Embed Size (px)
Citation preview

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Neuroimmunomodulation 2013;20:65–71 DOI: 10.1159/000345044
Oxytocin Affects Nitric Oxide and Cytokine
Production by Sepsis-Sensitized Macrophages
Gabriela Ravanelli Oliveira-Pelegrin a Rafael Simone Saia b Evelin Capellari Cárnio c
Maria José Alves Rocha a
a Departamento de Morfologia, Estomatologia e Fisiologia, Faculdade de Odontologia de Ribeirão Preto, b Departamento de Fisiologia, Faculdade de Medicina de Ribeirão Preto, and c Departamento de Enfermagem Geral e Especializada, Escola de Enfermagem de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto , Brazil
creased. Macrophages of the early phase of sepsis pretreat-ed with OXT and stimulated with lipopolysaccharide showed decreased nitrite, TNF- � and IL-1 � levels, but no alteration in IL-10 production. In the late phase, they showed reduction only on IL-1 � . Conclusions: OXT secretion during sepsis may represent a neuroendocrine response contributing to the overall host response to infection by decreasing the proin-flammatory response and oxidative stress.
Copyright © 2012 S. Karger AG, Basel
Introduction
Sepsis is a potentially deadly medical condition char-acterized by a whole-body inflammatory state called sys-temic inflammatory syndrome that is triggered by an in-fection. Clinical and experimental studies have revealed alterations in vasopressin secretion during sepsis. In the early phase, the levels of vasopressin (AVP) are increased, but in the late phase these are diminished. These tempo-ral hormonal changes following the onset of sepsis are believed to be an attempt of the organism to normalize blood pressure that is decreased due to the production of inflammatory mediators like cytokines and nitric oxide (NO) [1–4] .
Key Words
Cecal ligation and puncture � Hypothalamus � Neurohypophysis � Paraventricular nucleus � Supraoptic nucleus � Peritoneal macrophage culture � Cytokines � Nitric oxide
Abstract
Background/Aim: Oxytocin (OXT) secretion during cecal li-gation puncture (CLP)-induced sepsis has not yet been ex-amined. Although immune properties have been attributed to OXT, its effect on CLP-sensitized macrophages has never been investigated. We analyzed OXT secretion during CLP and its effect in CLP-sensitized macrophage cultures. Meth-
ods: Male Wistar rats were decapitated 4, 6 or 24 h after CLP surgery or sham operation and blood, brain and neurohy-pophyses were collected for OXT measurements. In another set of animals we studied the effect of OXT on nitrite, tumor necrosis factor (TNF- � ), interleukin (IL)-1 � and IL-10 produc-tion of peritoneal macrophages harvested at 6 and 24 h after CLP. Results: In the early phase of sepsis (4–6 h), OXT levels increased in plasma and decreased in hypothalamus and neurohypophysis. In the late phase (24 h), plasma and neu-rohypophyseal levels remained basal. In the paraventricular, the OXT content remained low, but in the supraoptic in-
Received: August 31, 2012 Accepted after revision: October 8, 2012 Published online: November 23, 2012
Maria José A. Rocha Departamento de MEF – Faculdade de Odontologia de Ribeirão Preto, USP Avenida do Café s/n CEP Ribeirão Preto, SP 14040-904 (Brazil) E-Mail mjrocha @ forp.usp.br
© 2012 S. Karger AG, Basel1021–7401/13/0202–0065$38.00/0
Accessible online at:www.karger.com/nim

Oliveira-Pelegrin /Saia /Cárnio /Rocha
Neuroimmunomodulation 2013;20:65–71 66
Proinflammatory cytokines and NO are important activators of immune cells, increasing their ability of bac-terial clearance. The exacerbated production of tumor necrosis factor (TNF)- � , interleukin (IL)-1 � and NO in the early stages of sepsis is important for activation of im-mune cells, increasing their ability of bacterial clearance, however, they may also be responsible for complications, including hypotension, depression of myocardium con-tractility and organ failure [5, 6] . On the other hand, the anti-inflammatory cytokine IL-10 is released later and is thought to be important for the modulation of the in-flammatory response and reduction of mortality in septic animals [7] .
Oxytocin (OXT) is a neurohypophyseal hormone synthesized in the paraventricular (PVN) and supraop-tic (SON) nuclei of the hypothalamus [8] . Besides its very well-known role in female physiology, in labor and lactation, OXT also contributes to cardiovascular regu-lation, feeding and gastric distension, body temperature and certain behaviors [9, 10] . Although there are no clinical reports during sepsis regarding OXT secretion, an increase followed by a decrease in plasma OXT levels is observed during shock experimentally induced bylipopolysaccharide (LPS) [11, 12] . Its secretory profile during polymicrobial sepsis has, however, not yet been reported.
Peritonitis induced by cecal ligation puncture (CLP) is a clinically relevant experimental model for understand-ing the pathophysiology of sepsis. It closely resembles the characteristics of human sepsis that result in a prolonged and diminished release of inflammatory mediators, dif-ferent from the response seen following LPS injection, which is the most commonly used experimental model of endotoxic shock [13–15] . Even though there are reports showing the expression of OXT receptor in monocytes/macrophages, the role of OXT in the inflammatory re-sponse remains unclear [16–18] . In human experimental endotoxemia, the infusion of a supraphysiological OXT dose attenuated the excessive synthesis of cytokines and chemokines [19] . Furthermore, the subcutaneous injec-tion of OXT in rats showed anti-inflammatory proper-ties, reducing paw edema induced by carrageenan [20] . Moreover, even though the hormone is claimed to have immune properties its effect on CLP-sensitized macro-phages has never been investigated. We herein analyzed the time course of OXT secretion during polymicrobial sepsis and its effect on the production of inflammatory mediators by sepsis-sensitized macrophages.
Material and Methods
Animals Sixty-one male Wistar rats (200–250 g) were used in this study.
The animals, provided by the Animal Facility of the Campus of Ribeirão Preto, University of São Paulo (USP), were housed in controlled temperature (25 8 1 ° C) and photoperiodic (12: 12 h night:day cycle) conditions, with food (Nuvilab CR-1, NUVITAL) and tap water available ad libitum. All experimental protocols were approved and performed according to the guidelines of the Ethics Committee of USP, Campus Ribeirão Preto. Humane end-points in shock research [21] were used as criterion to euthanize CLP-animals in high suffering, immediately before or soon after the studied time-points defined in this study.
CLP Surgery Animals were randomly assigned to one of two groups, a CLP
and a sham-operated group. All experimental surgeries were started between 9: 00 and 11: 00 a.m.. Induction of severe sepsis was by a CLP model as previously described [4] . Briefly, rats were anesthetized with a short-acting anesthetic agent (tribromoetha-nol; 2.5%, 250 mg/kg i.p.; Acros Organics). Under sterile surgical conditions, the animals were subjected to a midline laparotomy and the cecum was carefully isolated to avoid damage to blood vessels. Subsequently, the cecum was ligated below the ileocecal valve, without causing bowel obstruction, and punctured twenty times with a 16-gauge needle, allowing fecal contents to spill into the peritoneum. The abdominal cavity was closed in two layers and all animals received a subcutaneous injection of saline (20 ml/kg body weight). Sham-operated animals were submitted to laparotomy and the cecum was manipulated, but neither ligated nor punctured. The animals were allowed to recover in their cag-es with free access to food and water.
Experimental Protocol Following sham operation or CLP surgery with twenty punc-
tures one set of animals was decapitated at 4 or 6 h (early phase of sepsis), or 24 h (late phase of sepsis). Blood and neurohypophyses were collected for OXT measurements by a radioimmunoassay (RIA). Brains were rapidly removed, snap-frozen on dry ice and stored at –80 ° C until determination of SON and PVN OXT con-tent. Another set of animals was decapitated at 6 and 24 h and the peritoneal lavage fluid collected for peritoneal macrophage cul-ture.
Microdissection of the SON and PVN Nuclei In a cryostat, the brains were placed in a brain matrix (Insight
Equipment LTDA, Ribeirão Preto, Brazil) and cut based on the rat brain atlas coordinates [22] . Using the optic chiasm as an ana-tomical landmark for reproducibility among the dissection sin-gle, approximately 1-mm thick sections were taken and the SON and PVN regions were carefully dissected using a needle with in-ner diameter of 1,200 and 1,400 �m, respectively [23] . The tissue punches were placed in 200 � l of 0.1 N HCl for measuring hor-mone content by RIA and total protein content by the Bradford method [24] .
OXT Measurement The plasma and tissue OXT levels were measured by means of
a double antibody-specific RIA. Briefly, 1 ml plasma samples were

Oxytocin Affects Macrophages inCLP-Induced Sepsis
Neuroimmunomodulation 2013;20:65–71 67
extracted using acetone/petroleum ether, lyophilized and stored at –80 ° C until analysis. Neurohypophyses, SON and PVN punch-es were homogenized in 200 � l of 0.1 N HCl. A 50- � l aliquot of the homogenate was used to measure protein content by the Brad-ford method. At the moment of the assay, the homogenates were properly diluted in assay buffer. Standard reagents and incubation protocols were used for the peptide assays. For peptide labeling, 125 I was purchased from a commercial supplier (GE Healthcare). OXT antiserum (Peninsula Laboratories Inc., San Carlos, Calif., USA) was used at a final dilution of 1: 90,000 in a phosphate buffer (0.062 M , pH 7.5) supplemented with BSA (0.5%). A nonequilib-rium assay was used with an incubation volume of 500 � l and an incubation time of 4 days at 4 ° C. Bound hormone was separated from unbound by a secondary antibody produced in the labora-tory of José Antunes-Rodrigues and Lucila L. K. Elias (Faculty of Medicine of Ribeirão Preto, USP, São Paulo, Brazil), where the RIA was performed. The minimum detection limit was 0.7 pg/ml and the coefficients of intra-and inter-assay variation were 0.4 and 0.8%, respectively.
Peritoneal Macrophage Culture Resident peritoneal cells from naive rats and inflammatory
cells recruited into the peritoneal cavity at 6 or 24 h after CLP were collected through washing the peritoneum with 10 ml of cold RPMI 1640 medium. After centrifugation (100 ! g for 10 min), the cell pellets were suspended in hypotonic solution (0.84% NH4Cl) for erythrocytes lysis. The remaining leukocytes were re-suspended with RPMI 1640 medium supplemented with 10% heat inactivated fetal bovine serum, 100 U/ml penicillin and 100 � g/ml streptomycin (complete medium; Sigma-Aldrich, St. Louis, Mo., USA). Leukocytes were counted using a hemocytometer and the differential cells were determined on Cytospin slides stained with May-Grunwald and Giemsa stain. Subsequently, the per-centage of macrophages was calculated and 1 ! 10 6 macrophages per well were dispensed into 24-well plates containing 1 ml of complete medium. After incubation at 37 ° C for 2 h in a humid heating chamber and 5% CO 2 atmosphere, the nonadherent cells were removed by washing each well once with PBS and twice with RPMI 1640 medium. The remaining cells were at least 97% mac-rophages and a trypan blue exclusion test showed that viability was higher than 92%. After temperature stabilization, the cul-tures were treated with vehicle (saline) or three different concen-trations of OXT (10– 12 , 10– 10 or 10– 8 M ; Sigma) for 2 h before ex-posure to LPS (1 � g/ml; Escherichia coli serotype O111:B4, Sig-ma). The macrophage cultures were incubated for 24 h after LPS addition, and supernatant medium was recovered for cytokine (TNF- � , IL-1 � and IL-10) and nitrite determination.
Cytokine Level Measurements The IL-1 � , IL-10 and TNF- � concentrations in culture super-
natant were determined using specific enzyme-linked immuno-sorbent assay (ELISA) kits for each cytokine (R&D Systems, Min-neapolis, Minn., USA) according to the manufacturer’s instruc-tions. The detection limits for IL-1 � , IL-10 and TNF- � specific ELISA kits were 5, 10 and 5 pg/ml, respectively.
Nitrite Determination The stable product of NO oxidation, nitrite, was measured in
cell culture medium using the Griess reaction. Samples were mixed with an equal volume of Griess reagent (2% sulfanilamide,
5% H3PO4, 0.2% naphthyl ethylenediaminedihy drochloride; Sig-ma) and incubated at room temperature for 10 min. The absor-bance of each sample was measured spectrophotometrically at 540 nm using sodium nitrite as a standard.
Statistical Analysis All values are expressed as means 8 SEM. RIA data were an-
alyzed using a logit transformation of the raw data. All samples were measured in the same session to reduce experimental varia-tion. Tissue hormone content was expressed in relation to the pro-tein content of each sample. Statistical differences were inferred from analysis of variance (ANOVA) followed by a post hoc Tukey test, and significance was considered at p ! 0.05.
Results
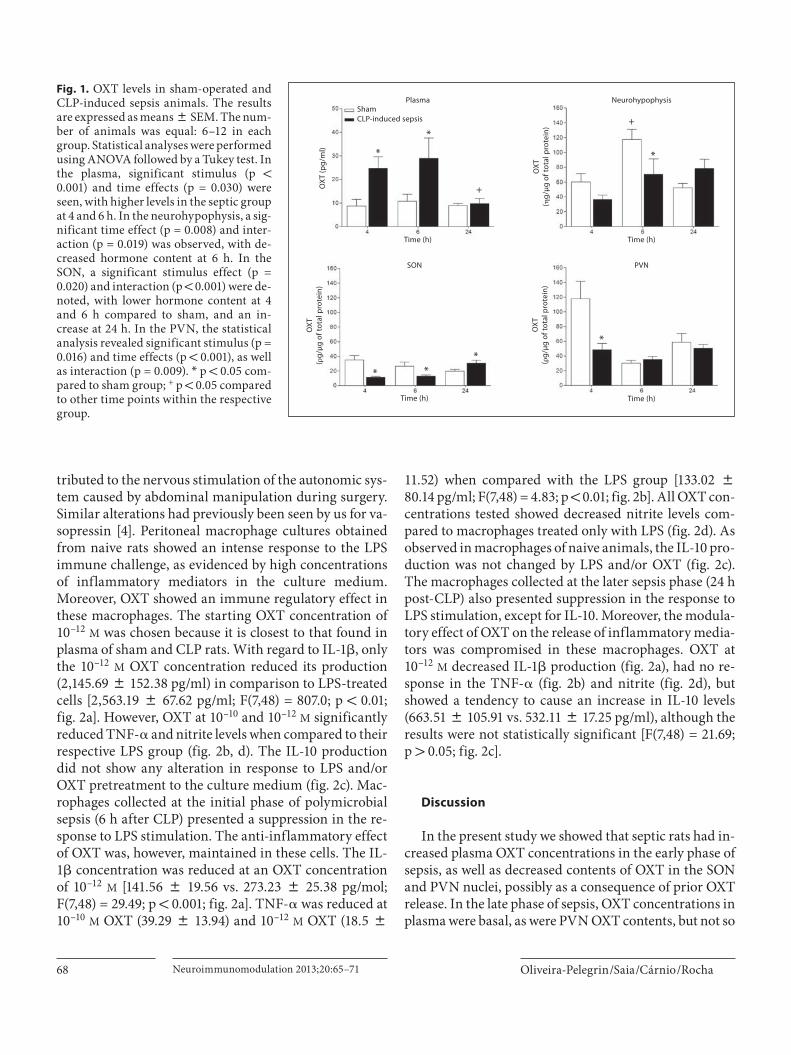
All animals submitted to CLP-induced sepsis devel-oped the typical early clinical signs of sepsis, including lethargy, piloerection and diarrhea as already reported in our previous work [25, 26] . Sham-operated animals, as expected, remained active in their cages without showing any such alterations. Plasma OXT values were higher in the CLP animals in the early phase (4 and 6 h) but re-turned to basal levels in the late phase of sepsis (24 h). Analysis by ANOVA revealed significant effects of stimu-lus [F(1,42) = 13.14; p ! 0.001] and time [F(2,42) = 3.83;p = 0.030] without interaction between the two. The post hoc test showed differences between sham and CLP at 4 and 6 h (p ! 0.05), and within the CLP group there was a significant decrease (p ! 0.05) at 24 h ( fig. 1 ). The OXT content in the neurohypophysis differed from the pattern seen for the plasma OXT levels, exhibiting a decrease in the early phase of sepsis and a tendency to increase inthe late phase (24 h). ANOVA showed significance in the time effect [F(2,41) = 5.37; p = 0.008] and interaction [F(2,41) = 4.39; p = 0.019] between time and stimulus with a significant decrease at 6 h after sepsis induction (117.4 8 13.5 ng/ � g CLP vs. 70.4 8 20.9 ng/ � g sham; p ! 0.05; fig. 1 ). In the early phase, the hypothalamic OXT content (pg/ � g protein) was significantly lower in the CLP com-pared to the sham group, in both SON [11.3 8 1.1 vs. 35.0 8 5.8; F(1,42) = 5.87; p = 0.020] and PVN [48.6 8 8.4 vs. 117.9 8 23.7; F(1,43) = 6.28; p = 0.016]. However, the pat-tern of changes in the two hypothalamic nuclei was dif-ferent. In the SON there was an increase in OXT content (pg/ � g protein) at 24 h after CLP [30.7 8 3.8 vs. 19.6 8 2.5; F(2,42) = 9.21; p ! 0.001], whereas in the PVN the dif-ferences in OXT content were not significant (50.4 8 5.1 vs. 58.7 8 12.0; fig. 1 ). Differences in OXT content were also seen in the sham group, both in the neurohypophysis (p ! 0.05) and PVN (p ! 0.001), and may possibly be at-
Oliveira-Pelegrin /Saia /Cárnio /Rocha
Neuroimmunomodulation 2013;20:65–71 68
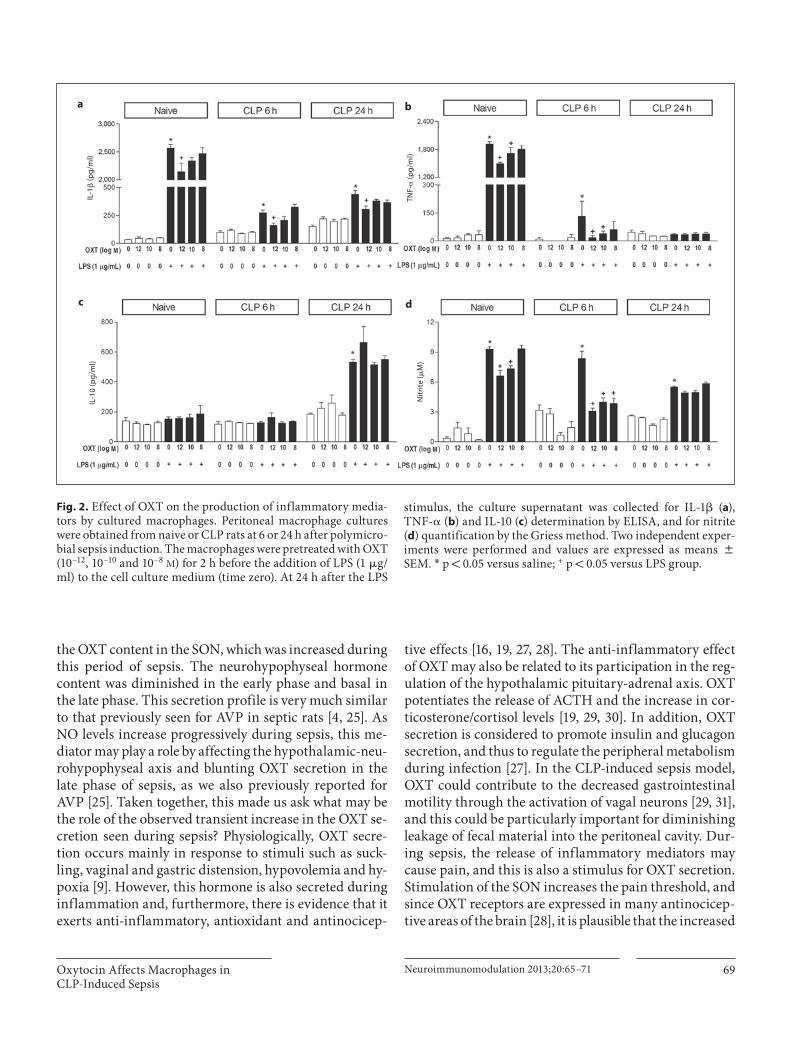
tributed to the nervous stimulation of the autonomic sys-tem caused by abdominal manipulation during surgery. Similar alterations had previously been seen by us for va-sopressin [4] . Peritoneal macrophage cultures obtained from naive rats showed an intense response to the LPS immune challenge, as evidenced by high concentrations of inflammatory mediators in the culture medium.Moreover, OXT showed an immune regulatory effect inthese macrophages. The starting OXT concentration of 10 –12 M was chosen because it is closest to that found in plasma of sham and CLP rats. With regard to IL-1 � , only the 10 –12 M OXT concentration reduced its production (2,145.69 8 152.38 pg/ml) in comparison to LPS-treated cells [2,563.19 8 67.62 pg/ml; F(7,48) = 807.0; p ! 0.01; fig. 2 a]. However, OXT at 10 –10 and 10 –12 M significantly reduced TNF- � and nitrite levels when compared to their respective LPS group ( fig. 2 b, d). The IL-10 production did not show any alteration in response to LPS and/or OXT pretreatment to the culture medium ( fig. 2 c). Mac-rophages collected at the initial phase of polymicrobial sepsis (6 h after CLP) presented a suppression in the re-sponse to LPS stimulation. The anti-inflammatory effect of OXT was, however, maintained in these cells. The IL-1 � concentration was reduced at an OXT concentration of 10 –12 M [141.56 8 19.56 vs. 273.23 8 25.38 pg/mol; F(7,48) = 29.49; p ! 0.001; fig. 2 a]. TNF- � was reduced at 10 –10 M OXT (39.29 8 13.94) and 10 –12 M OXT (18.5 8
11.52) when compared with the LPS group [133.02 8 80.14 pg/ml; F(7,48) = 4.83; p ! 0.01; fig. 2 b]. All OXT con-centrations tested showed decreased nitrite levels com-pared to macrophages treated only with LPS ( fig. 2 d). As observed in macrophages of naive animals, the IL-10 pro-duction was not changed by LPS and/or OXT ( fig. 2 c). The macrophages collected at the later sepsis phase (24 h post-CLP) also presented suppression in the response to LPS stimulation, except for IL-10. Moreover, the modula-tory effect of OXT on the release of inflammatory media-tors was compromised in these macrophages. OXT at10 –12 M decreased IL-1 � production ( fig. 2 a), had no re-sponse in the TNF- � ( fig. 2 b) and nitrite ( fig. 2 d), but showed a tendency to cause an increase in IL-10 levels (663.51 8 105.91 vs. 532.11 8 17.25 pg/ml), although the results were not statistically significant [F(7,48) = 21.69;p 1 0.05; fig. 2 c].
Discussion
In the present study we showed that septic rats had in-creased plasma OXT concentrations in the early phase of sepsis, as well as decreased contents of OXT in the SON and PVN nuclei, possibly as a consequence of prior OXT release. In the late phase of sepsis, OXT concentrations in plasma were basal, as were PVN OXT contents, but not so
OX
T (p
g/m
l)
OX
T(�
g/μ
g o
f tot
al p
rote
in)
OX
T(�
g/μ
g o
f tot
al p
rote
in)
OX
T(�
g/μ
g o
f tot
al p
rote
in)
ShamCLP-induced sepsis
Time (h) Time (h)
Time (h) Time (h)
Plasma Neurohypophysis
SON PVN
+
+
*
*
*
*
* **
Fig. 1. OXT levels in sham-operated and CLP-induced sepsis animals. The results are expressed as means 8 SEM. The num-ber of animals was equal: 6–12 in each group. Statistical analyses were performed using ANOVA followed by a Tukey test. In the plasma, significant stimulus (p ! 0.001) and time effects (p = 0.030) were seen, with higher levels in the septic group at 4 and 6 h. In the neurohypophysis, a sig-nificant time effect (p = 0.008) and inter-action (p = 0.019) was observed, with de-creased hormone content at 6 h. In the SON, a significant stimulus effect (p = 0.020) and interaction (p ! 0.001) were de-noted, with lower hormone content at 4 and 6 h compared to sham, and an in-crease at 24 h. In the PVN, the statistical analysis revealed significant stimulus (p = 0.016) and time effects (p ! 0.001), as well as interaction (p = 0.009). * p ! 0.05 com-pared to sham group; + p ! 0.05 compared to other time points within the respective group.
Oxytocin Affects Macrophages inCLP-Induced Sepsis
Neuroimmunomodulation 2013;20:65–71 69
the OXT content in the SON, which was increased during this period of sepsis. The neurohypophyseal hormone content was diminished in the early phase and basal in the late phase. This secretion profile is very much similar to that previously seen for AVP in septic rats [4, 25] . As NO levels increase progressively during sepsis, this me-diator may play a role by affecting the hypothalamic-neu-rohypophyseal axis and blunting OXT secretion in the late phase of sepsis, as we also previously reported for AVP [25] . Taken together, this made us ask what may be the role of the observed transient increase in the OXT se-cretion seen during sepsis? Physiologically, OXT secre-tion occurs mainly in response to stimuli such as suck-ling, vaginal and gastric distension, hypovolemia and hy-poxia [9] . However, this hormone is also secreted during inflammation and, furthermore, there is evidence that it exerts anti-inflammatory, antioxidant and antinocicep-
tive effects [16, 19, 27, 28] . The anti-inflammatory effect of OXT may also be related to its participation in the reg-ulation of the hypothalamic pituitary-adrenal axis. OXT potentiates the release of ACTH and the increase in cor-ticosterone/cortisol levels [19, 29, 30] . In addition, OXT secretion is considered to promote insulin and glucagon secretion, and thus to regulate the peripheral metabolism during infection [27] . In the CLP-induced sepsis model, OXT could contribute to the decreased gastrointestinal motility through the activation of vagal neurons [29, 31] , and this could be particularly important for diminishing leakage of fecal material into the peritoneal cavity. Dur-ing sepsis, the release of inflammatory mediators may cause pain, and this is also a stimulus for OXT secretion. Stimulation of the SON increases the pain threshold, and since OXT receptors are expressed in many antinocicep-tive areas of the brain [28] , it is plausible that the increased
IL-1
� (p
g/m
l)
TNF-
� (p
g/m
l)N
itri
te (�
M)
IL-1
0 (p
g/m
l)
a b
c d
Fig. 2. Effect of OXT on the production of inflammatory media-tors by cultured macrophages. Peritoneal macrophage cultures were obtained from naive or CLP rats at 6 or 24 h after polymicro-bial sepsis induction. The macrophages were pretreated with OXT (10–12, 10–10 and 10–8 M ) for 2 h before the addition of LPS (1 � g/ml) to the cell culture medium (time zero). At 24 h after the LPS
stimulus, the culture supernatant was collected for IL-1 � ( a ), TNF- � ( b ) and IL-10 ( c ) determination by ELISA, and for nitrite ( d ) quantification by the Griess method. Two independent exper-iments were performed and values are expressed as means 8 SEM. * p ! 0.05 versus saline; + p ! 0.05 versus LPS group.

Oliveira-Pelegrin /Saia /Cárnio /Rocha
Neuroimmunomodulation 2013;20:65–71 70
content of OXT in the SON in the late phase of CLP-in-duced sepsis may be related to antinociceptive effects of this hormone. OXT receptor expression has been report-ed in the thymus, lymphocytes and macrophages [17, 18] , and acute phase proteins and interleukins might affect the expression of this receptor [32, 33] . Taken together, this is all strong evidence for postulating a role of this hormone in the modulation of sepsis and other immune processes, including myocardial infarction [17, 18] . More-over, high doses of OXT prevented neutrophil influx and edema formation in paw inflammation induced by car-rageenan in rats, with the same potency as did the gluco-corticoid dexamethasone [20] . The particular effect of OXT on macrophage function is, however, not well dem-onstrated, despite the fact that OTX receptor expression in macrophages is almost 10 times higher than in mono-cytes [17] . During peritonitis, resident macrophages are important for sensing infections and producing proin-flammatory cytokines and chemokines that will assist in the recruitment of neutrophils and the clearance of mi-croorganisms from the infectious focus [34, 35] . In our study, the peritoneal resident and CLP-sensitized macro-phages obtained in the early phase of sepsis responded to OXT by reducing the LPS-induced synthesis of IL-1 � , TNF- � and nitrite. In the late phase of sepsis, however, macrophages showed poor responsiveness to OXT, except for IL-1 � . Our data is novel as it directly compares the anti-inflammatory effect of OXT with the time course of secretion of this hormone during polymicrobial sepsis in rats. Furthermore, the most important finding of our study is that naive and CLP-sensitized macrophages showed higher responses to OXT at concentrations close to physiological levels (approx. 10 –12 M ), suggesting an immunoregulatory role for this peptide in this experi-
mental model of sepsis. In the CLP-induced sepsis model, the injection of OXT appears to protect the organism against oxidative damage by acting as an antioxidant agent through inhibiting neutrophil infiltration and re-ducing lipid peroxidation, with all these effects contrib-uting to the maintenance of cellular integrity [16] . In fact, an antioxidant property of OXT could also be inferred in our study, seen as a decreased nitrite production by CLP-sensitized macrophages.
In the light of all these effects reported for OXT, our findings suggest that the transient increase in OXT secre-tion seen in the early phase of sepsis might be involved in modulating the inflammatory process. This clearly de-serves further study to elucidate the exact mode of action(s) of this hormone during sepsis.
Conclusion
OXT secretion during sepsis may represent a neuroen-docrine response contributing to the overall host re-sponse to infection by decreasing the proinflammatory response and oxidative stress.
Acknowledgments
The authors thank Nadir M. Fernandes, Milene M. Lopes and Maria Valci Silva for technical assistance. We also thank Dr. José Antunes-Rodrigues and Dr. Lucila L.K. Elias from the Depart-mento de Fisiologia, Faculdade de Medicina de Ribeirão Preto, USP, for providing the infrastructure for the RIA analyses. Finan-cial support from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) is gratefully acknowledged.
References
1 Landry DW, Levin HR, Gallant EM, Ashton RC Jr, Seo S, D’Alessandro D, Oz MC, Oliver JA: Vasopressin deficiency contributes to the vasodilation of septic shock. Circulation 1997; 95: 1122–1125.
2 Brierre S, Kumari R, Deboisblanc BP: The endocrine system during sepsis. Am J Med Sci 2004; 328: 238–247.
3 Maxime V, Siami S, Annane D: Metabolism modulators in sepsis: the abnormal pituitary response. Crit Care Med 2007; 35:S596–S601.
4 Oliveira-Pelegrin GR, Ravanelli MI, Branco LG, Rocha MJ: Thermoregulation and vaso-pressin secretion during polymicrobial sep-sis. Neuroimmunomodulation 2009; 16: 45–53.
5 Kumar A, Thota V, Dee L, Olson J, Uretz E, Parrillo JE: Tumor necrosis factor alpha and interleukin 1beta are responsible for in vitro myocardial cell depression induced by hu-man septic shock serum. J Exp Med 1996; 183: 949–958.
6 O’Dwyer MJ, Mankan AK, Stordeur P, O’Connell B, Duggan E, White M, Kelleher DP, McManus R, Ryan T: The occurrence of severe sepsis and septic shock are related to distinct patterns of cytokine gene expres-sion. Shock 2006; 26: 544–550.
7 Latifi SQ, O’Riordan MA, Levine AD: Inter-leukin-10 controls the onset of irreversible septic shock. Infect Immun 2002; 70: 4441–4446.
8 Cunningham ET Jr, Sawchenko PE: Reflex control of magnocellular vasopressin and oxytocin secretion. Trends Neurosci 1991; 14: 406–411.
9 Gutkowska J, Jankowski M, Mukaddam-Da-her S, McCann SM: Oxytocin is a cardiovas-cular hormone. Braz J Med Biol Res 2000; 33: 625–633.
10 Petersson M: Cardiovascular effects of oxy-tocin. Prog Brain Res 2002; 139: 281–288.
11 Borges BC, da Rocha MJ: Participation of the subfornical nucleus in hypothalamic-neuro-hypophyseal axis activation during the early phase of endotoxic shock. Neurosci Lett 2006; 404: 227–231.

Oxytocin Affects Macrophages inCLP-Induced Sepsis
Neuroimmunomodulation 2013;20:65–71 71
12 Stabile AM, Moreto V, Antunes-Rodrigues J, Carnio EC: Central but not systemic inhibi-tion of inducible nitric oxide synthase mod-ulates oxytocin release during endotoxemic shock. Peptides 2010; 31: 706–711.
13 Villa P, Sartor G, Angelini M, Sironi M, Con-ni M, Gnocchi P, Isetta AM, Grau G, Buur-man W, van Tits LJ, et al: Pattern of cytokines and pharmacomodulation in sepsis induced by cecal ligation and puncture compared with that induced by endotoxin. Clin Diagn Lab Immunol 1995; 2: 549–553.
14 Echtenacher B, Freudenberg MA, Jack RS, Mannel DN: Differences in innate defense mechanisms in endotoxemia and polymicro-bial septic peritonitis. Infect Immun 2001; 69: 7271–7276.
15 Dejager L, Pinheiro I, Dejonckheere E, Lib-ert C: Cecal ligation and puncture: the gold standard model for polymicrobial sepsis? Trends Microbiol 2011; 19: 198–208.
16 Iseri SO, Sener G, Saglam B, Gedik N, Ercan F, Yegen BC: Oxytocin protects against sep-sis-induced multiple organ damage: role of neutrophils. J Surg Res 2005; 126: 73–81.
17 Szeto A, Nation DA, Mendez AJ, Domin-guez-Bendala J, Brooks LG, Schneiderman N, McCabe PM: Oxytocin attenuates NADPH-dependent superoxide activity and IL-6 secretion in macrophages and vascular cells. Am J Physiol Endocrinol Metab 2008; 295:E1495–E1501.
18 Jankowski M, Bissonauth V, Gao L, Gangal M, Wang D, Danalache B, Wang Y, Stoya-nova E, Cloutier G, Blaise G, Gutkowska J: Anti-inflammatory effect of oxytocin in rat myocardial infarction. Basic Res Cardiol 2010; 105: 205–218.
19 Clodi M, Vila G, Geyeregger R, Riedl M, Stulnig TM, Struck J, Luger TA, Luger A: Oxytocin alleviates the neuroendocrine and cytokine response to bacterial endotoxin in healthy men. Am J Physiol Endocrinol Metab 2008; 295:E686–E691.
20 Petersson M, Wiberg U, Lundeberg T, Uv-nas-Moberg K: Oxytocin decreases carra-geenan induced inflammation in rats. Pep-tides 2001; 22: 1479–1484.
21 Nemzek JA, Xiao HY, Minard AE, Bolgos GL, Remick DG: Humane endpoints in shock research. Shock 2004; 21: 17–25.
22 Swanson LW: Brainmaps: Structure of the Rat Brain. Amsterdam, Elsevier Science, 1998.
23 Palkovits M: Isolated removal of hypotha-lamic or other brain nuclei of the rat. Brain Res 1973; 59: 449–450.
24 Bradford MM: A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976; 72: 248–254.
25 Correa PB, Pancoto JA, de Oliveira-Pelegrin GR, Carnio EC, Rocha MJ: Participation of iNOS-derived NO in hypothalamic activa-tion and vasopressin release during polymi-crobial sepsis. J Neuroimmunol 2007; 183: 17–25.
26 Oliveira-Pelegrin GR, Aguila FA, Basso PJ, Rocha MJ: Role of central NO-cGMP path-way in vasopressin and oxytocin gene ex-pression during sepsis. Peptides 2010; 31: 1847–1852.
27 Pittman DQ: A neuro-endocrine-immune symphony. J Neuroendocrinol 2011; 23: 1296–1297.
28 Yang J, Liang JY, Zhang XY, Qiu PY, Pan YJ, Li P, Zhang J, Hao F, Wang DX, Yan FL: Oxy-tocin, but not arginine vasopressin is involv-ing in the antinociceptive role of hypotha-lamic supraoptic nucleus. Peptides 2011; 32: 1042–1046.
29 Gimpl G, Fahrenholz F: The oxytocin recep-tor system: structure, function, and regula-tion. Physiol Rev 2001; 81: 629–683.
30 Ondrejcakova M, Bakos J, Garafova A, Ko-vacs L, Kvetnansky R, Jezova D: Neuroendo-crine and cardiovascular parameters during simulation of stress-induced rise in circulat-ing oxytocin in the rat. Stress 2010; 13: 314–322.
31 Rogers RC, Hermann GE: Oxytocin, oxyto-cin antagonist, TRH, and hypothalamic paraventricular nucleus stimulation effects on gastric motility. Peptides 1987; 8: 505–513.
32 Kimura T, Saji F: Molecular endocrinology of the oxytocin receptor. Endocr J 1995; 42: 607–615.
33 Elands J, Resink A, De Kloet ER: Neurohy-pophyseal hormone receptors in the rat thy-mus, spleen, and lymphocytes. Endocrinol-ogy 1990; 126: 2703–2710.
34 Henderson RB, Hobbs JA, Mathies M, Hogg N: Rapid recruitment of inflammatory monocytes is independent of neutrophil mi-gration. Blood 2003; 102: 328–335.
35 Knudsen E, Benestad HB, Seierstad T, Ivers-en PO: Macrophages in spleen and liver di-rect the migration pattern of rat neutrophils during inflammation. Eur J Haematol 2004; 73: 109–122.