3 vector (Invitrogen). Cells weretransfected with lipofectamine 2000 reagent (Invitrogen) ac-cording to the manufacturer’s instructions. Briefly, cells weretransfected at 80–90% confluence in 90-mm wells with 30 �llipofectamine 2000 and 15 �g of plasmid DNA. DNA-lipofectamine 2000-containing medium was replaced after 5 hrand experiments for raft isolation were carried out approxi-mately 30 hr later. Transfection efficiency was determined inparallel wells transfected with green fluorescent protein (GFP)inserted in the pcDNA3 vector and generally ranged from50–80%.
Protein Solubilization
CGNs (1 � 105 cells) at 7–8 days in vitro (DIV) werewashed twice with phosphate-buffered saline (PBS) alone, de-tached from dishes with the same buffer containing 2 mMEDTA, and pelleted. Cells were then resuspended in 50 �l ofPBS, with or without 0.5% saponin or 50 mM methyl-�-cyclodextrin. After 30 min at room temperature, cells werepelleted at 6,500 x g for 5 min and 100 �l of ice-cold lysis buffer(10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% Triton X-100,2 mM EDTA, 1 mM phenylmethylsulfonyl fluoride [PMSF],and 20 �M leupeptin) was added to the samples. After anadditional 30 min on ice, samples were centrifuged at 17,500 xg for 20 min at 4°C. Pellets were solubilized by boiling for 4 minwith 100 �l of 2� Laemmli sample buffer containing 8 M urea,and then diluted with 100 �l PBS. Supernatants were dilutedwith 100 �l of 2� Laemmli sample buffer with 8 M urea. Equalvolumes of pellets and supernatants were used for SDS-PAGE.
Preparation of Low-Density Membrane Fractions:Detergent-Containing Conditions
Detergent-insoluble glycolipid fractions were preparedfollowing an established method (Iwabuchi et al., 1998) withsome modifications. Briefly, CGNs (15 � 106 cells) at 7–8 DIVwere washed twice in PBS and then scraped in 700 �l ofice-cold lysis buffer (10 mM Tris-HCl pH 7.5, 150 mM NaCl,
654 Vacca et al.
1% Triton X-100, 2 mM EDTA, 1 mM PMSF, 20 �M leu-peptin). After 30 min, the lysate was centrifuged at 2,000 � g for10 min to remove nuclei and large debris. The resulting super-natant (500 �l) was mixed with an equal volume of 85% sucrosein Tris-buffered saline (TBS; 10 mM Tris-HCl, pH 7.5,150 mM NaCl), and placed at the bottom of the centrifuge tube.A volume of 700 �l of 35% sucrose in TBS and 400 �l of 5%sucrose were layered on the top of the lysate. The gradient wascentrifuged for 14 hr at 200,000 � g in a TLS 55 rotor (Beck-man Coulter, Palo Alto, CA). Four fractions of 150 �l and fivefractions of 300 �l were collected from the top of the tube. Theentire procedure was carried out at 4°C. The protein content ofeach fraction was determined by the Bradford method (Bio-Rad, Hercules, CA). For the preparation of low-density mem-brane fractions from total brain tissue, the brain was removedfrom 14-day-old Wistar rat, cut with a McIlwain tissue chopper(400 �m) in two orthogonal directions and resuspended in 10volumes (w/v) of lysis buffer with vortexing. After 60 min thelysate was passed five times through a 22-gauge needle andcentrifuged at 2,000 � g for 10 min. The remaining procedurewas identical to that for CGNs.
Preparation of Low-Density Membrane Fractions:Detergent-Free Conditions
Detergent-free preparation of low-density membranefractions was carried out following an established method (Songet al., 1996; Iwabuchi et al., 1998) with some modifications.Briefly, CGNs at 7–8 DIV (15 � 106 cells) or SH-SY5Y cells(5 � 106 cells) were washed twice in PBS and then scraped in700 �l of 500 mM Na2CO3 (pH 11) containing 2 mM EDTA,1 mM PMSF, 20 �M leupeptin. The cell suspension was ho-mogenized using a loose-fitting Dounce homogenizer (20strokes) and a sonicator (three 30-sec bursts). The homogenatewas mixed with an equal volume of 85% sucrose in MES-buffered saline (MBS) (25 mM MES, pH 6.5, 150 mM NaCl),and placed at the bottom of the centrifuge tube. A volume of
700 �l of 35% sucrose and 400 �l of 5% sucrose (both inMBS/500 mM Na2CO3, pH 11, 1:1 mixture) were layered onthe top of the lysate. The gradient was centrifuged for 14 hr at200,000 � g in a TLS 55 rotor (Beckman Instruments, PaloAlto, CA). Four fractions of 150 �l and five fractions of 300 �lwere collected from the top of the tube. The entire procedurewas carried out at 4°C. The protein content of each fraction wasdetermined with the Bradford method, using a Bio-Rad kit. Forpreparation of low-density membrane fractions from DRG, thefreshly isolated ganglia (30 for each preparation) were removedfrom 14-day-old Wistar rat, homogenized with a pestle andsonicated with 3 bursts of 20-sec in a bath sonicator. Thehomogenate was than centrifuged at 2,000 � g for 10 min toeliminate debris. The remaining procedure was identical to thatof CGNs.
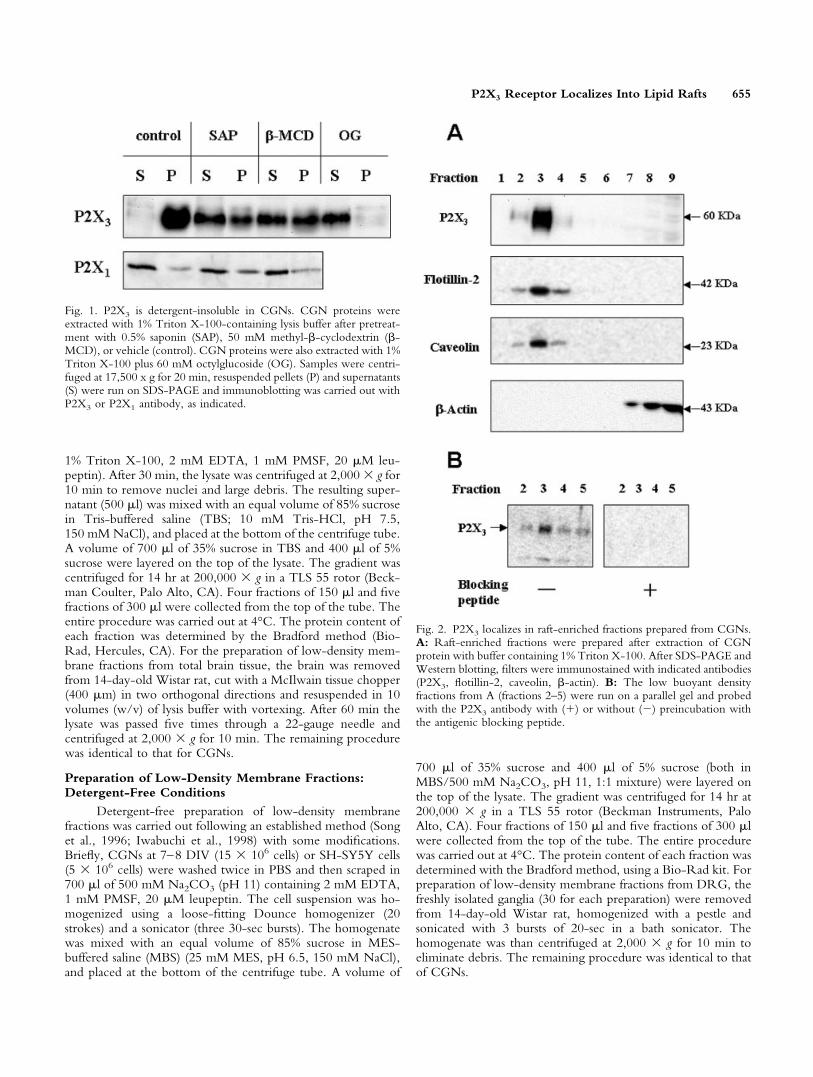
Fig. 1. P2X3 is detergent-insoluble in CGNs. CGN proteins wereextracted with 1% Triton X-100-containing lysis buffer after pretreat-ment with 0.5% saponin (SAP), 50 mM methyl-�-cyclodextrin (�-MCD), or vehicle (control). CGN proteins were also extracted with 1%Triton X-100 plus 60 mM octylglucoside (OG). Samples were centri-fuged at 17,500 x g for 20 min, resuspended pellets (P) and supernatants(S) were run on SDS-PAGE and immunoblotting was carried out withP2X3 or P2X1 antibody, as indicated.
Fig. 2. P2X3 localizes in raft-enriched fractions prepared from CGNs.A: Raft-enriched fractions were prepared after extraction of CGNprotein with buffer containing 1% Triton X-100. After SDS-PAGE andWestern blotting, filters were immunostained with indicated antibodies(P2X3, flotillin-2, caveolin, �-actin). B: The low buoyant densityfractions from A (fractions 2–5) were run on a parallel gel and probedwith the P2X3 antibody with (�) or without (�) preincubation withthe antigenic blocking peptide.
P2X3 Receptor Localizes Into Lipid Rafts 655

SDS-PAGE and Immunoblotting
Equal volumes (30 �l) from the sucrose gradient fractionswere analyzed by SDS-PAGE and immunoblotting (proteincontent ranging from 1–40 �g/fraction). Samples were dilutedin 4� Laemmli sample buffer, containing a final 2% (v/v)�-mercaptoethanol as reducing agent, and proteins were sepa-rated on 7.5 or 12.5% polyacrylamide gels. Blotting to nitrocel-lulose membranes (Amersham Biosciences, Uppsala, Sweden)was carried out using Tris-glycine transfer buffer with 20% (v/v)CH3OH. Nonspecific binding was prevented by incubating theblotted membranes in 5% (w/v) nonfat dry milk (NFDM) inTBS-T (Tris-buffered saline, 0.2% [v/v] Tween-20). Primaryantibodies were incubated for 2 hr at room temperature(flottillin-2, 1:5,000; caveolin, 1:5,000; �-actin, 1:2,500; P2X1,1:250; P2X2, 1:250; P2X4, 1:250; P2X7, 1:500) or overnight at4°C (P2X3, 1:3,000; P2Y4, 1:300). When indicated, a blockingantigenic peptide (10�5 M) was preincubated with the P2X3antibody for 30 min at room temperature, before hybridization.Secondary peroxidase-conjugated antibodies were used for50 min at room temperature. The antibodies were diluted inTBS-T with 3% (w/v) NFDM. Immunostained bands werevisualized using the ECL detection system (Amersham) andacquired with Kodak 440CF Image Station. Band intensitieswere measured with 1D image analysis software (EastmanKodak, New Haven, CT).
Immunofluorescence
CGNs at 7–8 DIV were fixed for 10 min in 4% parafor-maldehyde and permeabilized for 5 min with 0.1% TritonX-100 in PBS at room temperature. Nonspecific binding siteswere blocked with 10% normal donkey serum (Jackson Immu-
noresearch, West Baltimore Pike, PA) and 1% bovine serumalbumin (BSA) in PBS for 30 min at room temperature. Plateswere then incubated sequentially with anti-P2X3 (1:1,000) for12 hr at 4°C; anti-GFAP (1:400) for 1 hr at room temperature,a mixture of Cy2-conjugated donkey anti-mouse IgG (1:100),or Cy3-conjugated donkey anti-rabbit IgG (1:100) (JacksonImmunoresearch) for 1 hr at room temperature. All antibodieswere diluted in PBS/1% BSA. After three washes with PBS,plates were mounted with glass coverslips an antifading gel/mount (Biomeda, Foster City, CA). Double immunofluores-cence was analysed with a laser scanning confocal microscope(LSM 510; Zeiss, Jena, Germany).
RESULTSP2X3 Localizes in Lipid Rafts in CulturedCerebellar Granule Neurons
We observed that P2X3 was resistant to 1% TritonX-100 extraction in CGN cell extracts (Fig. 1). This is awell-recognized biochemical characteristic of proteins thatlocalize in lipid rafts and it is also well known that deter-gent insolubility of raft resident proteins is sensitive tocholesterol extraction from membranes. Cholesterol de-pletion was therefore carried out by pretreating CGNs at
Fig. 3. P2X3 also localizes in raft-enriched fractions when preparedwith a detergent-free procedure. Raft-enriched fractions were preparedfrom CGNs after extraction with carbonate buffer in detergent-freeconditions. After SDS-PAGE and Western blotting, filters were im-munostained with indicated antibodies (P2X3, flotillin-2, caveolin,�-actin).
Fig. 4. Not all P2 receptors localize in raft-enriched fractions. Raft-enriched fractions were prepared from CGNs after extraction with 1%Triton X-100-containing buffer. After SDS-PAGE and Western blot-ting, filters were immunostained with indicated antibodies (P2X1,P2X2, P2X4, P2X7, P2Y4, caveolin).
656 Vacca et al.

room temperature with either 0.5% saponin (Cerneus etal., 1993) or 50 mM methyl-�-cyclodextrin (Ilangumaranand Hoessli, 1998) before Triton extraction. In both con-ditions, a relevant fraction of the P2X3 was solubilized(60% with saponin; 50% with �-methyl-cyclodextrin)(Fig. 1), confirming that receptor insolubility could indeeddepend on lipid raft localization. Moreover, P2X3 wassolubilized almost completely by extracting CGN proteinswith 1% Triton X-100 plus 60 mM octyl-glucoside, aprocedure known to solubilize raft proteins (Brown andRose, 1992). To validate the detergent extraction proto-col, we tested the solubility of the P2X1 receptor, aprotein structurally similar to the P2X3 receptor. TheP2X1 receptor was solubilized mainly by 1% TritonX-100 extraction (over 90%). Moreover, the minor insol-uble fraction was not solubilized further by cholesterolextraction, suggesting that its insolubility could depend oncytoskeletal association and not on lipid raft localization(Fig. 1).
Together with detergent insolubility, the other maincharacteristic of lipid rafts is the low buoyant density insucrose gradient centrifugation. We prepared raft-enriched fractions from CGNs at 7–8 DIV, through su-crose gradient centrifugation, following a widely usedprocedure (Iwabuchi et al., 1998). We found that theP2X3 receptor was recovered almost completely from thelow-density fractions (Fig. 2A, fractions 2–4), togetherwith the lipid raft marker proteins caveolin and flotillin-2(Bickel et al., 1997), whereas the cytoskeletal protein�-actin was confined completely to the high-density frac-tions (fractions 7–9). The overall protein content of thelow-density fractions (fractions 1–4) was about 1.5% ofthe total (not shown), confirming that the raft-containingfractions excluded most cellular proteins. When the anti-serum used to identify P2X3 protein was preincubatedwith the immunizing peptide, the reactivity to this bandwas abolished completely, thereby confirming the speci-ficity of the antigen-antibody interaction (Fig. 2B).
As Triton X-100 can artefactually promote forma-tion of liquid-ordered raft-like domains in artificial lipidbilayers (Heerklotz, 2002), lipid rafts were prepared alsofrom CGNs following a well-established detergent-freeprocedure (Song et al., 1996). Under these conditions,P2X3 was also found in rafts-containing fractions (Fig. 3).The percent protein content of the low-density fractions(fractions 1–4) was not significantly different from thatobtained with the detergent-containing procedure (notshown). Sucrose gradient fractions, prepared underdetergent-containing conditions, were also probed for ad-ditional receptors of the P2X family, namely P2X1, P2X2,P2X4, and P2X7 and for the G protein-coupled P2Y4receptor. None of them localized in raft-containing frac-tions (Fig. 4). Even though CGN cultures were 95–98%pure, cells were immunostained with P2X3 and GFAPantibodies to confirm the localization of this receptor inneuronal cells. It was determined that P2X3 was indeedpresent in the cell bodies of CGNs, whereas it is absentfrom astrocytes, which represent the major contaminant inthese cell cultures (Levi et al., 1989) (Fig. 5).
Localization of P2X3 Receptor in Lipid RaftsPrepared From Rat Brain and Dorsal RootGanglia
We prepared lipid rafts from total brain and fromdorsal root ganglia. In total brains isolated from 14-day-oldrats, the distribution of P2X3 does not significantly differfrom that obtained in CGN preparations (Fig. 6A), al-though a small percentage of P2X3 was also detected in ahigh-density fraction (fraction 9). Similar results were ob-tained with lipid rafts prepared from newborn, 7-day-oldand 30-day-old rat brains or from 7- and 14-day-old ratcerebella (data not shown).
Lipid rafts were also prepared from dorsal root gan-glia, freshly isolated from 14-day-old rats. Different fromthe case with CGN and brain extracts, when thedetergent-containing procedure was used to isolate rafts,
Fig. 5. Immunochemical localization of P2X3 in CGNs. CGNs were immunostained with P2X3
(red) and GFAP (green). Immunofluorescence was detected with a laser scanning confocal micro-scope, using two independent channels. Figure can be viewed in color online via www.inter-science.wiley.com
P2X3 Receptor Localizes Into Lipid Rafts 657

the P2X3 receptor was almost completely segregated inhigh-density fractions (Fig. 6B). On the contrary, with thedetergent-free method, most P2X3 receptor was found inlow-density, raft-enriched fractions, together with caveo-lin (Fig. 6C). Similar results were obtained when lipid raftswere prepared from DRG isolated from 30-day-old rats(data not shown).
P2X3 Localizes Into Lipid Rafts Also WhenExogenously Expressed in SH-SY5Y HumanNeuroblastoma Cells
To confirm further the targeting of P2X3 into lipidrafts, we transiently expressed this receptor in the SH-
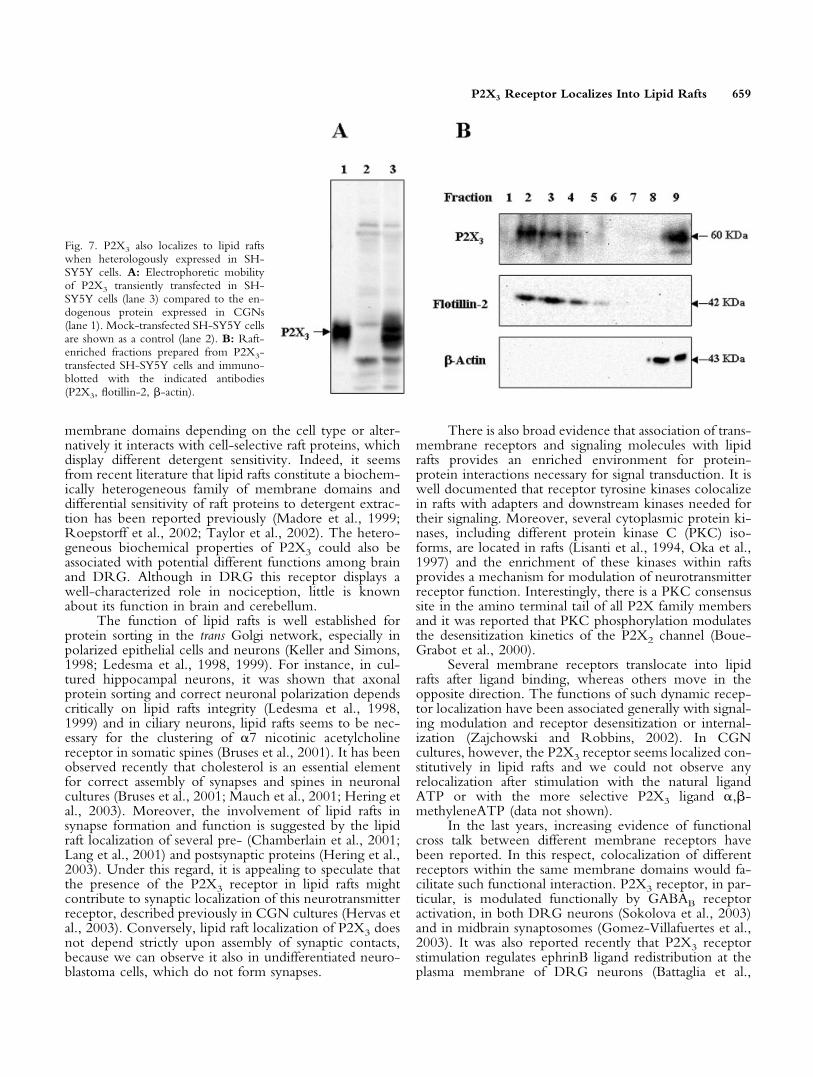
SY5Y neuroblastoma cell line, which does not endog-enously expresses appreciable levels of this protein. Thereceptor expressed in SH-SY5Y cells displays an electro-phoretic mobility similar to that of the endogenous recep-tor of CGNs (Fig. 7A). P2X3 more clearly appears as adoublet in SH-SY5Y, possibly resembling different post-traslational modifications. Lipid rafts were then preparedfrom SH-SY5Y cells, following both the detergent-freeand the detergent-containing procedures. As shown inFigure 7B, when the detergent-free procedure was used,the P2X3 receptor was recovered in the low-density frac-tions (fractions 2–4) also containing the raft markerflotillin-2, whereas �-actin was confined only to the high-density fractions (fractions 8, 9). Differently from thereceptor endogenously expressed in CGNs, a substantialfraction of the transfected receptor (�50% of total P2X3)was present in the high-density fraction (fraction 9). Whenthe detergent-containing method was used, P2X3 receptorwas confined completely to the high-density fractions(data not shown), as in the DRG.
DISCUSSIONThe results presented in this work provide direct
evidence that the P2X3 receptor localizes in membranedomains with biochemical characteristics distinctive oflipid rafts. In CGN primary cultures, we proved the spec-ificity of such localization by several different means. P2X3was resistant to detergent extraction at 4°C and this featurewas considerably sensitive to the extraction of cholesterolfrom the membranes, as expected for proteins associatedwith lipid rafts; P2X3 was highly enriched in detergent-resistant low buoyant density fractions, which instead ex-cluded additional P2 family members. Finally, P2X3 waspresent in lipid raft fractions prepared following a well-established detergent-free method. We used caveolin andflotillin-2 as controls for lipid raft preparation. Even ifcaveolin has been used already as a raft marker in neuronalcultures (Bruses et al., 2001; Vinson et al., 2003), itspresence on neuronal cells remains debated (Braun andMadison, 2000; Bu et al., 2003). It is beyond the scope ofthis work to show the presence of caveolin on CGNs. Weconfirmed by immunofluorescence, however, that P2X3receptor (Fig. 5) and caveolin (not shown) was expressedin CGNs and not on glial contaminants. We also observedthat P2X3 localized to raft-containing fractions preparedfrom total brain extracts and from dorsal root ganglia, andthat this localization was not modulated during postnataldevelopment of rat brain. P2X3 receptor was also associ-ated with lipid rafts when transiently transfected in SH-SY5Y neuroblastoma cells, favoring the hypothesis thatthis localization might be a molecular characteristic of theprotein, not strictly dependent on cell context.
It should be noted that the P2X3 receptor displaysdifferent sensitivity to Triton X-100 extraction dependingon the cells or tissues of origin. It is resistant to detergentextraction in CGN cultures and in brain tissue, whereas itis sensitive in neuroblastoma cells and in dorsal root gan-glia. This behavior has two possible explanations: theP2X3 receptor is localized to biochemically different
Fig. 6. P2X3 localizes in raft-enriched fractions prepared from total ratbrain and dorsal root ganglia. A: Raft-enriched fractions were preparedfrom total rat brain tissue obtained from 14-day-old animals extractedwith 1% Triton X-100-containing buffer. After SDS-PAGE and West-ern blotting, filters were immunostained with the indicated antibodies(P2X3, caveolin). Raft-enriched fractions were prepared from DRGobtained from 14-day-old animals following the detergent-containing(B) or detergent-free (C) procedures as described. After SDS-PAGEand Western blotting, the filters were immunostained with the indi-cated antibodies.
658 Vacca et al.
membrane domains depending on the cell type or alter-natively it interacts with cell-selective raft proteins, whichdisplay different detergent sensitivity. Indeed, it seemsfrom recent literature that lipid rafts constitute a biochem-ically heterogeneous family of membrane domains anddifferential sensitivity of raft proteins to detergent extrac-tion has been reported previously (Madore et al., 1999;Roepstorff et al., 2002; Taylor et al., 2002). The hetero-geneous biochemical properties of P2X3 could also beassociated with potential different functions among brainand DRG. Although in DRG this receptor displays awell-characterized role in nociception, little is knownabout its function in brain and cerebellum.
The function of lipid rafts is well established forprotein sorting in the trans Golgi network, especially inpolarized epithelial cells and neurons (Keller and Simons,1998; Ledesma et al., 1998, 1999). For instance, in cul-tured hippocampal neurons, it was shown that axonalprotein sorting and correct neuronal polarization dependscritically on lipid rafts integrity (Ledesma et al., 1998,1999) and in ciliary neurons, lipid rafts seems to be nec-essary for the clustering of �7 nicotinic acetylcholinereceptor in somatic spines (Bruses et al., 2001). It has beenobserved recently that cholesterol is an essential elementfor correct assembly of synapses and spines in neuronalcultures (Bruses et al., 2001; Mauch et al., 2001; Hering etal., 2003). Moreover, the involvement of lipid rafts insynapse formation and function is suggested by the lipidraft localization of several pre- (Chamberlain et al., 2001;Lang et al., 2001) and postsynaptic proteins (Hering et al.,2003). Under this regard, it is appealing to speculate thatthe presence of the P2X3 receptor in lipid rafts mightcontribute to synaptic localization of this neurotransmitterreceptor, described previously in CGN cultures (Hervas etal., 2003). Conversely, lipid raft localization of P2X3 doesnot depend strictly upon assembly of synaptic contacts,because we can observe it also in undifferentiated neuro-blastoma cells, which do not form synapses.
There is also broad evidence that association of trans-membrane receptors and signaling molecules with lipidrafts provides an enriched environment for protein-protein interactions necessary for signal transduction. It iswell documented that receptor tyrosine kinases colocalizein rafts with adapters and downstream kinases needed fortheir signaling. Moreover, several cytoplasmic protein ki-nases, including different protein kinase C (PKC) iso-forms, are located in rafts (Lisanti et al., 1994, Oka et al.,1997) and the enrichment of these kinases within raftsprovides a mechanism for modulation of neurotransmitterreceptor function. Interestingly, there is a PKC consensussite in the amino terminal tail of all P2X family membersand it was reported that PKC phosphorylation modulatesthe desensitization kinetics of the P2X2 channel (Boue-Grabot et al., 2000).
Several membrane receptors translocate into lipidrafts after ligand binding, whereas others move in theopposite direction. The functions of such dynamic recep-tor localization have been associated generally with signal-ing modulation and receptor desensitization or internal-ization (Zajchowski and Robbins, 2002). In CGNcultures, however, the P2X3 receptor seems localized con-stitutively in lipid rafts and we could not observe anyrelocalization after stimulation with the natural ligandATP or with the more selective P2X3 ligand �,�-methyleneATP (data not shown).
In the last years, increasing evidence of functionalcross talk between different membrane receptors havebeen reported. In this respect, colocalization of differentreceptors within the same membrane domains would fa-cilitate such functional interaction. P2X3 receptor, in par-ticular, is modulated functionally by GABAB receptoractivation, in both DRG neurons (Sokolova et al., 2003)and in midbrain synaptosomes (Gomez-Villafuertes et al.,2003). It was also reported recently that P2X3 receptorstimulation regulates ephrinB ligand redistribution at theplasma membrane of DRG neurons (Battaglia et al.,
Fig. 7. P2X3 also localizes to lipid raftswhen heterologously expressed in SH-SY5Y cells. A: Electrophoretic mobilityof P2X3 transiently transfected in SH-SY5Y cells (lane 3) compared to the en-dogenous protein expressed in CGNs(lane 1). Mock-transfected SH-SY5Y cellsare shown as a control (lane 2). B: Raft-enriched fractions prepared from P2X3-transfected SH-SY5Y cells and immuno-blotted with the indicated antibodies(P2X3, flotillin-2, �-actin).
P2X3 Receptor Localizes Into Lipid Rafts 659

2003). Interestingly, both GABAB receptor (Becher et al.,2001) and ephrinB ligands (Bruckner et al., 1999) havebeen identified as raft-resident proteins. The P2X3 local-ization in lipid rafts could therefore be an opportunity tointeract preferentially with regulatory proteins and to befunctionally coupled to additional receptors within thisspecific microenvironment. Moreover, the recognizedfunction of P2X3 receptor as mediator of pain-relatedsignals in DRG might suggest a novel mechanism involv-ing lipid rafts in the dynamic control of pain responses.
In conclusion, we have provided the novel demon-stration that a P2X family member is present in lipid rafts.The biological significance of this localization remains tobe elucidated for P2X3, as well as for other neurotrans-mitter receptors, but our finding might help to explain thephysiology of the P2X3 receptor, providing a novel po-tential mechanism for its correct targeting, dynamic acti-vation and function.
ACKNOWLEDGMENTSWe thank Prof. A. North for the generous gift of the
P2X3 expression plasmid. Fabrizio Vacca is the recipient ofa PhD fellowship from the University of Rome “La Sa-pienza”, Dottorato di Ricerca in “Neurofisiologia: basineurali di funzioni cognitive superiori.”
REFERENCESAmadio S, D’Ambrosi N, Cavaliere F, Murra B, Sancesario G, Bernardi G,
Burnstock G, Volonte C. 2002. P2 receptor modulation and cytotoxicfunction in cultured CNS neurons. Neuropharmacology 42:489–501.
Battaglia AA, Sehayek K, Grist J, McMahon SB, Gavazzi I. 2003. EphBreceptors and ephrin-B ligands regulate spinal sensory connectivity andmodulate pain processing. Nat Neurosci 6:339–340.
Becher A, White JH, McIlhinney RA. 2001. The gamma-aminobutyricacid receptor B, but not the metabotropic glutamate receptor type-1,associates with lipid rafts in the rat cerebellum. J Neurochem 79:787–795.
Bickel PE, Scherer PE, Schnitzer JE, Oh P, Lisanti MP, Lodish HF. 1997.Flotillin and epidermal surface antigen define a new family of caveolae-associated integral membrane proteins. J Biol Chem 272:13793–13802.
Boue-Grabot E, Archambault V, Seguela P. 2000. A protein kinase C sitehighly conserved in P2X subunits controls the desensitization kinetics ofP2X(2) ATP-gated channels. J Biol Chem 275:10190–10195.
Braun JE, Madison DV. 2000. A novel SNAP25-caveolin complex corre-lates with the onset of persistent synaptic potentiation. J Neurosci 20:5997–6006.
Brown DA, London E. 1998. Functions of lipid rafts in biological mem-branes. Annu Rev Cell Dev Biol. 14:111–136.
Brown DA, London E. 2000. Structure and function of sphingolipid- andcholesterol-rich membrane rafts. J Biol Chem 275:17221–17224.
Brown DA, Rose JK. 1992. Sorting of GPI-anchored proteins toglycolipid-enriched membrane subdomains during transport to the apicalcell surface. Cell 68:533–544.
Bruckner K, Pablo Labrador J, Scheiffele P, Herb A, Seeburg PH, Klein R.1999. EphrinB ligands recruit GRIP family PDZ adaptor proteins into raftmembrane microdomains. Neuron 22:511–524.
Bruses JL, Chauvet N, Rutishauser U. 2001. Membrane lipid rafts arenecessary for the maintenance of the (alpha)7 nicotinic acetylcholinereceptor in somatic spines of ciliary neurons. J Neurosci 21:504–512.
Bu J, Bruckner SR, Sengoku T, Geddes JW, Estus S. 2003 Glutamateregulates caveolin expression in rat hippocampal neurons. J Neurosci Res72:185–190.
Cavaliere F, Florenzano F, Amadio S, Fusco FR, Viscomi, MT, D’Ambrosi
N, Vacca F, Sancesario G, Bernardi G, Molinari M, Volonte C. 2003.Upregulation of P2X2 and P2X4 receptors during ischemic cell death:prevention by P2 antagonists. Neuroscience 120:85–98.
Cavaliere F, Sancesario G, Bernardi G, Volonte C. 2002. Extracellular ATPand nerve growth factor intensify hypoglycemia-induced cell death inprimary neurons: role of P2 and NGFRp75 receptors. J Neurochem83:1129–1138.
Cavaliere F, Amadio S, Sancesario G, Bernardi G, Volonte C. 2004.Synaptic P2X7 and oxygen/glucose deprivation in organotypic hip-pocampal cultures. J Cereb Blood Flow Metab (in press).
Cerneus DP, Ueffing E, Posthuma G, Strous GJ, van der Ende A. 1993.Detergent insolubility of alkaline phosphatase during biosynthetic trans-port and endocytosis. Role of cholesterol. J Biol Chem 268:3150–3155.
Chamberlain LH, Burgoyne RD, Gould GW. 2001. SNARE proteins arehighly enriched in lipid rafts in PC12 cells: implications for the spatialcontrol of exocytosis. Proc Natl Acad Sci USA 98:5619–5624.
Cockayne DA, Hamilton SG, Zhu QM, Dunn PM, Zhong Y, NovakovicS, Malmberg AB, Cain G, Berson A, Kassotakis L, Hedley L, LachnitWG, Burnstock G, McMahon SB, Ford AP. 2000. Urinary bladderhyporeflexia and reduced pain-related behaviour in P2X3-deficient mice.Nature 407:1011–1015.
Cook SP, Vulchanova L, Hargreaves KM, Elde R, McCleskey EW. 1997.Distinct ATP receptors on pain-sensing and stretch-sensing neurons.Nature 387:505–508.
Delling M, Wischmeyer E, Dityatev A, Sytnyk V, Veh RW, Karschin A,Schachner M. 2002. The neural cell adhesion molecule regulates cell-surface delivery of G-protein-activated inwardly rectifying potassiumchannels via lipid rafts. J Neurosci 22:7154–7164.
Garner CC, Nash J, Huganir RL. 2000. PDZ domains in synapse assemblyand signalling. Trends Cell Biol 10:274–280.
Gkantiragas I, Brugger B, Stuven E, Kaloyanova D, Li XY, Lohr K,Lottspeich F, Wieland FT, Helms JB. 2001 Sphingomyelin-enrichedmicrodomains at the Golgi complex. Mol Biol Cell 12:1819–1833.
Gomez-Villafuertes R, Pintor J, Gualix J, Miras-Portugal MT. 2003.GABAB receptor-mediated presynaptic potentiation of ATP ionotropicreceptors in rat midbrain synaptosomes. Neuropharmacology 44:311–323.
Heerklotz H. 2002. Triton promotes domain formation in lipid raft mix-tures. Biophys J 83:2693–2701.
Hering H, Lin CC, Sheng M. 2003. Lipid rafts in the maintenance ofsynapses, dendritic spines, and surface AMPA receptor stability. J Neu-rosci 23:3262–3271.
Hervas C, Perez-Sen R, Miras-Portugal MT. 2003. Coexpression of func-tional P2X and P2Y nucleotide receptors in single cerebellar granule cells.J Neurosci Res 73:384–399.
Ilangumaran S, Hoessli DC. 1998. Effects of cholesterol depletion bycyclodextrin on the sphingolipid microdomains of the plasma membrane.Biochem J 335:433–440.
Iwabuchi K, Yamamura S, Prinetti A, Handa K, Hakomori S. 1998.GM3-enriched microdomain involved in cell adhesion and signal trans-duction through carbohydrate-carbohydrate interaction in mouse mela-noma B16 cells. J Biol Chem 273:9130–9138.
Keller P, Simons K. 1998. Cholesterol is required for surface transport ofinfluenza virus hemagglutinin. J Cell Biol 140:1357–1367.
Kidd EJ, Miller KJ, Sansum AJ, Humphrey PP. 1998. Evidence for P2X3receptors in the developing rat brain. Neuroscience 87:533–539.
Lang T, Bruns D, Wenzel D, Riedel D, Holroyd P, Thiele C, Jahn R.2001. SNAREs are concentrated in cholesterol-dependent clusters thatdefine docking and fusion sites for exocytosis. EMBO J 20:2202–2213.
Ledesma MD, Brugger B, Bunning C, Wieland FT, Dotti CG.1999. Maturation of the axonal plasma membrane requires upregulationof sphingomyelin synthesis and formation of protein-lipid complexes.EMBO J 18:1761–1771.
660 Vacca et al.

Ledesma MD, Simons K, Dotti CG. 1998. Neuronal polarity: essential roleof protein-lipid complexes in axonal sorting. Proc Natl Acad Sci USA95:3966–3971.
Levi G, Aloisi F, Ciotti MT, Thangnipon W, Kingsbury A, Balazs R. 1989.Preparation of 98% pure cerebellar granule cell cultures. In: Shahar A,deVellis J, Vernadakis A, Haber B, editors. A dissection and tissue culturemanual of the nervous system. New York: Alan R. Liss, Inc. p 211–214.
Lisanti MP, Scherer PE, Vidugiriene J, Tang Z, Hermanowski-Vosatka A,Tu YH, Cook RF, Sargiacomo M. 1994. Characterization of caveolin-rich membrane domains isolated from an endothelial-rich source: impli-cations for human disease. J Cell Biol 126:111–126.
Llewellyn-Smith I, Burnstock G. 1998. Ultrastructural localization of P2X3receptors in rat sensory neurons. Neuroreport 9:2545–2550.
Madore N, Smith KL, Graham CH, Jen A, Brady K, Hall S, Morris R.1999. Functionally different GPI proteins are organized in different do-mains on the neuronal surface. EMBO J 18:6917–6926.
Marchand S, Devillers-Thiery A, Pons S, Changeux JP, Cartaud J. 2002.Rapsyn escorts the nicotinic acetylcholine receptor along the exocyticpathway via association with lipid rafts. J Neurosci 22:8891–8901.
Martens JR, Navarro-Polanco R, Coppock EA, Nishiyama A, Parshley L,Grobaski TD, Tamkun MM. 2000. Differential targeting of Shaker-likepotassium channels to lipid rafts. J Biol Chem 275:7443–7446.
Martens JR, Sakamoto N, Sullivan SA, Grobaski TD, Tamkun MM. 2001.Isoform-specific localization of voltage-gated K� channels to distinctlipid raft populations. Targeting of Kv1.5 to caveolae. J Biol Chem276:8409–844.
Mauch DH, Nagler K, Schumacher S, Goritz C, Muller EC, Otto A,Pfrieger FW. 2001. CNS synaptogenesis promoted by glia-derived cho-lesterol. Science 294:1354–1357.
Merlo D, Volonte C. 1996. Binding and functions of extracellular ATP incultured cerebellar granule neurons. Biochem Biophys Res Commun225:907–914.
Oka N, Yamamoto M, Schwencke C, Kawabe J, Ebina T, Ohno S, CouetJ, Lisanti MP, Ishikawa Y. 1997. Caveolin interaction with protein kinaseC. Isoenzyme-dependent regulation of kinase activity by the caveolinscaffolding domain peptide. J Biol Chem 272:33416–33421.
Ralevic V, Burnstock G. 1998. Receptors for purines and pyrimidines.Pharmacol Rev 50:413–492.
Robertson SJ, Ennion SJ, Evans RJ, Edwards FA. 2001. Synaptic P2Xreceptors. Curr Opin Neurobiol 11:378–386.
Roepstorff K, Thomsen P, Sandvig K, van Deurs B. 2002. Sequestration ofepidermal growth factor receptors in non-caveolar lipid rafts inhibitsligand binding. J Biol Chem 277:18954–18960.
Simons K, Ikonen E. 1997. Functional rafts in cell membranes. Nature387:569–572.
Smart EJ, Graf GA, McNiven MA, Sessa WC, Engelman JA, Scherer PE,Okamoto T, Lisanti MP 1999. Caveolins, liquid-ordered domains, andsignal transduction. Mol Cell Biol 19:7289–7304.
Sokolova E, Nistri A, Giniatullin R. 2003. The ATP-mediated fast currentof rat dorsal root ganglion neurons is a novel effector for GABA(B)receptor activation. Neurosci Lett 338:181–184.
Song KS, Li S, Okamoto T, Quilliam LA, Sargiacomo M, Lisanti MP.1996. Co-purification and direct interaction of Ras with caveolin, anintegral membrane protein of caveolae microdomains. Detergent-freepurification of caveolae microdomains. J Biol Chem 271:9690–9697.
Souslova V, Cesare P, Ding Y, Akopian AN, Stanfa L, Suzuki R, CarpenterK, Dickenson A, Boyce S, Hill R, Nebenuis-Oosthuizen D, Smith AJ,Kidd EJ, Wood JN. 2000. Warm-coding deficits and aberrant inflamma-tory pain in mice lacking P2X3 receptors. Nature 407:1015–1017.
Taylor CM, Coetzee T, Pfeiffer SE. 2002. Detergent-insolubleglycosphingolipid/cholesterol microdomains of the myelin membrane.J Neurochem 81:993–1004.
Tsui-Pierchala BA, Encinas M, Milbrandt J, Johnson EM Jr. 2002. Lipidrafts in neuronal signaling and function. Trends Neurosci 25:412–417.
Vinson M, Rausch O, Maycox PR, Prinjha RK, Chapman D, Morrow R,Harper AJ, Dingwall C, Walsh FS, Burbidge SA, Riddell DR. 2003.Lipid rafts mediate the interaction between myelin-associated glycopro-tein (MAG) on myelin and MAG-receptors on neurons. Mol Cell Neu-rosci 22:344–352.
Volonte C, Merlo D. 1996. Selected P2 purinoceptor modulators preventglutamate-evoked cytotoxicity in cultured cerebellar granule neurons.J Neurosci Res 45:183–193.
Vulchanova L, Riedl MS, Shuster SJ, Buell G, Surprenant A, North RA,Elde R. 1997. Immunohistochemical study of the P2X2 and P2X3receptor subunits in rat and monkey sensory neurons and their centralterminals. Neuropharmacology 36:1229–1242.
Zajchowski LD, Robbins SM. 2002. Lipid rafts and little caves. Compart-mentalized signalling in membrane microdomains. Eur J Biochem 269:737–752.
P2X3 Receptor Localizes Into Lipid Rafts 661





























