Embed Size (px)
Citation preview

"Os ovários: Estrutura Anatômica e Fisiologia; Implicações
Clínicas das Hipo e Hiperfunções; Condutas Terapêuticas”.
“The ovaries: Anatomical structure and physiology; Clinical
implications of hypo and hyper-functions; Therapeutic conducts”.
Lucas Viana Machado
Faculdade de Ciências Médicas de MG
Rua Maranhão 1567 Apto 1401- Funcionários, CEP 30150/331
Fone/fax: 0XX-31-3282 1945
E-mail: [email protected]

Resumo
Após breve introdução, realçando a importância das ciências básicas e um
conhecimento razoável de medicina biomolecular, são abordados os tópicos referentes à
estrutura anatômica dos ovários, seus compartimentos funcionais e a produção de
esteroides sexuais própria de cada um. Segue uma rápida explanação sobre o conceito
unitário da bioesteroidogênese, com as implicações sobre as diversas glândulas
produtoras de hormônios esteroides. É, a seguir, abordada a dinâmica folicular nas
diversas etapas da vida da mulher e a interação folículo/eixo H-H-O, desde a vida intra-
uterina e infância, adolescência (com especial destaque para a ocorrência dos ovários
policísticos), menácme (salientando o papel potencial do oócito no desenvolvimento
folicular) e concluindo com as alterações fisiológicas do climatério e transição
menopausal. Encerra, enfatizando a dinâmica hormonal do climatério, cujo
conhecimento é indispensável ao se indicar uma eventual terapia hormonal.
Palavras-chave: Fisiologia ovariana, dinâmica folicular ovariana, feedback hormonal.

Abstract After a short introduction highlighting the importance of basic sciences and a
reasonable knowledge of molecular medicine, topics on the ovarian anatomical
structures, its functional compartments and their respective steroids production are
discussed. It follows a short explanation on the unitarian concept of biosteroidogenesis
and it`s implications on the steroids producing glands. It is, them, focused on the
follicular dynamics during the periods of the women’s life and the interaction follicle/H-
H-O axis, from intrauterine life and infancy, adolescence (with special attention on the
occurrence of polycystic ovaries), menácme and the potential role of the oocyte on the
follicular development, ending with the physiologic changes of the menopausal
transition. The paper are closed emphasizing the climacteric hormonal dynamics, whose
knowledge is essential in prescribing an eventual hormone therapy.
Key-Words: Ovarian Physiology, Ovarian Follicular Dynamics, Hormonal Feedback.

A fisiologia ovariana é um tema vasto e aparentemente difícil, por lidar
fundamentalmente com pesquisas de laboratório, experiências em animais e in vitro.
Envolve diretamente as ciências básicas e especialmente a medicina biomolecular, o
que, para muitos, são assuntos teóricos, complexos e distantes da prática clínica diária.
Tentaremos sintetizar os conhecimentos atuais enfatizando o que, no momento,
parece ser aceito pela maioria dos pesquisadores. Por ser um texto dirigido aos
ginecologistas, evitaremos, tanto quanto possível, entrar em detalhes técnicos,
bioquímicos e experimentais. Lembremos porém, que uma atualização não dispensa os
trabalhos pioneiros sobre os quais construímos os avanços atuais.
ESTRUTURA ANATÔMICA DOS OVÁRIOS.
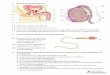
Destacam-se no ovário três regiões distintas: córtex, medula e hilo. A região
cortical, cuja porção periférica é denominada túnica albugínea, é revestida em sua
superfície por um epitélio de células cubóides denominado epitélio germinativo. Dele se
diferenciam as células da granulosa. Disseminados na córtex encontram-se os folículos
ovarianos envolvidos pelo estroma, que é formado por tecido conjuntivo e células
intersticiais derivadas do mesênquima embrionário. A medula ocupa a área central da
gônada e tem a sua origem junto com a córtex da supra-renal no blastema medial do
mesonefros. Ela fornece as células intersticiais que formarão a teca dos folículos. O
hilo é a região em que o ovário se liga ao mesovário. Ela contem nervos, vasos
sangüíneos e as células hilares, semelhantes às células de Leydig32.Na região hilar
concentram-se os remanescentes embrionários da medula da gônada primitiva, através
dos quais resultaria a diferenciação do testículo, caso o embrião fosse geneticamente
masculino. Estes remanescentes podem, eventualmente, originar tumores virilizantes do
ovário38.

Do ponto de vista funcional, o ovário pode ser dividido em três compartimentos
distintos: folicular, cujo principal produto de secreção são os estrogênios; corpo lúteo,
cujo principal produto é a progesterona e o estroma, fonte de produção dos
androgênios32.
O estroma ovariano tem sido objeto de investigação desde 1941, quando pela
primeira vez Smith35 relatou a ocorrência da chamada “hiperplasia do estroma cortical
ovariano”. A partir de então, o estroma deixou de ser um simples tecido conjuntivo de
sustentação dos elementos nobres do ovário para compartilhar com os mesmos em suas
atividades esteroidogênicas, mostrando tratar-se de tecido especializado complexo.
Um rápido resumo da origem dos elementos que compõem o ovário facilita a
compreensão das suas funções. As células germinativas tem origem extragonadal, sendo
identificadas em torno da 4ª semana de vida embrionária junto ao epitélio do saco
vitelino. Por meio de movimentos ameboides, migram para a crista genital para
completar a formação da gônada e induzir a sua diferenciação, que se inicia em torno da
6ª-7ª semana. São indispensáveis à organização gonadal pois sem elas não se
diferenciariam testículos ou ovários (uma das causas de disgenesia gonadal). As células
epiteliais que recobrem os corpos mesonéfricos e gônada primitiva darão origem às
células de Sertoli do testículo ou células da granulosa dos folículos ovarianos, enquanto
o mesênquima subjacente formará a medula da gônada da qual derivam as células
intersticiais de Leydig ou o estroma ovariano de onde se diferenciam as células da teca
dos folículos.
Tendo a mesma origem embriológica, ou seja, o mesênquima gonadal, não seria
estranho as células do estroma ovariano e as células intersticiais do testículo
compartilharem as mesmas potencialidades na síntese dos esteroides androgênicos. A
atividade secretora do estroma durante os ciclos ovulatórios normais é desprovida de

significância clínica perceptível, pela predominância estrogênica na fase folicular e da
associação estrogênio-progesterona na fase luteínica. É contudo nos distúrbios
anovulatórios, onde a maturação folicular é perturbada ou no ovário pós-menopausa,
que o estroma ovariano assume importância como fonte de esteroides capazes de
provocar manifestações clínicas detectáveis19.
ESTEROIDOGÊNESE OVARIANA
Quase todas as células do organismo são capazes de sintetizar o colesterol via
acetato, acetil coenzima-A, acido mevalônico, esqualeno, lanosterol e, finalmente
colesterol. Porém, somente determinadas células são capazes de reduzir a cadeia lateral
do colesterol, produzindo os esteróides pertencentes aos grupos do pregnano,
androstano e estrano. Estas células se encontram nas glândulas supra-renais, gônadas e
placenta. Embora as células destes órgãos sejam capazes de sintetizar o colesterol in
situ, via acetato, elas utilizam preferentemente o colesterol que lhes chega por via
sangüínea, através do LDL colesterol20.
A biosíntese dos esteroides foi extensamente estudada por Ryan31, e a maioria
dos autores segue, em linhas gerais, suas diretrizes. Este autor admite um conceito
unitário na esteroidogênese, no qual a supra-renal, ovário e testículo são capazes de
produzir as quatro classes de esteroides, ou seja, corticóides, progestogênios,
androgênios e estrogênios. O principal produto secretado por cada órgão é determinado
pela capacidade funcional das enzimas contidas dentro de suas células. Assim, o ovário
e o testículo não secretam mineralo e glicocorticoides porque suas células não possuem
as enzimas C-21 e C-11 beta hidroxilases. A supra-renal, por sua vez, não converte
normalmente androstenediona em testosterona nem aromatiza estes androgênios em
estrona e estradiol. Em situações anômalas, como nos tumores da supra-renal, este

órgão poderá produzir e secretar tanto testosterona como estradiol, indicando que os
sistemas de 17 beta redução e aromatização encontram-se inibidos na célula supra-renal
normal.
Durante a esteroidogênese, o número de átomos de carbono do esteroide poderá
ser reduzido, mas nunca aumentado, ou seja, os esteroides com 18 átomos de carbono
(estrogênios) derivam dos esteroides com 19 átomos de carbono (androgênios), que
derivam dos esteroides com 21 átomos de carbono (progestogênios), que derivam do
colesterol (27 átomos de carbono). Dependendo da concentração das enzimas, co-
fatores, substratos, receptores, estímulo gonadotrófico, corticotrófico e outros fatores, a
supra-renal e a gônada utilizarão diferentes vias a partir da pregnenolona para a síntese
dos corticóides, progestogênios, androgênios e estrogênios. Detalhes atualizados das
diversas etapas da esteroidogênese e as diversas enzimas nela envolvidas poderão ser
consultados no texto de Miller e Mellon27.
DINÂMICA FOLICULAR NAS DIVERSAS ETAPAS DA VIDA.
Toda fisiologia da reprodução está voltada para a liberação periódica de um
óvulo, cujo objetivo final é a perpetuação da espécie. Para que isto aconteça é necessária
uma função ovariana adequada, na qual a ovulação e a secreção de esteroides sexuais
ocorram de uma maneira regular e finamente sincronizada. Estes eventos dependerão da
presença de uma população folicular adequada, que obedecerá a estímulos específicos
de outros centros que integram o chamado eixo cortex-hipotálamo-hipófise-ovário. A
ovulação é desencadeada pela liberação do pico ovulatório do LH, que, por sua vez, será
induzido pelos níveis de estradiol circulantes secretados pelo folículo dominante, a
partir do momento em que eles reverterem o feed-back negativo para o LH em positivo.

É, portanto, o folículo dominante, mais precisamente o estradiol por ele secretado, que
irá reger a sincronização do ciclo reprodutivo21.
Para entendermos melhor a dinâmica folicular é necessário relembrarmos alguns
dados básicos. O ovário é um órgão dinâmico que nunca se encontra em repouso
absoluto. Desde a 20ª semana de vida intra uterina até a pós-menopausa tardia, ele
apresenta sinais de intensa e ininterrupta atividade. O fenômeno mais marcante desta
atividade é a maturação parcial e subsequente atresia folicular, e ela não depende da
presença de gonadotrofinas hipofisárias. Estas atuarão somente nos folículos
secundários, a partir do início da puberdade, quando a ovulação tornar-se o
epifenômeno da fisiologia reprodutiva feminina22. Portanto, a partir da 20ª semana da
vida intra-uterina até a menopausa, haverá sempre centenas de folículos em processo de
maturação parcial e outros tantos em regressão. No embrião de 20 semanas,
aproximadamente 7 milhões de células germinativas estarão presentes4 . Cada folículo
contem um oócito arrestado na profase da primeira divisão meiótica. Este pool de
folículos imaturos vai se reduzindo até que ao nascer, cada ovário terá de 266.000 a
472.000 folículos9. Por ocasião da menarca, eles se restringirão a um total aproximado
de 500.00012 . Durante o menácme, a depleção folicular ocorre numa média de 1.000
folículos/mês. Este processo se dá pela atresia ou pela entrada na fase de crescimento.
Esta média aumenta após a idade de 35 anos até a menopausa, quando o estoque de
folículos cairá para menos de 10034, 18. A imensa maioria dos folículos são consumidos
por um processo de morte celular programada denominada apoptose. Da população
folicular inicial, somente cerca de 400 folículos serão destinados atingir a maturação
completa e ovular.

INTERAÇÃO FOLÍCULO / EIXO HIPOTÁLAMO-HIPÓFISE-OVÁRIO NAS
DIVERSAS ETAPAS DA VIDA
VIDA INTRAUTERINA E INFÂNCIA
O eixo hipotálamo-hipófise torna-se funcionalmente ativo durante o segundo
trimestre da gestação, possibilitando a secreção das gonadotrofinas fetais sob o controle
do GnRH. Entretanto, o feedback negativo dos esteroides sobre o hipotálamo torna-se
operante próximo ao termo da gestação, causando a diminuição da secreção das
gonadotrofinas. Após o parto, o efeito supressivo dos esteroides placentários é
removido, e a secreção das gonadotrofinas é novamente ativado. Nos primeiros quatro
meses de vida, o LH sérico encontra-se em torno de 1 UI/l, mas os níveis de FSH estão
bem acima, excedendo aqueles observados na idade reprodutiva da mulher28. Estes
níveis elevados causam a ativação da gônada e uma elevação na secreção dos esteroides
ovarianos. Este efeito é, entretanto, de curta duração e quatro meses após, as
concentrações das gonadotrofinas e esteroides caem dramaticamente, atingindo níveis
basais aos 1-2 anos de idade. Na presença destes níveis extremamente baixos de
gonadotrofinas, a secreção de estradiol é suprimida. Apesar disto, a síntese dos
esteroides ovarianos permanece residualmente ativa, produzindo maiores concentrações
de estradiol (2.2 pmol/l) na meia infância feminina do que aquela observada no sexo
masculino (0.3 pmol/l)16.
Folículos medindo até 5mm de diâmetro são observados na infância, apesar dos
baixos níveis de gonadotrofinas, e o número de folículos maiores aumentam
significativamente após os 7 anos de idade6. Apesar de uma secreção noturna pulsátil,
de baixa amplitude e baixa freqüência de gonadotrofinas ocorrer na meia infância, as
concentrações séricas de FSH não são suficientes para sustentar o desenvolvimento
além deste estágio e estes folículos são fadados à atresia.

ADOLESCÊNCIA
Na puberdade, a remoção de um fator de restrição sobre o sistema nervoso
central ainda não definido (referido por alguns como gonadostato), possibilita a
completa reativação do gerador pulsátil do GnRH. À ultra-sonografia, os ovários
aparecem repletos de folículos medindo entre 5 e 12 mm, com escasso estroma entre
eles13. À medida que a puberdade progride, a amplitude da pulsatilidade das
gonadotrofinas e a produção dos hormônios esteroides aumentam, resultando no avanço
da maturação folicular. Antes de ocorrer a primeira ovulação, a hipófise terá que
desenvolver a habilidade de responder ao pico de estradiol circulante. Isto requer uma
diminuição na sensibilidade do sistema de feedback negativo para o LH, possibilitando
os níveis de GnRH e LH elevarem ao invés de caírem em resposta aos níveis crescentes
de estradiol. Assim, o estradiol exerce sempre um feedback negativo sobre o FSH, mas
condiciona uma ação bifásica em relação ao LH, ou seja, negativo em baixos níveis e
positivo em níveis elevados. O desenvolvimento desta resposta positiva sinaliza a
maturação do eixo cortex-hipotálamo-hipófise-ovário. Os ciclos ovulatórios regulares,
contudo, podem levar anos para se estabelecerem. No primeiro ano após a menarca, o
comprimento dos ciclos é longo e variado. Dosagens seriadas de progesterona
plasmática em adolescentes indicam que somente 15% dos ciclos são ovulatórios no
primeiro ano pós menarca, subindo para 41% no terceiro ano. Pelo sexto ano pós
menarca, 25% dos ciclos ainda são anovulatórios1-2. Segundo Metcalf, um terço de 209
mulheres entre 11 e 24 anos de idade permaneciam anovulatórias 5 anos após a
menarca26. A anovulação na adolescência parece ser um fenômeno limitado na maioria
dos casos e nos 9-12 anos subsequentes à menarca cerca de 80% das mulheres terão
ciclos ovulatórios regulares.

Obesidade e resistência periférica à insulina encontram-se associadas com um
prolongamento do período anovulatório na adolescência. Estas meninas também
mostram freqüentemente os ovários aumentados ao ultra-som, hiperandrogenismo
ovariano e LH sérico elevado3. Em algumas adolescentes estas anormalidades
endócrinas desaparecem espontaneamente, em outras elas persistem, levando ao quadro
de ovários policísticos.
Os dados acima nos levam a uma conclusão óbvia: se os ovários se encontram
em constante atividade desde a vida embrionária, exibindo uma maturação parcial de
folículos seguida de atresia, num dado momento, eles mostrarão invariavelmente
dezenas de folículos em estágios iniciais de desenvolvimento e outros tantos em
regressão. Na puberdade, enquanto não ocorrer a ovulação, eles terão um aspecto
policístico à ultra-sonografia. Não há nem necessidade da sua comprovação, bastam
estes conhecimentos básicos da fisiologia. Se a anovulação persistir por mais tempo,
eles manterão o mesmo aspecto, ou um pouco aumentados de tamanho, na dependência
da duração da anovulação e dos níveis de LH atuando sobre o estroma, daí, a equação:
anovulação = ovários policísticos e vice-versa23. Acrescente-se o fato de que as células
da teca interna, provenientes do estroma ovariano, possuem apenas receptores para o
LH, que por sua vez encontra-se aumentado na maioria destas pacientes. A elevação do
LH, que por si só já aumentaria a produção da testosterona e androstenediona pelo
estroma ovariano, atuaria associada à hiperinsulinemia, que potencializaria ainda mais a
produção destes androgênios. Qual o resultado inevitável desta situação? Anovulação
crônica, infertilidade, ovários policísticos e bilateralmente aumentados, hiperplasia do
estroma (hipertecose), hiperandrogenismo, freqüente obesidade, com eventual
surgimento de acanthosis nigricans. Este é o quadro que alguns autores ainda referem
como síndrome dos ovários policísticos, ou mais corretamente, anovulação

hiperandrogênica. É importante enfatizar que os ovários nestes casos encontram-se
absolutamente normais, tanto anatomicamente quanto fisiologicamente. Eles
simplesmente respondem de acordo com estímulos que atuam sobre os mesmos24.
Qualquer patologia ou interferência em qualquer nível do eixo córtex-hipotálamo-
hipófise-ovário poderá resultar em anovulação crônica e conseqüente formação de
ovários policísticos. A resistência periférica à insulina, embora freqüente, é apenas uma
delas. A sua importância é alertar-nos para as graves conseqüências metabólicas, caso
não sejam tomadas medidas terapêuticas adequadas.
MENÁCME
A maturação completa de um folículo primordial leva pelo menos 85 dias10. O
início do crescimento folicular ocorre continuamente e eles são selecionados
aleatoriamente. Entretanto, os fatores que determinam quando um folículo primordial
entrará em desenvolvimento e o levará a ovulação ou atresia não são ainda bem
conhecidos. O primeiro estágio da fase de crescimento envolve o aumento no tamanho
do oócito e proliferação das camadas de células da granulosa que o envolvem para
formar o folículo primário. Após desenvolver seu próprio suprimento sanguíneo, o
estroma peri-folicular se diferencia, formando a camada da teca interna. Este estágio de
desenvolvimento, denominado folículo secundário, é ainda caracterizado pela expressão
dos receptores para o LH nas células da teca interna7. As células da granulosa, por sua
vez, começam a expressar receptores para o FSH quando inicia a formação da cavidade
antral30. A duração do período de desenvolvimento do folículo primordial ao pré-antral
leva vários meses, e o período subsequente do desenvolvimento folicular, da fase antral
inicial ao folículo pre-ovulatório, leva aproximadamente 3 meses11. Apesar do
crescimento inicial do folículo ser regulado independentemente da estimulação

gonadotrófica, os estágios finais do desenvolvimento são FSH dependentes8. O número
de folículos que desenvolverão até o estagio pre-ovulatório é determinado pela duração
e magnitude do estímulo pelo FSH. Somente nos estágios mais avançados de
desenvolvimento folicular é que as células da granulosa tornam se sensíveis ao FSH e
são capazes de converter, através da ação das aromatases, a androstenediona e
testosterona produzidas pela teca em estradiol. O envolvimento dos dois tipos de células
(teca e granulosa) e dois hormônios (LH e FSH ) introduziu o conceito da “teoria das
duas células, duas gonadotrofinas” na produção dos estrogênios. Alem de estimular a
formação das aromatases, o FSH induz também a formação dos receptores de LH e um
maior aumento dos receptores do próprio FSH, ao mesmo tempo que estimula a síntese
de DNA, proteínas e receptores intracelulares dos esteroides sexuais14.
No ciclo ovariano normal, um folículo continuará desenvolvendo apesar da
queda do FSH, devido à sensibilidade aumentada pelo maior número de receptores de
FSH contidos nas células da granulosa do folículo dominante. Os demais folículos
daquele ciclo irão entrar em atresia, devido a queda do FSH.
A duração da elevação do FSH desempenha um papel crucial na determinação
do número de folículos que irão continuar o seu desenvolvimento. Foi proposto um
conceito da “janela do FSH”, acentuando a importância da duração da elevação do FSH
acima do limiar de estímulo, mais do que o nível da elevação do mesmo, para a seleção
de um único folículo dominante. Este conceito é substanciado pela demonstração de que
elevando-se os níveis de FSH bem acima do limiar de estímulo por um curto período de
tempo na fase folicular inicial, não aumenta o número de folículos dominantes.
Inversamente, quando a diminuição fisiológica do FSH em um ciclo normal é prevenida
pela administração de FSH na fase folicular tardia, a sensibilidade aumentada ao FSH
possibilitará vários folículos alcançarem a dominância33.

A diminuição dos níveis de FSH coincidindo com a seleção e desenvolvimento
do folículo dominante pode ser devido ao efeito de feedback negativo do estradiol sobre
o eixo hipotálamo-hipófise, coadjuvado pela grande elevação da inibina B, observada na
fase folicular. Este rápido aumento da inibina B ocorre logo após a elevação do FSH
entre os ciclos. Foi proposto que a inibina B limita a duração da elevação do FSH
(estreitando a janela do FSH) através do feedback negativo sobre a hipófise e pode ser
crucial para o desenvolvimento monofolicular36.
Apesar dos folículos vizinhos poderem exercer influencias recíprocas, o
crescimento de um folículo individual não é claramente sincronizado e vários folículos
no mesmo estágio de crescimento não terminam necessariamente da mesma maneira. O
número de folículos que iniciam o crescimento a cada ciclo está relacionado com o
número de folículos primordiais que permaneceram quiescentes até aquele momento.
Passado o período pré-natal e infância, onde a atividade ovariana é pontuada pela
redução do número de folículos através da apoptose, o ovário da adolescente responde
ao estímulo inicial do FSH através do crescimento de um número pequeno de folículos
antrais que escapam da atresia apoptótica. Estes folículos crescem sob o estímulo das
gonadotrofinas e estimulação hormonal local. Poucos folículos pré-antrais
eventualmente alcançam o estágio pré ovulatório e ovulam em resposta ao pico pré
ovulatório do LH. Após a liberação do oócito maduro, as células somáticas do folículo
se luteinizam e se transformam no corpo lúteo que secretará hormônios essenciais para a
manutenção da gravidez.
Papel potencial do oócito no desenvolvimento folicular
Os oócitos tem uma vida média prolongada; alguns podem sobreviver por
décadas antes de ovularem como nos folículos pre-ovulatórios, ou, mais provavelmente,

degenerarem como nos folículos atrésicos. Eles permanecem dormentes durante a maior
parte de suas vidas médias e encontram-se detidos no estágio dictiado da profase da
meiose até o início do pico pre-ovulatório das gonadotrofinas.
Durante o desenvolvimento folicular, a célula germinativa sofre um tremendo
crescimento, alcançando 100 vezes o seu volume inicial. Associado a este crescimento
ocorrem pronunciadas mudanças bioquímicas e morfológicas. O grande tamanho destas
células necessita, provavelmente, de uma simbiose metabólica com as células
foliculares vizinhas. Estudos anteriores demonstraram uma comunicação íntima entre o
oócito e as células vizinhas do “cumulus oophorus” através das “gap junctions”, que
permitem a troca de pequenas moléculas tais como o AMP cíclico e ions17. Estas “gap
junctions” são essenciais para o crescimento do oócito.
As células do cumulus que cercam o oócito são menos diferenciadas do que as
células altamente diferenciadas da granulosa próximas da lamina basal e mostram
menor produção de hormônios esteroides, bem como quantidades menores de receptores
para o LH. Este estado menos diferenciado das células do cumulus é provavelmente
necessária para um ótimo desenvolvimento do oócito, ao passo que as células
granulosas murais altamente diferenciadas, são essenciais para a esteroidogênese, bem
como, para a rotura folicular induzida pelo LH. Foi sugerido que fatores ovulares
podem inibir a diferenciação das células da granulosa e poderiam, portanto, ser
responsáveis pela formação de duas sub-populações de células da granulosa, do
cumulus e murais. Estudos morfológicos mostraram que a remoção do oócito leva à
luteinização das células da granulosa vizinhas e conseqüente produção de
progesterona29.
Inúmeros estímulos hormonais, fatores de crescimento locais e citocinas
exercem um papel importante no crescimento folicular e servem de base para a

formulação dos atuais protocolos de estimulação ovariana, nas tentativas de indução da
ovulação. O fator 9 de diferenciação de crescimento (GDF-9), um membro da família
dos fatores de crescimento de transformação (TGF beta), mostrou ser expresso
exclusivamente no oócito, sugerindo seu papel específico no desenvolvimento
folicular25. Estes achados indicam que o oócito, via secreção do GDF-9, pode favorecer
o crescimento folicular aumentando o número de células da granulosa, através da
inibição da diferenciação das células do cumulus que o cercam, levando a formação de
duas populações distintas de células da granulosa37.
CLIMATÉRIO/TRANSIÇÃO MENOPAUSAL
Apesar do esgotamento folicular ovariano ser o destino final do envelhecimento
reprodutivo, na realidade todo o eixo hipotálamo-hipófise-ovário está envolvido. O
envelhecimento de um órgão envolve um declínio na função fisiológica ao longo do
tempo.
A função primária do eixo H-H-O é a reprodução. Nas duas últimas décadas do
seu período reprodutivo, a mulher tem a sua fertilidade de leve a severamente
comprometida. As mudanças endócrinas ( na circulação e no folículo dominante) que
acompanham a diminuição do potencial de fertilidade são muito sutis até os
últimos estágios do envelhecimento reprodutivo. Em uma mulher ovulatória em seus
anos quarenta, a perda da fertilidade é muito mais dramática do que as alterações
endócrinas5. Nesta fase, o eixo H-H-O procura compensar esta perda da fertilidade
através do aumento do FSH, até que esta compensação não seja mais viável, devida a
quase total depleção folicular. O pensamento atual considera o oócito como a principal
causa da diminuição reprodutiva à medida que a mulher envelhece. Os oócitos não são
mais tão sadios, conforme refletido no aumento das aneuploidias, malformações

congênitas e dramático aumento de abortos espontâneos. Estas alterações ocorrem
quando há somente pequenas mudanças no ambiente endócrino.
Esgotado o pool folicular do ovário, a mulher entrará na menopausa, que será
precoce ou tardia, em função desta população folicular inicial .
Verifica-se que, com o correr dos anos, há uma diminuição progressiva
do número de folículos e consequentemente, um aumento relativo do estroma ovariano,
o que irá determinar as alterações da dinâmica hormonal do climatério. Às células da
teca interna dos folículos atrésicos, que são células diferenciadas do estroma ovariano,
se reincorporam ao mesmo e continuarão a secretar, sob estímulo do LH, os hormônios
que são próprios deste compartimento, ou seja, testosterona e androstenediona.
A redução numérica dos folículos resulta em gradual diminuição da
produção da inibina B (uma substância produzida pelas células da granulosa dos
folículos que atua especificamente inibindo a liberação do FSH hipofisário). A
diminuição da inibina B e consequentemente do efeito de feed-back negativo, resultará
na elevação do FSH, sendo esta, a primeira indicação laboratorial do climatério inicial.
O FSH elevado induzirá um desenvolvimento acelerado dos folículos,
provocando um encurtamento da primeira fase do ciclo menstrual, a primeira evidência
clínica do climatério inicial. À medida que a mulher se aproxima da menopausa, a
habilidade dos folículos em produzir níveis normais de estradiol e do corpo lúteo em
produzir progesterona são comprometidas. Neste estágio os ciclos tornam-se mais
irregulares e a fertilidade é praticamente nula. Cessadas as ovulações o LH começa a se
elevar, porém, sem atingir os níveis do FSH.
O conhecimento da fisiopatologia da “transição menopausal”, definida
por Burger como: o período da peri-menopausa compreendido entre o aparecimento das
primeiras irregularidades menstruais e a última regra – cuja duração média gira em

torno de quatro anos - é fundamental para se consolidar o diagnóstico de uma eventual
falência ovariana prematura ou orientar medidas terapêuticas visando a obtenção de
uma gestação.
Com relação à dinâmica folicular, verifica-se uma redução numérica dos
folículos primordiais de uma maneira mais ou menos constante, do nascimento até os 37
anos de idade, quando restarão cerca de 25.000 folículos ovarianos. Nos anos
subsequentes, até a menopausa, ocorrerá uma aceleração da depleção folicular. Por
ocasião da menopausa (última regra), eles estarão reduzidos a cerca de 1.000 folículos,
o que representaria o limite inferior necessário para manter os ciclos menstruais ou
apenas um resíduo de folículos funcionalmente incompetentes. Nos 2 a 3 anos
subseqüentes à menopausa, os folículos praticamente se esgotam, permanecendo
apenas alguns folículos degenerados, apresentando uma pobre camada de células da
granulosa, sem o “cumulus oophorus” e respectivo oócito e, eventualmente, alguns
cistos de inclusão, freqüentemente detectados à ultra-sonografia e sem nenhum
significado clínico-patológico.
A diminuição acelerada da população folicular após os 37 anos, irá
alterar a dinâmica hormonal do climatério, pois tanto o estradiol quanto a inibina são
produtos das células da granulosa dos folículos. Enquanto o folículo dominante é a
principal fonte do estradiol circulante, a inibina é produzida pelos demais folículos. É
interessante observar que a elevação do FSH começa em torno dos 40 anos, coincidindo
com o período de aceleração da atresia folicular. Como 90% do pico ovulatório de
estradiol é produzido pelo folículo dominante, enquanto a mulher estiver ovulando, não
haverá uma deficiência estrogênica, por conseguinte, a elevação do FSH não se deve à
falta do “feedback” negativo pela queda do estrogênio, mas reflete provavelmente a
diminuição acelerada do número de folículos e consequentemente da inibina. Tal

elevação do FSH pode, por sua vez, fornecer um estímulo adicional à maturação
folicular e manutenção do estradiol circulante em níveis fisiológicos, possibilitando a
ocorrência de ciclos ovulatórios até próximo aos 50 anos5.
Pelo exposto, admitindo-se que a secreção de inibina pelo folículo permaneça
inalterada, ela seria o marcador biológico mais sensível que o FSH na indicação do
envelhecimento ovariano. Contudo estes critérios podem ser enganosos, e a ciclicidade
algumas vezes pode retornar após longo período de refratariedade. O FSH é
fundamentalmente um indicador indireto da atividade secretora de estradiol dos
grandes folículos e não são afetados pelo número dos pequenos folículos, que estão
endocrinologicamente silenciosos.
O fato do ovário ter esgotado a sua população folicular e a mulher
entrado na menopausa não significa que não exista mais função ovariana. Na verdade o
ovário continua ativo, só que de uma maneira diferente daquela observada no menácme.
A menopausa sinaliza apenas o fim da função ovulatória (reprodutiva). Ao contrário do
que muitos imaginam, o ovário pós-menopausa não é um órgão falido, depositário
apenas das cicatrizes remanescentes de uma intensa atividade ovulatória durante o
menácme.
O exame histológico do ovário na pós-menopausa, revela um número
reduzido de folículos primordiais inativos. Folículos em fase de crescimento são raros,
porém, é comum encontrar alguns folículos císticos, sem o cumulus ooforus e
respectivas células germinativas, apresentando uma camada de células da granulosa
delgada e atípica e uma teca fibrosada. A histologia destes ovários sugere que a maioria,
senão todas as estruturas produtoras de estrogênios estão ausentes. Permanece apenas o
estroma, que é um tecido produtor de androgênios. Portanto, o ovário na pós-menopausa
continua ativo, secretando fundamentalmente testosterona e androstenediona.

Do ponto de vista clínico, é fundamental identificar o padrão endócrino da
paciente pós-menopausa, pois é exatamente este perfil que irá ditar a conduta clínica e
terapêutica.
Assim, segundo Howard Judd15, encontraremos um grupo de mulheres
que se apresentam hormonalmente equilibradas para a idade, sem manifestações clínicas
específicas. Nestas pacientes a terapia hormonal é questionável.
Um outro grupo apresentará manifestações clínicas que podem ser divididas em
três categorias:
1- Aquelas que apresentam sinais e sintomas de deficiência
estrogênica, como instabilidade vasomotora (fogachos, sudorese noturna,
palpitações, cefaléia), atrofia dos órgãos genitais (ressecamento vaginal,
dispareunia, sintomas urinários como frequência, urgência e incontinência) e
alterações da pele e esqueleto (osteoporose e perda de colágeno).
2- Aquelas que apresentam sinais e sintomas de excesso de
androgênios, como defeminização, hirsutismo e eventualmente virilização.
Estas são geralmente magras, pobres em tecido adiposo, mas podem também
ser obesas, o que, por outros mecanismos, favorece a hiperinsulinemia e
conseqüente estímulo do estroma ovariano a produzir androgênios, ao mesmo
tempo que diminui a síntese da SHBG, permitindo maior quantidade de
testosterona livre circulante.
3- Aquelas que apresentam sinais e sintomas paradoxais de excesso
de estrogênios, tais como sangramento uterino disfuncional, hiperplasia ou
adenocarcinoma do endométrio. São geralmente obesas e portanto, as que
apresentam maiores índices de aromatização.

É fundamental, portanto, identificarmos qual o padrão endócrino da paciente que
se apresenta diante de nós, para que possamos definir a necessidade ou não de uma
eventual terapia hormonal , bem como o tipo, via e esquema adequados.
LEITURAS SUPLEMENTARES
1. Apter D. Serum steroids and pituitary hormones in female puberty: a
partly longitudinal study. Clin Endocrinol 1980; 12: 107-20.
2. Apter D, Vihko R. Hormonal patterns of the first menstrual cycles. In:
Venturolli S, Flamigni C, Givens JR. Adolescence in Females. Chicago: Year Book
Medical Publishers, 1985. p. 215-38.
3. Apter D, Bützow T, Laughlin GA, et al. Metabolic features of polycystic
ovary syndrome are found in adolescent girls with hyperandrogenism. J Clin
Endocrinol Metab 1995; 80: 2966-73.
4. Baker TC. A quantitative and cytological study of germ cells in human
ovaries. Proc R Soc B. 1963; 158: 417-33.
5. Burger HG. The menopausal transition. In: Barlow DH. Bailliere`s
Clinical Obstetrics and Gynecology – The Menopause Key Issues. London: Bailliere
Tindall; 1996. p.347-59

6. Cacciatore B, Apter D, Althan H et al. Ultrasonic characteristics of the
uterus and ovaries in relation to pubertal development and serum LH, FSH and
estradiol concentrations. Adolesc Pediatr Gynecol 1991; 4: 15-20.
7. Channing CP, Kammerman S. Characteristics of the gonadotropin
receptors of porcine granulosa cells during folicle maturation. Endocrinology 1973;
92: 531-40.
8. Fauser BCJM, van Heusden AM. Manipulation of human ovarian
function: physiological concepts and clinical consequences. Endocr Rev 1997;18:
71-106.
9. Forabosco S. Sforza C, De Pol A, et al. Morphometric study of the
human neonatal ovary.Anat Rec 1991; 231: 201-08.
10. Gougeon A. Dynamics of follicular growth in the human: a model from
preliminary results. Hum Reprod 1986; 1: 81-87.
11. Gougeon A. Regulation of ovarian follicular development in primates:
facts and hypothesis. Endocr Rev 1996; 17: 121-55.

12. Himelstein-Braw R, Byskov AG, Peters H, Faber M. Follicular atresia in
the infant human ovary. J Rprod Fertil 1976; 46: 455-59.
13. Holm K, Mosfeldt Laursen E, Brocks V et al. Pubertal maturation of the
internal genitalia: an ultrasond evaluation of 166 healthy girls. Ultrasond Obstet
GYNECOL 1955; 6: 1 75-81.
14. Hsueh AJ, Bicsak TA, Jia XC, et al. Granulosa cells as hormone targets:
the role of biologically active follicle-stimulating hormone in reproduction. Recent
Prog Horm Res 1989; 45:209-73.
15. Judd H. Hormonal Dynamics associated with the menopause. Clin Obst
Gynecol 1976; 19: 775-88.
16. Klein K, Baron O, Colli MJ, et al. Estrogen levels in childhood
determined by na ultra-sensitive recombinant cell bioassay. J Clin Invest 1994; 6:
2475-80.
17. Lawrence TS, Beers WH, Gilula NB. Transmission of hormonal
stimulation by cell-to-cell communication. Nature 1979; 272: 501-6.
18. Linten-Moore Speters H, Moore GP et al. Follicular development in the
infant human ovary. J Reprod Fertil 1974; 39: 53-64.

19. Machado LV. O estroma ovariano e suas implicações clínicas. Femina
1976; 4: 680-85.
20. Machado LV. Bioesteroidogênese. Femina 1980; 8: 424-38.
21. Machado LV. Visão unitária da fisiopatologia ovariana. In: _______-
Endocrinologia Ginecológica. Rio de Janeiro: Medsi; 2000. p. 19-22.
22. Machado LV. Anovulação crônica/ovários policísticos: Causa e
consequencia. . In: _______- Endocrinologia Ginecológica. Rio de Janeiro: Medsi;
2000. p. 33-42.
23. Machado LV. Anovulação e ovários policisticos. Femina 1979; 7: 764-5.
24. Machado LV. Deixemos o ovário em paz II. Femina 1988; 16:1091-3.
25. McGrath AS, Esquela AF, Lee SJ. Oocyte-specific expression of
Growth/differentiation factor-9. Mol Endocrinol 1995; 9: 131-6.
26. Metcalf MG, Skidmore DS, LowryGF et al. Incidence of ovulation in the
years after the menarche. J Endocrinol 1983; 97: 213-19.

27. Miller WL, Mellon SH. Steroidogenesis. In: Fauser BCJM. Reproductive
Medicine. Molecular, Cellular, and Genetic Fundamentals. Boca Raton : Parthenon;
2003. p 245-66.
28. Neely EK, Hintz RL, Wilson DM et al. Normal rangesfor
immunochemiluminometric gonadotropin assays. J Pediatr 1995; 127: 40-6.
29. Nekola MV, Nalbandov AV. Morphological changesof rat follicular cells
as influenced by oocytes. Biol Reprod 1971; 4: 154-60.
30. Oktay K, Briggs D, Gosden RG. Ontogeny of follicle-stimulating
hormone receptor gene expression in isolated human ovarian follicles. J Clin
Endocrinol Metab 1997; 82: 3748-51.
31. Ryan KJ. Synthesis of Hormones in the Ovary. In: Grady HG, Smith DE.
The Ovary. International Academy of Pathology Monograph. Baltimore:Williams &
Wilkins Co; 1963. p 69-83.
32. Savard K.. The biogenesis of steroids in the human ovary. In: Mack HC.
The ovary. Proceedings of the Annual Symposium on the Physiology and Pathology
of Human Reproduction. Springfield: Charles C Thomas Publ; 1968. p.10-26
33. Schipper I, Hop WCJ, Fauser BCJM. The follicle-stimulating hormone
(FSH) threshold/window conceptexamined by differentialinterventions with
exogenous FSH during the follicular phase of the normal menstrual cicle: duration

rather than magnitude of FSH increse affects follicle development. J Clin
Endocrinol Med 1998; 83: 1292-8.
34. Schwartzman RA, Cidlowski J. Appoptosis: The biochemistry and
molecular biology of programed cell death. Endocr Rev 1993; 14:133-51.
35. Smith GVS. Carcinoma of the endometrium; review with results of the
treatment through 1935. N Engl J Med 1941; 225:608-15.
36. Van Santbrink EJP, Hop WC, van Dessel HJHM, et al. Decremental
follicle-stimulating hormone and dominant follicle development during the normal
menstrual cycle. Fertil Steril 1995; 64: 37-43.
37. Vitt VA, Hayashi M, Klein C. et al. Growth differentiation factor-9
stimulates proliferation but suppresses the follicle-stimulating hormone –induced
differentiation of cultured granulosa cells from small antral and preovulatory rat
follicles. Biol Reprod 2000; 62: 370-77.
38. Yeh J, Adashi EY. The ovarian life cycle. In: Yen SC, Jaffe RB, Barbieri
RL. Reproductive Endocrinology. 4th ed., Philadelphia: WB Saunders Co; 1999. p.
153-190.