Embed Size (px)
Citation preview

Int J Clin Exp Med 20169(7)12696-12704wwwijcemcom ISSN1940-5901IJCEM0021980
Original ArticleCurcumin prevents adriamycin-induced nephropathy MCP-1 expression through blocking histone acetylation
Jian Liu Fang Zhong Qin Dai Lili Xu Weiming Wang Nan Chen
Department of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China
Received December 15 2015 Accepted May 5 2016 Epub July 15 2016 Published July 30 2016
Abstract Objective To investigate the anti-MCP-1 expression effect of curcumin in kidney disease model and the possible mechanisms associated with histone acetylation Methods ADR nephropathy was induced in male SD rats by twice intravenous injections of ADR at day 0 (5 mgmiddotkg-1) and 1 week later (25 mgmiddotkg-1) Curcumin treatment was started at week 6 EP300 and MCP-1 mRNA transcription were assessed by real-time PCR The acetylation levels of the histones H3 and H4 in the promoters of MCP-1 were detected by ChIP-qPCR And mouse podocytes were cultured to investigate the effect of curcumin against ADR-induced injury Results In animal model curcumin inter-vention attenuated renal glomerulosclerosis and partially decreased transcription of EP300 and MCP-1 (P lt 005) mRNA Also curcumin prevented an increase of acetylation of MCP-1 promoters H3 and H4 in ADR-nephropathy In vitro cultured podocytes compared with the untreated ADR group pretreatment with low concentration of curcumin preserved podocyte cytoskeleton Curcumin significantly reduced EP300 and MCP-1 mRNA (P lt 001) Besides curcumin prevented ADR-induced MCP-1 promoter hyper-acetylation in ADR injured podocytes by ChIP-qPCR Conclusion Curcumin prevents kidney fibrosis and decreases MCP-1 expression through modification of histone acetylation and could be used as a therapy for patients with kidney disease
Keywords Curcumin renal fibrosis inflammation podocyte histone acetylation
Introduction
Renal fibrosis is a complicated and important pathological change in the development of kid-ney disease A variety of factors including the transforming growth factor-β pathway epitheli-al-mesenchymal transition and inflammation are involved in this process [1] Histone acetyla-tion is a recognized histone modification that can regulate the production of certain pro- teins and has spurred considerable interest [2 3] Studies have shown that histone activa-tion participates in a transforming growth fa- ctor-β-induced kidney injury and streptozotocin (STZ)-induced diabetic nephropathy model [4]
Curcumin is a compound purified from the rhi-zomes of the plant Curcuma longa L [5] Cur- cumin has been demonstrated to have an- ti-inflammatory antioxidant anti-proliferation anti-bacterial and anti-cancer activities [6] It could also protect against lipopolysaccharide- induced kidney injury by way of anti-inflamma-tion [7] In addition curcumin is a histone acet-ylase inhibitor [8]
Studies have confirmed that inflammation take part in the progression of this kidney disease in a ADR-induced glomerulosclerosis model [9] However the role of histone modification of curcumin in glomerular disease is still un- clear and its effect on MCP-1 expression also unknown Thus we utilized the ADR-induced nephropathy and evaluated the effect of cur-cumin on renal monocyte chemotactic protein 1 (MCP-1) expression and investigated the underlying mechanism of this effect
Materials and methods
Reagents and chemicals
Fetal bovine serum 1640 cell culture medium trypsin and Trizol RNA extraction reagent were purchased from Gibco Life Technologies (Grand Island NY USA) Reverse transcription-polymerase chain reaction (RT-PCR)-related reagents such as Moloney murine leukemia virus (M-MLV) reverse transcriptase deoxyribo-nucleotide triphosphate (dNTP) random prim-
Histone deacetylation effect of curcumin in Kidney disease
12697 Int J Clin Exp Med 20169(7)12696-12704
ers and RNase inhibitor were obtained from Promega (Madison Wisconsin USA) Curcumin and valproic acid were from Sigma-Aldrich (St Louis MO USA) Adriamycin was purchased from Sangon Biotech (Shanghai China) The SYBR Green Mix Kit was from TOYOBO (Osaka Japan) PCR primers of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) EP300 and MCP-1 were synthesized by Invitrogen (Shanghai China) The chromatin immunopre-cipitation (ChIP) and histone acetylation assay kits were purchased from Epigentek (NY USA) Fluorescein phalloidin for labelling F-actin was from Invitrogen (Carlsbad CA USA)
Animals
Specific pathogen free Sprague Dawley rats (180 mg) were purchased from the Laboratory Animal Center of Ruijin Hospital Shanghai Jiao Tong University School of Medicine (Shanghai China) The animals were maintained under climate-controlled conditions with a 12-h lightdark cycle and were fed standard rodent chow and water All experiments were con- ducted in accordance with in accordance with the National Institutes of Health Guide for the Care and with the approval of the In- stitutional Animal Care
Animal experiments
Rats were divided into three groups control group (animals that received an injection of saline by penis vein) ADR-induced kidney dis-ease model group (animals that were given intravenous ADR 5 mgmiddotkg-1 and one week later 25 mgmiddotkg-1) the intervention group (animals that were modeled as model group and 2 weeks later received curcumin 200 mgmiddotkg-1middotd-1)
Rats were sacrificed 8 weeks after the second time of ADR injection Kidney tissues were fixed in 10 formalin to prepare paraffin-embedded tissue sections for immunohistochemistry or used for MCP-1 mRNA detection and ChIP-qPCR Serum and urine were collected for detecting Albumin-Creatinine Ratio (ACR) and serum creatinine (Scr)
Detection of mRNA expression in renal tissue by real-time PCR
mRNA expression in kidneys was measured via quantitative reverse transcriptase PCR Total mRNA of the harvested kidneys was isolated using TRIzol and reverse-transcribed into cDNA as previously described SYBR Green Real-Time
PCR was performed in a 15-mL PCR mixture volume consisting of 75 mL of SYBR Green Real-Time PCR master mix (TOYOBO Osaka Japan) containing Hotmaster Taq DNA poly-merase and SYBR solution 03 mL (10 mM) of each forward and reverse primer 1 mL of cDNA and 59 mL of nuclease-free water PCR ampli-fication reactions were performed in a Chromo4 Four-color Real-Time PCR Detection System (Bio-Rad CA USA) with the following thermal cycle conditions initial denaturation 95degC for 3 min 40 cycles of amplification at 95degC for 15 s and annealing at 60degC for 45 s Each sample was analyzed in triplicate Data were normal-ized to GAPDH and calculated as the fold change in value of the treatment groups rela-tive to the control groups in accordance with the 2-ΔΔCT method
Primers were designed with Premier 50 soft-ware with the following sequences For tissue GAPDH forward 5rsquo-AGTATGACTCCACTCACGG- CAA-3rsquo and reverse 5rsquo-TCTCGCTCCTGGAAGAT- GGT-3rsquo MCP-1 forward 5rsquo-TAGCATCCACGTGCT- GTCTC-3rsquo and reverse 5rsquo-TGAGGTGGTTGTGG- AAAAGA-3rsquo EP300 forward 5rsquo-GGGACTAACCA- ATGGTGGTG-3rsquo and reverse 5rsquo-ATTGGGAGAA- GTCAAGCCTG-3rsquo For podocytes GAPDH for-ward 5rsquo-TCAAGCTCATTTCCTGGTAT-3rsquo and rever- se 5rsquo-GTCCAGGGTTTCTTACTCCT-3rsquo MCP-1 for-ward 5rsquo-TAGCATCCACGTGCTGTCTC-3rsquo and rever- se 5rsquo-TGAGGTGGTTGTGGAAAAGA-3rsquo EP300 forward 5rsquo-TTCAGCCAAGCGGCCTAAAC-3rsquo and reverse 5rsquo-GTTCCAGGTCAAACAGTGAACCAA-3rsquo
Histone acetylation assay
Kidney tissue (40 mg per sample) was applied and histone was withdrawn in accordance with the protocol in the Epigentek kit The histone acetylation level was detected at 450 nm wave-length with a microplate reader and calculated as acetylation = OD (sample-blank wells)OD (control group-blank wells) times 100 where OD is the optical density
ChIP
ChIP assays were performed in accordance with the protocol from Epigentek Real-time-qPCR was performed as in the procedure explained above Primers were designed with Premier 50 software with the following se- quences For tissue MCP-1 forward 5rsquo-GCA- CTTACTCAGCAGATTC-3rsquo and reverse 5rsquo-GCCT- CAGCCTTTTATTGT-3rsquo For podocytes MCP-1
Histone deacetylation effect of curcumin in Kidney disease
12698 Int J Clin Exp Med 20169(7)12696-12704
forward 5rsquo-ATTTCCACGCTCTTATCCTACTCTG-3rsquo and reverse 5rsquo-TCACCATTGCAAAGTGAATTGG-3rsquo
In vitro experiments
Cell culture Immortalized mouse podocytes were cultured in accordance with the standards outlined by Professor Mundel of Mount Sinai School of Medicine USA [10] Briefly the immortalized podocytes were cultured for 5 d at 33degC in 5 CO2 under permissive conditions and were subsequently cultured for 2 wk at 37degC in 5 CO2 By day 14 the podocytes had converted into spindle-shaped cells with small branches
Immunofluorescence assay Cells were washed once with PBS and fixed in 4 paraformalde-
hyde in PBS for 20 min at room temperature After the cells had been washed three times with PBS the nonspecific binding sites were blocked for 20 min with 3 bovine serum albu-min at room temperature before being incubat-ed with fluorescein-labelled phalloidin for F-actin detection for 20 min at room tempera-ture Subsequently nuclei were stained with 4rsquo6rsquo-diamidino-2-phenylindole dihydrochloride (DAPI) for 5 min at room temperature Images were obtained with an LSM 510 laser scanning confocal microscope (Carl Zeiss Thornwood NY USA) To visualize F-actin Fluorescence microscope (Olympus) was used The content of F-actin was determined by measuring the number of actin stress fibres Ten fields per group were used for the measurement The
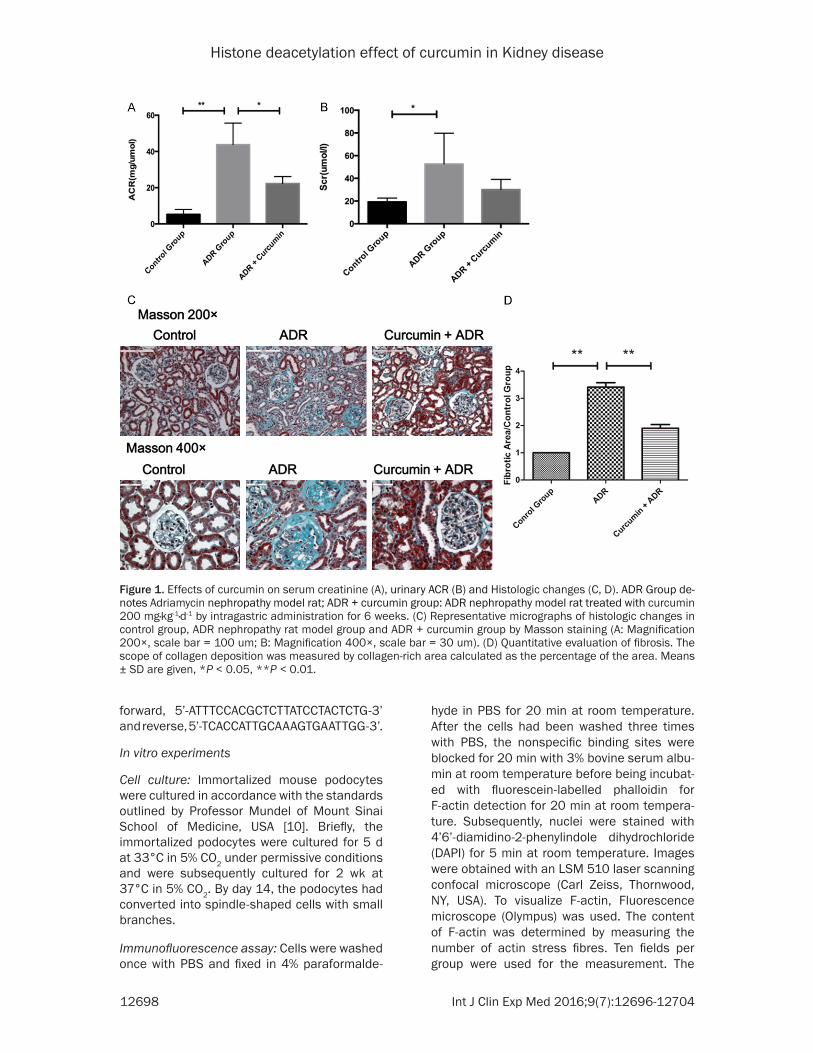
Figure 1 Effects of curcumin on serum creatinine (A) urinary ACR (B) and Histologic changes (C D) ADR Group de-notes Adriamycin nephropathy model rat ADR + curcumin group ADR nephropathy model rat treated with curcumin 200 mgmiddotkg-1middotd-1 by intragastric administration for 6 weeks (C) Representative micrographs of histologic changes in control group ADR nephropathy rat model group and ADR + curcumin group by Masson staining (A Magnification 200times scale bar = 100 um B Magnification 400times scale bar = 30 um) (D) Quantitative evaluation of fibrosis The scope of collagen deposition was measured by collagen-rich area calculated as the percentage of the area Means plusmn SD are given P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12699 Int J Clin Exp Med 20169(7)12696-12704
average content of F-actin in three independent experiments was determined
Statistical analyses
Results are expressed as mean plusmn standard deviation (SD) Significant differences in the mean values were evaluated using Studentrsquos t-test Intergroup variation was measured by one way analysis of variance (ANOVA) Statis- tical significance was considered P lt 005 Statistical analyses were performed with GraphPad Prism 5
Results
Curcumin treatment ameliorated biochemical parameters and renal pathology induced by ADR
In ADR Group urinary ACR were significant- ly higher than Control Group (4379plusmn6875 vs 5263plusmn1116 mgmiddotμmol-1 P lt 001) Urinary ACR in the Curucmin-treated group significantly decreased than that of the model group (2227plusmn2214 vs 4379plusmn6875 mgmiddotμmol-1 P lt 005) (Figure 1A) Compared with control group Serum creatinine increased (5266plusmn1568 vs 1925plusmn1367 μmolmiddotl-1 P lt 005) Curcumin treatment did not change Serum creatinine compared with model group (3012plusmn4498 vs 5266plusmn1568 μmolmiddotl-1 P = 0172) (Figure 1B)
Renal pathology was evaluated after ADR injec-tion Extracellular matrix (ECM) deposition
could be observed in the ADR-induced kidney disease model (both at 200times and 400times magni-fication) showing more fibrosis in the intersti-tial area and more sclerosis in the glomerular (Figure 1C) Quantitative data showed that the area of fibrosis was 343-fold that of the control group (P lt 001 Figure 1D) The above distribu-tion pattern of fibrosis in the ADR-induced kid-ney disease model was in accord with the liter-ature indicating the establishment of a renal fibrosis model
Treatment with curcumin significantly reduced renal ECM accumulation indicated by the smaller area of green color in tissue sections (Figure 1C) Quantitative data (Figure 1D) also showed that treatment with curcumin was associated with 19-fold less fibrosis (P lt 001)
Curcumin increased of EP300 mRNA tran-scription and decrease histone acetylation level
Renal EP300 mRNA levels were 3782-fold higher in the group treated with curcumin com-pared with the control group (P lt 001) while treatment with curcumin decreased renal EP300 mRNA expression to 1737-fold of the control (P lt 001 Figure 2A)
To further investigate the histone level of kid-ney cortex tissues treated with curcumin we measured the histone levels Both H3 (303-fold that of the control P lt 001) and H4 (16-fold that of the control P lt 005) acetylation
Figure 2 Levels of EP300 (A) and MCP-1 (B) mRNA in kidney of ADR nephropathy rats The level of mRNA was de-tectd by realtime-PCR and assessed by CT value with 2-ΔΔCT method Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12700 Int J Clin Exp Med 20169(7)12696-12704
levels were increased in the fibrosis model groups (Figure 3A 3B) Curcumin treatment could decrease both H3 and H4 acetylation lev-els to 21-fold (P lt 001) and 098-fold (P lt 001) of the control (Figure 3A 3B)
Curcumin decreased MCP-1 transcription and histone acetylation of the MCP-1 promoter in ADR-induced kidney injury model
Renal MCP-1 mRNA in the model group was sig-nificantly higher (8686-fold) than that in the
control (P lt 005) and curcumin treatment could decrease ADR-induced MCP-1 expres-sion to 4-fold (P lt 005 Figure 2B) These results indicated that curcumin could decrease the inflammation of renal disease to some extent
To understand the relationship between the changing histone acetylation function of cur-cumin and MCP-1 expression we applied the ChIP-qPCR assay by anti-Acetyl-H3 and anti-Acetyl-H4 antibody to detect the histone acety-
Figure 3 Effects of curcumin on ADR-induced renal histone acetylation levels A B Kidney cortex tissue (40 mg per sample) was applied and the histone acetylation levels were detected at 450 nm wavelengths with a microplate reader (details in methods) C D Rat cortex kidney tissues by different treatment were applied ChIP-qPCR assays were performed with anti-Acetyl-H3 anti-Acetyl-H4 and non-specific IgG antibodies and the binding was assessed on MCP-1 promoter region Values were presented as fold changes Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12701 Int J Clin Exp Med 20169(7)12696-12704
lation of the MCP-1 promoter H3 and H4 ace- tylation levels of the MCP-1 promoter were 36- and 45-fold increased that of the control respectively (P lt 001 and P lt 005 Figure 3C 3D) in ADR-nephropathy Curcumin treatment decreased ADR-induced histone acetylation (177- and 16-fold that of the control P lt 005 and P lt 001 Figure 3C 3D)
Curcumin relieved podocyte injury induced by ADR
Stimulation of mouse podocytes with ADR could induce cell death significantly (Figure 4A) Pretreatment with curcumin ameliorated ADR-induced podocyte injury partly To test whether curcumin has a role in the organization of the actin cytoskeleton we used fluorescent phalloidin-labelled F-actin to observe actin rearrangement After ADR stimulation for 24 h the filament was dissolved podocyte structure was disorganized and compared to the ADR
group curcumin intervention could preserve some structure (Figure 4B) All of the above indicated that curcumin could ameliorate podo-cyte injury induced by ADR to a certain degree
Curcumin changed EP300 and MCP-1 mRNA transcription and decreased MCP-1 promoter histone acetylation in podocyte
EP300 mRNA was 34-fold that of the control (P lt 005) by ADR-treated for 24 hours and com-pared to ADR group (Figure 5A) Compared with the ADR group curcumin could significantly reduce EP300 mRNA to 142-fold that of the control (P lt 005)
Compared with the ADR group curcumin (P lt 001) treatment reduced the level of MCP-1 mRNA expression to 072-fold of control respec-tively (Figure 5B) Also the MCP-1 promoter was hyperacetylated to 1997-fold that of the control in the ADR group detected by ChIP-
Figure 4 Curcumin protect ADR-caused podocyte injury A Podocytes were pretreated with curcumin (05 and 1 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h Podocytes were observed by inverted micro-scope B Podocytes were pretreated with curcumin (05 and 025 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The nulei were stained in blue and F-actin was stained in green (Original magnification times400 scale bar = 50 um) Data represent the mean plusmn SD P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780

Histone deacetylation effect of curcumin in Kidney disease
12697 Int J Clin Exp Med 20169(7)12696-12704
ers and RNase inhibitor were obtained from Promega (Madison Wisconsin USA) Curcumin and valproic acid were from Sigma-Aldrich (St Louis MO USA) Adriamycin was purchased from Sangon Biotech (Shanghai China) The SYBR Green Mix Kit was from TOYOBO (Osaka Japan) PCR primers of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) EP300 and MCP-1 were synthesized by Invitrogen (Shanghai China) The chromatin immunopre-cipitation (ChIP) and histone acetylation assay kits were purchased from Epigentek (NY USA) Fluorescein phalloidin for labelling F-actin was from Invitrogen (Carlsbad CA USA)
Animals
Specific pathogen free Sprague Dawley rats (180 mg) were purchased from the Laboratory Animal Center of Ruijin Hospital Shanghai Jiao Tong University School of Medicine (Shanghai China) The animals were maintained under climate-controlled conditions with a 12-h lightdark cycle and were fed standard rodent chow and water All experiments were con- ducted in accordance with in accordance with the National Institutes of Health Guide for the Care and with the approval of the In- stitutional Animal Care
Animal experiments
Rats were divided into three groups control group (animals that received an injection of saline by penis vein) ADR-induced kidney dis-ease model group (animals that were given intravenous ADR 5 mgmiddotkg-1 and one week later 25 mgmiddotkg-1) the intervention group (animals that were modeled as model group and 2 weeks later received curcumin 200 mgmiddotkg-1middotd-1)
Rats were sacrificed 8 weeks after the second time of ADR injection Kidney tissues were fixed in 10 formalin to prepare paraffin-embedded tissue sections for immunohistochemistry or used for MCP-1 mRNA detection and ChIP-qPCR Serum and urine were collected for detecting Albumin-Creatinine Ratio (ACR) and serum creatinine (Scr)
Detection of mRNA expression in renal tissue by real-time PCR
mRNA expression in kidneys was measured via quantitative reverse transcriptase PCR Total mRNA of the harvested kidneys was isolated using TRIzol and reverse-transcribed into cDNA as previously described SYBR Green Real-Time
PCR was performed in a 15-mL PCR mixture volume consisting of 75 mL of SYBR Green Real-Time PCR master mix (TOYOBO Osaka Japan) containing Hotmaster Taq DNA poly-merase and SYBR solution 03 mL (10 mM) of each forward and reverse primer 1 mL of cDNA and 59 mL of nuclease-free water PCR ampli-fication reactions were performed in a Chromo4 Four-color Real-Time PCR Detection System (Bio-Rad CA USA) with the following thermal cycle conditions initial denaturation 95degC for 3 min 40 cycles of amplification at 95degC for 15 s and annealing at 60degC for 45 s Each sample was analyzed in triplicate Data were normal-ized to GAPDH and calculated as the fold change in value of the treatment groups rela-tive to the control groups in accordance with the 2-ΔΔCT method
Primers were designed with Premier 50 soft-ware with the following sequences For tissue GAPDH forward 5rsquo-AGTATGACTCCACTCACGG- CAA-3rsquo and reverse 5rsquo-TCTCGCTCCTGGAAGAT- GGT-3rsquo MCP-1 forward 5rsquo-TAGCATCCACGTGCT- GTCTC-3rsquo and reverse 5rsquo-TGAGGTGGTTGTGG- AAAAGA-3rsquo EP300 forward 5rsquo-GGGACTAACCA- ATGGTGGTG-3rsquo and reverse 5rsquo-ATTGGGAGAA- GTCAAGCCTG-3rsquo For podocytes GAPDH for-ward 5rsquo-TCAAGCTCATTTCCTGGTAT-3rsquo and rever- se 5rsquo-GTCCAGGGTTTCTTACTCCT-3rsquo MCP-1 for-ward 5rsquo-TAGCATCCACGTGCTGTCTC-3rsquo and rever- se 5rsquo-TGAGGTGGTTGTGGAAAAGA-3rsquo EP300 forward 5rsquo-TTCAGCCAAGCGGCCTAAAC-3rsquo and reverse 5rsquo-GTTCCAGGTCAAACAGTGAACCAA-3rsquo
Histone acetylation assay
Kidney tissue (40 mg per sample) was applied and histone was withdrawn in accordance with the protocol in the Epigentek kit The histone acetylation level was detected at 450 nm wave-length with a microplate reader and calculated as acetylation = OD (sample-blank wells)OD (control group-blank wells) times 100 where OD is the optical density
ChIP
ChIP assays were performed in accordance with the protocol from Epigentek Real-time-qPCR was performed as in the procedure explained above Primers were designed with Premier 50 software with the following se- quences For tissue MCP-1 forward 5rsquo-GCA- CTTACTCAGCAGATTC-3rsquo and reverse 5rsquo-GCCT- CAGCCTTTTATTGT-3rsquo For podocytes MCP-1
Histone deacetylation effect of curcumin in Kidney disease
12698 Int J Clin Exp Med 20169(7)12696-12704
forward 5rsquo-ATTTCCACGCTCTTATCCTACTCTG-3rsquo and reverse 5rsquo-TCACCATTGCAAAGTGAATTGG-3rsquo
In vitro experiments
Cell culture Immortalized mouse podocytes were cultured in accordance with the standards outlined by Professor Mundel of Mount Sinai School of Medicine USA [10] Briefly the immortalized podocytes were cultured for 5 d at 33degC in 5 CO2 under permissive conditions and were subsequently cultured for 2 wk at 37degC in 5 CO2 By day 14 the podocytes had converted into spindle-shaped cells with small branches
Immunofluorescence assay Cells were washed once with PBS and fixed in 4 paraformalde-
hyde in PBS for 20 min at room temperature After the cells had been washed three times with PBS the nonspecific binding sites were blocked for 20 min with 3 bovine serum albu-min at room temperature before being incubat-ed with fluorescein-labelled phalloidin for F-actin detection for 20 min at room tempera-ture Subsequently nuclei were stained with 4rsquo6rsquo-diamidino-2-phenylindole dihydrochloride (DAPI) for 5 min at room temperature Images were obtained with an LSM 510 laser scanning confocal microscope (Carl Zeiss Thornwood NY USA) To visualize F-actin Fluorescence microscope (Olympus) was used The content of F-actin was determined by measuring the number of actin stress fibres Ten fields per group were used for the measurement The
Figure 1 Effects of curcumin on serum creatinine (A) urinary ACR (B) and Histologic changes (C D) ADR Group de-notes Adriamycin nephropathy model rat ADR + curcumin group ADR nephropathy model rat treated with curcumin 200 mgmiddotkg-1middotd-1 by intragastric administration for 6 weeks (C) Representative micrographs of histologic changes in control group ADR nephropathy rat model group and ADR + curcumin group by Masson staining (A Magnification 200times scale bar = 100 um B Magnification 400times scale bar = 30 um) (D) Quantitative evaluation of fibrosis The scope of collagen deposition was measured by collagen-rich area calculated as the percentage of the area Means plusmn SD are given P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12699 Int J Clin Exp Med 20169(7)12696-12704
average content of F-actin in three independent experiments was determined
Statistical analyses
Results are expressed as mean plusmn standard deviation (SD) Significant differences in the mean values were evaluated using Studentrsquos t-test Intergroup variation was measured by one way analysis of variance (ANOVA) Statis- tical significance was considered P lt 005 Statistical analyses were performed with GraphPad Prism 5
Results
Curcumin treatment ameliorated biochemical parameters and renal pathology induced by ADR
In ADR Group urinary ACR were significant- ly higher than Control Group (4379plusmn6875 vs 5263plusmn1116 mgmiddotμmol-1 P lt 001) Urinary ACR in the Curucmin-treated group significantly decreased than that of the model group (2227plusmn2214 vs 4379plusmn6875 mgmiddotμmol-1 P lt 005) (Figure 1A) Compared with control group Serum creatinine increased (5266plusmn1568 vs 1925plusmn1367 μmolmiddotl-1 P lt 005) Curcumin treatment did not change Serum creatinine compared with model group (3012plusmn4498 vs 5266plusmn1568 μmolmiddotl-1 P = 0172) (Figure 1B)
Renal pathology was evaluated after ADR injec-tion Extracellular matrix (ECM) deposition
could be observed in the ADR-induced kidney disease model (both at 200times and 400times magni-fication) showing more fibrosis in the intersti-tial area and more sclerosis in the glomerular (Figure 1C) Quantitative data showed that the area of fibrosis was 343-fold that of the control group (P lt 001 Figure 1D) The above distribu-tion pattern of fibrosis in the ADR-induced kid-ney disease model was in accord with the liter-ature indicating the establishment of a renal fibrosis model
Treatment with curcumin significantly reduced renal ECM accumulation indicated by the smaller area of green color in tissue sections (Figure 1C) Quantitative data (Figure 1D) also showed that treatment with curcumin was associated with 19-fold less fibrosis (P lt 001)
Curcumin increased of EP300 mRNA tran-scription and decrease histone acetylation level
Renal EP300 mRNA levels were 3782-fold higher in the group treated with curcumin com-pared with the control group (P lt 001) while treatment with curcumin decreased renal EP300 mRNA expression to 1737-fold of the control (P lt 001 Figure 2A)
To further investigate the histone level of kid-ney cortex tissues treated with curcumin we measured the histone levels Both H3 (303-fold that of the control P lt 001) and H4 (16-fold that of the control P lt 005) acetylation
Figure 2 Levels of EP300 (A) and MCP-1 (B) mRNA in kidney of ADR nephropathy rats The level of mRNA was de-tectd by realtime-PCR and assessed by CT value with 2-ΔΔCT method Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12700 Int J Clin Exp Med 20169(7)12696-12704
levels were increased in the fibrosis model groups (Figure 3A 3B) Curcumin treatment could decrease both H3 and H4 acetylation lev-els to 21-fold (P lt 001) and 098-fold (P lt 001) of the control (Figure 3A 3B)
Curcumin decreased MCP-1 transcription and histone acetylation of the MCP-1 promoter in ADR-induced kidney injury model
Renal MCP-1 mRNA in the model group was sig-nificantly higher (8686-fold) than that in the
control (P lt 005) and curcumin treatment could decrease ADR-induced MCP-1 expres-sion to 4-fold (P lt 005 Figure 2B) These results indicated that curcumin could decrease the inflammation of renal disease to some extent
To understand the relationship between the changing histone acetylation function of cur-cumin and MCP-1 expression we applied the ChIP-qPCR assay by anti-Acetyl-H3 and anti-Acetyl-H4 antibody to detect the histone acety-
Figure 3 Effects of curcumin on ADR-induced renal histone acetylation levels A B Kidney cortex tissue (40 mg per sample) was applied and the histone acetylation levels were detected at 450 nm wavelengths with a microplate reader (details in methods) C D Rat cortex kidney tissues by different treatment were applied ChIP-qPCR assays were performed with anti-Acetyl-H3 anti-Acetyl-H4 and non-specific IgG antibodies and the binding was assessed on MCP-1 promoter region Values were presented as fold changes Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12701 Int J Clin Exp Med 20169(7)12696-12704
lation of the MCP-1 promoter H3 and H4 ace- tylation levels of the MCP-1 promoter were 36- and 45-fold increased that of the control respectively (P lt 001 and P lt 005 Figure 3C 3D) in ADR-nephropathy Curcumin treatment decreased ADR-induced histone acetylation (177- and 16-fold that of the control P lt 005 and P lt 001 Figure 3C 3D)
Curcumin relieved podocyte injury induced by ADR
Stimulation of mouse podocytes with ADR could induce cell death significantly (Figure 4A) Pretreatment with curcumin ameliorated ADR-induced podocyte injury partly To test whether curcumin has a role in the organization of the actin cytoskeleton we used fluorescent phalloidin-labelled F-actin to observe actin rearrangement After ADR stimulation for 24 h the filament was dissolved podocyte structure was disorganized and compared to the ADR
group curcumin intervention could preserve some structure (Figure 4B) All of the above indicated that curcumin could ameliorate podo-cyte injury induced by ADR to a certain degree
Curcumin changed EP300 and MCP-1 mRNA transcription and decreased MCP-1 promoter histone acetylation in podocyte
EP300 mRNA was 34-fold that of the control (P lt 005) by ADR-treated for 24 hours and com-pared to ADR group (Figure 5A) Compared with the ADR group curcumin could significantly reduce EP300 mRNA to 142-fold that of the control (P lt 005)
Compared with the ADR group curcumin (P lt 001) treatment reduced the level of MCP-1 mRNA expression to 072-fold of control respec-tively (Figure 5B) Also the MCP-1 promoter was hyperacetylated to 1997-fold that of the control in the ADR group detected by ChIP-
Figure 4 Curcumin protect ADR-caused podocyte injury A Podocytes were pretreated with curcumin (05 and 1 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h Podocytes were observed by inverted micro-scope B Podocytes were pretreated with curcumin (05 and 025 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The nulei were stained in blue and F-actin was stained in green (Original magnification times400 scale bar = 50 um) Data represent the mean plusmn SD P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780
Histone deacetylation effect of curcumin in Kidney disease
12698 Int J Clin Exp Med 20169(7)12696-12704
forward 5rsquo-ATTTCCACGCTCTTATCCTACTCTG-3rsquo and reverse 5rsquo-TCACCATTGCAAAGTGAATTGG-3rsquo
In vitro experiments
Cell culture Immortalized mouse podocytes were cultured in accordance with the standards outlined by Professor Mundel of Mount Sinai School of Medicine USA [10] Briefly the immortalized podocytes were cultured for 5 d at 33degC in 5 CO2 under permissive conditions and were subsequently cultured for 2 wk at 37degC in 5 CO2 By day 14 the podocytes had converted into spindle-shaped cells with small branches
Immunofluorescence assay Cells were washed once with PBS and fixed in 4 paraformalde-
hyde in PBS for 20 min at room temperature After the cells had been washed three times with PBS the nonspecific binding sites were blocked for 20 min with 3 bovine serum albu-min at room temperature before being incubat-ed with fluorescein-labelled phalloidin for F-actin detection for 20 min at room tempera-ture Subsequently nuclei were stained with 4rsquo6rsquo-diamidino-2-phenylindole dihydrochloride (DAPI) for 5 min at room temperature Images were obtained with an LSM 510 laser scanning confocal microscope (Carl Zeiss Thornwood NY USA) To visualize F-actin Fluorescence microscope (Olympus) was used The content of F-actin was determined by measuring the number of actin stress fibres Ten fields per group were used for the measurement The
Figure 1 Effects of curcumin on serum creatinine (A) urinary ACR (B) and Histologic changes (C D) ADR Group de-notes Adriamycin nephropathy model rat ADR + curcumin group ADR nephropathy model rat treated with curcumin 200 mgmiddotkg-1middotd-1 by intragastric administration for 6 weeks (C) Representative micrographs of histologic changes in control group ADR nephropathy rat model group and ADR + curcumin group by Masson staining (A Magnification 200times scale bar = 100 um B Magnification 400times scale bar = 30 um) (D) Quantitative evaluation of fibrosis The scope of collagen deposition was measured by collagen-rich area calculated as the percentage of the area Means plusmn SD are given P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12699 Int J Clin Exp Med 20169(7)12696-12704
average content of F-actin in three independent experiments was determined
Statistical analyses
Results are expressed as mean plusmn standard deviation (SD) Significant differences in the mean values were evaluated using Studentrsquos t-test Intergroup variation was measured by one way analysis of variance (ANOVA) Statis- tical significance was considered P lt 005 Statistical analyses were performed with GraphPad Prism 5
Results
Curcumin treatment ameliorated biochemical parameters and renal pathology induced by ADR
In ADR Group urinary ACR were significant- ly higher than Control Group (4379plusmn6875 vs 5263plusmn1116 mgmiddotμmol-1 P lt 001) Urinary ACR in the Curucmin-treated group significantly decreased than that of the model group (2227plusmn2214 vs 4379plusmn6875 mgmiddotμmol-1 P lt 005) (Figure 1A) Compared with control group Serum creatinine increased (5266plusmn1568 vs 1925plusmn1367 μmolmiddotl-1 P lt 005) Curcumin treatment did not change Serum creatinine compared with model group (3012plusmn4498 vs 5266plusmn1568 μmolmiddotl-1 P = 0172) (Figure 1B)
Renal pathology was evaluated after ADR injec-tion Extracellular matrix (ECM) deposition
could be observed in the ADR-induced kidney disease model (both at 200times and 400times magni-fication) showing more fibrosis in the intersti-tial area and more sclerosis in the glomerular (Figure 1C) Quantitative data showed that the area of fibrosis was 343-fold that of the control group (P lt 001 Figure 1D) The above distribu-tion pattern of fibrosis in the ADR-induced kid-ney disease model was in accord with the liter-ature indicating the establishment of a renal fibrosis model
Treatment with curcumin significantly reduced renal ECM accumulation indicated by the smaller area of green color in tissue sections (Figure 1C) Quantitative data (Figure 1D) also showed that treatment with curcumin was associated with 19-fold less fibrosis (P lt 001)
Curcumin increased of EP300 mRNA tran-scription and decrease histone acetylation level
Renal EP300 mRNA levels were 3782-fold higher in the group treated with curcumin com-pared with the control group (P lt 001) while treatment with curcumin decreased renal EP300 mRNA expression to 1737-fold of the control (P lt 001 Figure 2A)
To further investigate the histone level of kid-ney cortex tissues treated with curcumin we measured the histone levels Both H3 (303-fold that of the control P lt 001) and H4 (16-fold that of the control P lt 005) acetylation
Figure 2 Levels of EP300 (A) and MCP-1 (B) mRNA in kidney of ADR nephropathy rats The level of mRNA was de-tectd by realtime-PCR and assessed by CT value with 2-ΔΔCT method Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12700 Int J Clin Exp Med 20169(7)12696-12704
levels were increased in the fibrosis model groups (Figure 3A 3B) Curcumin treatment could decrease both H3 and H4 acetylation lev-els to 21-fold (P lt 001) and 098-fold (P lt 001) of the control (Figure 3A 3B)
Curcumin decreased MCP-1 transcription and histone acetylation of the MCP-1 promoter in ADR-induced kidney injury model
Renal MCP-1 mRNA in the model group was sig-nificantly higher (8686-fold) than that in the
control (P lt 005) and curcumin treatment could decrease ADR-induced MCP-1 expres-sion to 4-fold (P lt 005 Figure 2B) These results indicated that curcumin could decrease the inflammation of renal disease to some extent
To understand the relationship between the changing histone acetylation function of cur-cumin and MCP-1 expression we applied the ChIP-qPCR assay by anti-Acetyl-H3 and anti-Acetyl-H4 antibody to detect the histone acety-
Figure 3 Effects of curcumin on ADR-induced renal histone acetylation levels A B Kidney cortex tissue (40 mg per sample) was applied and the histone acetylation levels were detected at 450 nm wavelengths with a microplate reader (details in methods) C D Rat cortex kidney tissues by different treatment were applied ChIP-qPCR assays were performed with anti-Acetyl-H3 anti-Acetyl-H4 and non-specific IgG antibodies and the binding was assessed on MCP-1 promoter region Values were presented as fold changes Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12701 Int J Clin Exp Med 20169(7)12696-12704
lation of the MCP-1 promoter H3 and H4 ace- tylation levels of the MCP-1 promoter were 36- and 45-fold increased that of the control respectively (P lt 001 and P lt 005 Figure 3C 3D) in ADR-nephropathy Curcumin treatment decreased ADR-induced histone acetylation (177- and 16-fold that of the control P lt 005 and P lt 001 Figure 3C 3D)
Curcumin relieved podocyte injury induced by ADR
Stimulation of mouse podocytes with ADR could induce cell death significantly (Figure 4A) Pretreatment with curcumin ameliorated ADR-induced podocyte injury partly To test whether curcumin has a role in the organization of the actin cytoskeleton we used fluorescent phalloidin-labelled F-actin to observe actin rearrangement After ADR stimulation for 24 h the filament was dissolved podocyte structure was disorganized and compared to the ADR
group curcumin intervention could preserve some structure (Figure 4B) All of the above indicated that curcumin could ameliorate podo-cyte injury induced by ADR to a certain degree
Curcumin changed EP300 and MCP-1 mRNA transcription and decreased MCP-1 promoter histone acetylation in podocyte
EP300 mRNA was 34-fold that of the control (P lt 005) by ADR-treated for 24 hours and com-pared to ADR group (Figure 5A) Compared with the ADR group curcumin could significantly reduce EP300 mRNA to 142-fold that of the control (P lt 005)
Compared with the ADR group curcumin (P lt 001) treatment reduced the level of MCP-1 mRNA expression to 072-fold of control respec-tively (Figure 5B) Also the MCP-1 promoter was hyperacetylated to 1997-fold that of the control in the ADR group detected by ChIP-
Figure 4 Curcumin protect ADR-caused podocyte injury A Podocytes were pretreated with curcumin (05 and 1 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h Podocytes were observed by inverted micro-scope B Podocytes were pretreated with curcumin (05 and 025 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The nulei were stained in blue and F-actin was stained in green (Original magnification times400 scale bar = 50 um) Data represent the mean plusmn SD P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780

Histone deacetylation effect of curcumin in Kidney disease
12699 Int J Clin Exp Med 20169(7)12696-12704
average content of F-actin in three independent experiments was determined
Statistical analyses
Results are expressed as mean plusmn standard deviation (SD) Significant differences in the mean values were evaluated using Studentrsquos t-test Intergroup variation was measured by one way analysis of variance (ANOVA) Statis- tical significance was considered P lt 005 Statistical analyses were performed with GraphPad Prism 5
Results
Curcumin treatment ameliorated biochemical parameters and renal pathology induced by ADR
In ADR Group urinary ACR were significant- ly higher than Control Group (4379plusmn6875 vs 5263plusmn1116 mgmiddotμmol-1 P lt 001) Urinary ACR in the Curucmin-treated group significantly decreased than that of the model group (2227plusmn2214 vs 4379plusmn6875 mgmiddotμmol-1 P lt 005) (Figure 1A) Compared with control group Serum creatinine increased (5266plusmn1568 vs 1925plusmn1367 μmolmiddotl-1 P lt 005) Curcumin treatment did not change Serum creatinine compared with model group (3012plusmn4498 vs 5266plusmn1568 μmolmiddotl-1 P = 0172) (Figure 1B)
Renal pathology was evaluated after ADR injec-tion Extracellular matrix (ECM) deposition
could be observed in the ADR-induced kidney disease model (both at 200times and 400times magni-fication) showing more fibrosis in the intersti-tial area and more sclerosis in the glomerular (Figure 1C) Quantitative data showed that the area of fibrosis was 343-fold that of the control group (P lt 001 Figure 1D) The above distribu-tion pattern of fibrosis in the ADR-induced kid-ney disease model was in accord with the liter-ature indicating the establishment of a renal fibrosis model
Treatment with curcumin significantly reduced renal ECM accumulation indicated by the smaller area of green color in tissue sections (Figure 1C) Quantitative data (Figure 1D) also showed that treatment with curcumin was associated with 19-fold less fibrosis (P lt 001)
Curcumin increased of EP300 mRNA tran-scription and decrease histone acetylation level
Renal EP300 mRNA levels were 3782-fold higher in the group treated with curcumin com-pared with the control group (P lt 001) while treatment with curcumin decreased renal EP300 mRNA expression to 1737-fold of the control (P lt 001 Figure 2A)
To further investigate the histone level of kid-ney cortex tissues treated with curcumin we measured the histone levels Both H3 (303-fold that of the control P lt 001) and H4 (16-fold that of the control P lt 005) acetylation
Figure 2 Levels of EP300 (A) and MCP-1 (B) mRNA in kidney of ADR nephropathy rats The level of mRNA was de-tectd by realtime-PCR and assessed by CT value with 2-ΔΔCT method Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12700 Int J Clin Exp Med 20169(7)12696-12704
levels were increased in the fibrosis model groups (Figure 3A 3B) Curcumin treatment could decrease both H3 and H4 acetylation lev-els to 21-fold (P lt 001) and 098-fold (P lt 001) of the control (Figure 3A 3B)
Curcumin decreased MCP-1 transcription and histone acetylation of the MCP-1 promoter in ADR-induced kidney injury model
Renal MCP-1 mRNA in the model group was sig-nificantly higher (8686-fold) than that in the
control (P lt 005) and curcumin treatment could decrease ADR-induced MCP-1 expres-sion to 4-fold (P lt 005 Figure 2B) These results indicated that curcumin could decrease the inflammation of renal disease to some extent
To understand the relationship between the changing histone acetylation function of cur-cumin and MCP-1 expression we applied the ChIP-qPCR assay by anti-Acetyl-H3 and anti-Acetyl-H4 antibody to detect the histone acety-
Figure 3 Effects of curcumin on ADR-induced renal histone acetylation levels A B Kidney cortex tissue (40 mg per sample) was applied and the histone acetylation levels were detected at 450 nm wavelengths with a microplate reader (details in methods) C D Rat cortex kidney tissues by different treatment were applied ChIP-qPCR assays were performed with anti-Acetyl-H3 anti-Acetyl-H4 and non-specific IgG antibodies and the binding was assessed on MCP-1 promoter region Values were presented as fold changes Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12701 Int J Clin Exp Med 20169(7)12696-12704
lation of the MCP-1 promoter H3 and H4 ace- tylation levels of the MCP-1 promoter were 36- and 45-fold increased that of the control respectively (P lt 001 and P lt 005 Figure 3C 3D) in ADR-nephropathy Curcumin treatment decreased ADR-induced histone acetylation (177- and 16-fold that of the control P lt 005 and P lt 001 Figure 3C 3D)
Curcumin relieved podocyte injury induced by ADR
Stimulation of mouse podocytes with ADR could induce cell death significantly (Figure 4A) Pretreatment with curcumin ameliorated ADR-induced podocyte injury partly To test whether curcumin has a role in the organization of the actin cytoskeleton we used fluorescent phalloidin-labelled F-actin to observe actin rearrangement After ADR stimulation for 24 h the filament was dissolved podocyte structure was disorganized and compared to the ADR
group curcumin intervention could preserve some structure (Figure 4B) All of the above indicated that curcumin could ameliorate podo-cyte injury induced by ADR to a certain degree
Curcumin changed EP300 and MCP-1 mRNA transcription and decreased MCP-1 promoter histone acetylation in podocyte
EP300 mRNA was 34-fold that of the control (P lt 005) by ADR-treated for 24 hours and com-pared to ADR group (Figure 5A) Compared with the ADR group curcumin could significantly reduce EP300 mRNA to 142-fold that of the control (P lt 005)
Compared with the ADR group curcumin (P lt 001) treatment reduced the level of MCP-1 mRNA expression to 072-fold of control respec-tively (Figure 5B) Also the MCP-1 promoter was hyperacetylated to 1997-fold that of the control in the ADR group detected by ChIP-
Figure 4 Curcumin protect ADR-caused podocyte injury A Podocytes were pretreated with curcumin (05 and 1 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h Podocytes were observed by inverted micro-scope B Podocytes were pretreated with curcumin (05 and 025 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The nulei were stained in blue and F-actin was stained in green (Original magnification times400 scale bar = 50 um) Data represent the mean plusmn SD P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780

Histone deacetylation effect of curcumin in Kidney disease
12700 Int J Clin Exp Med 20169(7)12696-12704
levels were increased in the fibrosis model groups (Figure 3A 3B) Curcumin treatment could decrease both H3 and H4 acetylation lev-els to 21-fold (P lt 001) and 098-fold (P lt 001) of the control (Figure 3A 3B)
Curcumin decreased MCP-1 transcription and histone acetylation of the MCP-1 promoter in ADR-induced kidney injury model
Renal MCP-1 mRNA in the model group was sig-nificantly higher (8686-fold) than that in the
control (P lt 005) and curcumin treatment could decrease ADR-induced MCP-1 expres-sion to 4-fold (P lt 005 Figure 2B) These results indicated that curcumin could decrease the inflammation of renal disease to some extent
To understand the relationship between the changing histone acetylation function of cur-cumin and MCP-1 expression we applied the ChIP-qPCR assay by anti-Acetyl-H3 and anti-Acetyl-H4 antibody to detect the histone acety-
Figure 3 Effects of curcumin on ADR-induced renal histone acetylation levels A B Kidney cortex tissue (40 mg per sample) was applied and the histone acetylation levels were detected at 450 nm wavelengths with a microplate reader (details in methods) C D Rat cortex kidney tissues by different treatment were applied ChIP-qPCR assays were performed with anti-Acetyl-H3 anti-Acetyl-H4 and non-specific IgG antibodies and the binding was assessed on MCP-1 promoter region Values were presented as fold changes Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12701 Int J Clin Exp Med 20169(7)12696-12704
lation of the MCP-1 promoter H3 and H4 ace- tylation levels of the MCP-1 promoter were 36- and 45-fold increased that of the control respectively (P lt 001 and P lt 005 Figure 3C 3D) in ADR-nephropathy Curcumin treatment decreased ADR-induced histone acetylation (177- and 16-fold that of the control P lt 005 and P lt 001 Figure 3C 3D)
Curcumin relieved podocyte injury induced by ADR
Stimulation of mouse podocytes with ADR could induce cell death significantly (Figure 4A) Pretreatment with curcumin ameliorated ADR-induced podocyte injury partly To test whether curcumin has a role in the organization of the actin cytoskeleton we used fluorescent phalloidin-labelled F-actin to observe actin rearrangement After ADR stimulation for 24 h the filament was dissolved podocyte structure was disorganized and compared to the ADR
group curcumin intervention could preserve some structure (Figure 4B) All of the above indicated that curcumin could ameliorate podo-cyte injury induced by ADR to a certain degree
Curcumin changed EP300 and MCP-1 mRNA transcription and decreased MCP-1 promoter histone acetylation in podocyte
EP300 mRNA was 34-fold that of the control (P lt 005) by ADR-treated for 24 hours and com-pared to ADR group (Figure 5A) Compared with the ADR group curcumin could significantly reduce EP300 mRNA to 142-fold that of the control (P lt 005)
Compared with the ADR group curcumin (P lt 001) treatment reduced the level of MCP-1 mRNA expression to 072-fold of control respec-tively (Figure 5B) Also the MCP-1 promoter was hyperacetylated to 1997-fold that of the control in the ADR group detected by ChIP-
Figure 4 Curcumin protect ADR-caused podocyte injury A Podocytes were pretreated with curcumin (05 and 1 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h Podocytes were observed by inverted micro-scope B Podocytes were pretreated with curcumin (05 and 025 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The nulei were stained in blue and F-actin was stained in green (Original magnification times400 scale bar = 50 um) Data represent the mean plusmn SD P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780

Histone deacetylation effect of curcumin in Kidney disease
12701 Int J Clin Exp Med 20169(7)12696-12704
lation of the MCP-1 promoter H3 and H4 ace- tylation levels of the MCP-1 promoter were 36- and 45-fold increased that of the control respectively (P lt 001 and P lt 005 Figure 3C 3D) in ADR-nephropathy Curcumin treatment decreased ADR-induced histone acetylation (177- and 16-fold that of the control P lt 005 and P lt 001 Figure 3C 3D)
Curcumin relieved podocyte injury induced by ADR
Stimulation of mouse podocytes with ADR could induce cell death significantly (Figure 4A) Pretreatment with curcumin ameliorated ADR-induced podocyte injury partly To test whether curcumin has a role in the organization of the actin cytoskeleton we used fluorescent phalloidin-labelled F-actin to observe actin rearrangement After ADR stimulation for 24 h the filament was dissolved podocyte structure was disorganized and compared to the ADR
group curcumin intervention could preserve some structure (Figure 4B) All of the above indicated that curcumin could ameliorate podo-cyte injury induced by ADR to a certain degree
Curcumin changed EP300 and MCP-1 mRNA transcription and decreased MCP-1 promoter histone acetylation in podocyte
EP300 mRNA was 34-fold that of the control (P lt 005) by ADR-treated for 24 hours and com-pared to ADR group (Figure 5A) Compared with the ADR group curcumin could significantly reduce EP300 mRNA to 142-fold that of the control (P lt 005)
Compared with the ADR group curcumin (P lt 001) treatment reduced the level of MCP-1 mRNA expression to 072-fold of control respec-tively (Figure 5B) Also the MCP-1 promoter was hyperacetylated to 1997-fold that of the control in the ADR group detected by ChIP-
Figure 4 Curcumin protect ADR-caused podocyte injury A Podocytes were pretreated with curcumin (05 and 1 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h Podocytes were observed by inverted micro-scope B Podocytes were pretreated with curcumin (05 and 025 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The nulei were stained in blue and F-actin was stained in green (Original magnification times400 scale bar = 50 um) Data represent the mean plusmn SD P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780
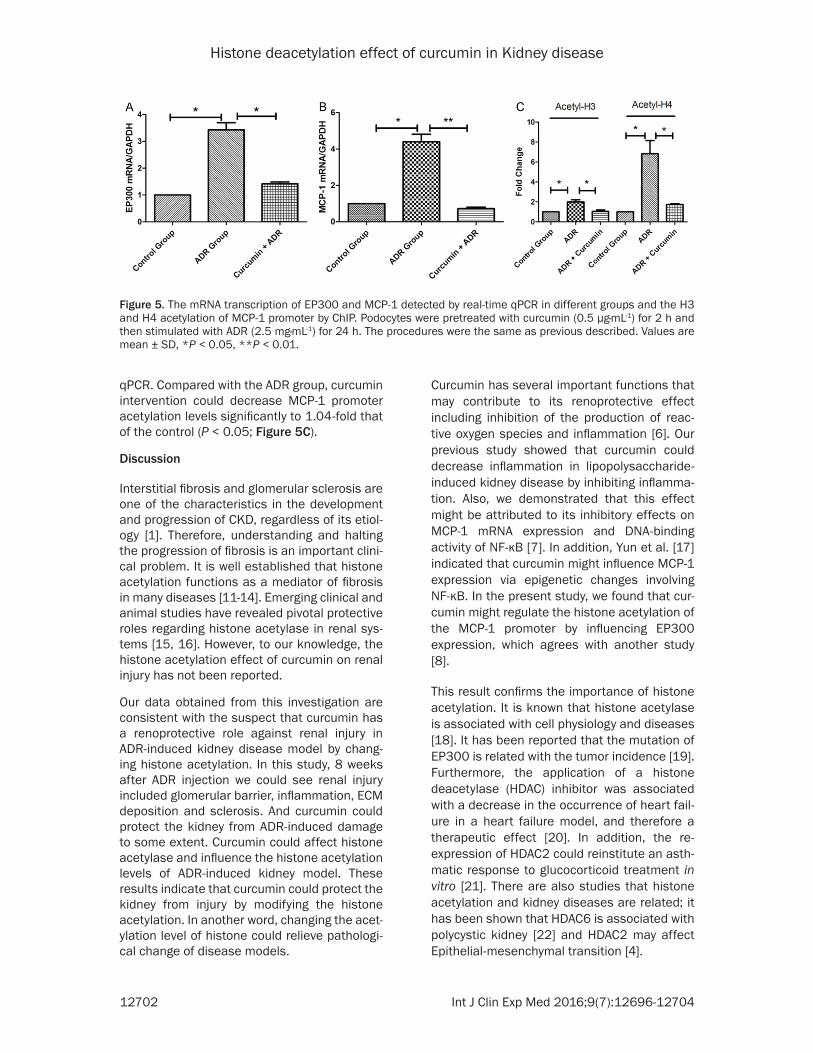
Histone deacetylation effect of curcumin in Kidney disease
12702 Int J Clin Exp Med 20169(7)12696-12704
qPCR Compared with the ADR group curcumin intervention could decrease MCP-1 promoter acetylation levels significantly to 104-fold that of the control (P lt 005 Figure 5C)
Discussion
Interstitial fibrosis and glomerular sclerosis are one of the characteristics in the development and progression of CKD regardless of its etiol-ogy [1] Therefore understanding and halting the progression of fibrosis is an important clini-cal problem It is well established that histone acetylation functions as a mediator of fibrosis in many diseases [11-14] Emerging clinical and animal studies have revealed pivotal protective roles regarding histone acetylase in renal sys-tems [15 16] However to our knowledge the histone acetylation effect of curcumin on renal injury has not been reported
Our data obtained from this investigation are consistent with the suspect that curcumin has a renoprotective role against renal injury in ADR-induced kidney disease model by chang-ing histone acetylation In this study 8 weeks after ADR injection we could see renal injury included glomerular barrier inflammation ECM deposition and sclerosis And curcumin could protect the kidney from ADR-induced damage to some extent Curcumin could affect histone acetylase and influence the histone acetylation levels of ADR-induced kidney model These results indicate that curcumin could protect the kidney from injury by modifying the histone acetylation In another word changing the acet-ylation level of histone could relieve pathologi-cal change of disease models
Curcumin has several important functions that may contribute to its renoprotective effect including inhibition of the production of reac-tive oxygen species and inflammation [6] Our previous study showed that curcumin could decrease inflammation in lipopolysaccharide-induced kidney disease by inhibiting inflamma-tion Also we demonstrated that this effect might be attributed to its inhibitory effects on MCP-1 mRNA expression and DNA-binding activity of NF-κB [7] In addition Yun et al [17] indicated that curcumin might influence MCP-1 expression via epigenetic changes involving NF-κB In the present study we found that cur-cumin might regulate the histone acetylation of the MCP-1 promoter by influencing EP300 expression which agrees with another study [8]
This result confirms the importance of histone acetylation It is known that histone acetylase is associated with cell physiology and diseases [18] It has been reported that the mutation of EP300 is related with the tumor incidence [19] Furthermore the application of a histone deacetylase (HDAC) inhibitor was associated with a decrease in the occurrence of heart fail-ure in a heart failure model and therefore a therapeutic effect [20] In addition the re-expression of HDAC2 could reinstitute an asth-matic response to glucocorticoid treatment in vitro [21] There are also studies that histone acetylation and kidney diseases are related it has been shown that HDAC6 is associated with polycystic kidney [22] and HDAC2 may affect Epithelial-mesenchymal transition [4]
Figure 5 The mRNA transcription of EP300 and MCP-1 detected by real-time qPCR in different groups and the H3 and H4 acetylation of MCP-1 promoter by ChIP Podocytes were pretreated with curcumin (05 microgmiddotmL-1) for 2 h and then stimulated with ADR (25 mgmiddotmL-1) for 24 h The procedures were the same as previous described Values are mean plusmn SD P lt 005 P lt 001
Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780

Histone deacetylation effect of curcumin in Kidney disease
12703 Int J Clin Exp Med 20169(7)12696-12704
ADR-induced kidney injury is a model of human FSGS characterized by glomerular sclerosis and tubulointerstitial fibrosis In our study we indicated that curcumin may protect podocytes in vitro by inhibiting MCP-1 expression and by restoring cytoskeleton which may be due to changing histone acetylation levels Studies has shown that podocyte injury was associated with proteinuria and the impaired podocyte may undergo glomerular base membrane dam-age cytoskeletal disorder changes of foot pro-cess motility podocyte apoptosis inflamma-tion and Epithelial-mesenchymal transition [23 24] MCP-1 as an important inflammatory cytokine can influence cell adhesion and in- duce inflammation and injury [25] Therefore MCP-1 could initiate and promote kidney dam-age and is associated with proteinuria [26 27] The most important cytoskeletal compo-nents of podocytes revealed recently includeα-actinin-4 podocin nephrin and integrin-link- ed kinase [28 29] Therefore we predict that the cytoskeleton rescued by curcumin may be associated with recovery of the above proteins
Although curcumin could protect podocytes from injury in the present study we found that curcumin in high concentrations could lead to podocyte death This is in accord with other lit-erature We presume that high doses of cur-cumin may cause cytoskeletal disorder and cell death However the mechanism of this oppo-site effect is unknown
Conclusion
In summary our study herein indicates that curcumin was able to attenuate ADR-induced kidney injury by changing histone acetylation levels and inhibiting MCP-1 expression Mech- anistic studies revealed that curcumin signifi-cantly changed histone acetylation of the MCP-1 promoter Result of our in vitro study suggests that low doses of curcumin could pro-tect podocytes from cytoskeletal disorder and MCP-1 expression caused by ADR and these effects may be linked to changing histone acet-ylation These findings provide a theoretical basis for further utilization of curcumin in renal disease
Acknowledgements
This work was supported by the Major State Basic Research Development Program of China
(973 Program 2012CB517700 and 2012CB- 517604) the National Natural Science Found- ation of China (81270782 30771000) and the Doctoral Innovation Fund Projects from Shanghai Jiao Tong University School of Medi- cine (BXJ201409)
Disclosure of conflict of interest
None
Address correspondence to Weiming Wang De- partment of Nephrology Ruijin Hospital Shanghai Jiao Tong University School of Medicine Shanghai 200025 PR China Tel 021-64370025665232 Fax 021-64370025665232 E-mail wweim-ing01126com
References
[1] Liu Y Cellular and molecular mechanisms of renal fibrosis Nat Rev Nephrol 2011 7 684-696
[2] Berger SL Histone modifications in transcrip-tional regulation Curr Opin Genet Dev 2002 12 142-148
[3] Graumlff J and Tsai LH Histone acetylation mo-lecular mnemonics on the chromatin Nat Rev Neurosci 2013 14 97-111
[4] Noh H Oh EY Seo JY Yu MR Kim YO Ha H and Lee HB Histone deacetylase-2 is a key regula-tor of diabetes-and transforming growth factor-β1-induced renal injury Am J Physiol Renal Physiol 2009 297 F729-F739
[5] Ammon HP Anazodo MI Safayhi H Dhawan BN Srimal RC Curcumin A potent inhibitor of leukotriene B4 formation in rat peritoneal poly-morphonuclear neutrophils (PMNL) Planta Med 1992 58 226
[6] Epstein J Sanderson IR and MacDonald TT Curcumin as a therapeutic agent the evidence from in vitro animal and human studies Br J Nutr 2010 103 1545-1557
[7] Zhong F Chen H Han L Jin Y and Wang W Curcumin attenuates lipopolysaccharide-in-duced renal inflammation Biol Pharm Bull 2011 34 226-232
[8] Balasubramanyam K Varier RA Altaf M Swaminathan V Siddappa NB Ranga U and Kundu TK Curcumin a novel p300CREB-binding protein-specific inhibitor of acetyltrans-ferase represses the acetylation of histonenonhistone proteins and histone acetyltrans-ferase-dependent chromatin transcription J Biol Chem 2004 279 51163-51171
[9] Bertani T Rocchi G Sacchi G Mecca G and Remuzzi G Adriamycin-induced glomeruloscle-rosis in the rat Am J Kidney Dis 1986 7 12-19
Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780

Histone deacetylation effect of curcumin in Kidney disease
12704 Int J Clin Exp Med 20169(7)12696-12704
[10] Mundel P Reiser J Borja AZMa Pavenstaumldt H Davidson GR Kriz W and Zeller R Rearrangements of the cytoskeleton and cell contacts induce process formation during dif-ferentiation of conditionally immortalized mouse podocyte cell lines Exp Cell Res 1997 236 248-258
[11] Backs J and Olson EN Control of cardiac growth by histone acetylationdeacetylation Circ Res 2006 98 15-24
[12] Barnes PJ Adcock IM and Ito K Histone acety-lation and deacetylation importance in inflam-matory lung diseases Eur Respir J 2005 25 552-563
[13] Coward WR Watts K Feghali-Bostwick CA Knox A and Pang L Defective histone acetyla-tion is responsible for the diminished expres-sion of cyclooxygenase 2 in idiopathic pulmo-nary fibrosis Mol Cell Biol 2009 29 4325-4339
[14] Pang M and Zhuang S Histone deacetylase a potential therapeutic target for fibrotic disor-ders J Pharmacol Exp Ther 2010 335 266-272
[15] Marumo T Hishikawa K Yoshikawa M Hirahashi J Kawachi S and Fujita T Histone deacetylase modulates the proinflammatory and-fibrotic changes in tubulointerstitial injury Am J Physiol Renal Physiol 2010 298 F133-F141
[16] Pang M Kothapally J Mao H Tolbert E Ponnusamy M Chin YE and Zhuang S Inhibition of histone deacetylase activity at-tenuates renal fibroblast activation and inter-stitial fibrosis in obstructive nephropathy Am J Physiol Renal Physiol 2009 297 F996-F1005
[17] Yun JM Jialal I and Devaraj S Epigenetic regu-lation of high glucose-induced proinflammato-ry cytokine production in monocytes by cur-cumin J Nutr Biochem 2011 22 450-458
[18] Khan SN and Khan AU Role of histone acetyla-tion in cell physiology and diseases An update Clin Chim Acta 2010 411 1401-1411
[19] Gayther SA Batley SJ Linger L Bannister A Thorpe K Chin SF Daigo Y Russell P Wilson A and Sowter HM Mutations truncating the EP300 acetylase in human cancers Nat Genet 2000 24 300-303
[20] McKinsey TA Therapeutic potential for HDAC inhibitors in the heart Annu Rev Pharmacol Toxicol 2012 52 303-319
[21] Barnes PJ Role of HDAC2 in the pathophysiol-ogy of COPD Annu Rev Physiol 2009 71 451-464
[22] Liu W Fan LX Zhou X Sweeney WE Jr Avner ED and Li X HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic traf-ficking and degradation in renal epithelial cells PLoS One 2012 7 e49418
[23] Li Y Kang YS Dai C Kiss LP Wen X and Liu Y Epithelial-to-mesenchymal transition is a po-tential pathway leading to podocyte dysfunc-tion and proteinuria Am J Pathol 2008 172 299-308
[24] Mundel P and Reiser J Proteinuria an enzy-matic disease of the podocyte Kidney Int 2010 77 571-580
[25] Viedt C and Orth SR Monocyte chemoattrac-tant protein-1 (MCP-1) in the kidney does it more than simply attract monocytes Nephrol Dial Transplant 2002 17 2043-2047
[26] Tesch GH MCP-1CCL2 a new diagnostic marker and therapeutic target for progressive renal injury in diabetic nephropathy Am J Physiol Renal Physiol 2008 294 F697-F701
[27] Eardley KS Zehnder D Quinkler M Lepenies J Bates RL Savage CO Howie AJ Adu D and Cockwell P The relationship between albumin-uria MCP-1CCL2 and interstitial macro-phages in chronic kidney disease Kidney Int 2006 69 1189-1197
[28] Welsh GI and Saleem MA The podocyte cyto-skeleton-key to a functioning glomerulus in health and disease Nat Rev Nephrol 2011 8 14-21
[29] Menzel S and Moeller MJ Role of the podo- cyte in proteinuria Pediatr Nephrol 2011 26 1775-1780





























