Embed Size (px)
Citation preview

ORIGINAL PAPER
Overexpression of PeHSF mediates leaf ROS homeostasisin transgenic tobacco lines grown under salt stress conditions
Zedan Shen • Mingquan Ding • Jian Sun • Shurong Deng • Rui Zhao •
Meijuan Wang • Xujun Ma • Feifei Wang • Huilong Zhang • Zeyong Qian •
Yue Hu • Rui Yu • Xin Shen • Shaoliang Chen
Received: 31 January 2013 / Accepted: 26 July 2013 / Published online: 6 August 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Transcriptional responses of a heat-shock tran-
scription factor from the salt-resistant Populus euphratica,
PeHSF, were assessed in transgenic tobacco seedlings sub-
jected to salinity stress. Moreover, their relevance to NaCl-
induced anti-oxidative defense responses in higher plants
was also explored. Expression of PeHSF was induced by
high-salinity (150 mM NaCl) in leaves and callus cultures of
P. euphratica. The salt-stimulated PeHSF expression in P.
euphratica callus cultures was markedly inhibited by DPI
(an inhibitor of plasma membrane NADPH oxidase) and
LaCl3 (an inhibitor of plasma membrane Ca2?-permeable
channels), indicating the involvement of reactive oxygen
species (ROS) and Ca2? in HSF activation under NaCl stress.
Sequence analysis showed that PeHSF is a typical class A
HSF in higher plants. PeHSF overexpression in tobacco
significantly improved the salt tolerance of transgenic plants,
in term of seed germination and root growth. Compared to
the wild-type plants, the PeHSF-transgenic lines exhibited
an increased capacity to control ROS homeostasis, but not an
enhanced capacity to retain ionic homeostasis under salt
stress. PeHSF expression in tobacco enhanced the activities
of ascorbate peroxidase, glutathione peroxidase, and gluta-
thione reductase, due to upregulated transcriptional levels.
We conclude that PeHSF plays a crucial role in ROS
detoxification under salt stress, rather than in maintenance of
the K?/Na?. It is likely that the salt-induced H2O2 and
cytosolic Ca2? led to transcriptional activation of HSF,
which initiated the transcription of genes encoding antioxi-
dant enzymes in P. euphratica, thus contributing to ROS
homeostasis control under saline conditions.
Keywords Populus euphratica � Heat shock
transcription factor � NaCl � Antioxidant enzyme �H2O2 � Tobacco
Introduction
Salinity leads to reduced plant growth and crop produc-
tivity (Mahajan and Tuteja 2005), and some plants have
developed a signaling network to sense salinity stress. Heat
shock transcription factors (Hsfs) participate in multiple
stress responses, mediating plant responses to heat (Nover
et al. 1996; Swindell et al. 2007), low temperature (Sabehat
et al. 1998), osmotic stress (Sun et al. 2001), salt (Ogawa
et al. 2007), oxidative stress (Banzet et al. 1998; Lee et al.
2000; Volkov et al. 2006), desiccation (Liu et al. 2006),
intense light (Hihara et al. 2001; Rossel et al. 2002),
wounding (Cheong et al. 2002), and heavy metals
(Gyorgyey et al. 1991). Hsfs have a DNA-binding domain
(DBD), containing a helix-turn-helix motif that recog-
nizes and binds the heat shock elements (HSEs) of
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11240-013-0362-7) contains supplementarymaterial, which is available to authorized users.
Z. Shen � M. Ding � J. Sun � S. Deng � R. Zhao � M. Wang �X. Ma � F. Wang � H. Zhang � Z. Qian � Y. Hu � R. Yu �X. Shen � S. Chen (&)
College of Biological Sciences and Technology,
Beijing Forestry University, Box 162, Beijing 100083,
People’s Republic of China
e-mail: [email protected]
M. Ding
College of Agricultural and Food Science, Zhejiang Agricultural
and Forestry University, Hangzhou 311300, Zhejiang Province,
People’s Republic of China
J. Sun
College of Life Science, Jiangsu Normal University,
Xuzhou 221116, Jiangsu Province, People’s Republic of China
123
Plant Cell Tiss Organ Cult (2013) 115:299–308
DOI 10.1007/s11240-013-0362-7

Hsf-responsive genes (Damberger et al. 1994; Harrison et al.
1994; Vuister et al. 1994; Schultheiss et al. 1996). HSEs are
found in the promoter regions of a variety of defense genes
(Rizhsky et al. 2004). Upon stress, inactive Hsf monomers
form a homo-trimer, which can then be further modified by
phosphorylation or conformational changes to activate target
genes (Larson et al. 1988; Lee et al. 2000).
Higher plants possess a large Hsf family, with 18
members in tomato and 34 in soybean (Nover et al. 1996,
2001). The multigene Hsf family appears to benefit plants
that must respond to environmental variations. Different
Hsf family members have distinct roles in responding to
stress and activating target genes (Miller and Mittler 2006),
presumably due to their different structural characteristics.
There are three different classes of Hsfs: Class A, B, and C
(Nover et al. 2001). Class A Hsfs contain a C-terminal
activation domain and are thought to function as tran-
scriptional activators, while Class B and Class C Hsfs lack
a defined activation domain (Nover et al. 1996). Class B
Hsfs are reportedly transcription repressors (Boscheinen
et al. 1997; Czarnecka-Verner et al. 2000, 2004); however,
HsfB1 was recently demonstrated to function as a novel co-
regulator of tomato HsfA1 or HsfA2, enhancing their
transcriptional activity (Bharti et al. 2004). It has been
proposed that Hsfs target heat shock proteins (HSPs) (Ra-
bindran et al. 1993; Milioni et al. 2001). Different types of
Hsfs likely activate various defense pathways, assisting
plant adaptation to adverse environments; however, the
contribution Hsfs to salt tolerance in woody plants is
unknown.
Populus euphratica exhibits higher salt tolerance than
other poplars and is thus considered a model species for
studying tree stress physiology (Chen and Polle 2010).
There are several well-documented molecular and physio-
logical mechanisms underlying the salt tolerance of P.
euphratica (Chen et al. 2001, 2002a, b, 2003; Ottow et al.
2005; Sun et al. 2009a, b; Chen and Polle 2010). Tran-
scriptome analysis reveals that P. euphratica exhibits
permanent activation of control mechanisms for osmotic
adjustment (sugar and sugar alcohols) and ion compart-
mentalization (Ding et al. 2010; Janz et al. 2010). Fur-
thermore, salt stress signals (e.g., extracellular ATP, H2O2,
nitric oxide, and Ca2?) are involved in controlling ionic
homeostasis in P. euphratica (Sun et al. 2010a, b, 2012;
Zhang et al. 2007). These plants also exhibit a high
capacity to maintain anti-oxidant enzyme activity under
NaCl stress (Wang et al. 2007, 2008). Microarray analysis
has shown that salt stress upregulates the expression of a
class A HSF in P. euphratica (our unpublished data).
However, the link between HSF and oxidative stress has
not yet been established in P. euphratica.
The present study aimed to evaluate the transcriptional
response of PeHSF to salinity, and the relevance of this
response to NaCl-induced anti-oxidative defense. We
cloned the PeHSF gene from P. euphratica, and transferred
it to the model species Nicotiana tabacum to investigate the
role of this transcription factor in salinity tolerance. In
wild-type tobacco and PeHSF-overexpressing lines, we
examined ion relations, reactive oxygen species (ROS)
accumulation, and activities of antioxidant enzymes—such
as superoxide dismutase (SOD), catalase (CAT), ascorbate
peroxidase (APX), glutathione peroxidase (GPX), and
glutathione reductase (GR). Real-time PCR was used to
quantify the relative changes in the mRNA expression
levels of these antioxidant enzymes. Moreover, we inves-
tigated the correlations between ROS, Ca2?, and PeHSF
expression in P. euphratica callus cultures.
Materials and methods
Plant materials and treatments
One-year-old Populus euphratica Oliver (P. euphratica)
seedlings were obtained from the Xinjiang Uygur Auton-
omous Region of China. They were planted in individual
10-L pots containing loam soil, and placed in a greenhouse
at Beijing Forestry University. The potted plants were
irrigated two to three times per week according to evapo-
ration demand, and watered with 1 L full-strength Hoa-
gland’s nutrient solution every 2 weeks. The greenhouse
temperature was maintained at 20–25 �C, with a 16-h
photoperiod (7:00 AM–11:00 PM). Photosynthetically
active radiation was 150–600 lmol m-2 s-1 during the
2-month culture period. Seedlings were subjected to
4 weeks of NaCl stress by watering with 2 L of 150 mM
NaCl in full-strength Hoagland’s nutrient solution on days
1, 8, 15, and 23. For RNA extraction, upper mature leaves
(leaf index numbers 4–20 from shoot apex) were sampled
on days 2, 9, 22, and 29 after salt treatment.
P. euphratica callus cultures were started from shoots,
as described by Sun et al. (2010a, b). The calluses were
grown in Murashige and Skoog (1962) solid medium
(2.5 % sucrose, pH 5.7) supplemented with 0.25 mg L-1
benzyl adenine (BA) and 0.50 mg L-1 naphthalene acetic
acid (NAA). Callus cultures were raised in the dark at
25 �C, and subcultured every 15 days. Ten days after
transplantation, the calluses were sampled for salt and
inhibitor treatments on fresh propagation medium. P. eu-
phratica callus cultures were subjected to 150 mM NaCl,
with or without 100 lM DPI (diphenylene iodonium, an
inhibitor of PM NADPH oxidase) and/or 5 mM LaCl3 (an
inhibitor of PM Ca2? channels). Control callus cultures
were not subjected to the application of NaCl or inhibitors.
For RNA extraction, calluses were harvested after 1 and
12 h of salt treatment.
300 Plant Cell Tiss Organ Cult (2013) 115:299–308
123

Full-length PeHSF gene cloning and sequence analysis
Total RNA was isolated from P. euphratica leaves using
TRIzol reagent (Invitrogen) according to the manufac-
turer’s instructions. First-strand cDNA was synthesized
from 1 lg total RNA using the SuperScriptTM First Strand
Synthesis System. The 1437-nt fragment containing the
PeHSF ORF was amplified using specific primers (forward,
50-ATG GAT GGT TCA CAG AGT AA-30; reverse, 50-TCT GAC ATT ATG ACC TAT AC-30), and PCR prod-
ucts were cloned into the pMD18-T vector for sequencing.
The poplar putative amino acid sequences were aligned
using the multiple sequence alignment function of the
DNAMAN software. The phylogenetic tree was created
using the neighborhood-joining bootstrap (bootstrap NJ
Tree) function of the MEGA program (version 5.0).
The full-length PeHSF cDNA contained a 1437-bp open
reading frame encoding a putative protein of 479 amino
acids (Fig. S1). Phylogenetic analysis revealed PeHSF to
be a typical class A HSF (Doring et al. 2000; Fig. S2). The
putative PeHSF amino acid sequence contains a DNA-
binding domain (DBD), an adjacent domain with heptad
hydrophobic repeats (HR-A/B) involved in oligomeriza-
tion, nuclear localization sequences (NLS), two aromatic
hydrophobic acidic (AHA)-type activation domain, and a
nuclear export sequence (NES; Fig. S1). The putative Pe-
HSF protein from P. euphratica was highly homologous to
RcHsfA4 at position 20–120, which functions as a DNA-
binding domain (Fig. S1). Therefore, PeHSF can be clas-
sified as an HSFA4 with an E-Value of 9.3e-176 (http://
www.cibiv.at/services/hsf/).
Yeast one-hybrid assay
To construct the yeast-hybrid vector, the 1335-bp PeHSF
ORF was amplified using specific primers (forward, 50-CATG GAG GCC GAATTC-ATG GAT GGT TCA CAG
AGT AA-30; and reverse, 50-GC AGGTCGACGGATCC-
TCA TGA TCT TTC AGC TGT CGC AATG-30). Purified
PCR products were then fused in-frame with the GAL4BD
in the pGBKT7 vector by cloning into EcoRI and BamHI
sites. The recombinant plasmids were transformed into the
yeast strain Y2HGold using the YeastmakerTM Yeast
Transformation System 2. Yeast cells containing pGBKT7
were used as negative controls. The strains were tested
using synthetic defined (SD) media lacking histidine and
tryptophan (SD/-T-H), followed by X-a-Gal analysis.
X-a-Gal was directly added to the culture media to test
positive colonies (blue, direct visualization on the plate).
The transcriptional activity of PeHSF was tested with a
yeast one-hybrid assay. In contrast to the negative control,
the pGBKT7-PeHSF strain grew on SD media lacking
histidine and tryptophan (SD/-T-H; Fig. S3a). Moreover,
the pGBKT7-HSF strain also turned blue in the X-a-Gal
analysis (Fig. S3b). These results confirmed the transcrip-
tional activity of the PeHSF protein.
Overexpression of PeHSF in transgenic tobacco lines
To construct the 35S:PeHSF overexpression vector, a
1437-nt fragment containing the PeHSF ORF was ampli-
fied from P. euphratica cDNA using specific primers
(forward, 50-ATG GAT GGT TCA CAG AGT AA-30;reverse, 50-TCT GAC ATT ATG ACC TAT AC-30), and
then cloned into the pMD18-T vector. Next, the PeHSF
ORF was subcloned into the EcoRI and BamHI sites of the
pCABIA2300 vector driven by the 35S promoter. The
35S:PeHSF-2300 construct was introduced into Agrobac-
terium tumefaciens strain LBA4404, and then transformed
into wild-type N. tabacum using an Agrobacterium-medi-
ated method. The blank pCABIA2300 vector was intro-
duced into wild-type tobacco plants as a control. Ten lines
(#1–#10) of transgenic tobacco were obtained. The seeds of
T1 transformants were screened on half-strength MS agar
medium supplemented with 100 mg/L kanamycin. Four
positive transgenic lines #1, #2, #6, and #10 were con-
firmed by semi-PCR and real-time PCR using PeHSF-
specific primers (see below).
Among T1 progeny, the segregation ratio of kanamycin-
resistant (KanR) to kanamycin-sensitive (KanS) seedlings
ranged from 2.7 to 3.47 (Fig. S4a; Table S1), close to the
typical Mendelian ratio of 3:1, suggesting that the selected
transgenic tobacco lines (#2 and #6) harbored a single copy
of the transgene. We also examined the continuation of the
transgene expression. T2 seedlings of lines #2 and #6 could
grow on MS medium containing kanamycin (Fig. S4b), and
real-time PCR analysis showed PeHSF expression in both
lines (Fig. S4c). Wild-type and transgenic plants (T2) on
MS agar medium were exposed to short-term (24 h) and
long-term (10-15 days) of NaCl stress (100 mM). Seed
germination, root growth, ion relations, hydrogen peroxide
level, activity of antioxidant enzymes and gene expression
were examined at indicated time. The tobacco plants were
grown in a growth chamber at 25 ± 1 �C with relative
humidity of 50–60 %, a light intensity of 50 lmol m-2 s-1,
and a photoperiod of 16 h light/8 h dark.
Real-time PCR analysis
To examine the salt-induced expression of PeHSF in P. eu-
phratica, total RNA was isolated from P. euphratica leaves
and callus cultures grown under normal and saline condi-
tions. RNA was extracted with the Plant RNA Kit (QBio
Technologies Inc., Beijing, China) following the manufac-
turer’s introductions. The RNA was then treated with RNase-
free DNase (Promega, Madison, WI, USA), and first-strand
Plant Cell Tiss Organ Cult (2013) 115:299–308 301
123

cDNA was prepared as described above. One microliter of
cDNA product was used for SYBR Green-based real-time
PCR analysis in a Real-Time PCR System (MJ option2; Bio-
Rad, Hercules, CA, USA). Each sample was run in triplicate.
ACT7 (a housekeeping gene in P. euphratica; NCBI Ref Seq
acc. XM_002322628) was used as an internal control. ACT7
forward and reverse primers were 50-CAC ACT GGA GTG
ATG GTT GG-30 and 50-ATT GGC CTT GGG GTT AAG
AG-30, respectively. PeHSF forward and reverse primers
were 50-AACGCCCAGTTATCTCGGTGAT-30 and 50-AG
TCAGGAAGTGCTGCCAGAA-30, respectively. The PCR
running conditions were: 95 �C for 5 min, followed by 34
cycles of 94 �C for 30 s, 55 �C for 30 s, and 72 �C for 30 s,
with a final step of 72 �C for 10 min.
Real-time PCR was also performed on selected transgenic
tobacco lines with higher PeHSF expression. EF1a (a
housekeeping gene in tobacco; GenBank acc. D63396) was
used as an internal control. EF1a forward and reverse primers
were 50-GCT GTG AGG GAC ATG CGT CAA A-30 and 50-GTA GTA GAT ATC GCG AGT ACC ACC A-30, respec-
tively (Wang et al. 2013). Real-time quantification was based
on Ct values and the expression data was normalized to the
EF1a expression level. For semi-PCR, the PCR products were
separated in 2 % agarose gels, stained with ethidium bromide,
and then photographed under UV illumination.
Tissue ion analysis
Wild-type and transgenic seedlings were oven-dried (65 �C
for 4 days), ground, and passed through a 1.0-mm sieve for
mineral analysis. Na?, K?, Ca2? were analyzed as previ-
ously described (Chen et al. 2003; Wang et al. 2013).
H2O2 measurement
The H2O2-specific fluorescent probe H2DCF-DA (green;
Molecular Probe) was used to measure H2O2 (Leshem et al.
2007; Sun et al. 2010b). Roots were treated with 50 lM
H2DCF-DA (prepared in a Mes–KCl buffer, pH 5.7) for
5 min at room temperature in the dark. Then, the H2DCF-
DA-loaded plants were washed three to four times with the
basic solution. DCF-dependent fluorescence was measured
with a fluorescence microscope.
Activity and transcriptional analyses of ROS-
scavenging enzymes
Transgenic seedlings and wild-type tobacco (0.2 g) were
harvested, ground to a fine powder in liquid N2, and then
immediately homogenized in 2 mL of 50 mM potassium
phosphate buffer (pH 7.0) containing 1 mM ethylenediami-
netetraacetic acid (EDTA) and 1 % polyvinylpyrrolidone
(PVP; Jiang and Zhang 2003). The homogenate was centri-
fuged at 10,000g for 20 min at 4 �C, and the supernatant was
used to examine the antioxidant enzymes CAT, GR, APX, and
SOD, as described by Wang et al. (2008). Total GPX activity
was measured using the Cellular Glutathione Peroxidase
Assay Kit (Sigma), and the transcript levels of the antioxidant
enzymes were quantified by real-time PCR using gene-specific
primers (described in Supplementary Table S2).
Data analysis
Data were subjected to analysis of variance. Differences
between means were evaluated with Duncan’s multiple
range test. Unless otherwise stated, differences were con-
sidered statistically significant when P \ 0.05.
Results
Salt-induced PeHSF expression in P. euphratica
To clarify whether PeHSF was transcriptionally responsive
to high salt stress in P. euphratica leaves, we used real-
time PCR to examine the PeHSF transcript abundance
during NaCl treatment (150 mM, 29 days). The PeHSF
transcript level significantly increased by threefold on the
first day of salt treatment, followed by a gradual decline,
reaching a level similar to that in control plants at the end
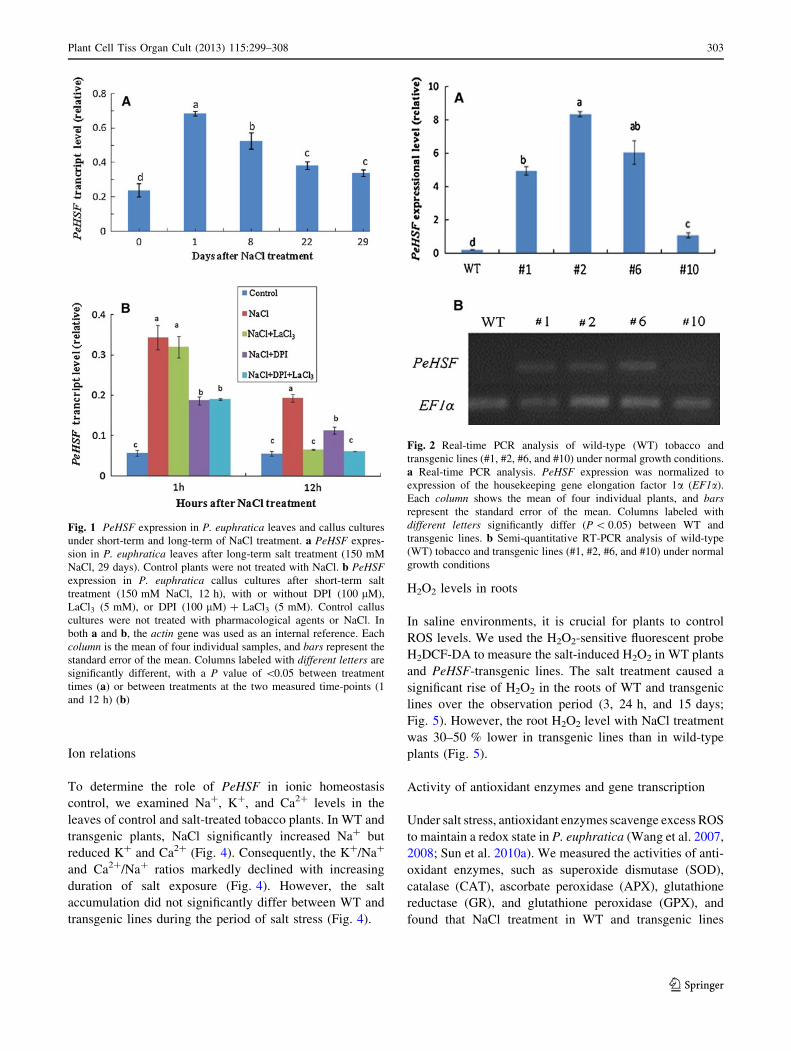
of the experiment (Fig. 1a).
Pharmacological experiments showed that PeHSF
expression in NaCl-treated P. euphratica callus cultures
was regulated by H2O2 and Ca2?. The PM NADPH oxidase
inhibitor DPI and the PM Ca2?-permeable channel blocker
LaCl3 each had a marked inhibitory effect on salt-induced
PeHSF expression at the two measured time-points (1 and
12 h; Fig. 1b).
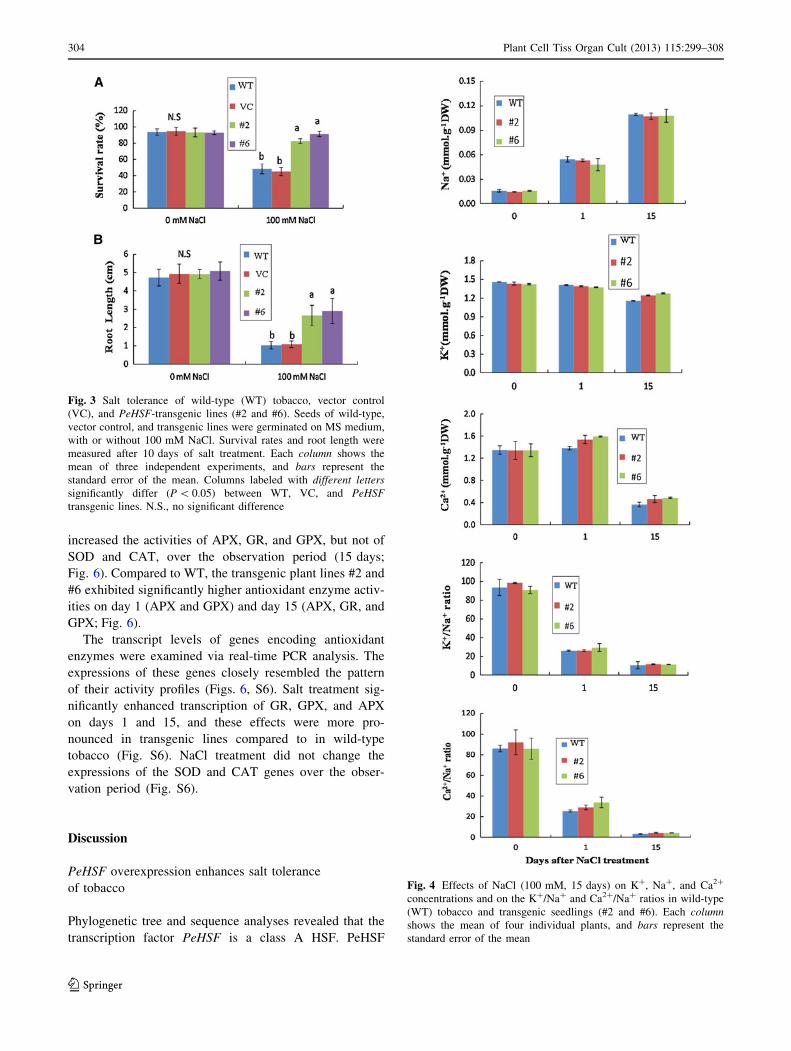
Seed germination, root growth, and salinity tolerance
To determine the role of the transcriptional factor in the salt
tolerance of P. euphratica, the PeHSF gene was trans-
formed into wild-type tobacco. Four positive transgenic
plant lines were subjected to semi-quantitative RT-PCR and
real-time PCR to check transgene expression (Fig. 2). The
two transgenic lines with higher PeHSF transcription levels
(#2 and #6, Fig. 2) were used for salt tolerance tests. Seeds
from wild-type tobacco and transgenic lines #2 and #6 (T2
generation) were germinated on MS medium without or
with NaCl (100 mM). Under NaCl stress, the two transgenic
lines showed significantly higher germination rates than
WT plants (Figs. 3a, S5a). Moreover, root length was less
reduced by salt treatment in the salinized #2 and #6 plants
than in WT plants (Figs. 3b, S5b).
302 Plant Cell Tiss Organ Cult (2013) 115:299–308
123
Ion relations
To determine the role of PeHSF in ionic homeostasis
control, we examined Na?, K?, and Ca2? levels in the
leaves of control and salt-treated tobacco plants. In WT and
transgenic plants, NaCl significantly increased Na? but
reduced K? and Ca2? (Fig. 4). Consequently, the K?/Na?
and Ca2?/Na? ratios markedly declined with increasing
duration of salt exposure (Fig. 4). However, the salt
accumulation did not significantly differ between WT and
transgenic lines during the period of salt stress (Fig. 4).
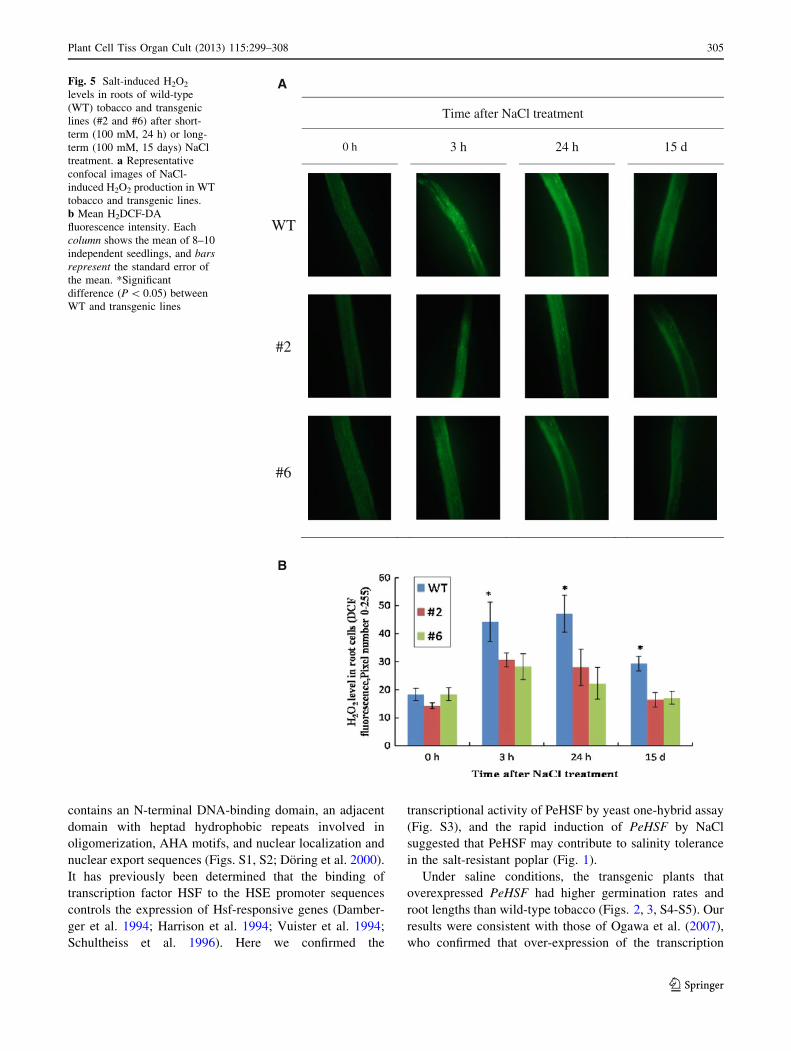
H2O2 levels in roots
In saline environments, it is crucial for plants to control
ROS levels. We used the H2O2-sensitive fluorescent probe
H2DCF-DA to measure the salt-induced H2O2 in WT plants
and PeHSF-transgenic lines. The salt treatment caused a
significant rise of H2O2 in the roots of WT and transgenic
lines over the observation period (3, 24 h, and 15 days;
Fig. 5). However, the root H2O2 level with NaCl treatment
was 30–50 % lower in transgenic lines than in wild-type
plants (Fig. 5).
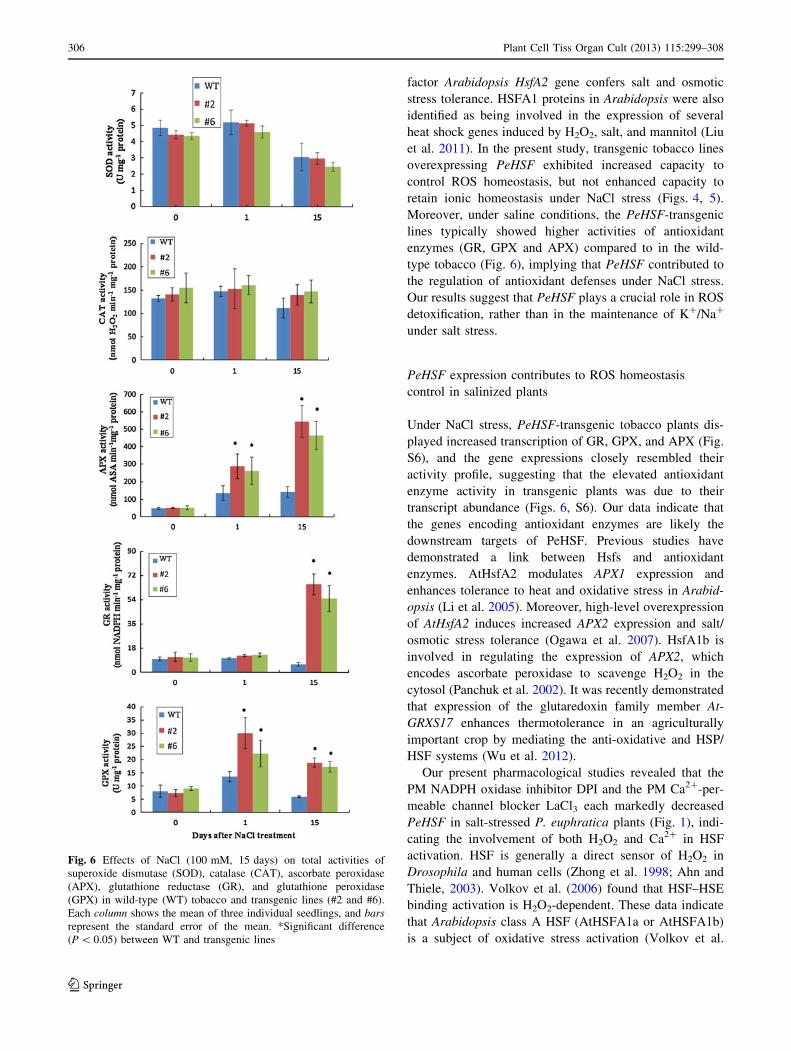
Activity of antioxidant enzymes and gene transcription
Under salt stress, antioxidant enzymes scavenge excess ROS
to maintain a redox state in P. euphratica (Wang et al. 2007,
2008; Sun et al. 2010a). We measured the activities of anti-
oxidant enzymes, such as superoxide dismutase (SOD),
catalase (CAT), ascorbate peroxidase (APX), glutathione
reductase (GR), and glutathione peroxidase (GPX), and
found that NaCl treatment in WT and transgenic lines
Fig. 1 PeHSF expression in P. euphratica leaves and callus cultures
under short-term and long-term of NaCl treatment. a PeHSF expres-
sion in P. euphratica leaves after long-term salt treatment (150 mM
NaCl, 29 days). Control plants were not treated with NaCl. b PeHSF
expression in P. euphratica callus cultures after short-term salt
treatment (150 mM NaCl, 12 h), with or without DPI (100 lM),
LaCl3 (5 mM), or DPI (100 lM) ? LaCl3 (5 mM). Control callus
cultures were not treated with pharmacological agents or NaCl. In
both a and b, the actin gene was used as an internal reference. Each
column is the mean of four individual samples, and bars represent the
standard error of the mean. Columns labeled with different letters are
significantly different, with a P value of \0.05 between treatment
times (a) or between treatments at the two measured time-points (1
and 12 h) (b)
Fig. 2 Real-time PCR analysis of wild-type (WT) tobacco and
transgenic lines (#1, #2, #6, and #10) under normal growth conditions.
a Real-time PCR analysis. PeHSF expression was normalized to
expression of the housekeeping gene elongation factor 1a (EF1a).
Each column shows the mean of four individual plants, and bars
represent the standard error of the mean. Columns labeled with
different letters significantly differ (P \ 0.05) between WT and
transgenic lines. b Semi-quantitative RT-PCR analysis of wild-type
(WT) tobacco and transgenic lines (#1, #2, #6, and #10) under normal
growth conditions
Plant Cell Tiss Organ Cult (2013) 115:299–308 303
123
increased the activities of APX, GR, and GPX, but not of
SOD and CAT, over the observation period (15 days;
Fig. 6). Compared to WT, the transgenic plant lines #2 and
#6 exhibited significantly higher antioxidant enzyme activ-
ities on day 1 (APX and GPX) and day 15 (APX, GR, and
GPX; Fig. 6).
The transcript levels of genes encoding antioxidant
enzymes were examined via real-time PCR analysis. The
expressions of these genes closely resembled the pattern
of their activity profiles (Figs. 6, S6). Salt treatment sig-
nificantly enhanced transcription of GR, GPX, and APX
on days 1 and 15, and these effects were more pro-
nounced in transgenic lines compared to in wild-type
tobacco (Fig. S6). NaCl treatment did not change the
expressions of the SOD and CAT genes over the obser-
vation period (Fig. S6).
Discussion
PeHSF overexpression enhances salt tolerance
of tobacco
Phylogenetic tree and sequence analyses revealed that the
transcription factor PeHSF is a class A HSF. PeHSF
Fig. 3 Salt tolerance of wild-type (WT) tobacco, vector control
(VC), and PeHSF-transgenic lines (#2 and #6). Seeds of wild-type,
vector control, and transgenic lines were germinated on MS medium,
with or without 100 mM NaCl. Survival rates and root length were
measured after 10 days of salt treatment. Each column shows the
mean of three independent experiments, and bars represent the
standard error of the mean. Columns labeled with different letters
significantly differ (P \ 0.05) between WT, VC, and PeHSF
transgenic lines. N.S., no significant difference
Fig. 4 Effects of NaCl (100 mM, 15 days) on K?, Na?, and Ca2?
concentrations and on the K?/Na? and Ca2?/Na? ratios in wild-type
(WT) tobacco and transgenic seedlings (#2 and #6). Each column
shows the mean of four individual plants, and bars represent the
standard error of the mean
304 Plant Cell Tiss Organ Cult (2013) 115:299–308
123
contains an N-terminal DNA-binding domain, an adjacent
domain with heptad hydrophobic repeats involved in
oligomerization, AHA motifs, and nuclear localization and
nuclear export sequences (Figs. S1, S2; Doring et al. 2000).
It has previously been determined that the binding of
transcription factor HSF to the HSE promoter sequences
controls the expression of Hsf-responsive genes (Damber-
ger et al. 1994; Harrison et al. 1994; Vuister et al. 1994;
Schultheiss et al. 1996). Here we confirmed the
transcriptional activity of PeHSF by yeast one-hybrid assay
(Fig. S3), and the rapid induction of PeHSF by NaCl
suggested that PeHSF may contribute to salinity tolerance
in the salt-resistant poplar (Fig. 1).
Under saline conditions, the transgenic plants that
overexpressed PeHSF had higher germination rates and
root lengths than wild-type tobacco (Figs. 2, 3, S4-S5). Our
results were consistent with those of Ogawa et al. (2007),
who confirmed that over-expression of the transcription
Time after NaCl treatment
0 h 3 h 24 h 15 d
WT
#2
#6
A
B
Fig. 5 Salt-induced H2O2
levels in roots of wild-type
(WT) tobacco and transgenic
lines (#2 and #6) after short-
term (100 mM, 24 h) or long-
term (100 mM, 15 days) NaCl
treatment. a Representative
confocal images of NaCl-
induced H2O2 production in WT
tobacco and transgenic lines.
b Mean H2DCF-DA
fluorescence intensity. Each
column shows the mean of 8–10
independent seedlings, and bars
represent the standard error of
the mean. *Significant
difference (P \ 0.05) between
WT and transgenic lines
Plant Cell Tiss Organ Cult (2013) 115:299–308 305
123
factor Arabidopsis HsfA2 gene confers salt and osmotic
stress tolerance. HSFA1 proteins in Arabidopsis were also
identified as being involved in the expression of several
heat shock genes induced by H2O2, salt, and mannitol (Liu
et al. 2011). In the present study, transgenic tobacco lines
overexpressing PeHSF exhibited increased capacity to
control ROS homeostasis, but not enhanced capacity to
retain ionic homeostasis under NaCl stress (Figs. 4, 5).
Moreover, under saline conditions, the PeHSF-transgenic
lines typically showed higher activities of antioxidant
enzymes (GR, GPX and APX) compared to in the wild-
type tobacco (Fig. 6), implying that PeHSF contributed to
the regulation of antioxidant defenses under NaCl stress.
Our results suggest that PeHSF plays a crucial role in ROS
detoxification, rather than in the maintenance of K?/Na?
under salt stress.
PeHSF expression contributes to ROS homeostasis
control in salinized plants
Under NaCl stress, PeHSF-transgenic tobacco plants dis-
played increased transcription of GR, GPX, and APX (Fig.
S6), and the gene expressions closely resembled their
activity profile, suggesting that the elevated antioxidant
enzyme activity in transgenic plants was due to their
transcript abundance (Figs. 6, S6). Our data indicate that
the genes encoding antioxidant enzymes are likely the
downstream targets of PeHSF. Previous studies have
demonstrated a link between Hsfs and antioxidant
enzymes. AtHsfA2 modulates APX1 expression and
enhances tolerance to heat and oxidative stress in Arabid-
opsis (Li et al. 2005). Moreover, high-level overexpression
of AtHsfA2 induces increased APX2 expression and salt/
osmotic stress tolerance (Ogawa et al. 2007). HsfA1b is
involved in regulating the expression of APX2, which
encodes ascorbate peroxidase to scavenge H2O2 in the
cytosol (Panchuk et al. 2002). It was recently demonstrated
that expression of the glutaredoxin family member At-
GRXS17 enhances thermotolerance in an agriculturally
important crop by mediating the anti-oxidative and HSP/
HSF systems (Wu et al. 2012).
Our present pharmacological studies revealed that the
PM NADPH oxidase inhibitor DPI and the PM Ca2?-per-
meable channel blocker LaCl3 each markedly decreased
PeHSF in salt-stressed P. euphratica plants (Fig. 1), indi-
cating the involvement of both H2O2 and Ca2? in HSF
activation. HSF is generally a direct sensor of H2O2 in
Drosophila and human cells (Zhong et al. 1998; Ahn and
Thiele, 2003). Volkov et al. (2006) found that HSF–HSE
binding activation is H2O2-dependent. These data indicate
that Arabidopsis class A HSF (AtHSFA1a or AtHSFA1b)
is a subject of oxidative stress activation (Volkov et al.
Fig. 6 Effects of NaCl (100 mM, 15 days) on total activities of
superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase
(APX), glutathione reductase (GR), and glutathione peroxidase
(GPX) in wild-type (WT) tobacco and transgenic lines (#2 and #6).
Each column shows the mean of three individual seedlings, and bars
represent the standard error of the mean. *Significant difference
(P \ 0.05) between WT and transgenic lines
306 Plant Cell Tiss Organ Cult (2013) 115:299–308
123

2006). Furthermore, HsfA4a is a candidate for a sensor
molecule involved in H2O2 perception in Arabidopsis
(Davletova et al. 2005), and Ca2? and calmodulin modulate
DNA-binding activity of Hsf in maize (Li et al. 2004).
Our previous studies have repeatedly shown that salt
stress results in an early H2O2 burst and elevation of
cytosolic Ca2? ([Ca2?]cyt) in P. euphratica callus cells
(Sun et al. 2010a, b, 2012). P. euphratica plants rapidly up-
regulated anti-oxidant enzyme activity upon NaCl expo-
sure (Wang et al. 2007, 2008). Overall, based on our
present data, we conclude that the salt-induced H2O2 and
[Ca2?]cyt led to transcriptional activation of HSF, which
initiated transcription of target genes encoding antioxidant
enzymes in P. euphratica, thus contributing to ROS
homeostasis control. However, future research is needed to
explore how HSFs respond to salt-elicited signals and
regulate the expression of downstream target genes.
Acknowledgments This research was supported jointly by the
National Natural Science Foundation of China (Grant Nos. 31270654,
31170570, and 31200470), the Key Grant Project of the Chinese Min-
istry of Education (Grant No. 2013001), the Beijing Natural Science
Foundation (Grant No. 6112017), the Program of Introducing Talents of
Discipline to Universities (111 Project, Grant No. B13007), Funda-
mental Research Funds for the Central Universities (Grant Nos.
JC2011-2 and TD-2012-04), the Foundation for the Supervisors of
Beijing Excellent Doctoral Dissertations (Grant No. YB20081002201).
References
Ahn SG, Thiele DJ (2003) Redox regulation of mammalian heat
shock factor 1 is essential for Hsp gene activation and protection
from stress. Genes Dev 17:516–528
Banzet N, Richaud C, Deveaux Y, Kazmaier M, Gagnon J,
Triantaphylides C (1998) Accumulation of small heat shock
proteins, including mitochondrial HSP22, induced by oxidative
stress and adaptive response in tomato cells. Plant J 13:519–527
Bharti K, Von Koskull-Doring P, Bharti S, Kumar P, Tintschl-
Korbitzer A, Treuter E, Nover L (2004) Tomato heat stress
transcription factor HsfB1 represents a novel type of general
transcription coactivator with a histone-like motif interacting
with the plant CREB binding protein ortholog HAC1. Plant Cell
16:1521–1535
Boscheinen O, Lyck R, Queitsch C, Treuter E, Zimarino V, Scharf
KD (1997) Heat stress transcription factors from tomato can
functionally replace HSF1 in the yeast Saccharomyces cerevi-
siae. Mol Gen Genet 255:322–331
Chen SL, Polle A (2010) Salinity tolerance of Populus. Plant Biol
12:317–333
Chen SL, Li JK, Wang SS, Huttermann A, Altman A (2001) Salt,
nutrient uptake and transport, and ABA of Populus euphratica; a
hybrid in response to increasing soil NaCl. Trees 15:186–194
Chen SL, Li J, Wang T, Wang S, Polle A, Huttermann A (2002a)
Osmotic stress and ion-specific effects on xylem abscisic acid
and the relevance to salinity tolerance in poplar. J Plant Growth
Regul 21:224–233
Chen SL, Li JK, Fritz E, Wang SS, Huttermann A (2002b) Sodium
and chloride distribution in roots and transport in three poplar
genotypes under increasing NaCl stress. For Ecol Manage
168:217–230
Chen SL, Li JK, Wang SS, Fritz E, Huttermann A, Altman A (2003)
Effects of NaCl on shoot growth, transpiration, ion compart-
mentation, and transport in regenerated plants of Populus
euphratica and Populus tomentosa. Can J For Res 33:967–975
Cheong Y, Chang H, Gupta R, Wang X, Zhu T, Luan S (2002)
Transcriptional profiling reveals novel interactions between
wounding, pathogen, abiotic stress, and hormonal responses in
Arabidopsis. Plant Physiol 129:661–677
Czarnecka-Verner E, Yuan CX, Scharf KD, Englich G, Gurley WB
(2000) Plants contain a novel multi-member class of heat shock
factors without transcriptional activator potential. Plant Mol Biol
43:459–471
Czarnecka-Verner E, Pan S, Salem T, Gurley WB (2004) Plant class
B HSFs inhibit transcription and exhibit affinity for TFIIB and
TBP. Plant Mol Biol 56:57–75
Damberger FF, Pelton JG, Harrison CJ, Nelson HC, Wemmer DE
(1994) Solution structure of the DNA-binding domain of the heat
shock transcription factor determined by multidimensional
heteronuclear magnetic resonance spectroscopy. Protein Sci
3:1806–1821
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ, Coutu J,
Shulaev V, Schlauch K, Mittler R (2005) Cytosolic ascorbate
peroxidase 1 is a central component of the reactive oxygen gene
network of Arabidopsis. Plant Cell 17:268–281
Ding MQ, Hou PC, Shen X, Wang MJ, Deng SR, Sun J, Xiao F,
Wang RG, Zhou XY, Lu CF, Zhang DQ, Zheng XJ, Hu ZM,
Chen SL (2010) Salt-induced expression of genes related to Na?/
K? and ROS homeostasis in leaves of salt-resistant and salt-
sensitive poplar species. Plant Mol Biol 73:251–269
Doring P, Treuter E, Kistner C, Lyck R, Chen A, Nover L (2000) The
role of AHA motifs in the activator function of tomato heat stress
transcription factors HsfA1 and HsfA2. Plant Cell 12:265–278
Gyorgyey J, Gartner A, Nemeth K, Magyar Z, Hirt H, Heberle-Bors
E, Dudits D (1991) Alfalfa heat shock genes are differentially
expressed during somatic embryogenesis. Plant Mol Biol
16:999–1007
Harrison CJ, Bohm AA, Nelson HC (1994) Crystal structure of the
DNA binding domain of the heat shock transcription factor.
Science 263:224–227
Hihara Y, Kamei A, Kanehisa M, Kaplan A, Ikeuchi M (2001) DNA
microarray analysis of cyanobacterial gene expression during
acclimation to high light. Plant Cell 13:793–806
Janz D, Behnke K, Schnitzler J-P, Kanawati B, Schmitt-Kopplin P,
Polle A (2010) Pathway analysis of the transcriptome and
metabolome of salt sensitive and tolerant poplar species reveals
evolutionary adaption of stress tolerance mechanisms. BMC
Plant Biol 10:150
Jiang M, Zhang J (2003) Cross-talk between calcium and reactive
oxygen species originated from NADPH oxidase in abscisic
acid-induced antioxidant defence in leaves of maize seedlings.
Plant, Cell Environ 26:929–939
Larson JS, Schuetz TJ, Kingston RE (1988) Activation in vitro of
sequence-specific DNA binding by a human regulatory factor.
Nature 335:372–375
Lee BH, Won SH, Lee HS, Miyao M, Chung W-II, Kim IJ, Jo J
(2000) Expression of the chloroplast-localized small heat shock
protein by oxidative stress in rice. Gene 245:283–290
Leshem Y, Seri L, Levine A (2007) Induction of phosphatidylinositol
3-kinase-mediated endocytosis by salt stress leads to intracellular
production of reactive oxygen species and salt tolerance. Plant J
51:185–197
Li B, Liu H, Sun D, Zhou R (2004) Ca2? and calmodulin modulate
DNA-binding activity of maize heat shock transcription factor
in vitro. Plant Cell Physiol 45:627–634
Plant Cell Tiss Organ Cult (2013) 115:299–308 307
123

Li C, Chen Q, Gao X, Qi B, Chen N, Xu S, Chen J, Wang X (2005)
AtHsfA2 modulates expression of stress responsive genes and
enhances tolerance to heat and oxidative stress in Arabidopsis.
Sci China (Ser C, Life Sci) 48:540–550
Liu NY, Ko SS, Yeh KC, Charng YY (2006) Isolation and
characterization of tomato Hsa32 encoding a novel heat-shock
protein. Plant Sci 170:976–985
Liu HC, Liao HT, Charng YY (2011) The role of class A1 heat shock
factors (HSFA1s) in response to heat and other stresses in
Arabidopsis. Plant, Cell Environ 34:738–751
Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses: an
overview. Arch Biochem Biophys 444:139–158
Milioni D, Franz G, Sung R, Hatzopoulos P (2001) Gene expression
during heat-shock in embryogenic carrot cell lines. Plant Cell,
Tissue Organ Cult 65:221–228
Miller G, Mittler R (2006) Could heat shock transcription factors
function as hydrogen peroxide sensors in plants? Ann Bot
98:279–288
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bio-assays with tobacco tissue cultures. Physiol Plant 15:473–497
Nover L, Scharf KD, Gagliardi D, Vergne P, Czarnecka-Verner E,
Gurley WB (1996) The Hsf world: classification and properties
of plant heat stress transcription factors. Cell Stress Chaperon
1:215–223
Nover L, Bharti K, Doring P, Mishra SK, Ganguli A, Scharf KD
(2001) Arabidopsis and the heat stress transcription factor world:
how many heat stress transcription factors do we need? Cell
Stress Chaperon 6:177–189
Ogawa D, Yamaguchi K, Nishiuchi T (2007) High-level overexpres-
sion of the Arabidopsis HsfA2 gene confers not only increased
themotolerance but also salt/osmotic stress tolerance and
enhanced callus growth. J Exp Bot 58:3373–3383
Ottow EA, Brinker M, Teichmann T, Fritz E, Kaiser W, Brosche M,
Kangasjarvi J, Jiang XN, Polle A (2005) Populus euphratica
displays apoplastic sodium accumulation, osmotic adjustment by
decreases in calcium and soluble carbohydrates, and develops
leaf succulence under salt stress. Plant Physiol 139:1762–1772
Panchuk I, Volkov R, Schoffl F (2002) Heat stress- and heat shock
transcription factor-dependent expression and activity of ascor-
bate peroxidase in Arabidopsis. Plant Physiol 129:838–853
Rabindran S, Haroun R, Clos J, Wisniewski J, Wu C (1993)
Regulation of heat-shock factor trimer formation: role of the
conserved leucine zipper. Science 259:230–234
Rizhsky L, Davletova S, Liang H, Mittler R (2004) The zinc finger
protein Zat12 is required for cytosolic ascorbate peroxidase 1
expression during oxidative stress in Arabidopsis. J Biol Chem
279:11736–11743
Rossel JB, Wilson IW, Pogson BJ (2002) Global changes in gene
expression in response to high light in Arabidopsis. Plant Physiol
130:1109–1120
Sabehat A, Lurie S, Weiss D (1998) Expression of small heat-shock
proteins at low temperatures—a possible role in protecting
against chilling injuries. Plant Physiol 117:651–658
Schultheiss J, Kunert O, Gase U, Scharf KD, Nover L, Ruterjans H
(1996) Solution structure of the DNA-binding domain of the
tomato heat-stress transcription factor HSF24. Eur J Biochem
236:911–921
Sun WN, Bernard C, van de Cotte B, Van Montagu M, Verbruggen N
(2001) At-HSP17.6A, encoding a small heat-shock protein in
Arabidopsis, can enhance osmotolerance upon overexpression.
Plant J 27:407–415
Sun J, Chen SL, Dai SX, Wang RG, Li NY, Shen X, Zhou XY, Lu
CF, Zheng XJ, Hu ZM, Zhang ZK, Song J, Xu Y (2009a) NaCl-
induced alternations of cellular and tissue ion fluxes in roots of
salt-resistant and salt-sensitive poplar species. Plant Physiol
149:1141–1153
Sun J, Dai SX, Wang RG, Chen SL, Li NY, Zhou XY, Lu CF, Shen
X, Zheng XJ, Hu ZM, Zhang ZK, Song J, Xu Y (2009b) Calcium
mediates root K?/Na? homeostasis in poplar species differing in
salt tolerance. Tree Physiol 29:1175–1186
Sun J, Li LS, Liu MQ, Wang MJ, Ding MQ, Deng SR, Lu CF, Zhou
XY, Shen X, Zheng XJ, Chen SL (2010a) Hydrogen peroxide
and nitric oxide mediate K?/Na? homeostasis and antioxidant
defense in NaCl-stressed callus cells of two contrasting poplars.
Plant Cell, Tissue Organ Cult 103:205–215
Sun J, Wang MJ, Ding MQ, Deng SR, Liu MQ, Lu CF, Zhou XY,
Shen X, Zheng XJ, Zhang ZK, Song J, Hu ZM, Xu Y, Chen SL
(2010b) H2O2 and cytosolic Ca2? signals triggered by the PM
H?-coupled transport system mediate K?/Na? homeostasis in
NaCl-stressed Populus euphratica cells. Plant, Cell Environ
33:943–958
Sun J, Zhang X, Deng S, Zhang C, Wang M, Ding M, Zhao R, Shen
X, Zhou X, Lu C, Chen S (2012) Extracellular ATP signaling is
mediated by H2O2 and cytosolic Ca2? in the salt response of
Populus euphratica cells. PLoS ONE 7:e53136
Swindell WR, Huebner M, Weber AP (2007) Transcriptional profiling
of Arabidopsis heat shock proteins and transcription factors
reveals extensive overlap between heat and non-heat stress
response pathways. BMC Genomics 8:125
Volkov RA, Panchuk II, Mullineaux PM, Schoffl F (2006) Heat
stress-induced H2O2 is required for effective expression of heat
shock genes in Arabidopsis. Plant Mol Biol 61:733–746
Vuister GW, Kim SJ, Wu C, Bax A (1994) NMR evidence for
similarities between the DNA-binding regions of Drosophila
melanogaster heat shock factor and the helix-turn-helix and
HNF-3/forkhead families of transcription factors. Biochemistry
33:10–16
Wang RG, Chen SL, Deng L, Fritz E, Huttermann A, Polle A (2007)
Leaf photosynthesis, fluorescence response to salinity and the
relevance to chloroplast salt compartmentation and anti-oxida-
tive stress in two poplars. Trees 21:581–591
Wang RG, Chen SL, Zhou XY, Shen X, Deng L, Zhu HJ, Shao J, Shi
Y, Dai SX, Fritz E, Huttermann A, Polle A (2008) Ionic
homeostasis and reactive oxygen species control in leaves and
xylem sap of two poplars subjected to NaCl stress. Tree Physiol
28:947–957
Wang F, Deng S, Ding M, Sun J, Wang M, Zhu H, Han Y, Shen Z,
Jing X, Zhang F, Hu Y, Shen X, Chen SL (2013) Overexpression
of a poplar two-pore K? channel enhances salinity tolerance in
tobacco cells. Plant Cell, Tissue Organ Cult 112:19–31
Wu QY, Lin J, Liu JZ, Wang XF, Lim W, Oh M, Park J, Rajashekar
CB, Whitham SA, Cheng NH, Hirschi KD, Park S (2012)
Ectopic expression of Arabidopsis glutaredoxin AtGRXS17
enhances thermotolerance in tomato. Plant Biotechnol J
10(8):945–955
Zhang F, Wang Y, Wang D (2007) Role of nitric oxide and hydrogen
peroxide during the salt resistance response. Plant Signal Behav
2:473–474
Zhong M, Orosz A, Wu C (1998) Direct sensing of heat shock and
oxidation by Drosophila heat shock transcription factor. Mol
Cell 2:101–108
308 Plant Cell Tiss Organ Cult (2013) 115:299–308
123










![Overexpression of AtMYB44 Enhances Stomatal Closure to ... · Overexpression of AtMYB44 Enhances Stomatal Closure to Confer Abiotic Stress Tolerance in Transgenic Arabidopsis1[C][W][OA]](https://img.pdfslide.us/doc/110x75/5cac6d4e88c993220b8be0a0/overexpression-of-atmyb44-enhances-stomatal-closure-to-overexpression-of.jpg)


















