Embed Size (px)
Citation preview

HORTSCIENCE 47(12):1832–1836. 2012.
Chilades pandava Damage among85 Cycas Species in a CommonGarden SettingThomas E. Marler1
Western Pacific Tropical Research Center, College of Natural and AppliedSciences, University of Guam, UOG Station, Mangilao, Guam 96923
Anders J. LindstromNong Nooch Tropical Botanical Garden, 34/1 Sukhumvit Hgw, Najomtien,Sattahip, Chonburi 20250, Thailand
L. Irene TerryWestern Pacific Tropical Research Center, College of Natural and AppliedSciences, University of Guam, UOG Station, Mangilao, Guam 96923
Additional index words. Cycadaceae, leaf traits, plant–arthropod interactions, plant defense,volatiles
Abstract. The extent of Chilades pandava Horsfield herbivory among 85 Cycadaceaespecies was determined by three evaluators in a common garden setting in Thailand toidentify patterns that may improve horticultural and conservation managementpractices. The significant differences in herbivory damage from this invasive lepidop-teran pest ranged 8.7-fold among the species. Phylogenetic sections of this monogenericcycad family did not correspond to the relative differences among the species, andcountry of nativity was also not informative for this purpose. We suggest the Cycas L.species that share native habitat with this butterfly or the closely related Theclinesthesonycha Hewitson are among the least damaged taxa when they are comingled with otherCycas species in a common landscape. Grouping the most damaged Cycas speciestogether in a managed landscape may reduce costs associated with plant protection. Theinclusion of non-native Cycas plants in gardens nearby native Cycas habitats carries thepotential of disrupting the delicate specialist relationship that native butterfly popula-tions have with host Cycas species.
Invasive arthropod herbivores compriseone of the greatest threats to cycad conserva-tion both in situ and ex situ. The lepidopteranChilades pandava (Plains Cupid Butterfly,Fig. 1B) is a specialist insect native to southernAsia that invaded Guam in recent years (Mooreet al., 2005), and it is among the invasivespecies that are collectively threatening theendemic Cycas micronesica K.D. Hill (Marlerand Lawrence, 2012). This insect depends on asustained close relationship with native Cycaswithin its indigenous range. It causes ex-tensive damage to Cycas populations withinhabitats in its invasive range like Guam. Thesefactors engender a horticultural conflict be-tween the need to protect Cycas species fromdamage within garden settings and within thebutterfly’s invasive range and the arguableneed to conserve its dependence on Cycasplants in situ.
An extensive ex situ germplasm collectionof Guam’s C. micronesica was established at
Nong Nooch Tropical Botanical Garden inThailand as part of our efforts to conserve thisendangered cycad. In this setting where thebutterfly is native, herbivory is highly hetero-geneous among comingled Cycas species.Some species like Cycas revoluta Thunb.(Fig. 1C) and C. micronesica (Fig. 1E) exhibitextensive, unsightly damage, whereas otherspecies exhibit minimal damage (Figs. 1D and1F). Management decisions for this situationwould benefit from understanding how thebutterfly relates to its native host Cycasspecies in natural habitats and how it relatesas a pest to novel Cycas host species in hor-ticultural settings.
Most of the 107 described Cycas species(Osborne et al., 2012) are being conserved inthe ex situ cycad germplasm collections inNong Nooch Tropical Botanical Garden. Pes-ticide protection of susceptible Cycas taxaresulting from this pest represents one of thegreatest costs for success of these conservationefforts. We used this germplasm collectiongrowing in a common garden setting to de-termine relative butterfly herbivory at thespecies level. Our objective was to then minethese data for linkages between resistance todamage and group traits that may reduceuncertainty in predictions of and exposemechanisms that explain disparity in butterflydamage.
Materials and Methods
Three evaluators conducted this survey toquantify the extent of damage to representa-tive field-planted Cycas species at NongNooch Tropical Botanical Garden in Thai-land. The survey included 85 species growingin homogeneous abiotic and insect pressureconditions. Two arthropods exhibit chronicpest pressures on the Cycas plants in thisgarden, Chilades pandava and Aulacaspisyasumatsui Takagi. The butterfly can feedon several tissue types, but for every sub-strate, it requires expanding tissue for ovipo-sitioning and caterpillar food. The variousgroups of Cycas species produce vegetativeflushes at varied times of the year, and someexotic Cycas taxa produce leaves throughoutthe year. These characteristics provide theresident butterfly population with a sustainedsupply of caterpillar food. Therefore, the pestpressures are relentless and constant through-out the year so that no matter when a Cycashas a flush of leaves, it is subjected to adultbutterfly ovipositioning. A. yasumatsui, thesecond resident Cycas pest in our garden, isan armored scale that infests any organ thatpresents live exposed surfaces. Unidentifiedbiological control organisms keep the wide-spread A. yasumatsui infestations from irrupt-ing. Confusing the plant damage imposed bythese two arthropods is not possible; therefore,identifying butterfly damage is unambiguous.
Individual plants were ranked for butterflydamage in Mar. 2012. We assigned 0 fora plant exhibiting no visible damage and 10for a plant with butterfly damage on everyleaf. A total of 673 plants in 85 species,representing all five Cycas sections, wereevaluated. A Poisson regression model wasfitted on log-transformed damage evaluationscores using SAS GLIMMIX procedure inVersion 9.3 (SAS Institute Inc., Cary, NC). AType III test of fixed main effects was usedto determine significance of evaluators andspecies. The damage differences among thespecies were determined using pairwise com-parison of least squares means on log scale.
Results and Discussion
The mean damage score among specieswas significant based on Type III test (P #0.0001) after removing the evaluator differ-ences. The log-transformed mean damagescores were back-transformed and presentedas a LSMEANs table (Table 1). Butterflydamage of the 85 Cycas species in this commongarden setting exhibited an overall mean of2.093, and mean damage ranged 8.7-foldamong the species (0.5947 to 5.1502). Thedamage scores for these species partitioned into18 overlapping groups. Clearly, whether Cycasplants succumb to or resist damage by chronicbutterfly pressure is governed by differencesexpressed at the species level.
Phylogeny. Cycas micronesica, C. petraeaA. Lindstr. & K.D. Hill, C. revoluta,C. seemannii A. Braun, C. tansachana K.D.Hill & S.L. Yang, and C. thouarsii R.Br. exGaudich. emerged as the most damaged
Received for publication 22 Aug. 2012. Acceptedfor publication 10 Oct. 2012.Support provided by U.S. Forest Service ProjectNo. 10-DG-11059702-095.We thank George Fernandez for statistical analyses.1To whom reprint requests should be addressed;e-mail [email protected].
1832 HORTSCIENCE VOL. 47(12) DECEMBER 2012

species and C. aculeata K.D. Hill & T.H.Nguyen, C. circinalis L., C. condaoensis K.D.Hill & S.L. Yang, C. pachypoda K.D. Hill,C. pruinosa Maconochie, C. taitungensis C.F.Shen, K.D. Hill, C.H. Tsou & C.J. Chen, C.tuckeri K.D. Hill, and C. wadei Merrillexhibited the least amount of damage amongthe 85 species (Table 1). Exploring character-istics that explain these two extreme groupsmay establish a foundation for understandingthe controlling mechanisms of the disparity indamage within the genus. Two obvious grouptraits to explore for this purpose are phyloge-netic sections and native biogeographic range.
The 107 species (Osborne et al., 2012)that comprise the Cycadaceae are classifiedinto five sections (Hill, 2004) based primarilyon morphological characters. Mean damagescore for these sections exhibited a 2-foldnon-significant range (Fig. 1A). The sectionAsiorientales, distinguished by tomentoseovules, provides an example of the immense
variation within each of these sections. Asior-ientales exhibited a mean score that wasgreater than the overall survey mean, con-tained C. taitungensis in the least damagedgroup, and C. revoluta in the most damagedgroup. These major Cycas sections do notadequately inform our understanding of whatunderlies the differences in butterfly damageamong the 85 species that we observed.
Our results do not broadly correspond withphylogeny. However, the non-significant lowerdamage scores in this garden survey for thespecies in the Wadeanae section indicate thatdirect bioassays may show this section to bemore resistant than the other sections. Further-more, one smaller group of species emerges ashighly damaged relative to the overall list ofCycas species. The Rumphiae subsection ofthe Cycas section (Hill, 2004) is distinguishedby spongy tissue within the seed sclerotesta.When fully developed, this tissue enablesoceanic seed dispersal (Dehgan and Yuen,
1983). The mean for the eight species fromthis subsection in our survey was 3.1281, andhalf of the Cycas species ranked as mostdamaged (Table 1) were from this smallsubsection. An interesting example that un-derscores these observations is the Philippineisland of Culion, where the strict endemic C.wadei (Wadeanae section without floatingseeds) and the widespread C. edentata deLaub. (Rumphiae subsection within Cycassection with floating seeds) live on the mainisland and its barrier islands (Lindstrom et al.,2008). Damage to C. edentata was three timesgreater than that to C. wadei, and it is rankedamong the top 10 most damaged species(Table 1). Although this efficient long-distanceseed dispersal mechanism enables coloniza-tion of remote islands to establish an extensiveindigenous range, dispersal to an uninfestedlocation may preclude or at least delay thebutterfly or any other specialist from establish-ing in the new location.
Fig. 1. Relationship of Cycas species with the specialist herbivore Chilades pandava. (A) Extent of herbivory damage for 85 Cycas species sorted into the fiveCycadaceae sections, mean ± SD. (B) Female butterfly ovipositioning eggs on Cycas revoluta leaf. (C) C. revoluta plant showing typical extensive butterflydamage to leaflets. Damage score�9. (D) C. pachypoda plant with undamaged leaves. Damage score 0. (E) Production beds of the susceptible C. micronesicashowing plant mortality and heterogeneous growth. (F) Production beds of the resistant C. chamaoensis showing homogeneous growth.
HORTSCIENCE VOL. 47(12) DECEMBER 2012 1833
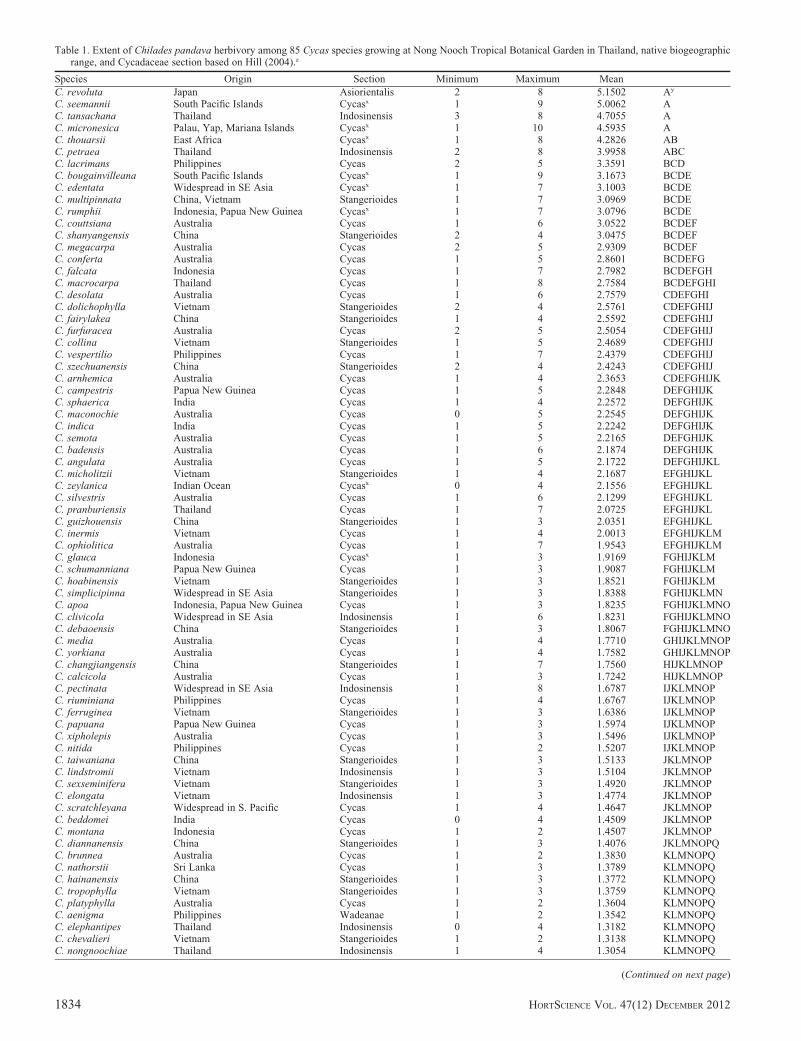
Table 1. Extent of Chilades pandava herbivory among 85 Cycas species growing at Nong Nooch Tropical Botanical Garden in Thailand, native biogeographicrange, and Cycadaceae section based on Hill (2004).z
Species Origin Section Minimum Maximum Mean
C. revoluta Japan Asiorientalis 2 8 5.1502 Ay
C. seemannii South Pacific Islands Cycasx 1 9 5.0062 AC. tansachana Thailand Indosinensis 3 8 4.7055 AC. micronesica Palau, Yap, Mariana Islands Cycasx 1 10 4.5935 AC. thouarsii East Africa Cycasx 1 8 4.2826 ABC. petraea Thailand Indosinensis 2 8 3.9958 ABCC. lacrimans Philippines Cycas 2 5 3.3591 BCDC. bougainvilleana South Pacific Islands Cycasx 1 9 3.1673 BCDEC. edentata Widespread in SE Asia Cycasx 1 7 3.1003 BCDEC. multipinnata China, Vietnam Stangerioides 1 7 3.0969 BCDEC. rumphii Indonesia, Papua New Guinea Cycasx 1 7 3.0796 BCDEC. couttsiana Australia Cycas 1 6 3.0522 BCDEFC. shanyangensis China Stangerioides 2 4 3.0475 BCDEFC. megacarpa Australia Cycas 2 5 2.9309 BCDEFC. conferta Australia Cycas 1 5 2.8601 BCDEFGC. falcata Indonesia Cycas 1 7 2.7982 BCDEFGHC. macrocarpa Thailand Cycas 1 8 2.7584 BCDEFGHIC. desolata Australia Cycas 1 6 2.7579 CDEFGHIC. dolichophylla Vietnam Stangerioides 2 4 2.5761 CDEFGHIJC. fairylakea China Stangerioides 1 4 2.5592 CDEFGHIJC. furfuracea Australia Cycas 2 5 2.5054 CDEFGHIJC. collina Vietnam Stangerioides 1 5 2.4689 CDEFGHIJC. vespertilio Philippines Cycas 1 7 2.4379 CDEFGHIJC. szechuanensis China Stangerioides 2 4 2.4243 CDEFGHIJC. arnhemica Australia Cycas 1 4 2.3653 CDEFGHIJKC. campestris Papua New Guinea Cycas 1 5 2.2848 DEFGHIJKC. sphaerica India Cycas 1 4 2.2572 DEFGHIJKC. maconochie Australia Cycas 0 5 2.2545 DEFGHIJKC. indica India Cycas 1 5 2.2242 DEFGHIJKC. semota Australia Cycas 1 5 2.2165 DEFGHIJKC. badensis Australia Cycas 1 6 2.1874 DEFGHIJKC. angulata Australia Cycas 1 5 2.1722 DEFGHIJKLC. micholitzii Vietnam Stangerioides 1 4 2.1687 EFGHIJKLC. zeylanica Indian Ocean Cycasx 0 4 2.1556 EFGHIJKLC. silvestris Australia Cycas 1 6 2.1299 EFGHIJKLC. pranburiensis Thailand Cycas 1 7 2.0725 EFGHIJKLC. guizhouensis China Stangerioides 1 3 2.0351 EFGHIJKLC. inermis Vietnam Cycas 1 4 2.0013 EFGHIJKLMC. ophiolitica Australia Cycas 1 7 1.9543 EFGHIJKLMC. glauca Indonesia Cycasx 1 3 1.9169 FGHIJKLMC. schumanniana Papua New Guinea Cycas 1 3 1.9087 FGHIJKLMC. hoabinensis Vietnam Stangerioides 1 3 1.8521 FGHIJKLMC. simplicipinna Widespread in SE Asia Stangerioides 1 3 1.8388 FGHIJKLMNC. apoa Indonesia, Papua New Guinea Cycas 1 3 1.8235 FGHIJKLMNOC. clivicola Widespread in SE Asia Indosinensis 1 6 1.8231 FGHIJKLMNOC. debaoensis China Stangerioides 1 3 1.8067 FGHIJKLMNOC. media Australia Cycas 1 4 1.7710 GHIJKLMNOPC. yorkiana Australia Cycas 1 4 1.7582 GHIJKLMNOPC. changjiangensis China Stangerioides 1 7 1.7560 HIJKLMNOPC. calcicola Australia Cycas 1 3 1.7242 HIJKLMNOPC. pectinata Widespread in SE Asia Indosinensis 1 8 1.6787 IJKLMNOPC. riuminiana Philippines Cycas 1 4 1.6767 IJKLMNOPC. ferruginea Vietnam Stangerioides 1 3 1.6386 IJKLMNOPC. papuana Papua New Guinea Cycas 1 3 1.5974 IJKLMNOPC. xipholepis Australia Cycas 1 3 1.5496 IJKLMNOPC. nitida Philippines Cycas 1 2 1.5207 IJKLMNOPC. taiwaniana China Stangerioides 1 3 1.5133 JKLMNOPC. lindstromii Vietnam Indosinensis 1 3 1.5104 JKLMNOPC. sexseminifera Vietnam Stangerioides 1 3 1.4920 JKLMNOPC. elongata Vietnam Indosinensis 1 3 1.4774 JKLMNOPC. scratchleyana Widespread in S. Pacific Cycas 1 4 1.4647 JKLMNOPC. beddomei India Cycas 0 4 1.4509 JKLMNOPC. montana Indonesia Cycas 1 2 1.4507 JKLMNOPC. diannanensis China Stangerioides 1 3 1.4076 JKLMNOPQC. brunnea Australia Cycas 1 2 1.3830 KLMNOPQC. nathorstii Sri Lanka Cycas 1 3 1.3789 KLMNOPQC. hainanensis China Stangerioides 1 3 1.3772 KLMNOPQC. tropophylla Vietnam Stangerioides 1 3 1.3759 KLMNOPQC. platyphylla Australia Cycas 1 2 1.3604 KLMNOPQC. aenigma Philippines Wadeanae 1 2 1.3542 KLMNOPQC. elephantipes Thailand Indosinensis 0 4 1.3182 KLMNOPQC. chevalieri Vietnam Stangerioides 1 2 1.3138 KLMNOPQC. nongnoochiae Thailand Indosinensis 1 4 1.3054 KLMNOPQ
(Continued on next page)
1834 HORTSCIENCE VOL. 47(12) DECEMBER 2012

Biogeography. Cycas is a widespread ge-nus comprised of species that range geo-graphically from strict endemics to somewidespread species (Table 1). Native geo-graphic range at the country or regional levelis also not useful for explaining the differ-ences that we found among these 85 species.Although the gross geographic region of ori-gin does not appear to be useful for under-standing the differences in butterfly damageamong Cycas species, there may be explana-tions for the diversity in damage noted withina country or region. We propose that identi-fying the Cycas species possessing a nativehabitat that is shared with the butterfly may beinformative. Four subspecies of this cycadspecialist Lycaenidae species are currentlyrecognized. Chilades pandava lanka is knownfrom Sri Lanka, Chilades pandava peripatriaoccurs exclusively in Taiwan, Chilades pan-dava vapanda occurs in the Philippines, andthe widespread Chilades pandava pandava isfound in mainland China and Southeast Asia(Hsu, 2002; Igarashi and Fukuda, 2000; Wuet al., 2010). The precise genotype of thepopulation we studied at Nong Nooch Trop-ical Botanical Garden is not known but islikely an admixture of Chilades pandavapandava introductions from numerous loca-tions. The closely related Lycaenidae special-ist Theclinesthes onycha occurs in Australia(Braby, 2004; Forster and Machin, 1994). Theeight Cycas species that assemble as the leastdamaged species (Table 1) occur within theseprescribed geographic ranges.
For the Australian Cycas species exhibit-ing greater than average butterfly damage inour survey, these may occur outside theconstrictions of Queensland and New SouthWales that comprise the contiguous nativerange of T. onycha (Braby, 2004). Further-more, any evolved reactions of a Cycas spe-cies to T. onycha herbivory may not translateto how that Cycas reacts to a different genus orspecies of herbivore. A full understanding ofwhy some Cycas species are susceptible tobutterfly damage despite their nativity withinknown butterfly range is more elusive. Anexample is C. tansachana, which is heavilydamaged by butterfly relative to the entire listof Cycas (Table 1). Cycas tansachana is
endemic to a small limestone outcrop withextremely high surface temperatures. Wesuspect that this butterfly cannot thrive in thishabitat despite its location within the indige-nous range for the butterfly. The details of fieldconditions to validate these habitat-specificrelationships will require direct observationsof butterfly behavior in situ.
Mechanisms and caveats. The specificityof lepidopteran larvae to feed on particularhost plant genotypes is well studied (Ehrlichand Raven, 1964). We have discussed poten-tial underlying mechanisms that explain thedisparity in butterfly feeding behavior in re-lation to Cycas species (Marler et al., 2012).One mechanism may be an information-mediated system that is founded in differencesin constitutive or induced leaf volatiles thatsearching female butterflies may use as adver-tisements for suitable tissue for larval devel-opment (Bernays and Chapman, 1994). Thismechanism could be tested with ovipositionchoice tests whereby reproductive female but-terflies are simultaneously provisioned withexpanding leaves of least-damaged vs. most-damaged Cycas species. An alternative mech-anism may be a resource-mediated system thatis founded in differences among the Cycasspecies for leaf palatability and defense traits.This mechanism may be expressed within themost damaged species as greater abundance ordiversity of phagostimulants for feeding cater-pillars and/or slow development of leaf tough-ness as leaves expand. Alternatively, it may beexpressed within the least damaged species asconstitutive or induced plant substances thatinhibit caterpillar feeding and/or rapid leafgrowth out of the soft, vulnerable tissue stage.Relevance of this resource-mediated mecha-nism could be tested with caterpillar bioassaysthat determine survival and growth rates amongcaterpillars reared on expanding leaf tissue ofvarious Cycas species. We note that manyspecies have individuals exhibiting no dis-cernible butterfly damage (indicated by 0 inTable 1) despite high-density butterfly popula-tion pressure and our observation of oviposi-tioning on all Cycas species. Therefore, theseassays should include first instar performancebecause this stage is highly vulnerable for manyLepidoptera species (Zalucki et al., 2002).
The extant literature illuminates severalissues that deserve mention as possible rele-vant factors for interpreting the results of ourstudy. For this Cycas–Chilades system, wehave mentioned (Marler et al., 2012) thattritrophic relations may partly mediate therelationship of the herbivore with the Cycasplant (e.g., Harrewijn et al., 1994). Activityof parasitoids or other natural enemies ofbutterfly eggs or caterpillars may interplay toexplain the highly heterogeneous extent ofherbivory at the Cycas species level. Inaddition, the step of ovipositioning by itselfmay suppress emission of both constitutiveand herbivore-induced volatiles in someplants (Penaflor et al., 2011). Therefore,disparity in timing of ovipositioning amongCycas species within close proximity mayinfluence the attraction of subsequent gravidfemale butterflies. Moreover, genotype ofneighboring plants may influence herbivoreabundance on focal plants (Genung et al.,2012). Consequently, herbivory of an indi-vidual Cycas tree within a common gardensetting may be influenced by layout of thespecies within the planting. Our understand-ing of how these sorts of interactions affectplant–pest relations in cycad biology is nil.
The current literature is also revealing moreabout belowground and aboveground interac-tions in relation to arthropod herbivory. Forexample, leaf herbivory may directly influenceroot microbiota (Landgraf et al., 2012), yet rootmicrobiota may directly influence abovegroundvolatile infochemicals produced by plants(Pineda et al., 2012; Schausberger et al.,2012). The Cycas plant rhizosphere is char-acterized by three-way relations among plantroots, mycorrhizae, and cyanobacteria (Fisherand Vovides, 2004; Muthukumar and Udaiyan,2002; Norstog and Nicholls, 1997). Little isknown about the genotype diversity and func-tion of the microbiome of Cycas in a commongarden setting. Preliminary work indicates thatcyanobacteria genotype within each coralloidroot complex is homogeneous, but within oneplant’s root system, each coralloid root can becolonized by different genotypes (Costa et al.,1999). The relationship among an in situpopulation of the butterfly, its host Cycaspopulation, and the natural root symbiont
Table 1. (Continued) Extent of Chilades pandava herbivory among 85 Cycas species growing at Nong Nooch Tropical Botanical Garden in Thailand, nativebiogeographic range, and Cycadaceae section based on Hill (2004).z
Species Origin Section Minimum Maximum Mean
C. balansae Vietnam Stangerioides 1 2 1.2935 KLMNOPQC. cairnsiana Australia Cycas 1 2 1.2931 KLMNOPQC. panzhihuaensis China Asiorientalisw 0 3 1.2769 LMNOPQC. chamaoensis Thailand Indosinensis 1 3 1.2647 MNOPQC. tuckeri Australia Cycas 1 2 1.1559 MNOPQRC. pruinosa Australia Cycas 1 2 1.1144 NOPQRC. circinalis India Cycas 1 2 1.0967 OPQRC. wadei Philippines Wadeanae 1 2 1.0598 OPQRC. pachypoda Vietnam Indosinensis 0 2 1.0592 PQRC. condaoensis Vietnam Indosinensis 0 2 0.9792 PQRC. taitungensis Taiwan Asiorientalis 0 1 0.8389 QRC. aculeata Vietnam Stangerioides 0 1 0.5947 RzMeans of three evaluators, 0 = no damage, 10 = extensive damage.yMeans with the same letter are not significantly different.xSubsection Rumphiae.wRecent evidence supports change to Stangerioides section.
HORTSCIENCE VOL. 47(12) DECEMBER 2012 1835

populations is not influenced by anthropo-genic introductions of non-native organisms.
The caveats discussed here illuminatesome of the challenges involved in interpret-ing research conducted in botanic gardensettings. For example, a botanic garden inwhich leaf damage by the butterfly is highlyheterogeneous among closely planted Cycasindividuals provides a situation in whichthese feedback loops between abovegroundand belowground relations may become a partof the mechanisms that foster sustaineddisparity in foliar damage among the Cycasspecies. Moreover, fertilization and irrigationregimes in managed landscapes may influ-ence the relations of cultivated Cycas plantsin a manner that deviates from that in naturalhabitats. In the current study, microsite var-iation throughout the garden and disparity inage and size of the trees are among otherpotentially confounding factors.
We have focused exclusively on oneLycaenidae pest in this discussion, but in thereal world, a cycad plant is attacked bymultiple herbivores. In Guam, for example,C. micronesica is threatened by a growing listof invasive and native specialist arthropodsliving in sympatry (Marler and Muniappan,2006; Marler et al., 2011). The armored scaleA. yasumatsui deserves mention as a rapidlyinvading pest that infests Cycas populationsthroughout much of the native and invasiveChilades pandava range. Most Cycas trees inThailand, including within our garden setting,experience minor infestations of this scalepest. An understanding of butterfly herbivorycannot be fully attained if the influences of A.yasumatsui and other pests are ignored in theprocess. For example, Gomez et al. (2012)used individual and combined attack of Tsugacanadensis (L.) Carriere by a scale insect anda gall-forming insect to show that feeding byone arthropod herbivore altered the tissuequality for and damage by the other herbivore.
Horticulture and conservation. Nativeand invasive populations of Chilades pan-dava affect horticultural decisions in severalways. First, the aesthetic appeal of suscepti-ble Cycas trees can be entirely removed bythe damage imposed on susceptible species(Fig. 1C). In some locations this pest cannotbe ignored by landscape managers. Second,the desired homogeneous growth ratesneeded to reduce costs in any horticulturalproduction system are difficult to achieve forthe susceptible Cycas species when this pestis present (Fig. 1E). The growth rates ofresistant Cycas species are homogeneousamong plants despite chronic butterfly pres-sure (Fig. 1F). These two factors underscorethe importance of understanding the biologyof the pest for the best horticultural outcomesand defining the most appropriate means ofcontrol. Third, horticulturists would do wellto acknowledge their role in conservation,and this case study provides a good exampleof the complications in fulfilling that role.Introducing non-native Cycas trees into land-scapes close to Cycas habitats, especially
those of endemic Cycas species, has thepotential of disrupting the specialist depen-dence of the habitat’s natural butterfly pop-ulation on its co-occurring Cycas species. Forexample, Wu et al. (2010) reported that hor-ticultural plantings of C. revoluta in Taiwanlead to irruptions of the native Chiladespandava that in turn increased the threatsto the endemic, endangered C. taitungensispopulations. Cycad conservation plans maybe more effective if the role of cycad trees inthe life histories of other native organisms isprominently figured into the plans.
Expanding leaves on all Cycas plants ina garden may be frequently sprayed with in-secticides for protection against butterfly dam-age as one means of managing the pest. AtNong Nooch Tropical Botanical Garden, theseexpensive sprays are applied at weekly in-tervals. Our results and field observations in-dicate that one means of improving efficiencywould be to eliminate a fixed spray regime onthe most resistant Cycas species. If the Cycasspecies comprising the least damaged taxacould be positioned together in the gardenlandscape, then this section of the garden couldbe avoided during sprays applied on a fixedschedule. This may result in substantial re-duction in pesticide costs and would also allowlimited human resource effort to focus moreheavily on protecting the susceptible species.
Some or all of the disparate mechanismsand caveats that we discussed here maymediate the relationships between Chiladespandava and comingled Cycas species ina garden landscape. The real situation likelyis a conflation of multiple mechanisms.
Literature Cited
Bernays, E.A. and R.F. Chapman. 1994. Host-plantselection by phytophagous insects. Chapmanand Hall, New York, NY.
Braby, M.F. 2004. The complete field guide tobutterflies of Australia. CSIRO Publishing,Collingwood, Australia.
Costa, J.-L., P. Paulsrud, and P. Lindblad. 1999.Cyanobiont diversity within coralloid roots ofselected cycad species. FEMS Microbiol. Ecol.28:85–91.
Dehgan, B. and C.K.K.H. Yuen. 1983. Seed mor-phology in relation to dispersal, evolution andpropagation of Cycas L. Bot. Gaz. 144:412–418.
Ehrlich, P.R. and P.H. Raven. 1964. Butterflies andplants: A study in coevolution. Evolution 18:586–608.
Fisher, J.B. and A.P. Vovides. 2004. Mycorrhizaeare present in cycad roots. Bot. Rev. 70:16–23.
Forster, P.I. and P.J. Machin. 1994. Cycad hostplants for Lilioceris nigripes (Fabricius) (Co-leoptera: Chrysomelidae) and Theclinesthesonycha (Hewitson) (Lepidoptera: Lycaenidae).Aust. Entomol. 21:99–102.
Genung, M.A., G.M. Crutsinger, J.K. Bailey, J.A.Schweitzer, and N.J. Sanders. 2012. Aphid andladybird beetle abundance depend on the in-teraction of spatial effect and genotypic di-versity. Oecologia 168:167–174.
Gomez, S., C.M. Orians, and E.L. Preisser. 2012.Exotic herbivores on a shared native host: Tissuequality after individual, simultaneous, and se-quential attack. Oecologia 169:1015–1024.
Harrewijn, P., A.K. Minks, and C. Mollema. 1994.Evolution of plant volatile production ininsect–plant relationships. Chemoecology 5:55–73.
Hill, K.D. 2004. Character evolution, species rec-ognition and classification concepts in the Cyca-daceae, p. 23–44. In: Walters, T. and R. Osborne(eds.). Cycad classification: Concepts and rec-ommendations. CABI Publishing, Cambridge,MA.
Hsu, Y.F. 2002. Butterflies of Taiwan. NationalFonghuanggu Bird Park, Nantou, Taiwan.
Igarashi, S. and H. Fukuda. 2000. The life historiesof Asian butterflies. Tokai University Press,Tokyo, Japan.
Landgraf, R., R.S. Schaarschmidt, and B. Hause.2012. Repeated leaf wounding alters the colo-nization of Medicago truncatula roots by ben-eficial and pathogenic microorganisms. PlantCell Environ. 35:1344–1357.
Lindstrom, A.J., K.D. Hill, and L.C. Stanberg.2008. The genus Cycas (Cycadaceae) in thePhilippines. Telopea (Syd.) 12:119–145.
Marler, T.E. and J.H. Lawrence. 2012. Demogra-phy of Cycas micronesica on Guam followingintroduction of the armoured scale Aulacaspisyasumatsui. J. Trop. Ecol. 28:233–242.
Marler, T.E., A. Lindstrom, and L.I. Terry. 2012.Information-based or resource-based systemsmay mediate Cycas herbivore interactions.Plant Signal. Behav. 7:760–762.
Marler, T.E. and R. Muniappan. 2006. Pests ofCycas micronesica leaf, stem, and male re-productive tissues with notes on current threatstatus. Micronesica 39:1–9.
Marler, T.E., L.S. Yudin, and A. Moore. 2011.Schedorhinotermes longirostris (Isoptera: Rhi-notermitidae) on Guam adds to assault on theendemic Cycas micronesica. Fla. Entomol. 94:699–700.
Moore, A., T.E. Marler, R. Miller, and R. Muniappan.2005. Biological control of cycad aulacaspis scaleon Guam. The Cycad Newsletter 28:6–8.
Muthukumar, T. and K. Udaiyan. 2002. Arbuscu-lar mycorrhizas in cycads of southern India.Mycorrhiza 12:213–217.
Norstog, K.J. and T.J. Nicholls. 1997. The biologyof the cycads. Cornell Univ. Press, Ithaca, NY.
Osborne, R., M. Calonje, K. Hill, L. Stanberg, andD.W. Stevenson. 2012. The world list ofcycads. Mem. N. Y. Bot. Gard. 106:480–508.
Penaflor, M.F., M. Erb, C.A. Robert, L.A. Miranda,A.G. Werneburg, F.C. Dossi, T.C. Turlings,and J.M. Bento. 2011. Oviposition by a mothsuppresses constitutive and herbivore-inducedplant volatiles in maize. Planta 234:207–215.
Pineda, A., R. Soler, B.T. Weldegergis, M.M.Shimwela, J.J.A. van Loon, and M. Dicke.2012. Non-pathogenic rhizobacteria interferewith the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid sig-naling. Plant Cell Environ. (in press).
Schausberger, P., S. Peneder, S. J}urschik, and D.Hoffmann. 2012. Mycorrhiza changes plantvolatiles to attract spider mite enemies. Funct.Ecol. 26:441–449.
Wu, L.-W., S.-H. Yen, D.C. Lees, and Y.-F. Hsu.2010. Elucidating genetic signatures of na-tive and introduced populations of the CycadBlue, Chilades pandava to Taiwan: A threatboth to Sago Palm and to native Cycas popu-lations worldwide. Biol. Invasions 12:2649–2669.
Zalucki, M.P., A.R. Clarke, and S.B. Malcolm.2002. Ecology and behavior of first instar larvalLepidoptera. Annu. Rev. Entomol. 47:361–393.
1836 HORTSCIENCE VOL. 47(12) DECEMBER 2012





























