Embed Size (px)
Citation preview

HERPETOZOA 11 (1/2): 47 - 70Wien, 30. Juli 1998
Osteology of the Pyrenean Mountain Lizardsand comparison with other species
of the collective genus Archaeolacerta MERTENS, 1921 S. 1.from Europe and Asia Minor
(Squamata: Sauna: Lacertidae)
Osteologie der Pyrenäen-Gebirgseidechsen und Vergleich mit anderen Arten derSammelgattung Archaeolacerta MERTENS, 1921 S. 1. aus Europa und Kleinasien
(Squamata: Sauria: Lacertidae)
OSCAR J. ARRIBAS
KURZFASSUNG
Die Pyrenäenarten von Archaeolacerta s. 1. (A. aranica, A. aurelioi und A. bonnali) unterscheiden sich vonden übrigen Arten der Eurasiatischen Radiationslinie (sensu MAYER & BENYR 1994) durch den Besitz knöchernerRippen im Bereich des dritten Präsakralwirbels. Die drei Pyrenäenarten zeigen in der Morphologie von Postorbitaleund Postfrontale jeweils osteologische Verschiedenheiten, die Artniveau erreichen und ihre zweifelsfreie Unterschei-dung erlauben. Auch finden sich Unterscheidungsmerkmale im Schultergürtel (Klavikel und Stemalfontanelle).
Apomorphe Merkmale innerhalb der kaukasischen 'Lacerta' saxicola - Gruppe stellen deutliche Unterschiedesowohl gegenüber den Archaeolacerten Europas (fast immer ein zusätzlicher Präsakralwirbel) als auch gegenüberApathya (ein zusätzlicher Präsakralwirbel und eine verminderte Anzahl von Prämaxillarzähnen) dar. 'L. ' chlorogasterunterscheidet sich von den übrigen kaukasischen Formen klar durch die Zahl der Prämaxillarzahne und Form der In-terklavikel. Die parthenogenetischen Taxa zeigen deutliche Ähnlichkeiten (Modalwerte der Wirbelzahl, Morphologiedes Postorbitale) zu ihren Eltemarten, besonders zur weiblichen Elternart.
Aufgrund der osteologischen Befunde wird deutlich, daß die europäischen, vorläufig in der SammelgattungArchaeolacerta s. 1. vereinigten Taxa keine natürliche Einheit darstellen. Die Formen der Pyrenäen und des Kaukasuserscheinen osteologisch deutlich differenziert, während die iberischen, balkanischen und tyrrhenischen Taxa in denuntersuchten Strukturen nur synplesiomorphe Merkmalsausprägungen gemeinsam haben, die keine Aussagen überihre Verwandtschaft ermöglichen.
ABSTRACT
The Pyrenean species of Archaeolacerta s. 1. (A. aranica, A. aurelioi and A. bonnali) are different from theother species of the Eurasian radiation (sensu MAYER & BENYR 1994) by the possession of bony ribs associated withthe third presacrai vertebra. The three Pyrenean species can be unequivocally distinguished at a species level by themorphology of the postorbital and postfrontal bones. Other distinctive character states are found in the pectoral girdle(clavicle and sternal fontanelle).
In their apomorphic characters, the Caucasian taxa of the 'L.' saxicola group present clear differences whencompared to the Archaeolacertae of Europe (almost always an extra presacrai vertebra) and to Apathya (an extra pre-sacral vertebra and a minor number of premaxillary teeth). 'L.' chlorogaster appears clearly differentiated from theother taxa of the Caucasian group by the number of premaxillary teeth and shape of interclavicle. The parthenogeneticforms present characteristic features (modal numbers of vertebrae, postorbital bone morphology) of their original pa-rental species, the female parental species in particular.
In view of the osteological results, it becomes evident that the European species currently assigned to the col-lective genus Archaeolacerta s. I. do not constitute a natural cluster. The Pyrenean and Caucasian species are oste-ological ly clearly differentiated while among the structures studied in the Iberian, Balcanic and Tyrrhenian taxa onlysymplesiomorphic character states were found which do not permit conclusions on their relationship.
KEY WORDS
Squamata: Sauria: Lacertidae; Archaeolacerta, A. aranica, A. aurelioi, A. bonnali, A. monticola, A. cyreni,Lacerta, Pyrenees, Iberian Peninsula, Europe, Caucasus, Asia Minor, osteology, phylogeny.
INTRODUCTION
Bibliography on lacertid osteology is tile families of similar or even smallerscarce when compared to the number of range (BARBADILLO & SANZ 1983). Apartosteological papers dedicated to other rep- from classic works on general osteology of
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

48 O S C A R J.ARRTOAS
the reptiles (e.g. CAMP 1923; ROMER 1956),comparatively little attention has been de-voted to lacertid osteological variation oftaxonomic relevance.
Standard reference publication forlacertid skull morphology is the detailedwork by MÉHELY (1909) which pays specialattention to the form of the postorbital andpostfrontal region and the number of pre-maxillary, maxillary, and dentary teeth.KLEMMER'S (1957) paper highlights sexualdimorphism in the development of theskull osteoderms (crusta calcarea) and des-cribes different degrees of platycephalyfound in the various species as a functionof their more or less saxicolous habitat.The outstanding publications of SIEBEN-ROCK (1894), KÜHNE (1913), and HOFF-STETTER & GASC (1969) deal with the ver-tebral column while the publication ofETHERIDGE (1967) concentrates on the cau-dal region. More recently, ARNOLD (1973,1989) reviewed a good number of osteo-logical characters for a multitude of species(many of which find their first and uniqueosteological reference there). Together withother morphological information he util-
ized these character states for the construc-tion of lacertid phylogenies and for defini-tion and differential diagnosis of somegenera. Further references concerning oste-ological characters were recently publishedin PÉREZ-MELLADO & al. (1993) and ARRI-BAS(1993, 1994, 1996).
In the present paper the author in-tends to (i) indicate osteological charactersof diagnostic relevance for the three Pyre-nean species of Archaeolacerta s. 1. (= Ar-chaeolacertae); (ii) compare the Pyreneanspecies with Archaeolacerta s. 1. speciesfrom Spain and Europe concerning thestates of their osteological characters; (iii)study Caucasian species of the 'L.' saxicolaEVERSMANN, 1834 group, usually assignedto Archaeolacerta (ARNOLD 1973, 1989;DAREVSKY 1967, 1972; MAYER & BENYR1994) or Apathya (BISCHOFF 1991); thepresent paper submits first osteologicaldata for a number of species; (iv) obtain asound series of osteological characters forfuture phylogenetic analyses; (v) evaluatevarious osteological characters used for thereconstruction of lacertid phylogenies.
MATERIAL AND METHODS
Specimens studied (table 1)
As was done by ARRIBAS (1996), in thepresent paper the European species studiedare provisionally assigned to the genus Ar-chaeolacerta in a wide sense. Species ofuncertain generic status, like the Caucasianones are referred to as 'Lacerta' sens. lat.For generic nomenclature of other taxa ofthe Eurasian radiation, the proposal of AR-NOLD (1973, 1989), updated by MAYER &BISCHOFF (1996) is followed. Apathya istreated as a full genus due to its clear mor-phological differences to the Caucasianspecies of the 'L.' saxicola group and theother nominal lacertid genera recognizeduntil now. Lacerta andreansky is still as-signed to 'Lacerta' of unclear taxonomicposition although MAYER & BISCHOFF(1996) classified it as belonging to the ge-nus Teira.
The following taxa which are clearlydifferent from Archaeolacerta s. 1. were in-cluded in this study for comparison: Lacer-ta agilis LINNAEUS, 1758, L bilineata DAU-
DIN, 1802, L. schreiben BEDRIAGA, 1878,Zootoca vivipara (JACQUIN, 1787 (ovi-parous form), Podarcis muralis (LAURENTI,1768), P. hispanica STEINDACHNER, 1870,P. bocagei SEOANE, 1884, P. pityusensisBoscÂ, 1883, P. lilfordi (GÜNTHER, 1874),Algyroides marchi VAL VERDE, 1958, Teiradugesii MILNE-EDWARDS, 1829, 'Lacerta*andreanskyi WERNER, 1929 and Timon lepi-dus (DAUDIN, 1802). Gallotia galloti (Du-MÉRIL & BiBRON, 1839) and Psammodro-mtnus algirus (LINNAEUS, 1758), belongingto the subfamily Gallotiinae MAYER &BENYR, 1994 were used as outgroup taxa inthe discussion of the osteological charac-ters.
Methods
X-ray photographs were taken with aPhilips Super 100 cp ® apparatus. Best re-sults were obtained at 40 kV, 4 mA/sec,and 19 milliseconds time of exposure. X-ray photographs were used to study thenumber of presacrai vertebrae without dis-
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
Osteology of Archaeolacerta s. 1. 49
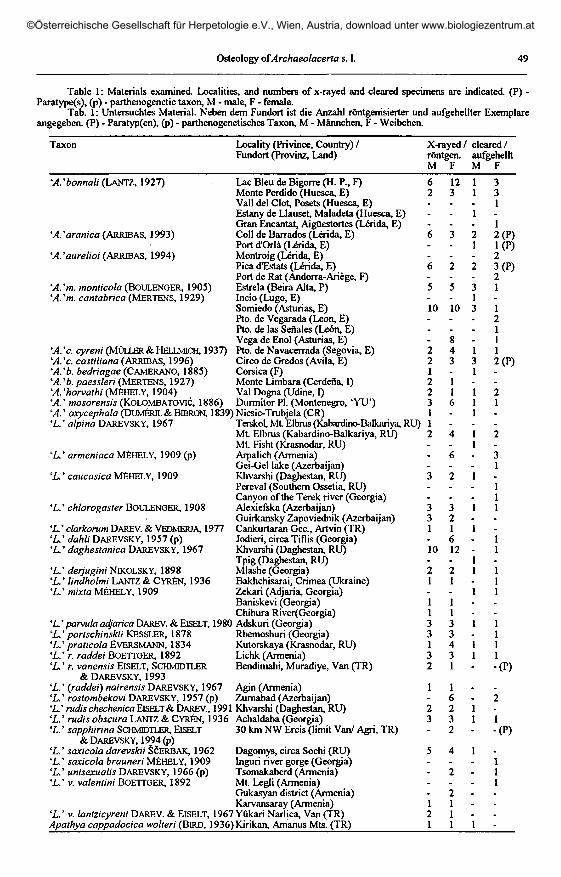
Table 1 : Materials examined. Localities, and numbers of x-rayed and cleared specimens are indicated. (P) -Paratype(s), (p) - parthenogenetic taxon, M - male, F - female.
Tab. 1 : Untersuchtes Material. Neben dem Fundort ist die Anzahl röntgenisierter und aufgehellter Exemplareangegeben. (P) - Paratyp(en), (p) - parthenogenetisches Taxon, M - Männchen, F - Weibchen.
Taxon Locality (Privince, Country) /Fundort (Provinz, Land)
X-rayed/ cleared/röntgen, aufgehelltM F M F
1927)
'A.1 aranica (ARRIBAS, 1993)
'A'awre/Zo/(ARRIBAS, 1994)
'A.'m. monticola (BOULENGER, 1905)'A.'m. cantabrica (MERTENS, 1929)
'A. 'c. cyreni (MÜLLER & HELLMICH, 1937)cA.'c. castiliana (ARRIBAS, 1996)lA:b. bedriagae (CAMERANO, 1885)'A. 'b. paessleri (MERTENS, 1927)'A. 'horvathi (MÉHELY, 1904)' A ' mosorensis (KOLOMBATOVIC, 1886)
Lac Bleu de Bigorre (H. P., F)Monte Perdido (Huesca, E)Vail del Clot, Posets (Huesca, E)Estany de Llauset, Maladeta (Huesca, E)Gran Encantat, AigOestortes (Lérida, E)Coli de Barrados (Lérida, E)Port d'Orlà (Lérida, E)Montroig (Lérida, E)Pica d'Estats (Lérida, E)Port de Rat (Andorra-Ariège, F)Estrela (Beira Alta, P)Incio (Lugo, E)Somiedo (Asturias, E)Pto. de Vegarada (Leon, E)Pto. de las Sefiales (Leon, E)Vega de Enol (Asturias, E)Pto. de Navacerrada (Segovia, E)Circo de Gredos (Avila, E)Corsica (F)Monte Limbara (Cerdefia, I)Val Dogna (Udine, I)Durmitor PI. (Montenegro, 'YU')
' A ' oxycephala (DUMÉRIL& BlBRON, 1839)Nicsic-Trubjela (CR)'L.' alpina DAREVSKY, 1967 Terskol, Mt Elbrus (Kabardino-Balkariya, RU)
ML Elbrus (Kabardino-Balkariya, RU)Mt. Fisht (Krasnodar, RU)Arpalich (Armenia)Gei-Gel lake (Azerbaijan)Khvarshi (Daghestan, RU)Pereval (Southern Ossetia, RU)Canyon of the Terek river (Georgia)Alexiefska (Azerbaijan)Guirkansky Zapoviednik (Azerbaijan)Cankurtaran Gee, Artvin (TR)Jodieri, circa Tiflis (Georgia)Khvarshi (Daghestan, RU)Tpig (Daghestan, RU)Mlashe (Georgia)Bakhchisarai, Crimea (Ukraine)Zekari (Adjaria, Georgia)Baniskevi (Georgia)Chihura River(Georgia)
'pannila adjarica DAREV. & ElSELT, 1980 Adskuri (Georgia)
'L.' armeniaca MÉHELY, 1909 (p)
'L.' caucasico MÉHELY, 1909
'Z,.' chlorogaster BOULENGER, 1908
'L' clarkorum DAREV. & VEDMERJA, 1977'L.' dahli DAREVSKY, 1957 (p)'L.' daghestanica DAREVSKY, 1967
'L.' derjugini NDCOLSKY, 1898'L.' lindholmi LANTZ & CYRÉN, 1936'L.' mixta MÉHELY, 1909
'Z,.' portschinskii KESSLER, 1878'Z,.'praticola EVERSMANN, 1834'Z,.' r. raddei BOETTGER, 1892'Z..' r. vanensis ElSELT, SCHMIDTLER
& DAREVSKY, 1993'Z,.' (raddei) nairensis DAREVSKY, 1967
rostombekovi DAREVSKY, 1957 (p)
Rbemoshuri (Georgia)Kutorskaya (Krasnodar, RU)Lichk (Armenia)Bendimahi, Muradiye, Van (TR)
Agin (Armenia)Zurnabad (Azerbaijan)
'L.' rudis chechenica ElSELT & DAREV., 1991 Khvarshi (Daghestan, RU)'L. ' rudis obscura LANTZ & CYRÉN, 1936 Achaldaba (Georgia)'L.' sapphirina SCHMIDTLER, ElSELT
& DAREVSKY, 1994 (p)'L.' saxicola darevskii SÒERBAK, 1962'L.' saxicola brauneri MÉHELY, 1909'Z,.' unisexualis DAREVSKY, 1966 (p)'Z,.' v. valentini BOETTGER, 1892
30 km NW Ercis (limit Van/ Agri, TR)
Dagomys, circa Sochi (RU)Inguri river gorge (Georgia)Tsomakaberd (Armenia)Mt. Legli (Armenia)Gukasyan district (Armenia)Karvansaray (Armenia)
'L.' v. lantzicyreni DAREV. & ElSELT, 1967 Yûkari Narlica, Van (TR)Apathya cappadocica wolteri (BIRD, 1936)Kirikan, Amanus Mts. (TR)
62
6-
6-5.10
.221223112.--3
-331.10.21.1133132
1.23-
5-
.121
123
3-
2-5.10
.843.116
4.6.2
321612.21.1133431
16232
4
2.2111
11.1.21-2-313
.131.111.11
1
1.1
11.1
1.11-
11-
1
1
331.12(P)1(P)23(P)21.121112(P)
.21
2.31.111.
11.111
-1111-(P)
.2.1"(P)
.111
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

50 OSCAR J. ARRTOAS
secting the specimens, as demonstrated byseveral authors (e. g. ARNOLD 1973).
Most specimens were preserved in 70%ethanol and thereafter cleared by means ofpotash lye (KOH, 1%). Bones were stainedwith alizarine red and transferred to glycer-ine for permanent storage (TAYLOR 1967;DURFORT 1978).
Characters studied
Skull characters: (i) shape of the na-sal process of premaxilla, (ii) number ofpremaxillary teeth, (iii) number of maxil-lary teeth, (iv) number of dentary teeth, (v)shape of margo ocularis of maxilla ("stepedjugal" sensu ARNOLD 1989), (vi) conditionof antero-distal process of the postfrontal,(vii) condition of antero-medial process of
the postorbital, (viii) length ratio of post-orbital and postfrontal, (ix) degree of over-lap of postorbital and squamosal, (x) con-dition of postorbital and postfrontal (fusedor separated) at birth.
Postcranial characters: (xi) presenceof visible (ossified) ribs associated withthird presacral vertebra, (xii) shape of ribsassociated with sixth presacral vertebra,(xiii) sternal costal formula, (xiv) numberof presacral vertebrae, (xv) number ofshort-ribbed posterior presacral vertebrae,(xvi) expression and shape of sternal fon-tanelle, (xvii) shape (emarginate or not) ofclavicle, (xviii) shape of interclavicle, (xix)comparative lengths of interclavicularbranches, (xx) morphology of first preauto-tomic tail vertebrae (see ARNOLD 1973,1989).
RESULTS AND DISCUSION
SKULL CHARACTERS
Shape of nasal processof premaxilla
In all Pyrenean species, the nasal pro-cess, extending from the premaxillary to-wards the nasal bones, is of elongate undif-ferentiated form in that its lateral marginsare parallel. In the oldest specimens itsometimes appears spoon-shaped due to in-creased growth in the distal part above thenasals.
In the other closely related Iberianspecies, the nasal process can be narrow atits base and arrow-shaped in the apex. Thissituation is predominant in A. cyreni, andalso appears frequently in A. monticolamonticola and some Gallician specimens ofA. monticola cantabrica (in one studiedspecimen from Incio, Lugo). Specimensstudied from the other parts of the Cantab-rian Mountains present undifferentiatedand slender nasal processes, equaling thePyrenean taxa. All other European speciesof Archaeolacerta as well as of the 'Z,.'saxicola group and Apathya cappadocicapresent nasal processes of undifferentiatedshape, as the Pyrenean species.
From all other taxa studied, only 77-mon lepidus possesses an arrow-shaped na-sal process, while in Podarcis, Teira, Algy-roides, Lacerta (s. Str.), Zootoca, Psammo-
dromus and Gallotia nasal processes aresimple and undifferentiated.
The presence of an undifferentiatedelongate nasal process shall be consideredto be the most primitive condition. It isfound in almost all outgroup species, and isthe structurally most simple and wide-spread type in the Eurasian lacertid radia-tion (sensu MAYER & BENYR 1994).
Numberof premaxillary teeth
It is difficult to determine the primi-tive number of premaxillary teeth per sidein Lacertidae. The two most common num-bers, 7 and 9, appear more or less mingledamong the members of the Eurasian radia-tion. In the outgroup, both numbers appear(7 in Gallotia, 9 in Psammodromus). InAcanthodactylus, a member of the Africanradiation, the usual number is 7, while inLacerta s. str. is 9, and 7 again in Zootoca.
The three Pyrenean species usuallydevelop seven premaxillary teeth. In A. mon-ticola cantabrica, this number is omni-present in all Cantabrian specimens studied(however, PÉREZ-MELLADO & al., 1993indicated 9 and 10 as the number of pre-maxillary teeth in this species!), while 7 or8 are typical to A. monticola monticola.Archaeolacerta cyreni usually presents 8 or9 premaxillary teeth, rarely 7.
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Osteology of Archaeolacerta s. 1. 51
The number of 9 is found in A. bed-riagae (9) and in A. mosorensis (8 or 9),while A. horvathi and A. oxycephala have7 each. According to MÉHELY (1909), Amosorensis also possesses 7 teeth, which isin discrepancy to the data of SIEBENROCK(1894), KLEMMER (1957), and of my own.The species of the 'Z..1 saxicola group usu-ally have 7 teeth, rarely 8, except 'Z,.1 der-jugini (8) and the uncertain 'L' chlorogas-ter which shows 9 or 10. Apathya cappa-docica also has 10.
Number of maxillary teeth
In A bonnali there are 15 to 18 teethin each maxilla (x = 15,7; stD = 0,9, n =19), in A aranica 14 to 17 (x= 15,41; stD= 1,18, n = 12) and in A aurelioi 15 to 17(x= 15.95; stD = 0.66, n = 20). Other Ibe-rian species show numbers just sjighty su-perior, as A monticola (15 - 18, x= 16.41,n = 26), or clearly superior, as A cyreni(16 - 20, x = 18.07, n = 14). In the otherspecies, numbers appear rather variableand are usually higher than those found inthe Pyrenean taxa.
Number of dentary teeth
The three Pyrenean species displayvery similar values, which are relativelylow within the species studied Archaeo-lacerta bonnali presents 17 to 21 (x =19.05; stD = 1.20, n = 20). In A aranicathey number from 16 to 20 (x = 18.58; stD= 1.03, n = 12) and in A aurelioi from 16to 22 (x = 19.66, stD = 1.94, n = 18). Thelowest values are found in A aranica,which proportionately has a slightly shorterhead and therefore shorter maxilla anddentary.
Other closely related species, as Amonticola show slightly higher values (17 -23, x = 20.53, stD = 1.49, n = 20), orclearly superior ones like A cyreni (20 -24, x = 22.07, stD = 1.03, n = 16). Theother European species of Archaeolacertaalso have higher numbers (21 - 23), exceptA horvathi which is very similar to the Py-renean species in this respect. The 'L' saxi-cola group presents variable values: insmall species, like 'Z,.1 derjugini, 'Z,.1 dagh-estanica and 'Z..' rostombekovi (and in thesmall-headed fL.' praticola) numbers of
teeth are as low as in the Pyrenean species.Within the Lacertidae there seems to
be a clear positive relationship between thefinal size of the individual and the numberof teeth in the dentary. Juveniles and adultsof a given species show just small dif-ferences in the number of teeth, withhigher values in adults (see e. g., ROCEK1980). According to differences in allo-metric growth of the male head which isincreased in almost all species, taxa can besubdivided into two types: (i) species inwhich males develop allometrically bigheads (with more teeth), and (ii) species inwhich males develop relatively short orsmall heads (with fewer teeth). Withinclosely related species only one type seemsto occur, indicating that this feature couldbe of some phylogenetic significance. How-ever, tooth numbers are not subject to un-hindered variation in Lacertidae as one cansee in the large Gallotia simonyi (STEIN-DACHNER, 1889) and T. lepidus, in whichthe tooth number does not differ signifi-cantly from that found in smaller species.The numbers of premaxillary and maxil-lary teeth, however, do not seem to be sub-ject to the ontogenetic variation (ROCEK1980).
Shape of maxillary margo ocularis
The shape of the margo ocularis ofthe maxilla ("stepped jugal", sensu AR-NOLD 1989) has been utilized as one char-acter for the reconstruction of phylogenies.According to ARNOLD (1989), the suturebetween maxilla and jugal is clearly sinous("stepped") in Lacerta s. str. and Podarcis,and might represent the derived state of thecaracter. However, in the sample of Ibero-balearic Podarcis, the above character stateis not clearly more pronounced than in therest of the Lacertidae examined. ThereforeI think that the character is of limited valuein phylogenetic reconstructions. In Lacertas. str. the stepped route of the suture isclearly visible.
In the three Pyrenean species studied,the suture between maxilla and jugal israther smoothly stepped, and not very pro-nounced. The same situation is found in allspecies of Archaeolacerta s. 1. (EuropeanArchaeolacertae, 'Z,.' saxicola group andApathya cappadocicd), and in the other
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

52 OSCAR J. ARRTOAS
lacertid species of the Eurasian radiationstudied. Only in Lacerta s.str. this suturewas strongly angulose.
Antero-distal process of postfrontal(plates 1-4)
This process is present in the majorityof the Lacertidae and its lack should beconsidered a derived character state.
Only A. aure Hoi lacks this process (inall specimens studied), with the unique ex-ception of an individual from Andorrawhich is considered an atavistic return tothe primitive character state (plate 2). In A.bonnali,A. aranica, and in all other lacer-tid species included in this study, this proc-ess is present and well developed. Its re-duction and disappearance in A aurelioi isan autapomorphy.
Antero-medial process of postorbital(plates 1-4)
The expression of this process ismore subject to variation than the previouscharacter. The presence of a well developedantero-medial postorbital process indicatesthe primitive character state, as is fre-quently found in all genera studied inwhich the postfrontal and postorbital areseparated from birth on (e. g., most Lac-erta, Podarcis, Archaeolacerta). In somespecies belonging to different phyleticlines, this process is missing or present inreduced form.
Among the Pyrenean species, the an-tero-medial process of the postorbital ap-pears very well developed in A. aranica,while it is absent in both A. bonnali and A.aurelioi. These observations apply to allPyrenean specimens studied and permit - incombination with the previous character -the specific distinction of all Pyrenean Ar-chaeolacertae from an osteological point ofview.
In most other taxa studied here, out-group taxa included, this process is clearlydeveloped, except in some species of the'Z,.' saxicola complex. It frequently lacks in'Z,.1 mixta and 'L' raddei, which are mater-nal species of the initial crossings that ledto the parthenogenetic 'Z,.' armeniaca ('Z,.1
valentini M x 'L' mixta F), 'L' dahli ÇL'portschiskii M x 1 ' mixta F), 'Z,.' rostom-
bekovi ('Z,.' portschinskii Mx'L' raddei F)and 'L.' unisexualis ('Z,.' valentini M x 'Z,.'raddei nairensis F) (UzzELL & DAREVSKY1975; DAREVSKY & DANIELYAN 1977; DA-REVSKY & al. 1985, 1986; KUPRIYANOVA1989; MORITZ & al. 1992). As this processlacks or is extremely reduced in all theseparthenogenetic species, there seems to bedominancy or at least strong maternal in-fluence in the heredability of this character.In this respect it could be enlightening tostudy 'Z,.' uzzelli, with the same parentalspecies like 'L' unisexualis but with 'Z,.'valentini as maternal species.
Lengths of postorbital and postfrontal(plates 1-4)
In general, both bones are more orless subequal in length (primitive characterstate), although there is some secondarydeviation in several species (e.g., in casesof reduced cranial ossification related tosaxicolous life).
In the Pyrenean species A. aurelioiand A. aranica, the postorbital bone some-times clearly surpasses the posterior end ofthe postfrontal. This situation should beconsidered the derived state, while sub-equal length of the elements as in A. bon-nali would represent the secondary returnto a primitive state.
In the other European Archaeolacer-tae, there are two exceptions from the ruleof subequal length: A. horvathi, in whichthe postorbital is notably longer than thepostfrontal, and A. oxycephala, in whichthe reverse situation is found.
In the species of the 'Z..' saxicolagroup, however, both bones can be sube-qual, but there is a marked tendency to-wards a postorbital which is shorter thanthe postfrontal ('Z,.1 saxicola ssp., 'Z,.1 lind-holmi, 'L' alpina, 'Z,.' daghestanica, 'L.'raddei, 'Z,.'parvula, 'L' mixta).
Overlap of postorbitaland squamosal
Among the species studied, the lengthof the contact suture between squamosaland postorbital varies from something lessthan a third to near half of the length of thepostorbital. In general, variation is fairlyrestricted within a species. It therefore
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Osteology of Archaeolacerta s. 1. 53
makes sense to indicate whether the lengthof the contact zone is closer to one half orone third of the length of the postorbitalbone. The most frequently found conditionis one third of overlap (as is present in theoutgroup and is the most common situationin the ingroup), which is therefore consid-ered the primitive character state.
In A. bonnali the length of overlap isabout 1/3 of the postorbital length (20 % ofthe specimens approach to 1/2). In A. ara-nica, the overlap is close to 1/2 (100% ofthe specimens) and in A. aurelioi also 1/2(30% of the specimens approach to 1/3).
Fusion or separationof postorbital and postfrontal
Depending on the species, postfrontaland postorbital are either fused or sepa-rated from birth. In species in which thesebones are united from birth, they form anentire piece, possible constituents of whichremain completely undifferentiable (seee.g. the development of Z. vivipara inRIEPPEL 1992). In all outgroup taxa, post-orbital and postfrontal appear as a singleelement, which consequently is consideredthe primitive character state and the pres-ence of separated bones as the derived one.Nevertheless, ARNOLD (1989) observed thatthese bones were separated in juveniles andfused in the oldest individuals of some lac-ertid species. By finding this, he concluded- based on an ontogenetic criterion - thatthe presence of two individualized boneswould represent the primitive state. I think,however, that the secondary fusion men-tioned is something different from having asingular bone from birth. In the first case,the suture of the bones is markedly imbri-cated and interdigitated. This joining proc-ess is part of the development of a robustskull (including proliferation of cranial osteo-derms - crusta calcarea) which leaves bothbones still distinguisliable (coalescence of thetwo bones during ontogeny). This processis to be distinguished from the primordialexistence of a singular bone.
All three Pyrenean species have sep-arated bones, hatchlings as well as old in-dividuals. A specimen of A. aranica pres-ents an aberrant fusion of the bones in oneside of the head, while the other side showsthem perfectly separated.
All other species of Archaeolacerta s.1. display both bones separated, as most ofthe species of the Eurasian radiation do.Within the Lacertidae, the possession ofbones fused from birth is found in the out-group genera Gallotia and Psammodromus,but is also present in Zootoca and someLacerta s. str.(e.g., L. schreiben).
POSTCRANIAL CHARACTERS
Bony ribs on third presacrai vertebra
According to the osteological litera-ture on Lacertidae, the fourth presacraivertebra in the cervical region is the first tobear ribs implying that the third vertebralacks ribs (e.g., BARBADILLO & SANZ 1983).However, I found that in those species(outgroup species included) qualified as themost primitive lacertids, small bony ribswere associated with the third presacraivertebra. In the more advanced lacertids,these ribs frequently do also exist but arenot that apparent because of being reducedto their proximal part and/or not ossified.
Bony ribs associated with the thirdvertebra were found to be present in Gal-lotia, Psammodromus, Z. vivipara and L.agilis, and in the three Pyrenean Archaeo-lacerta and were found to lack in mostgenera of the the Eurasian radiation (JPod-arcis, Te ira, Algyroides, Archaeolacerta s.1. - except the Pyrenean ones).
Stability of this character state is vari-able. In A. aranica, bony ribs associatedwith the third vertebra were expressed in90 % of the specimens examined; in A. au-relioi in 70 %, and in A. bonnali in some-what more than 50 %. Hence, their pres-ence is the standard in the Pyrenean Ar-chaeolacertae suggesting their close inter-relationship as against the other EuropeanArchaeolacertae s. 1.
All other Iberian and European Ar-chaeolacerta s. 1. as well as the species ofthe 'Z,.' saxicola group lack bony ribs asso-ciated with the third presacrai vertebra.Very occasionally I found specimens withinthese taxa that presented these ribs ossified(one specimen of A. horvathi and one of X.1
unisexualis). Both cases should be classi-fied as atavism which was also observed ina few exceptional specimens among other
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

54 OSCAR J. ARRIBAS
genera studied (Podarcis, Teira). In the Py-renean species, the presence of these bonyribs could be interpreted as a secondary re-turn as well.
Ribs of sixth presacrai vertebra
The sixth presacrai vertebra is thelast short-ribbed vertebra previous to thosethat bear long ribs the first of which are notlinked to the sternal apparatus. In somespecies (P. algirus, L. agilis, Z. vivipara),the ribs of the sixth vertebra are elongateand relatively undifferentiated (primitivestate), while in most Lacertidae (e. g., Ar-chaeolacerta s. 1., Podarcis, Teira, 'L.' sa-xicola group, Algyroides Gallotia) theseribs are short and spoon-shaped with twosmall processes extending towards the su-prascapula (derived state). Exceptionally,individuals can be found among them (e.g., a specimen of 'L.' daghestanicd) whichapproach the primitive state.
Sternal-xiphisternal costal formula
Sternal-xiphisternal costal formula isrepresented by the term (x+y), where x andy are the numbers of ribs with sternal andxiphisternal contact, respectively. In Lacerti-dae, three sternal formulae are found: (3 +1),(3+2) and (3+3), the second being the mostcommon type. Two formulae can co-occurin one species: e.g., (3+2) [frequently] and(3+1) [more rarely] in Gallotia and Zoo-toca (primitive condition) or (3+2) [fre-quently] and (3+3) [more rarely] in allother taxa studied (derived character state).
In the three Pyrenean species, theformula is (3+2) [(3+3) in one A. aurelioispecimen from Montroig Massif]. The otherspecies of Archaeolacerta s. 1. includingthe Caucasian species, also present (3+2),rarely (3+3).
Number of presacrai vertebrae
The majority of Lacertidae displayssexual dimorphism in the number of pre-sacral vertebrae (primitive condition) whileGallotia and some Acanthodactylus speciesdo not (derived condition). The modalnumber of vertebrae is superior in femalesin that they have one to three vertebraemore than males. According to ARNOLD
(1983), there is a clear although not rigidcorrelation between number of vertebraeand habitat: high vertebral counts are pref-erentially found in species colonizing me-sic habitats, while low counts are typical tospecies from more arid, open and lessvegetated places. The above author sug-gests that mesic habitat could requiregreater inflection of the body in order tomove among vegetation and that a greaternumber of vertebrae facilitates this move-ment. Therefore, sexual dimorphism in thenumber of vertebrae would reflect differ-ences between sexes in microhabitat selec-tion, in that the males would spend moretime in open areas due to territorial activi-ties. An alternative theory by the sameauthor which is more in accordance withmy observations says that extra presacraivertebrae would allow expansion of thebody volume in gravid females and, thus,increased clutch volume. However, I thinkthere is an important phylogenetic back-ground reflected by the state of this charac-ter in that the number of presacrai verte-brae is similar in phylogenetically relatedspecies.
In A. bonnali, males always present26, and females 27 presacrai vertebrae. Thesame numbers apply to A. aranica and A.aurelioi, but in each of the latter two spe-cies one female was found to have 28 ver-tebrae. Males of some European Archaeo-lacerta s. 1. (A. monticola, A. cyreni, A.bedriagae, A. horvathi) have modal countsof 26 vertebrae, while others frequentlyshow 25 only (A. mosorensis, A. oxyceph-ala; versus 26 found by ARNOLD 1973 fromspecimens of other localities). The femalesare more variable in this respect: 28 verte-brae are frequently encountered in A mon-ticola and A. cyreni, while the numbers of26, 27 and 28 appear in A. mosorensis.Female A horvathi, A. oxycephala and Abedriagae usually present 27 vertebrae.
In Podarcis the number of presacraivertebrae is 26 or 27 in males; in Algyroi-des and Apathya, reduction to 25 is notrare in males, whereas all males of the 'L'saxicola group present 27 (except 'Z-.' rad-dei which has 28). Females usually haveone more vertebra than males (very rarelytwo or even three).
In the males of the outgroup genera{Gallotia, Psammodromus), 26 vertebrae
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Osteology of Archaeolacerta s. 1. 55
were observed. This number is consideredto be the plesiomorphic character state,while reduction to 25 as well as increase to27 or 28 (as found in male 'Z,.1 parva and'L' fraasi) are apomorphic character stateswhich evolved independently.
Number of short-ribbed posteriorpresacrai vertebrae
ARNOLD (1989) compares the numberof these posterior presacrai vertebrae ("num-ber of short free dorsal ribs") with thenumber of anterior dorsal ribs, and con-cludes that low numbers indicate the primi-tive state and high numbers the derivedone. (i) both conditions have been found inthe outgroup, and (ii) elevated numbers ofshort vertebrae are widespread in the mostadvanced species of the African radiation.To my opinion, these facts make it difficultto speculate on the direction of the evolu-tionary development of this character.
The Pyrenean species usually showsix short-ribbed posterior presacrai verte-brae in both sexes.
Archaeolacerta bonnali of Bigorre isoutstanding in that it presents a constantnumber of five short-ribbed posterior pre-sacral vertebrae, while the only clearedspecimen of Aigiiestortes has seven. Bothcases refer to relatively isolated populationsin which the establishment of accidentalvariations may be more likely. Most impor-tant, the reduction to five short-ribbed ver-tebrae in the posterior presacrai region isnot paralleled by the total number of pre-sacral vertebrae, implying that this popula-tion is distinguished by one more long dor-sal rib. Given that the long ribs are impor-tant structures in flattening the body forbasking, the genetic fixation of this charac-teristic in the Bigorre population couldconstitute an adaptive advantage in ther-morégulation for these individuals whichlive on the northern slope of the Pyreneanmountains. A detailed study of a greatersample including neighboring populationsmight throw more light on this topic.
All A. aranica and A. aurelioi speci-mens studied present 6 short-ribbed poste-rior presacrai vertebrae, with the exceptionof a young male aranica of Port d'Orlawhich has 5.
In the other Archaeolacerta s. 1., sixis also the common number, e. g. in Amonticola and A. cyreni (7 in a female ofGuadarrama), however, in some individu-als of A horvathi, A. mosorensis and A.oxycephala the number of short-ribbedposterior presacrai vertebrae is 5.
Six is the usual count for the Cauca-sian species of the 'L' saxicola group;numbers of 5 or 7 occur less frequently. 'Z.'portschinskii which is distinguished by 7short-ribbed posterior presacrai vertebrae,is the parental species (with still somedoubts) of the parthenogenetic '£.' dahliand '£.' rostombekovi, which frequentlyalso present 7.
Expression and shapeof sternal fontanelle
The sternal fontanelle is a foramen inthe sternal plate, present from birth andfound in most lacertids. Its shape can berounded, more or less oval, slightly orclearly heart-shaped (cordiform, as in Pod-arcis, see ARNOLD, 1973, 1989). Obviouslythe rounded and oval, ill differentiated andvery widespread types, represent the primi-tive character state, while the cordiform fon-tanelles are derived (ARNOLD, 1973, 1989).As it is not clear whether the slightly cor-diform type represents an intermediate typeor a secondary regression to the primitivestate, this type is independently treatedhere, as was done by ARNOLD (1989).
In A bonnali the sternal fontanelle isoval or rounded (cordiform in one speci-men from Bigorre) and rounded in all Aaranica studied.
Almost 50 % of the A aurelioispecimens (from three different massifs)lack the fontanelle, and 20 % present it re-duced to a pair of small foramina, whileabout 30 % show the typical oval form.This absence or reduction of the fontanelleis supposed to be a secondary loss, as sug-gested by the fact that there is a continuousseries of character states ranging from ovalfontanelles through small holes to the ab-sence of perforation. It may be allowed tospeculate here, that the cordiform fontan-elle could be a transitional stage to partialclosing of the fontanelle.
The other European Archaeolacertas. 1. studied present oval fontanelles. Spo-
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

56 OSCAR I. ARRIBAS
radically, you find individuals of A. cyreniand A. horvathi with reduced fontanelles.The fontanelle is oval in the 'Z.1 saxicolagroup, although reduced but still visible insome individuals (as in a specimen of 'Z.'chlorogaster) or clearly cordiform (as in anindividual of 'L' derjugini, '£.' armeniacaand 'Z.' mixta, and several specimens of'Z.1
alpina). In general, the other species in-cluding the outgroup (but with the excep-tion of Podarcis, 'Z.' laevis, 'Z.1 danfordi,some A moreoticus, A. marchi and 'Z.' an-dreanskyi) exhibit clearly oval or roundedfontanelles, wherefore this state is consid-ered the most primitive.
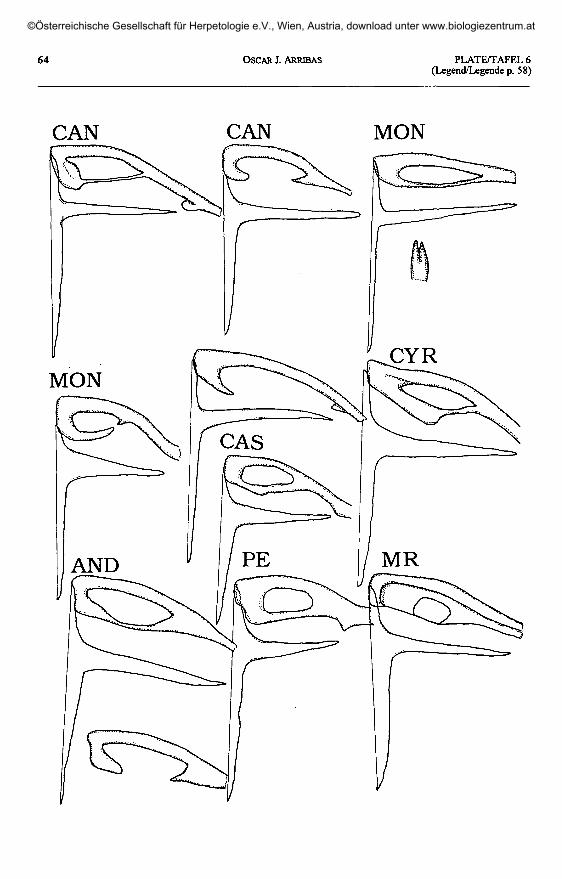
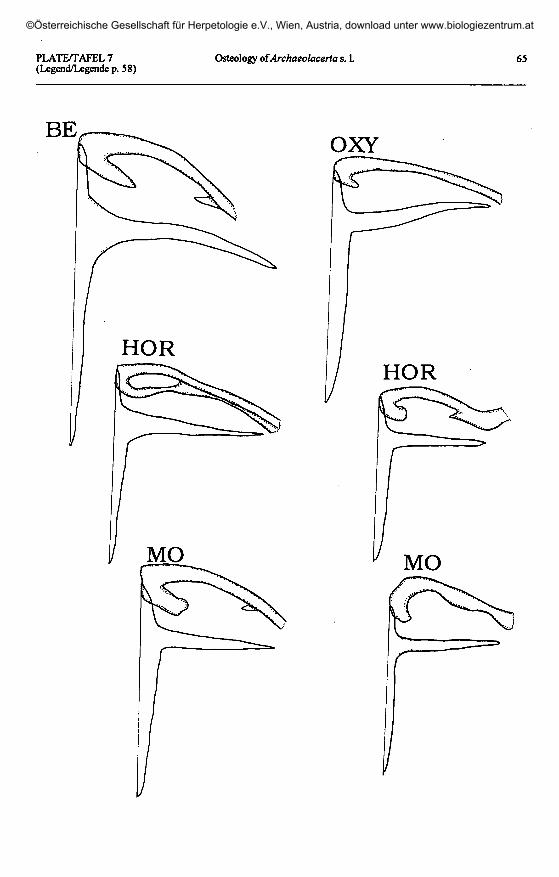
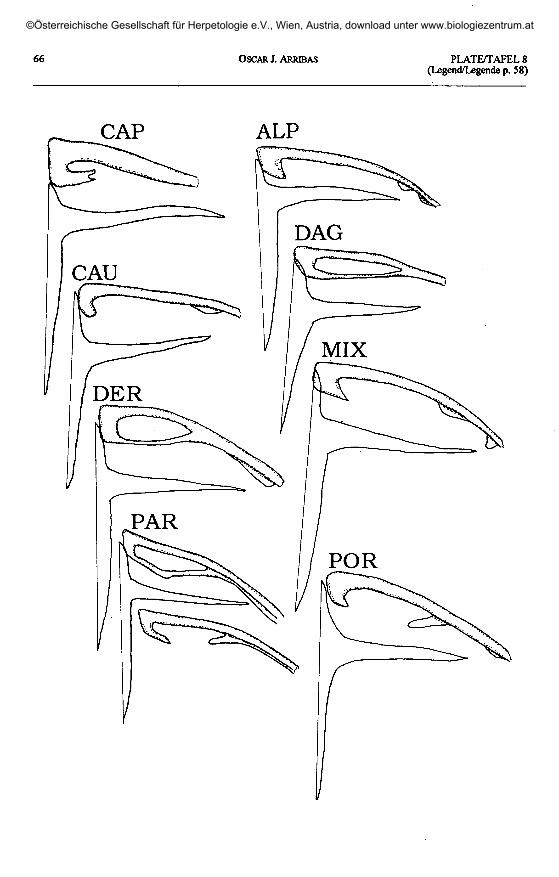
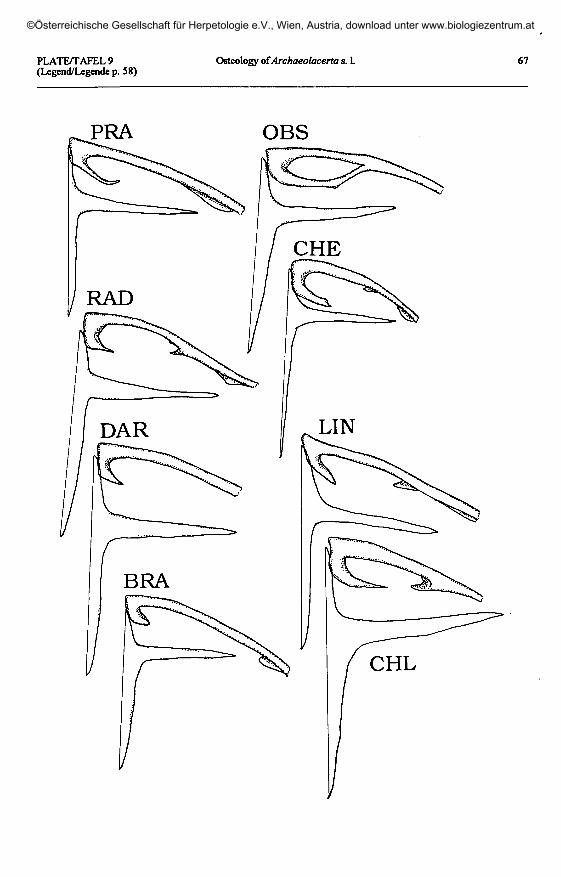
Shape of clavicle(plates 5 -10)
In general, the clavicle of Lacertidaeis characterized by the presence of a greatforamen in its proximal expanded part.This aperture is bordered by an anteriorand a posterior clavicular branch (un-emarginated or closed type). In many cases,the posterior branch is incompletely formed(emarginated or open type). Obviously, theclosed type seems to be the primitive but inmany species one can encounter both types.Several genera and species mainly presentthe closed condition (Psammodromus, Gal-lotia) while others the open type (e. g. A.bedriagae, A. oxycephala). Some authors(e. g. ARNOLD 1989) consider the presenceof both types as the primitive situation andthe exclusive presence of one or the otheras the derived condition.
All A. aranica and A. bonnali speci-mens, except a bonnali hatchling fromAigiiestortes (the one that has seven small-ribbed posterior presacrai vertebrae), showopen clavicles, while all A. aurelioi but onepresent closed clavicles. In this discordantspecimen (which also has an aberrantpostfrontal, plate 2) from Port de Rat (An-dorra), the clavicle is open. Both peculiari-ties can be interpreted as atavisms in thisindividual.
In the other European species of Ar-chaeolacerta s. 1., the situation is variable.Archaeolacerta montico!a montico la pres-ents closed clavicles, while in A. monticolacantabrica there are almost twice as muchopen clavicles than closed (this observationis contrary to the findings of PÉREZ-MEL-
LADO & al. 1993). Clavicles are mainlyclosed in A. cyreni, open in A. bedriagae,A. oxycephala and A. mosorensis, andvariable in A. horvathi. In the 'Z.1 saxicolagroup and in Apathya cappadocica, bothtypes of clavicles appear blended in almostall species.
Shape of interclavicle(plates 5 -10)
In all lacertids, the interclavicle is across-shaped (cruciform) element. The an-terior process is very short in relation to theposterior one, and the lateral extensions areintermediate in length.
In general, the lateral extensions aremore or less perpendicular to the antero-posterior axis; occasionally they are some-what or clearly inclined in anterior or pos-terior direction. Lateral extensions whichare more or less perpendicular representthe primitive character state, as present inthe outgroup species and in the majority ofthe lacertids of the Eurasian radiation.Deviations from this most common stateshall be considered as derived and arefound in 'Z,.' chlorogaster (clearly directedanteriorly), A. oxycephala (moderately di-rected anteriorly), T. dugesii, [and T. per-spicillata - not studied here] (clearly direc-ted posteriorly), and 'Z.1 andreanskyi (lessclearly directed posteriorly - but comp. AR-NOLD 1973, 1989).
The three Pyrenean species displaythe primitive type, without sensible devia-tions. The same situation is found in allother European Archaeolacerta s. 1., andthe Caucasus and Asia Minor species, withthe exception of 'Z.1 chlorogaster. I couldnot find large lateral processes in the in-terclavicle of A. bonnali (as described byPÉREZ-MELLADO & al. 1993) which is notdifferent from those of the other taxa stud-ied here (comp. plate 5).
Length of anterior and posteriorinterclavicular process
Proportion of the length of the ante-rior and posterior interclavicular processvaries within and between species. In Psam-modromus algirus for instance, the anteriorprocess is quite long, attaining almost 50%of the length of the posterior.
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Osteology of Archaeolacerta s. 1. 57
Within the Pyrenean species, ratios(anterior/posterior) are 0.21-0.38 (x= 0.29,StD = 0.54, n = 6) in A. bonnali, 0.20 -0.35 (x = 0.26, StD = 0.53, n = 6) in A.aranica, and 0.22 - 0.35 (x = 0.29, StD =0.39, n = 9) in A aurelioi.
In the other Archaeolacerta s. 1. spe-cies and P. algirus, ratios range from 0.25to 0.46 and display great individual vari-ability within populations. The index seemsnot to be related to the habitat type, sinceground dwellers and saxicolous species canhave similar values. I therefore hypothe-size, that (except in very peculiar cases) thelength ratio of the anterior and posteriorinterclavicular processes is subject to con-siderable variation and of little use in thestudy of lacertid phylogeny.
Preautotomic caudal vertebrae
ARNOLD (1973, 1989) distinguishedthree types of vertebrae in the immediatepreautotomic caudal region: Type A showsone pair of perpendicular processes in theanterior part of the vertebra; type B withtwo pairs of processes, a bigger anteriorand a smaller posterior one; type C displaystwo pairs of processes, a smaller anteriorand a bigger posterior one.
Types A and B frequently co-occurand are very widespread among the Lacer-tidae of the Eurasian radiation, all speciesof Archaeolacerta s. 1., Apathy a included.Type C appears in Podarcis, some speciesfrom Asia Minor ('Z,.1 danfordi, 'L' ana-tolicd) and in the outgroup species Gallotiaand Psammodromus. According to ARNOLD(1989), 'L' parva, 'L' fraasi, and 'L'brandtii are intermediate between types Band C. Although this author considers
types A and B to represent primitive char-acter states, I think that type C as found inthe outgroup taxa and the species of theAfrican radiation (Acanthodactylus) shouldbe regarded the primitive state. Never-theless, homology and direction of evol-ution of the different types is not clearlyunderstood.
The Pyrenean species A. bonnali, A.aranica and A. aurelioi present type A with-out exception.
In the other Archaeolacertae of theIberian Peninsula the situation is verysimilar. Archaeolacerta montico la monti-cola^, m. cantabrica, and A cyreni showtype A vertebrae. However, in 30 % of thelatter two taxa I encountered additionalprocesses reminiscent of type B (whichmeans that there are vertebrae of interme-diate morphology between A and B, withan anterior process in one side and twoprocesses in the other).
In the other European Archaeola-certae the situation is very similar. In A.bedriagae, and A. horvathi I only foundtype A (but this could be due to the smallsample size), while A. mosorensis, and A.oxycephala present type A vertebrae with afew processes of type B in some individu-als.
Apathya cappadocica shows verte-brae of type A, while in the species of theof 'L' saxicola group, the situation is vari-able within the above spectrum. Vertebraeof type A (frequently associated with someof type B) are the most common combina-tion. The reverse situation (type B verte-brae with some of type A) is found in 'L. 'mixta, 'L.1 parvula, L.x praticola, and '£.'raddei.
CONCLUSIONS
(i) The Pyrenean species of Ar-chaeolacerta are unequivocally distin-guished from all other species of the Eura-sian radiation (sensu MAYER & BENYR1994) by the possession of bony ribs asso-ciated with the third presacrai vertebra.This is a plesiomorphic character statewithin the lacertids, but can be interpretedas a secondary return to the primitive statein this group.
(ii) Within the Pyrenean group, A.aranica is characterized by the presence ofa well developed antero-distal process ofthe postfrontal as well as antero-medialprocess of the postorbital, and by havingopen (emarginated) clavicles, and oval orrounded sternal fontanelles.
(iii) Within the Pyrenean group, A.bonnali is characterized by the possessionof the antero-distal process in the postfron-
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

58 OSCAR J. ARRIBAS
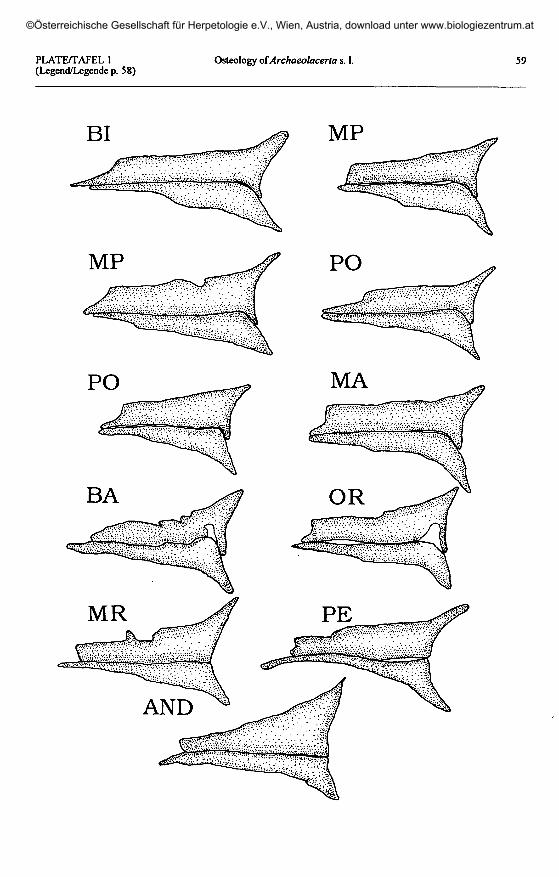
Legends to plates 1 to 10Legenden zu den Tafeln 1 bis 10
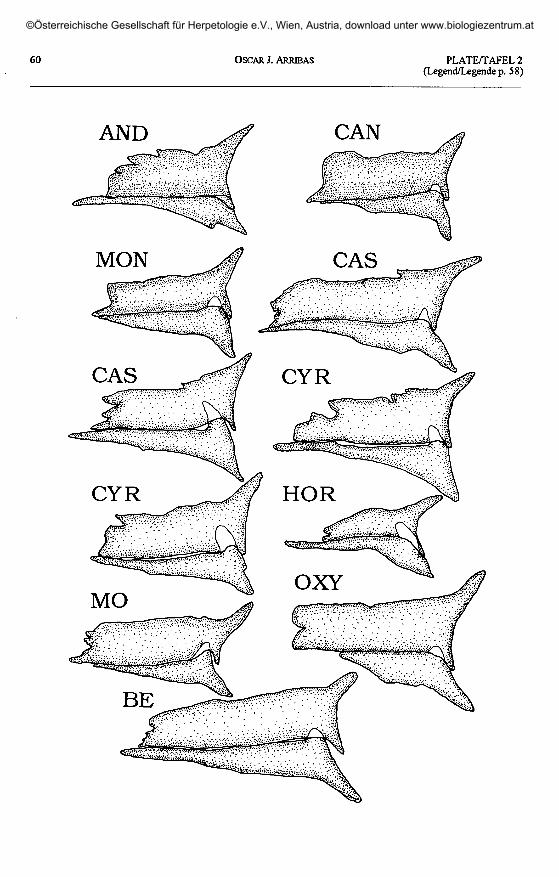
Plate I (opposite page): Dextral postfrontal and postorbital bones in Archaeolacerta s. 1.The part of the anteromedial postorbital process covered by the postfrontal is shown in white.
Taf. 1 (gegenüberliegende Seite): Postfrontale und Postorbitale der rechten Seite bei Archaeolacerta s. 1.Der vom Postfrontale verdeckte Teil des antero-medialen Postorbitalfortsatzes ist weiß dargestellt
A. bonnali: BI - Lac Bleu de Bigorre; MP - Monte Perdido; PO - Vail del Clot, Posets; MA - Estany de Llauset,Maladeta. A. aranica: BA - Coli de Barradós; OR - Port d'Orla. A. aurelioi: MR - Estany de Calberante, Montroig;
PE - Estany de Sotllo, Pica d'Estats; AND - Port de Rat, Andorra-Ariège.
Plate 2 (plate I continued, page 60) / Taf. 2 (Taf. 1 fortgesetzt, Seite 60).
A. aurelioi: AND - Port de Rat, Andorra (aberrant morphology, atavism).A. monticola cantabrica: CAN - Somiedo. A. monticola monticola: MON - A Torre, Estrela, Portugal.
A. cyreni castiliana: CAS - Laguna Grande, Gredos. A. cyreni cyreni: CYR - Pto. de Navacerrada.A. horvathi: HOR - Val Dogna, Italy. A. mosorensis: MO - Zmisko Jezero, Montenegro.A. oxycephala: OXY - Nicsic, Croatia. A. bedriagae bedriagae: BE - Corsica, France.
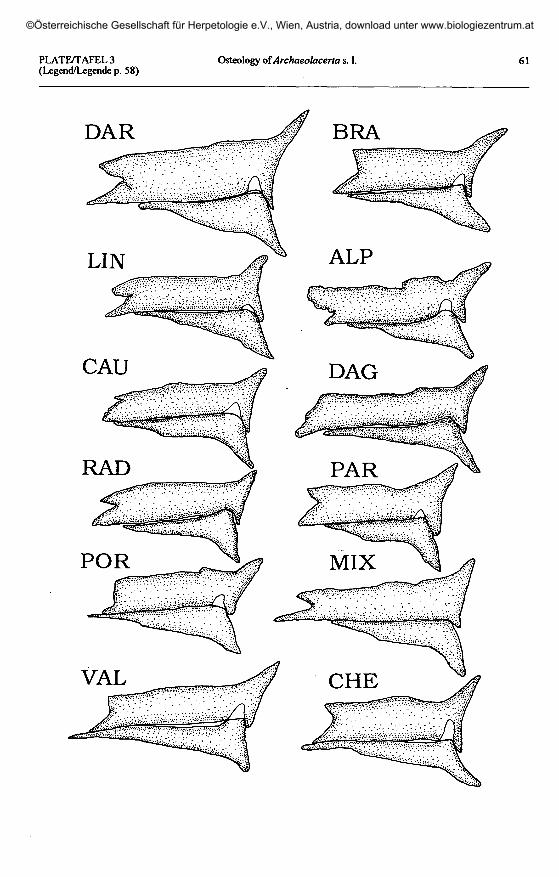
Plate 3 (plates 1 & 2 continued, page 61) / Taf. 3 (Taf. 1 & 2 fortgesetzt, Seite 61).'L.' saxicola darevskii: DAR - Dagomys, Sochi, Russia. 'L.' saxicola brauneri: BRA - Inguri River, Georgia.
'L.' lindholmi: LIN - Bakhchisaray, Georgia. 'L.' alpina: ALP - Fisht Mount, Krasnodar, Russia.'L.' caucasico: CAU - Pereval, Southern Ossetia. 'L.' daghestanica: DAG - Khvarshi, Daghestan, Russia.
'L.' raddei raddei: RAD - Lichk, Armenia. 'L.'pannila: PAR - Adskuri, Georgia.'L.' portschinskii: POR - Rbemoshuri, Georgia. 'L.' mixta: MIX - Baniskevi, Georgia.
'L.' valentini: VAL - Legli Mt., Armenia. 'L.' rudis chechenica: CHE - Khvarshi, Daghestan, Russia.
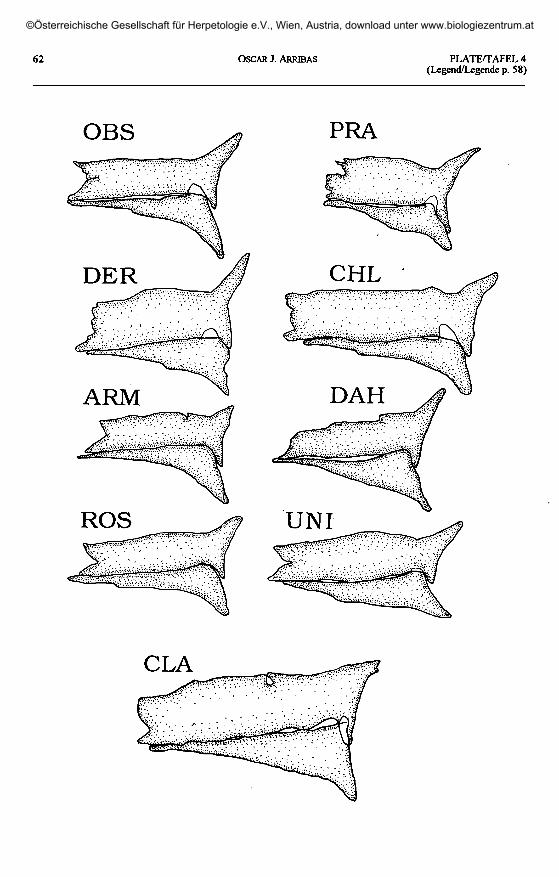
Plate 4 (plates 1 - 3 continued, page 62) / Taf. 4 (Taf. 1 - 3 fortgesetzt, Seite 62).'L' rudis obscura: OBS - Achaldaba, Georgia. 'L.'praticola: PRA - Kutorskaya,
Krasnodar, Russia. 'L.' derjugini: DER - Mlashe, Georgia. 'L.' chlorogaster: CHL - Alexiefska, Azerbaijan.'L.' armeniaca: ARM - Arpalich, Armenia. 'L.' dahli: DAH - Jodieri, Georgia. 'L.' rostombekovi: ROS - Zumabad,
Azerbaijan. 'L.' unisexualis: UNI - Tsomakaberd, Armenia. 'L.' clarkorum: CLA - Cankurtaran Gee, Turkey.
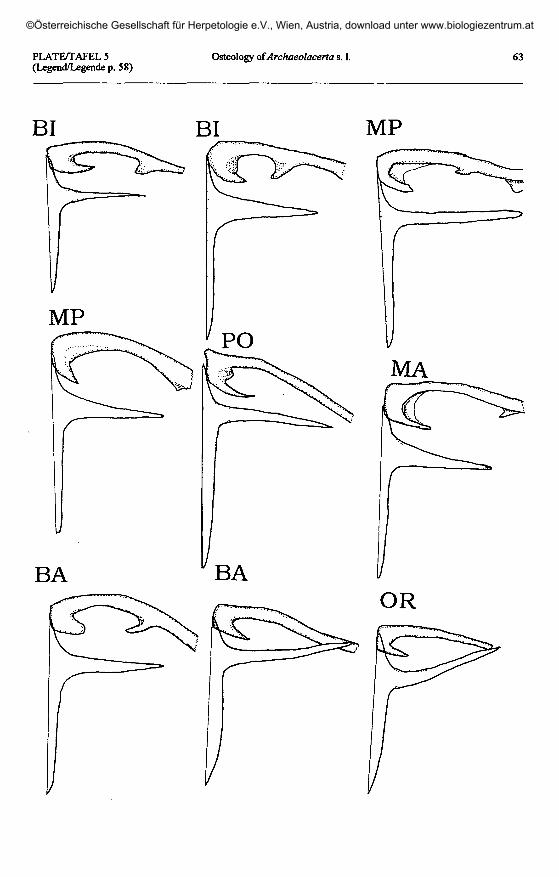
Plate 5 (page 63): Clavicle and interclavicle in Archaeolacerta s.l. The left ventral side of the apparatus is shown.Taf. 5 (Seite 63): Klavikel und Interklavikel bei Archaeolacerta s. 1. Linke Seite des Apparates von ventral.
A. bonnali: BI - Lac Bleu de Bigorre; MP - Monte Perdido; PO - Vail del Clot, Posets; MA - Estany de Llauset,Maladeta. A. aranica: BA - Coli de Barradós; OR - Port d'Orla.
Plate 6 (plate 5 continued, page 64) / Taf. 6 (Abb 5 fortgesetzt, Seitee 64).
A. monticola cantabrica: CAN - Somiedo. A. monticola monticola: MON - A Torre, Estrela, Portugal.A. cyreni castiliana: CAS - Laguna Grande, Gredos. A. cyreni cyreni: CYR - Pto. de Navacerrada.
A. aurelioi: MR - Estany de Calberante, Montroig; PE - Pica d'Estats; AND - Port de Rat, Andorra-Ariège.
Plate 7 (plates 5 & 6 continued, page 65) / Taf. 7 (Taf. 5 & 6 fortgesetzt, Seite 65).A. bedriagae bedriagae: BE - Corsica, France. A. oxycephala: OXY - Nicsic, Croatia.
A. horvathi: HOR - Val Dogna, Italy. A. mosorensis: MO - Zmisko Jezero, Montenegro, Yugoslavia.
Plate 8 (plates 5 - 7 continued, page 66) / Taf. 8 (Taf. 5 - 7 fortgesetzt, Seite 66).
Apathya cappadocica wolteri: CAP - Kirikan, Amanus Mts, Turkey. 'L.' alpina: ALP - Fisht Mt., Krasnodar, Russia.'L.' caucasico: CAU - Pereval, Southern Ossetia, Russia. 'L.' daghestanica: DAG - Khvarshi, Daghestan, Russia.
'L.' derjugini: DER - Mlashe, Georgia. 'L.' mixta: MIX - Baniskevi, Georgia. 'L.'pannila: PAR - Adskuri, Georgia.'L.' portschinskii: POR - Rbemoshuri, Georgia.
Plate 9 (plates 5 - 8 continued, page 67) / Taf. 9 (Taf. 5 - 8 fortgesetzt, Seite 67).'L.'praticola: PRA - Kutorskaya, Krasnodar, Russia. 'L.' rudis obscura: OBS - Achaldaba, Georgia.
'L.' rudis chechenica: CHE - Khvarshi, Daghestan, Russia. 'L.' raddei raddei: RAD - Lichk, Armenia.'L.' saxicola darevskii: DAR - Dagomys, Sochi, Russia. 'L.' saxicola brauneri: BRA - Inguri River, Georgia.
'L.' lindholmi: LIN - Bakhchisaray, Georgia. 'L.' chlorogaster: CHL - Alexiefska, Azerbaijan.
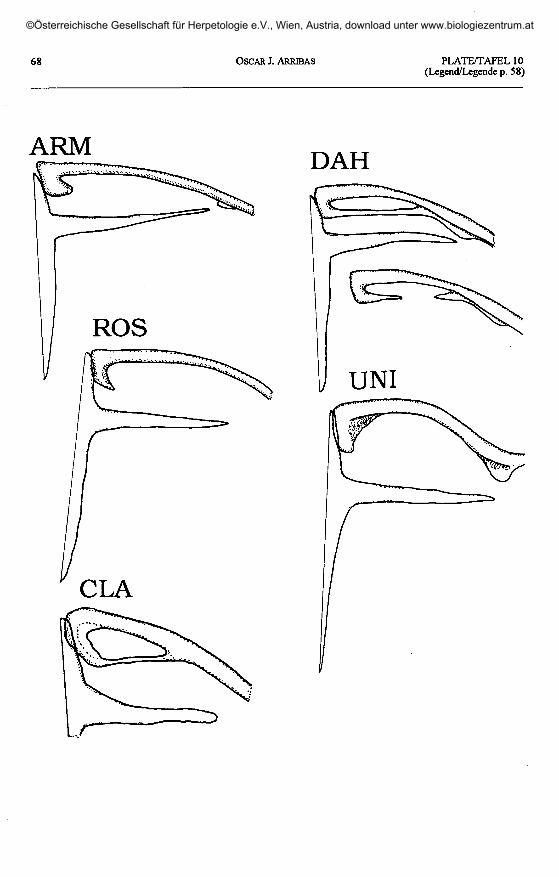
Plate 10 (plates 5 - 9 continued, page 68) / Taf. 10 (Taf. 5 - 9 fortgesetzt, Seite 68).
'L' armeniaca: ARM - Arpalich, Armenia. 'L.' dahli: DAH - Jodieri, Georgia. 'L.' rostombekovi: ROS - Zumabad,Azerbaijan. 'L.' unisexualis: UNI - Tsomakaberd, Armenia. 'L.' clarkorum: CLA - Cankurtaran Gee, Turkey.
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
PLATE/TAFEL 1 Osteology of Archaeolacerta s. 1.(Legend/Legende p. 58)
59
MP
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
60 OSCAR J. ARRIBAS PLATE/TAFEL2(Legend/Legende p. 58)
AND
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
PLATE/TAFEL3(Legend/Legende p. 58)
Osteology of Archaeolacerta s. I. 61
DAR
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
62 OSCAR J. ARRIBAS PLAT&TAFEL4(Legend/Legende p. 58)
OBS PRA
CLA
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
PLATE/TAFEL 5 Osteology oîArchaeolacerta s. 1.(Legend/Legende p. 58)
63
MP
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
64 OSCAR J.ARRiBAS PLATE/TAFEL6(Legend/Legende p. 58)
CAN CAN MON
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
PLATE/TAFEL7(Legend/Legende p. 38)
Osteology of Archaeolacerta s. 1. 63
OXY
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
66 OSCAR J. ARMBAS PLATEvTAFEL 8(Legend/Legende p. 58)
CAP ALP
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
PLATE/TAFEL 9(Legend/Legende p. 58)
Osteology of Archaeolacerta s. 1. 67
PRA OBS
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at
68 OSCAR J. ARRIBAS PLATE/TAFEL 10(Legend/Legende p. 58)
ARM DAH
r
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

Osteology of Archaeolacerta s. 1. 69
tal, and by the lack of the antero-medialprocess of the postorbital. The lizard hasopen (emarginated) clavicles and oval orrounded sternal fontanelles.
(iv) Within the Pyrenean group, A.aurelioi is characterized by the lack of bothpostfrontal and postorbital processes, by thepossession of closed clavicles (unemargin-ated) and the frequent lack or reduction ofthe sternal fontanelle.
(v) All above character states areconstantly expressed throughout the spe-cies' range. But we can also find charactersthe expression of which varies greatly, es-pecially when peripheral or isolated popu-lations are included, e. g., acquisition of anadditional long rib in disfavour of thenumber of short ribs, as occurs in theBigorre population of A. bonnali (but alsoin other species).
(vi) In view of the osteological re-sults, it becomes clear that the Europeanspecies so far assigned to Archaeolacerta s.1. do not constitute a natural cluster. Onone hand, the Pyrenean species appear dif-ferentiated on the basis of the charactersstudied, while the Iberian, Balcanic andTyrrhenic taxa share symplesiomorphiccharacter states only which do not permitconclusions on their relationship.
(vii) The Caucasian species of the'Z,.' saxicola group, currently assigned toArchaeolacerta s. 1., are different from theEuropean Archaeolacertae by the apomor-
phic character of an extra presacrai verte-bra and from Apathy'a by both an extra pre-sacral vertebra and a minor number ofteeth in the premaxila.
(viii) Within the Caucasian group, '£.'chlorogaster appears clearly differentiatedfrom the other taxa by the number of pre-maxillary teeth and the shape of the inter-clavicle, which coincides with the lizard'suncertain systematic position (see BISCHOFF1991).
(ix) The parthenogenetic Caucasianspecies present characteristic features oftheir parental species, the female parentalspecies of the original crossing in particu-lar. Lack or extreme reduction of the an-tero-medial process of the postorbital in theoriginal maternal species 'L.' raddei and'Z,.' mixta is also expressed in the partheno-genetic descendant species studied here.Likewise, the modal number of vertebrae ofthe parthenogenetic females of the 'L' rad-dei cluster is somewhat increased as com-pared to the other species of the '£.' saxi-cola group, with the exception of the ma-ternal species, which also show thesehigher vertebral counts.
(x) '£.' portschinskii frequently pre-sents the exceptional number of 7 short-ribbed posterior presacrai vertebrae, whichspeaks in favour of this taxon to be anoriginal parental species of the partheno-genetic *L' dahli and 'L' rostombekovi,which also present this elevated number.
AKNOWLEDGMENTS
I want to express my sincere thanks to the follow-ing colleagues (in alphabetical order): To W. BÖHME(Bonn, D), I. S. DAREVSKY (St. Petersburg, RU), J. El-SELT (Wien, A), H. GRILLITSCH (Wien, A), C. P. GUIL-LAUME (Montpellier, F), B. LANZA (Firenze, I), E. S.ROYTBERG (Halle, D), N. SZCZERBAK (Kiev, Ukr), J. F.SCHMIDTLER (München, D), F. TŒDEMANN (Wien, A),
A. TOKAR (Kiev, Ukr) and B. TUNIYEV (Sochi, Ru), forproviding literature and valuable specimens; to C. ARRI-BAS for facilities in radiographizing specimens. Allspecimens from the very localized Pyrenean Mountainlizards have been collected with permissions of the Ara-gonian and Catalonian Autonomie governments (wildlifeprotection sections).
REFERENCES
ARNOLD, E. N. (1973): Relationships of the pa-laearctic lizards assigned to the genera Lacerta, Algyro-ides and Psammodromus (Reptilia: Lacertidae).- Bull.British Mus. nat. Hist. (Zool.), London; 25: 289-366.
ARNOLD, E. N. (1983): Osteology, genitalia andthe relationships of Acanthodactylus (Reptilia: Lacerti-dae).- Bull. British Mus. nat. Hist. (Zool.), London; 44(5): 291-339.
ARNOLD, E. N. (1989): Towards a phylogenyand biogeography of the Lacertidae: relationships withinan Old-World family of lizards derived from morphol-ogy.- Bull. British Mus. nat. Hist. (Zool.), London; 55(2): 209-257.
ARRIBAS, O. J. (1993): Intraspecific variability ofLacerta {Archaeolacerta) bonnali LANTZ, 1927 (Squama-ta: Sauria: Lacertidae).- Herpetozoa, Wien; 6 (3-4): 129-140.
ARRIBAS, O. (1994): Una nueva especie de la-gartija de los Pirineos Orientales: Lacerta {Archaeo-lacerta) aurelioi sp. nov. (Reptilia: Lacertidae).- Boll.Mus. reg. Sci. nat., Torino; 12 (1): 327-351.
ARRIBAS, O. (1996): Morfologia, filogenia y bio-geografìa de las lagartijas de alta montana de losPirineos. PhD Thesis. Pub. Universität Autònoma deBarcelona (1977). Bellaterra. Vili+ 353 pp.
BARBADILLO, L. J. & SANZ, J. L. (1983): Analisis osteométrico de las regiones sacra y presacra de la
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at

70 OSCAR J. ARRIBAS
columna vertebral en los lagartos Ibéricos Lacerta viri-dis LAURENT!, Lacerta lepida DAUDIN y Lacerta schreibenBEDRIAGA.- Amphibia-Reptilia, Bonn; 4: 215-239.
BISCHOFF, W. (1991): Übersicht der Arten undUnterarten der Familie Lacertidae 3. Die Gattung Lacer-ta.- Die Eidechse; 3: 5-16.
CAMP, C. L (1923): Clasification of the lizards.-Bull. American Mus. Nat Hist; 48: 289-481.
DAREVSKY, I. S. (1967): Rock lizards of theCaucasus. Systematics, ecology and phylogenesis of thepolymorphic groups of Rock lizards of the subgenus Ar-chaeolacerta. Leningrad (Nauka press).
DAREVSKY, I. S. (1972): Zur Verbreitung einigerFelseidechsen des Subgenus Archaeolacerta in der Tür-kei.- Bonner zool. Beitr.; 23 (4): 347-351.
DAREVSKY, I. S. & DANIELYAN, F. D. (1977):Lacerta uzzelli sp. nov. (Sauria, Lacertidae). A newparthenogenetic species of rock lizard from Eastern Tur-key, (in Russian).- Trudy Zool. Inst. Leningrad; 76: 55-59.
DAREVSKY, I. S. & KUPRIYANOVA, L. & Uz-ZELL, T. (1985): Parthenogenesis in reptiles. In: GANS,F. & BILLET, F. (eds.): Biology of the Reptilia. Vol 15(Development B): 411-526. New York, London (Acad-emic press)
DAREVSKY, I. S. & KUPRIYANOVA, L. A. & DA-NIELYAN, F. D. (1986): New evidence of hybrid malesof parthenogenetic lizards. In: ROCEK, Z. (ed.): Studiesin Herpetology, Prague, pp. 207-211.
DURFORT, M. (1978): Tècniques de transparen-tat d'invertebrats i d'esquelets de vertebrate: aplicacions.-Circ. Inst. Catalan Hist. Nat; 1:1-9.
ETHERIDGE, R. (1967): Lizard caudal vertebrae.-Copeia 1967 (4): 699-721.
HOFFSTETTER, R. & GASC, J. P. (1969): Verte-brae and ribs. In: GANS, C. & PARSONS, T. S. (eds.):Biology of the Reptilia. Vol. 1 (Morphology A): 201-310. New York, London (Academic press).
KLEMMER, K. (1957): Untersuchungen zur Oste-ologie und Taxonomie der europäischen Mauereidech-sen.- Abh. senckenberg. naturforsch. Ges., Frankfurt amMain; 496: 1-56.
KÜHNE, K. (1913): Über die Variationen derWirbelsäule und der Extremitätenplexus bei Lacertaviridis GESSN. und Lacerta agilis LlNN.- Morphol. Jb.;46: 563-592.
KUPRIYANOVA, L. A. (1989): Cytogenetic evi-dence for genome interaction in hybrid lacertid lizards,pp. 236-240. In: DAWLEY, R. M. & BOGART, J. P.
(eds.): Evolution and ecology of unisexual vertebrates. -Bull. New York State Mus<; 466.
MAYER, W. & BENYR, G. (1994): Albumin-Evolution und Phylogenese in der Familie Lacertidae(Reptilia: Sauria).- Ann. Naturhist Mus. Wien; (B) 96:621-648.
MAYER, W. & BISCHOFF, W. (1986): Beiträgezur taxonomischen Revision der Gattung Lacerta(Reptilia: Lacertidae). Teil 1: Zootoca, Omanosaura,Timon und Teira als eigenständige Gattungen.- Sala-mandra; 32 (3): 163-170.
MÉHELY, L. (1909): Materialien zu einer Syste-matik und Phylogenie der Muralis-ähnlichen Lacerten.-Ann. Hist Nat Mus. Natn., Budapest; 5: 84-88.
MORITZ, C. & UZZELL, T. & SPOLSKY, C. &HOTZ, H. & DAREVSKY, I. & KUPRIYANOVA, L. & DA-NIELYAN, F. (1992): The material ancestry and approxi-mate age of parthenogenetic species of Caucasian rocklizards (Lacerta : Lacertidae).- Genetica; 87: 53-62.
PEREZ MELLADO, V & BARBADILLO, L. J. &BARAHONA, F. & BROWN, R. P. & CORTL C. & GUER-RERO, F. & LANZA, B. (1993): A systematic survey ofthe Iberian Rock Lizard, Lacerta monticola. pp. 85-105.In: VALAKOS E. & BÖHME, W. & PEREZ MELLADO, V.& MARAGOU, P (eds.): Lacertids of the mediterraneanRegion. A biological approach. Athens, Bonn, Alicante(Hellenic Zool. Soc).
REPPEL, O. (1992): Studies on skeleton formation inreptiles. III. Patterns of ossification in the skeleton ofLacerta vivipara JACQUIN (Reptilia, Squamata).- Field-iana, Zoology. N.S.; 68: 1-25.
ROCEK, Z. (1980): Intraspecific and ontogeneticvariation of the dentition in the green lizard Lacertaviridis (Reptilia, Squamata).- Vëstnik CeskoslovenskéSpoleônosti. Zool.; 44: 272-277.
ROMER, A. S. (1956): Osteology of the reptiles.Chicago, London (University of Chicago press).
SIEBENROCK, F. (1894): Das Skelett der Lacertasimonyi und der Lacertidenfamilie überhaupt- Sitz.-ber.kais. Akad. Wiss. Wien, Math.-Naturwiss. Kl.; 103:205-292.
TAYLOR, W. R. (1967): An enzyme method ofClearing and staining small vertebrates.- Proc. U S Na-tional Mus., Smithsonian Inst; 122 (3596): 1-17.
UZZELL, T. & DAREVSKY, I. S. (1975): Bio-chemical evidence for the hybrid origin of the partheno-genetic species of the Lacerta saxicola complex (Sauna,Lacertidae), with a discussion of some ecological andevolutionary implications.- Copeia; 1975: 204-222.
DATE OF SUBMISSION: November 18th, 1997 Corresponding editor: Heinz Grillitsch
AUTHOR: Dr. OSCAR J. ARRIBAS, Department of Animal Biology (Zoology), Universität Autònoma de Bar-celona, E-08193, Bellaterra, Barcelona, Spain.
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at