Embed Size (px)
Citation preview

Outline
•A Biological Perspective
•The Cell
•The Cell Cycle
•Modeling
•Mathematicians I have known

Cancer
Heart disease
Neurodegenerative illnesses
Molecular Basis of Disease


If we can understand disruption of molecular events at the cellular level we can perhaps prevent or stop disease manifestation at the
organismal level

The Cell

A variety of membrane-bounded compartments exist within eucaryotic cells, each specialized to perform a different function.

DNA•Information is contained in the primary structure (the sequence of bases).
Protein•Information is contained at multiple structural levels (primary, secondary, tertiary, quaternary)
Forms of Biological Information
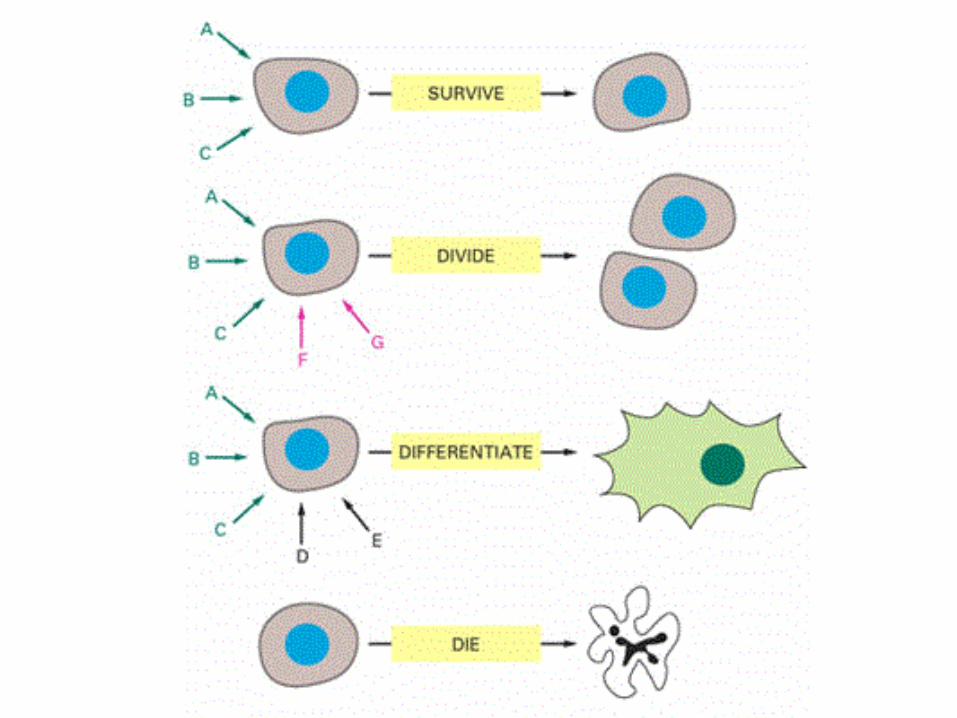




Two processes must alternate during eukaryotic cell division
•Genome must be replicated in S phase
•Genome must be halved during M phase
The Cell Cycle


Cell cycle events must be highly regulated in a temporal manner

Genetic and molecular studies in diverse biological systems have resulted in identification and characterization of the cell cycle machinery

Mitotic spindle
DNA replication Chiasmata
Dynamic instabilty
Cdc mutantsCell-cycle controlMaturation-promoting factor
Regulation of Cdc2
Cyclin characterization
Checkpoint controlp53
The mitotic checkpointThe APC and proteolysis SCF and F-box proteins
The restriction point
Yeast centromeresCell-cycle conservation
Replication originsRetinoblastoma/E2FBody-plan regulationA new class of cyclinsCDK inhibitors
Sister-chromatid cohesion


The cell cycle engines
•Cyclin Dependent Kinases (CDKs)
CD
K
cyclin
substrate
ATP
P
product + ADP
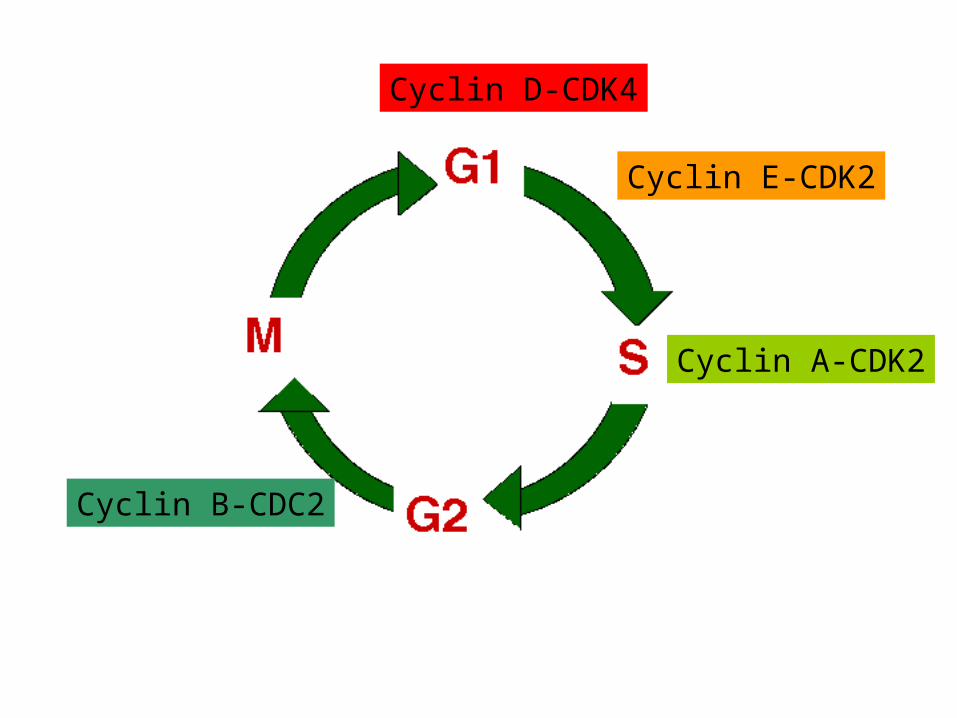
Cyclin D-CDK4
Cyclin E-CDK2
Cyclin A-CDK2
Cyclin B-CDC2

CDK activity

asyn 0 4 8 12 14 16 20 24 28 32 hours
cyclin A
cyclin E
Cdk2
Cyclin and CDK expression as cells re-enter the cell cycle
G0 G1 S cell cycle phases

G1
S
G2/M
cyclin Eprotein
Cdk2 boundto cyclin E
cyclin Eassociatedkinase activity
1 2 3 4 5 6 7 8


Cyclin D-CDK4
Cyclin E-CDK2
Cyclin A-CDK2
Cyclin B-CDC2
CDK inhibitors



The Cell Cycle
•Complex system
•Components are identified
•Highly regulated
•Defined parameters

Cell Cycle Characteristics
•Temporally ordered events
•Irreversibility
•Oscillations
•Checkpoints
•Positive and negative feedback loops

Positive Feedback Loop

70kg human ~ 1013 cells

Overall properties not predictable from what is known about
constituent parts
Complexity

Reductionist-analytical strategies focus on component properties and actions, but do not necessarily describe dynamic behavior
of the larger system.

The best test of our understanding of cells will be to make quantitative predictions about their behavior and test them. This will require
detailed simulations of the biochemical processes taking place within cells…
Hartwell, Hopfield, Leibler, and Murray

What’s the problem?
•Cartoons are cartoons
•They do not quantitatively describe the experimental data they summarize
•Used in a loose qualitative manner
•Informal, verbal
•Not reliable for judging accuracy of mechanistic proposals

•Notion of mathematical modeling adding value to standard approaches
•Help to formalize and predict behavior, suggest experiments
Bioessays 24, 2002.
Can Mathematical Modeling Help?

•Start from a grocery list of parts
•Break down large scale systems into smaller functional modules
•Simulate steady states, oscillations, sharp transitions
Modeling the Cell Cycle

•Formulate interactions as precise molecular mechanisms.
•Convert the mechanism into a set of nonlinear ordinary differential equations.
•Study the solutions of the differential equations by numerical simulation.
•Use bifurcation theory to uncover the dynamical principles of control systems.

Cells progressing through the cell cycle must commit irreversibly to mitosis.

•What causes cyclin degradation to turn on and off periodically?
•Why don’t rates of synthesis and degradation balance each other?
•There must be some mechanism for switching irreversibly between phases of net cyclin synthesis and net cyclin degradation.
Questions

•Many competing models because the degrees of freedom were unbounded.
•Could occur by hysteresis (ie toggle-like switching behavior in a dynamical system).
•Time delayed negative feedback loops.
Models, models, everywhere

vs

•Describes a network of interlocking positive and negative feedback loops controlling cell cycle progression.
•Proposes a bistable switch is created by the positive feedback loops involving cyclin B-cdc2 and its regulatory proteins.
The Hysteresis Model of Novak and Tyson

•It takes more of something to push a system from state A to B than it does to keep the system in B.
•Creates a bistable system with a rachet to prevent slippage backwards.
•Irreversibility was proposed to arise on transversing a hysteresis loop
Hysteresis

•Using Xenopus egg extracts to demonstrate the cell cycle exhibits hysteresis
•The amount of cyclin required to induce entry into mitosis is larger than the amount of cyclin needed to keep the extract in mitosis.
Experimental System
Need pic of xenopus

Steady state cdc2 kinase activity as a function of [cyclin]
Black dots=experimental
Gray dots=proposed
Ti=inactivation threshold
Ta=activation threshold

The hysteresis model made nonintuitive predications that were confirmed
experimentally.
• [cyclin B] to drive mitosis > [cyclin B] to stay in mitosis.
• Unreplicated DNA elevates the cyclin B threshold for cdc2 activation; ie checkpoints enlarge the hysteresis loop.
• Cdc2 activation slows down at cyclin B concentrations marginally above the threshold.

Mathematicians I have known

Cyclin D-CDK4
Cyclin E-CDK2
Cyclin A-CDK2
Cyclin B-CDC2
p27kip1

CD
Kcyclin
substrate
ATP
P
product + ADP
p27kip1
CD
K
cyclin Inhibited
p27kip1
Model of p27kip1 Function

wtp27 p27(87-198)
cyclin E
wt
87-198
- + - +
Cyclin E-CDK2 can phosphorylate p27kip1

50 10 15 20
time (min)
No pre-incubation
p27 pre-incubation
phos. p27
time
p27
No pre-incubation p27 pre-incubation
time

E-K2 + p27 E-K2-p27 (E-K2-p27)*
rapid slow
loose binding tight binding
Two Distinct Binding Modes between p27 and Cyclin E-Cdk2
rapid

K2E
p27
ATP
(K2E-p27)loose
(K2E-ATP)
(K2E-p27-ATP)loose
ATP
p27
(K2E-p27)tight
K2E-p27P + ADPK2E + p27P
Slow
Fast
InhibitoryInteraction
CatalyticCycle

20151050
0
1000
2000
3000
4000
5000 1000 μM ATP
500 μM ATP
150 μM ATP
50 μM ATP
phos. HH1
Increasing [ATP] Drives p27 Phosphorylation
P27-P
Time (min)

Inhibited
p27kip1
CD
K
cyclin Switch
Active
CD
K
cyclin p27kip1
P
p27kip1
P
Cell cycleprogression
Switching between Inhibitor and substrate functions

Mathematical analysis of binary activation of a cell cycle kinase which
down-regulates its own inhibitorC.D. Thron

•P27 binds and inhibits cyclin E-CDK2
•Cyclin E-CDK2 phosphorylates and deactivates p27
•This creates a positive feedback loop
Experimental Observations

Is the release of EK2 binary (all-or-none)?
•Binary enzyme activation implies an abrupt switch from a stable steady state with a low level of free active enzyme.
•Implies a bistable system. Small parameter change causes low activity steady state to be extinguished in a saddle-node bifurcation.
•Mathematical analysis of the biochemical kinetics required for binary activation.

Conclusions
An enzyme that attacks and deactivates its own inhibitor is not released from inhibitor
binding in an all-or-none fashion unless certain kinetic features are present.

•If you want to communicate with someone, you need to speak their language
•Convert math to cartoons
•Seek out collaborations/sabbaticals
•The burden of proof is on you
•Look outward as well as inward (kinetics and physiology)
You say tomato, I say tomahto





























